
Evolutionary Analysis of StSnRK2 Family Genes and Their Overexpression in Transgenic Tobacco Improve Drought Tolerance

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
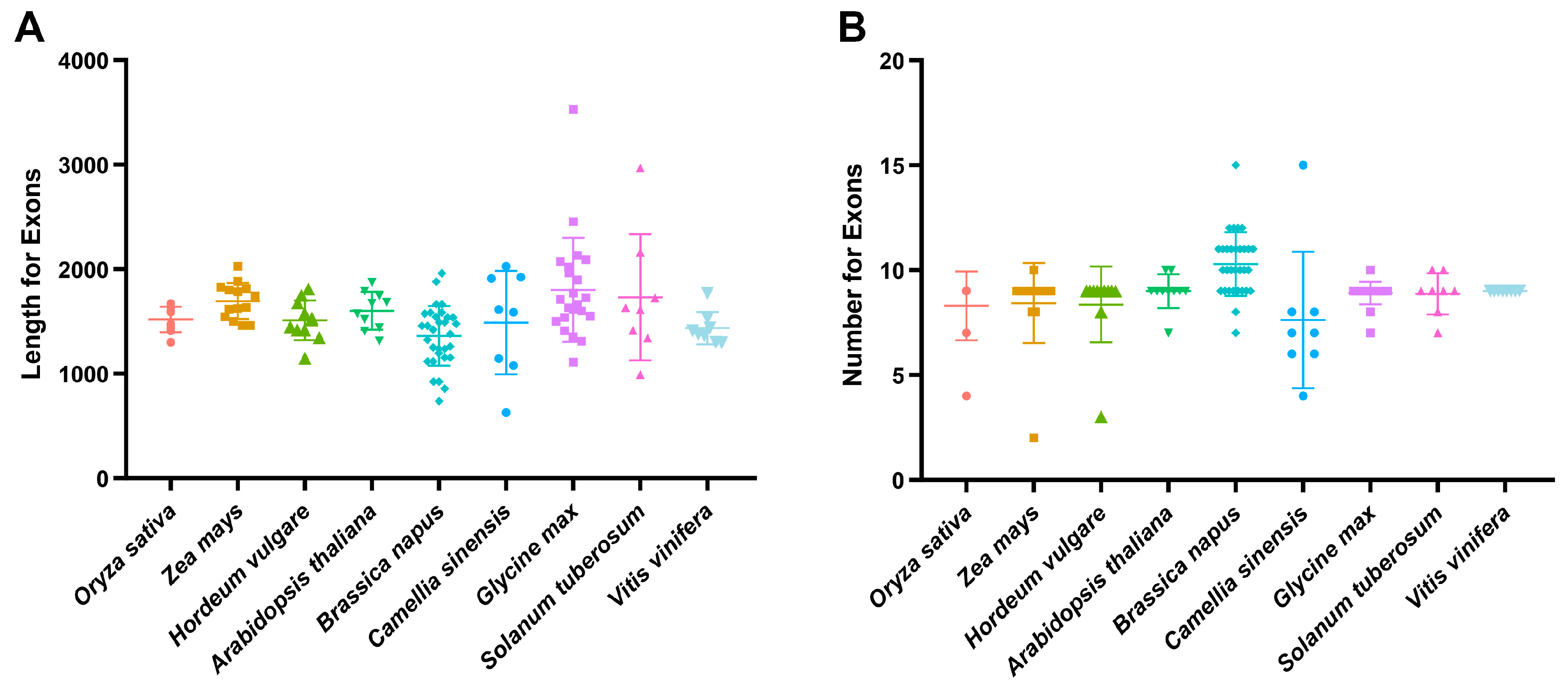
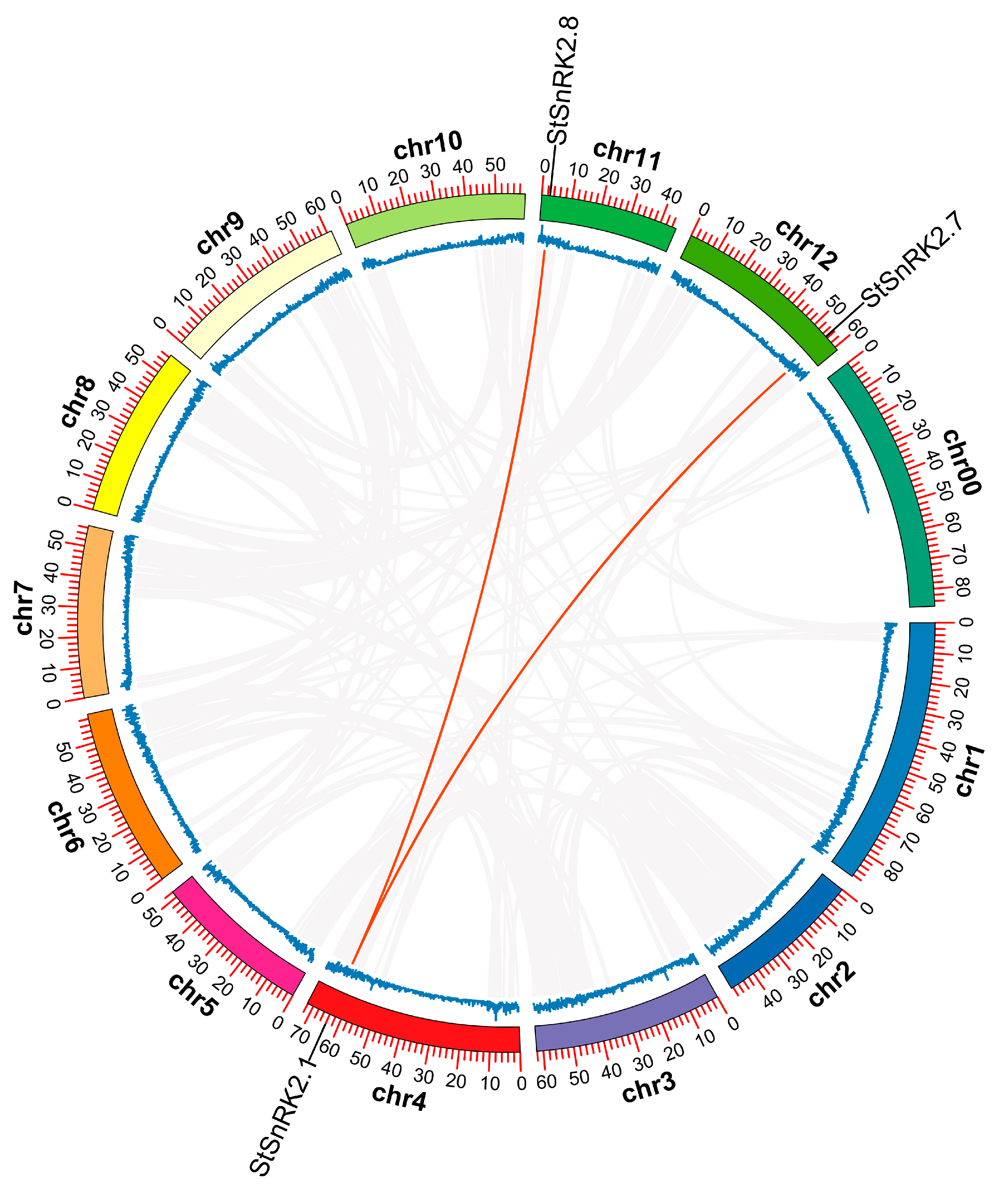
2.1. Gene Structure, Motif Composition and Chromosomal Distribution of StSnRK2 Genes
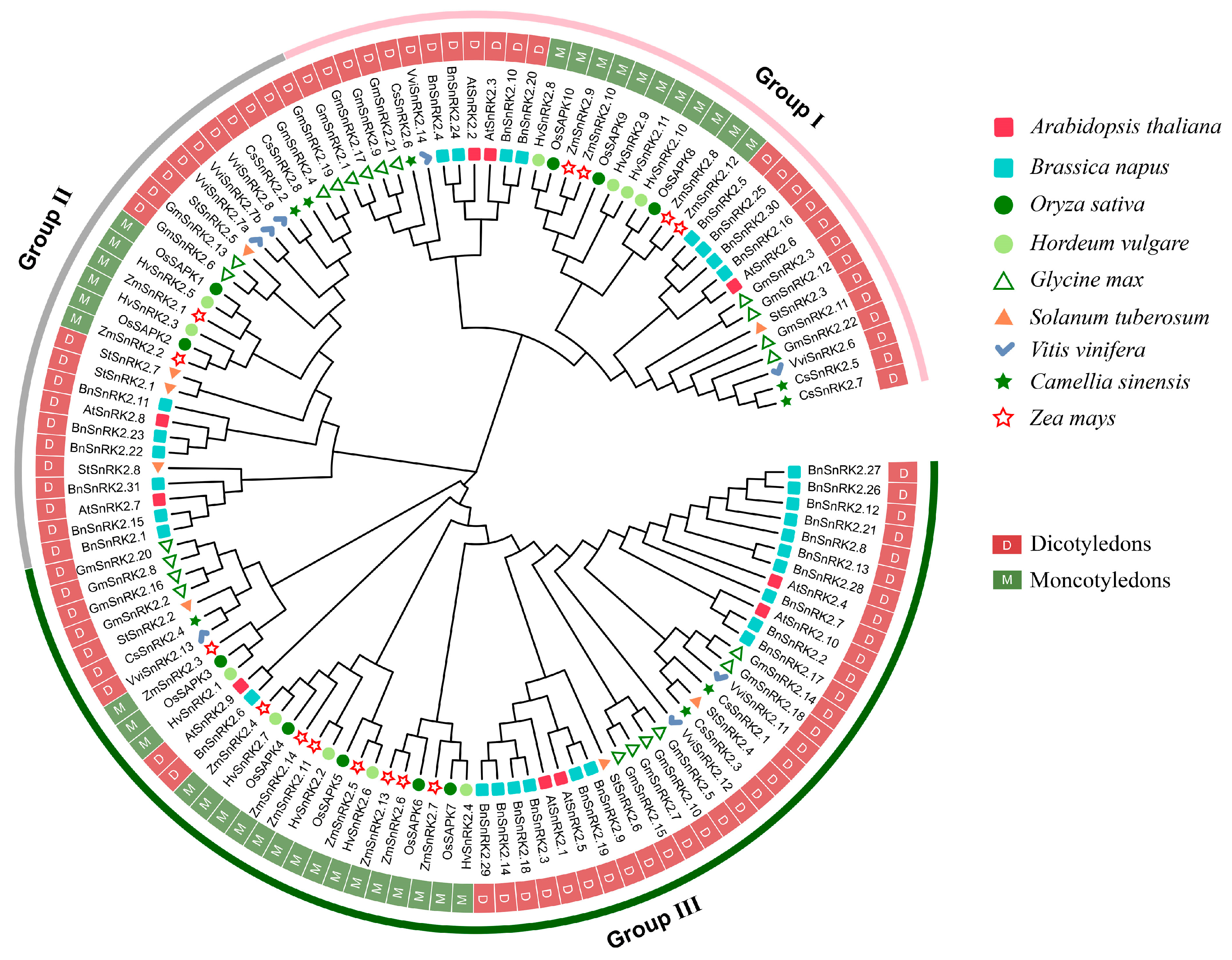
2.2. Phylogenetic Analysis Divides StSnRK2s into Three Subgroups
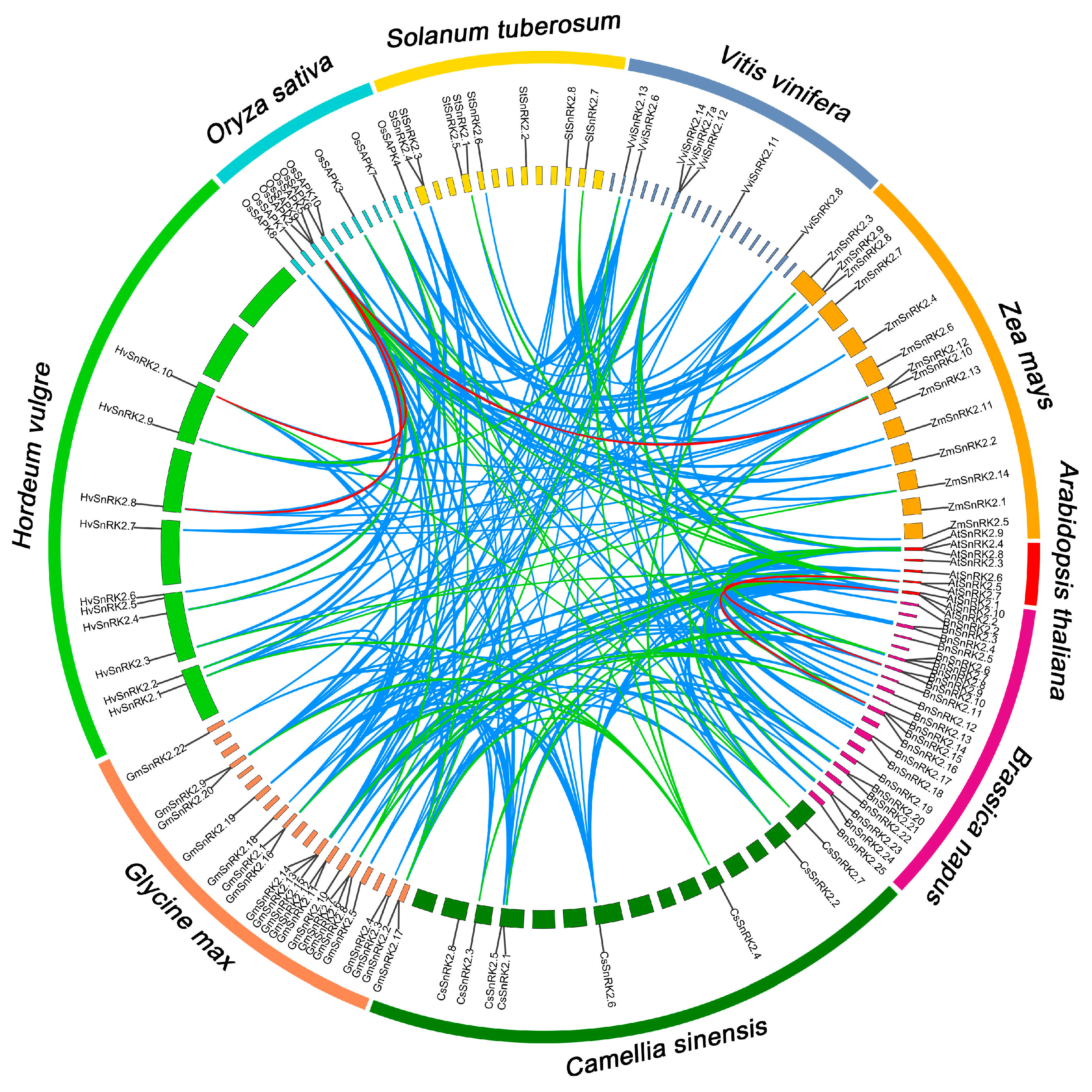
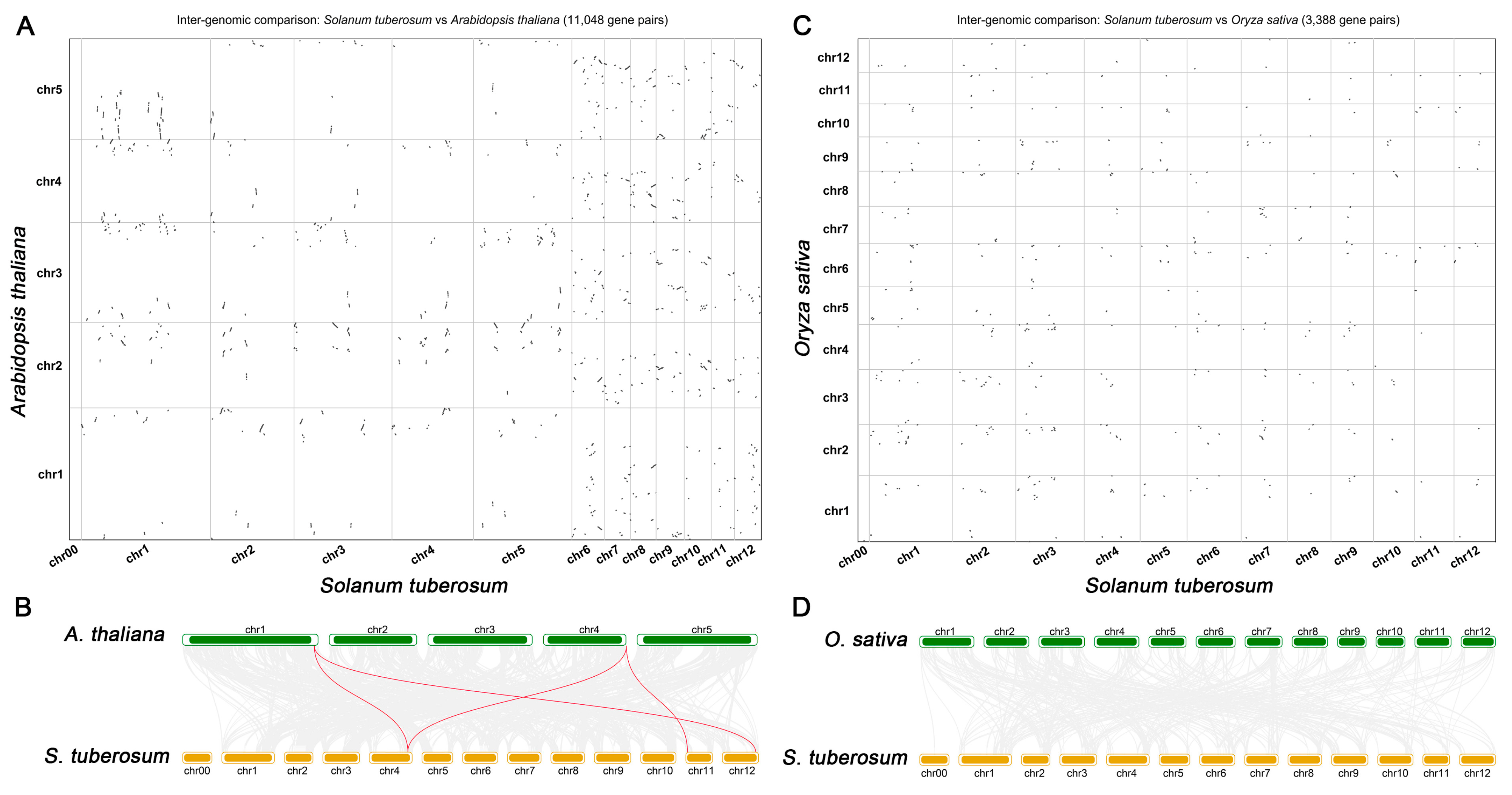
2.3. Synteny Analysis of StSnRK2 Genes
2.4. Evolutionary Selection Pressure and Codon Usage Bias Analysis
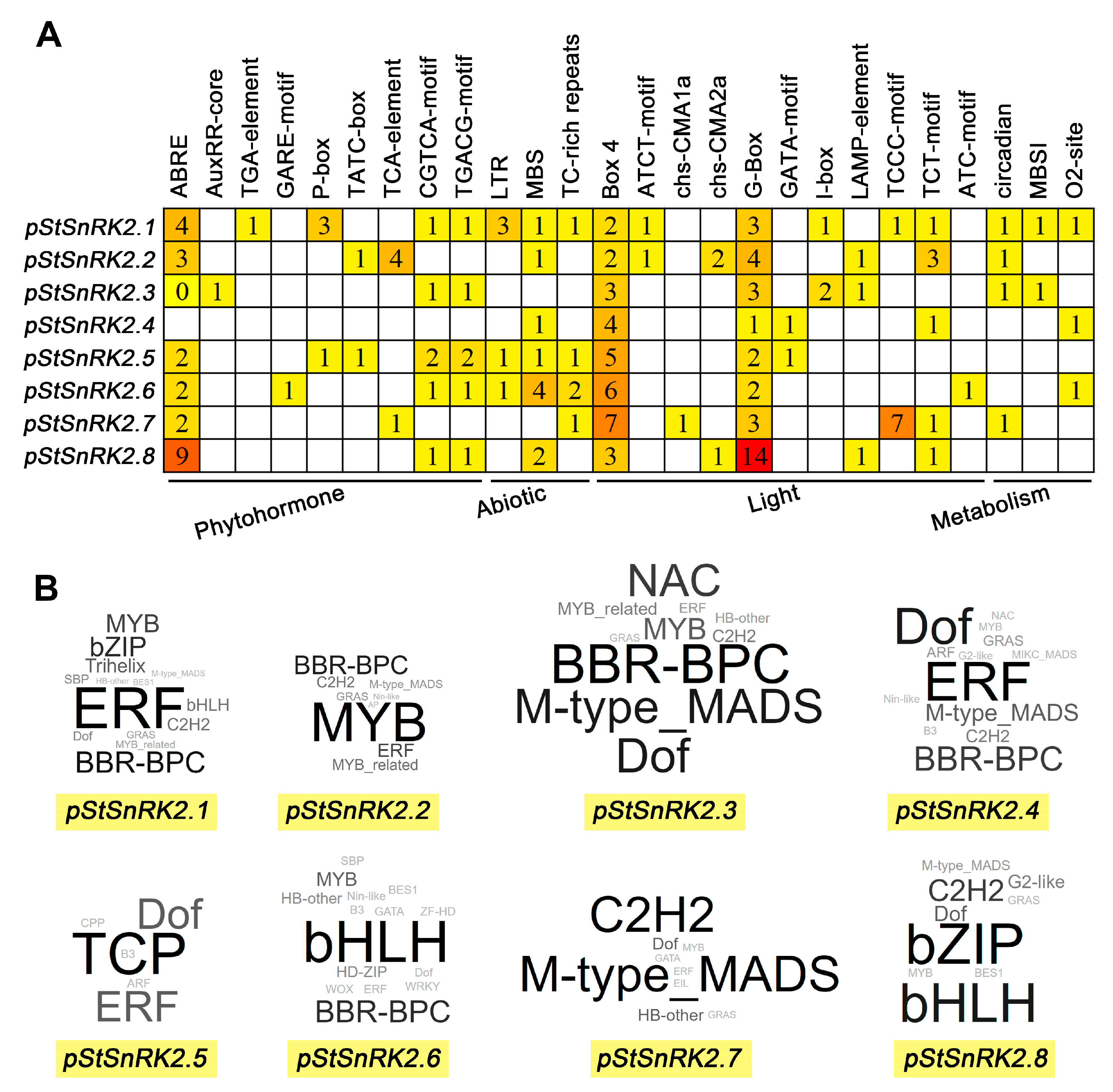
2.5. Prediction of Cis-Acting Elements and Binding Proteins of StSnRK2 Gene Promoters
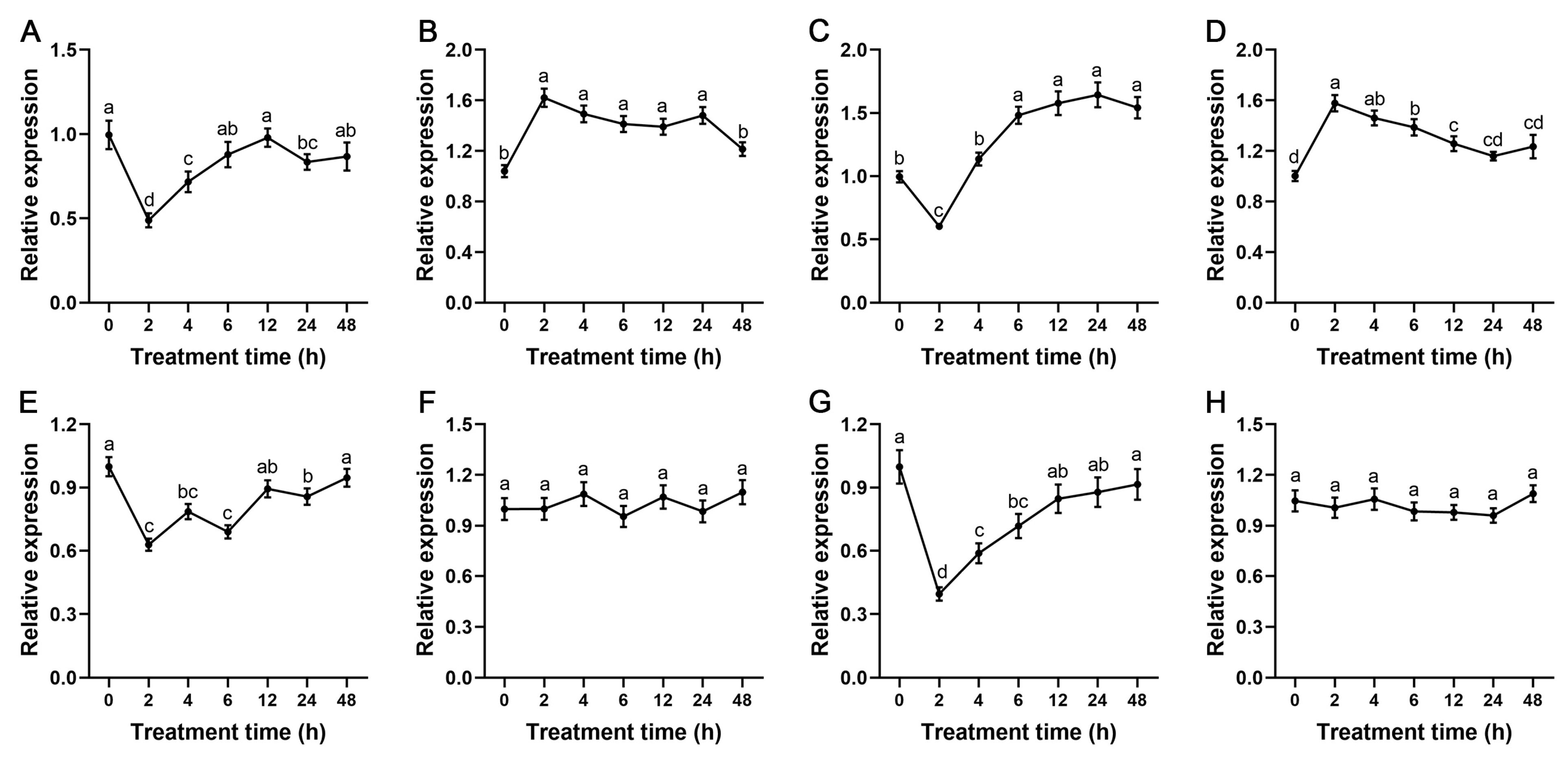
2.6. Expression Patterns of StSnRK2 Genes under ABA and Drought Treatments
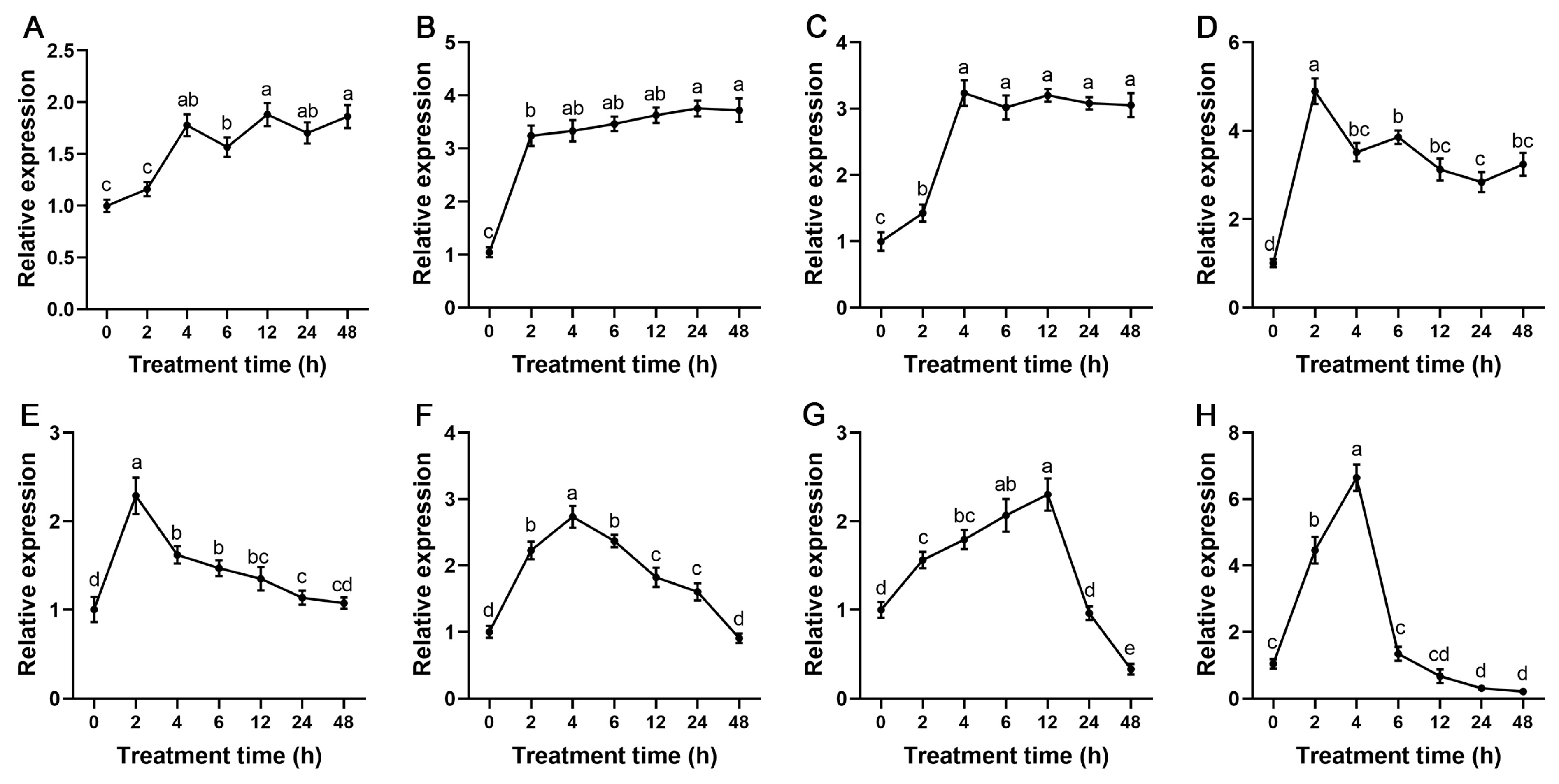
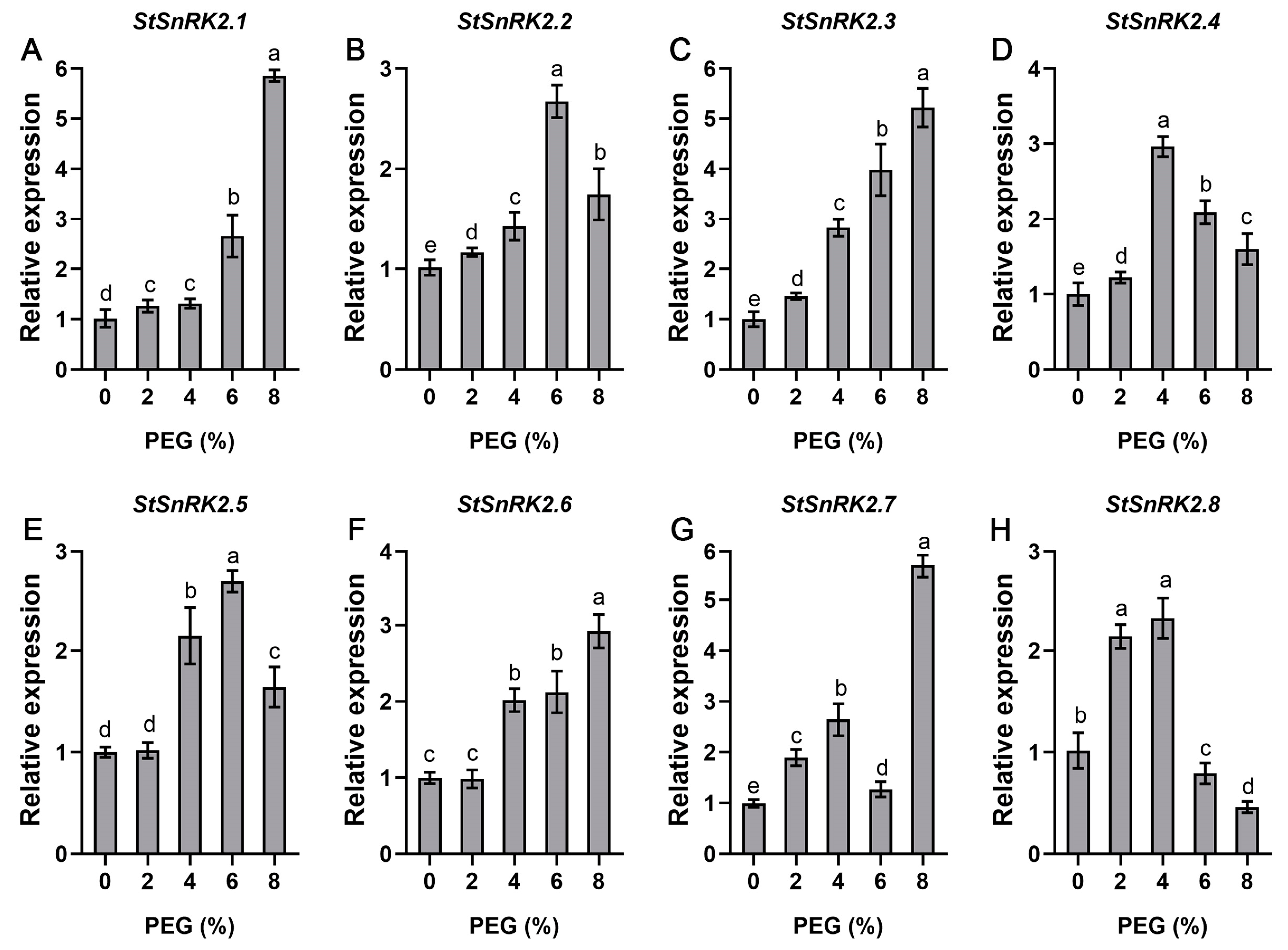
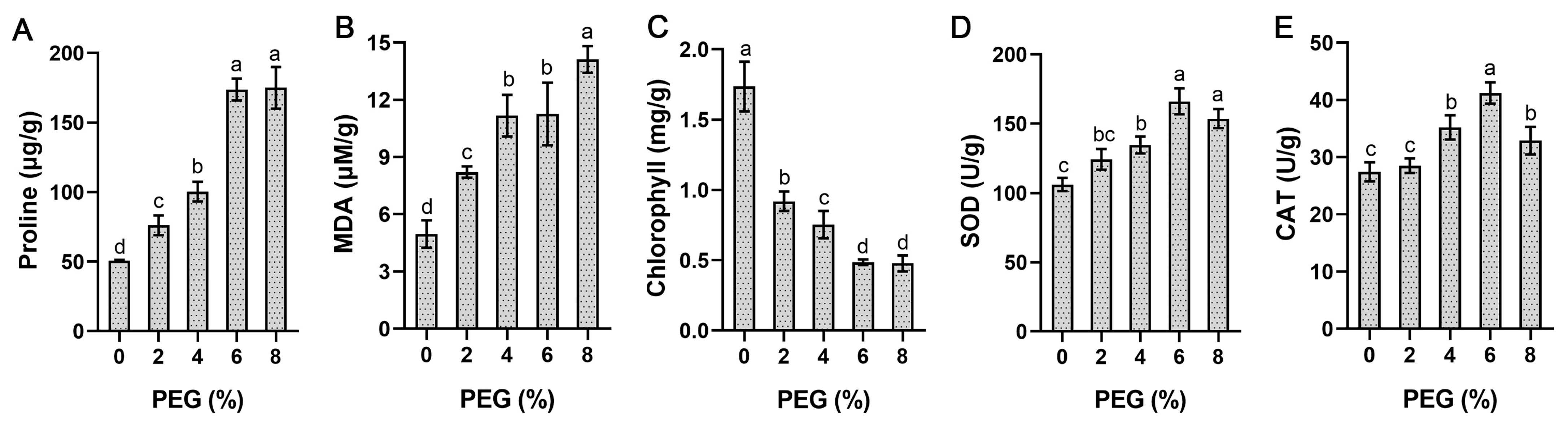
2.7. StSnRK2 Gene Expression and Physiological Changes under Different Degrees of Drought Stress
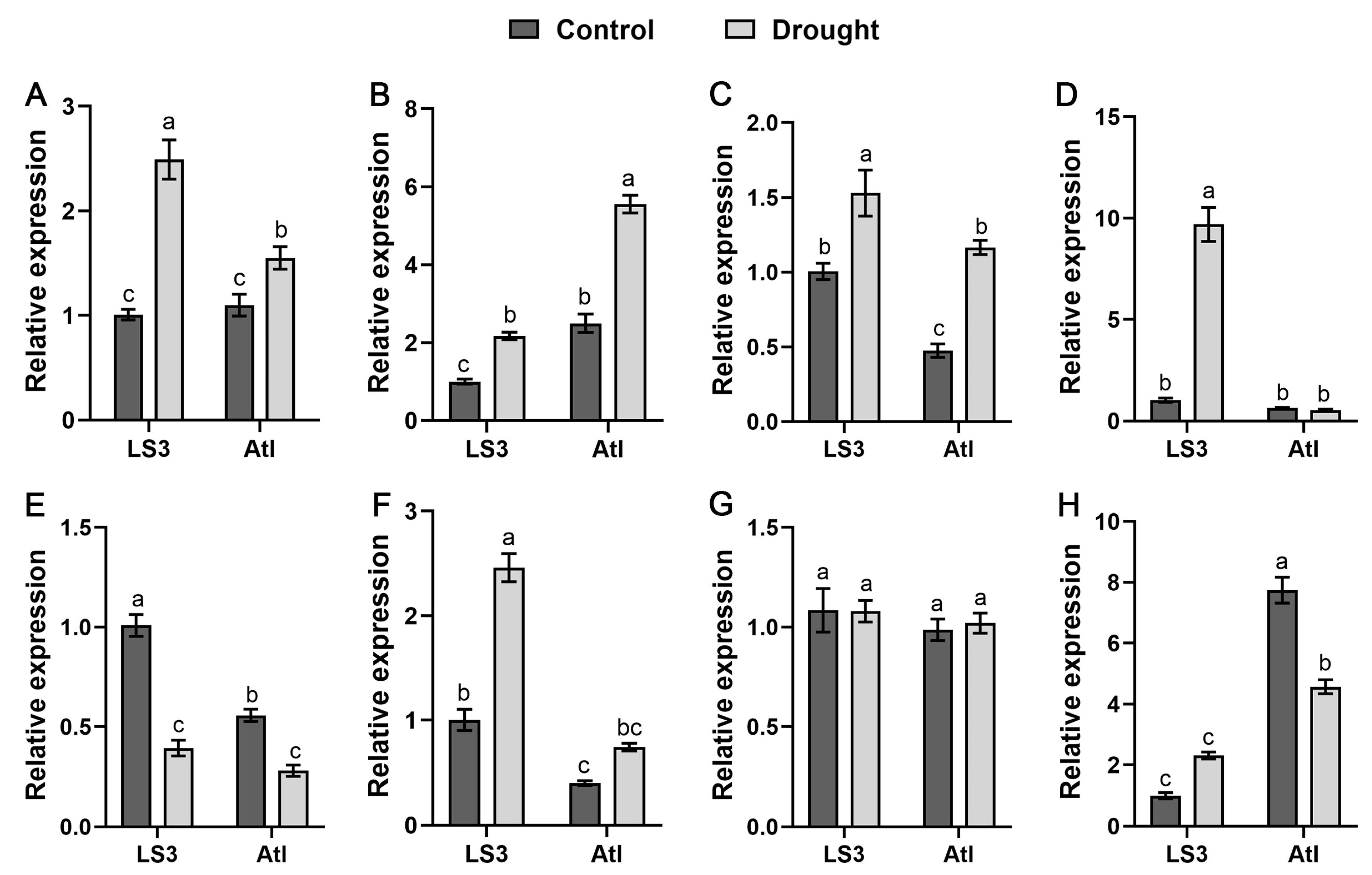
2.8. Response of StSnRK2 Genes to Drought Stress in Different Drought-Resistant Cultivars
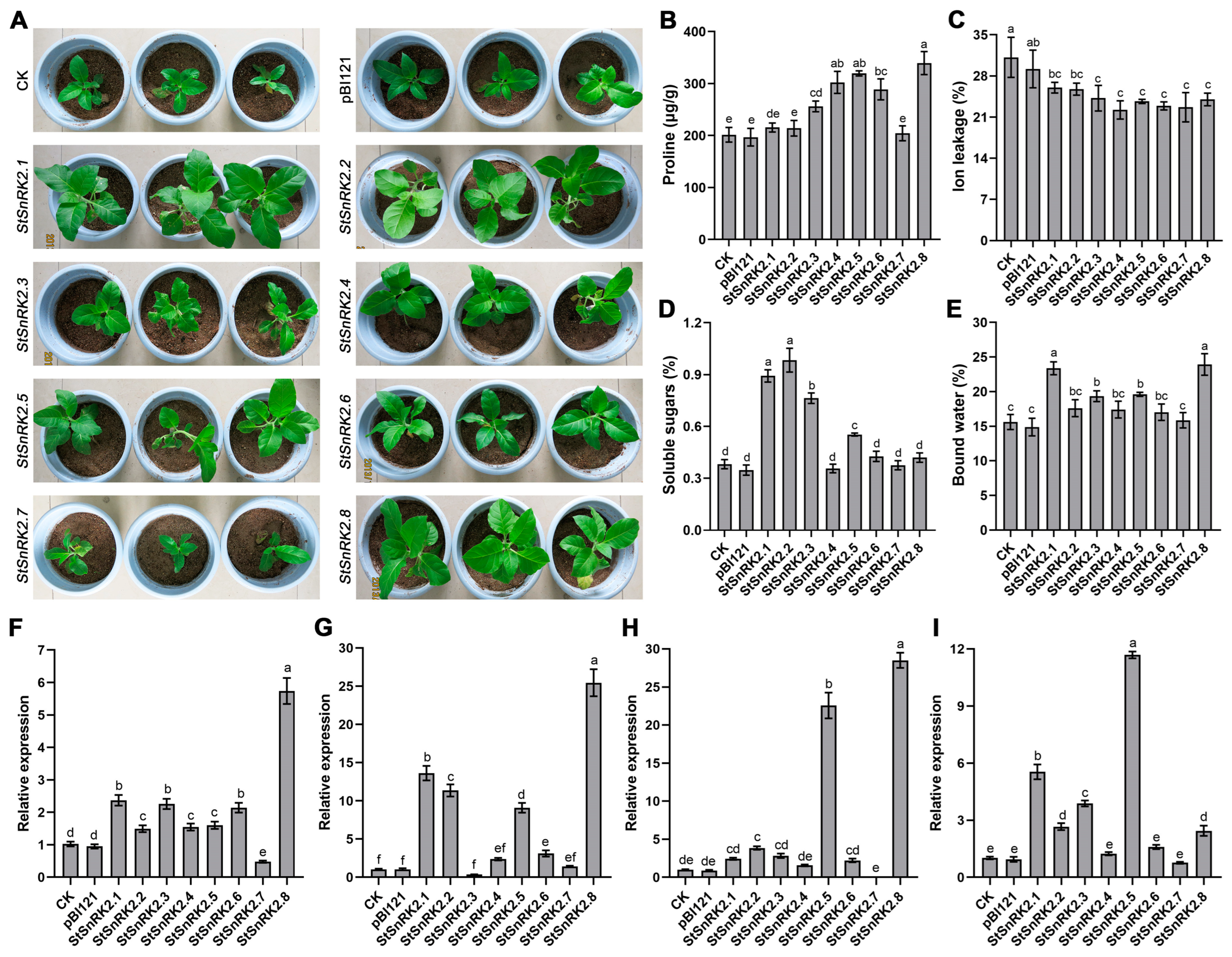
2.9. Effect of StSnRK2s on Drought Tolerance of Transgenic Tobacco
2.10. Physiological Characteristics of StSnRK2 Transgenic Plants under Drought Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Exogenous ABA and Drought Treatments
4.3. Tobacco Transformation
4.4. Drought Stress of Transgenic Tobacco
4.5. qRT-PCR
4.6. Intro/Exon Structures, conserved Motifs/Domains and Chromosomal Distribution of Potato SnRK2 Genes
4.7. Phylogenetic Analysis of Potato SnRK2 Proteins
4.8. Intraspecific/Interspecific Collinearity Analysis of Potato SnRK2 Genes
4.9. Selection Pressur, and Codon Usage Bias Analysis of Potato SnRK2 Genes
4.10. Cis-Element Analysis of Potato SnRK2 Genes
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef] [PubMed]
- Simelton, E.; Fraser, E.D.; Termansen, M.; Forster, P.M.; Dougill, A.J. Typologies of crop-drought vulnerability: An empirical analysis of the socio-economic factors that influence the sensitivity and resilience to drought of three major food crops in China (1961–2001). Environ. Sci. Policy 2009, 12, 438–452. [Google Scholar] [CrossRef]
- Chen, G.; Zhou, Y.; Kishchenko, O.; Stepanenko, A.; Jatayev, S.; Zhang, D.; Borisjuk, N. Gene editing to facilitate hybrid crop production. Biotechnol. Adv. 2021, 46, 107676. [Google Scholar] [CrossRef] [PubMed]
- Georges, F.; Ray, H. Genome editing of crops: A renewed opportunity for food security. Gm. Crops Food. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Wu, J.; Jiang, M.; Wang, Y. Plant mitogen-activated protein kinase cascades in environmental stresses. Int. J. Mol. Sci. 2021, 22, 1543. [Google Scholar] [CrossRef] [PubMed]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.i.; Kolapo, K. Drought resistance in rice from conventional to molecular breeding: A review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Zhang, J.P.; Yan, K.; Zhou, Z.G.; Zhao, W.Q.; Zhang, X.D.; Pu, Y.H.; Yu, R.X. Beneficial effects of abscisic acid and melatonin in overcoming drought stress in cotton (Gossypium hirsutum L.). Physiol. Plantarum. 2021, 173, 2041–2054. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Harshavardhan, V.T.; Govind, G.; Seiler, C.; Kohli, A. Contrapuntal role of ABA: Does it mediate stress tolerance or plant growth retardation under long-term drought stress? Gene 2012, 506, 265–273. [Google Scholar] [CrossRef]
- Duarte, K.E.; Souza, W.R.; Santiago, T.R.; Sampaio, B.L.; Ribeiro, A.P.; Cotta, M.G.; Da, C.B.A.D.B.; Marraccini, P.R.R.; Kobayashi, A.K.; Molinari, H.B.C. Identification and characterization of core abscisic acid (ABA) signaling components and their gene expression profile in response to abiotic stresses in Setaria viridis. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Miyazono, K.I.; Miyakawa, T.; Sawano, Y.; Kubota, K.; Kang, H.J.; Asano, A.; Miyauchi, Y.; Takahashi, M.; Zhi, Y.; Fujita, Y. Structural basis of abscisic acid signalling. Nature 2009, 462, 609–614. [Google Scholar] [CrossRef]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F.F. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yan, H.; Qiu, Z.; Hu, B.; Zeng, B.; Zhong, C.; Fan, C. Comprehensive analysis of SnRK gene family and their responses to salt stress in Eucalyptus grandis. Int. J. Mol. Sci. 2019, 20, 2786. [Google Scholar] [CrossRef] [Green Version]
- Saha, J.; Chatterjee, C.; Sengupta, A.; Gupta, K.; Gupta, B. Genome-wide analysis and evolutionary study of sucrose non-fermenting 1-related protein kinase 2 (SnRK2) gene family members in Arabidopsis and Oryza. Comput. Biol. Chem. 2014, 49, 59–70. [Google Scholar] [CrossRef]
- Xu, M.R.; Huang, L.Y.; Zhang, F.; Zhu, L.H.; Zhou, Y.L.; Li, Z.K. Genome-wide phylogenetic analysis of stress-activated protein kinase genes in rice (OsSAPKs) and expression profiling in response to Xanthomonas oryzae pv. oryzicola infection. Plant Mol. Biol. Rep. 2013, 31, 877–885. [Google Scholar] [CrossRef]
- Long, T.; Xu, B.; Hu, Y.; Wang, Y.; Mao, C.; Wang, Y.; Zhang, J.; Liu, H.; Huang, H.; Liu, Y. Genome-wide identification of ZmSnRK2 genes and functional analysis of ZmSnRK2.10 in ABA signaling pathway in maize (Zea mays L.). BMC Plant Biol. 2021, 21, 1–17. [Google Scholar] [CrossRef]
- Zhao, W.; Cheng, Y.H.; Zhang, C.; Shen, X.J.; You, Q.B.; Guo, W.; Li, X.; Song, X.J.; Zhou, X.A.; Jiao, Y.Q. Genome-wide identification and characterization of the GmSnRK2 family in soybean. Int. J. Mol. Sci. 2017, 18, 1834. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Song, J.; Li, C.; Ma, J.; Liu, J.; Zhu, X.; Li, J.; He, F.; Yang, C. Genome-wide identification and expression profile analysis of the SnRK2 gene family in Nicotiana tabacum. Biochem. Genet. 2022, 60, 1–16. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Wan, S.Q.; Wang, W.D.; Chen, J.F.; Huang, L.L.; Duan, M.S.; Yu, Y.B. Genome-wide identification and characterization of the CsSnRK2 family in Camellia sinensis. Plant Physiol. Bioch. 2018, 132, 287–296. [Google Scholar] [CrossRef]
- Liu, Z.; Ge, X.; Yang, Z.; Zhang, C.; Zhao, G.; Chen, E.; Liu, J.; Zhang, X.; Li, F. Genome-wide identification and characterization of SnRK2 gene family in cotton (Gossypium hirsutum L.). BMC Genet. 2017, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Cheng, J.; Hu, F.; Qin, C.; Xu, X.; Hu, K. The SnRK2 family in pepper (Capsicum annuum L.): Genome-wide identification and expression analyses during fruit development and under abiotic stress. Genes Genom. 2020, 42, 1117–1130. [Google Scholar] [CrossRef]
- Liu, J.Y.; Chen, N.N.; Cheng, Z.M.; Xiong, J.S. Genome-wide identification, annotation and expression profile analysis of SnRK2 gene family in grapevine. Aust. J. Grape Wine Res. 2016, 22, 478–488. [Google Scholar] [CrossRef]
- Fatima, A.; Khan, M.J.; Awan, H.M.; Akhtar, M.N.; Bibi, N.; Sughra, K.; Khan, M.R.; Ahmad, R.; Ibrahim, M.; Hussain, J. Genome-wide identification and expression analysis of SnRK2 gene family in mungbean (Vigna radiata) in response to drought stress. Crop Pasture Sci. 2020, 71, 469–476. [Google Scholar] [CrossRef]
- Song, J.; Sun, P.; Kong, W.; Xie, Z.; Li, C.; Liu, J.H. SnRK2.4-mediated phosphorylation of ABF2 regulates ARGININE DECARBOXYLASE expression and putrescine accumulation under drought stress. New Phytol. 2022, 23, 3053–3057. [Google Scholar]
- Zhang, X.; Wu, S.; Liu, S.; Takano, T. The Arabidopsis sucrose non-fermenting-1-related protein kinase AtSnRK2.4 interacts with a transcription factor, AtMYB21, that is involved in salt tolerance. Plant Sci. 2021, 303, 110685. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhang, B.; Qin, F. Arabidopsis RZFP34/CHYR1, a ubiquitin E3 ligase, regulates stomatal movement and drought tolerance via SnRK2.6-mediated phosphorylation. Plant Cell. 2015, 27, 3228–3244. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yu, B.; Wu, Q.; Min, Q.; Zeng, R.; Xie, Z.; Huang, J. OsMADS23 phosphorylated by SAPK9 confers drought and salt tolerance by regulating ABA biosynthesis in rice. PLoS Genet. 2021, 17, 1009699. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, X.; Wang, C.; Jing, R. Overexpression of a common wheat gene TaSnRK2.8 enhances tolerance to drought, salt and low temperature in Arabidopsis. PLoS ONE 2010, 5, 16041. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Hu, J.; Fan, W.; Zhu, P.; Cao, B.; Zheng, S.; Xia, Z.; Zhu, Y.; Zhao, A. Heterotrimeric G-protein γ subunits regulate ABA signaling in response to drought through interacting with PP2Cs and SnRK2s in mulberry (Morus alba L.). Plant Physiol. Bioch. 2021, 161, 210–221. [Google Scholar] [CrossRef]
- Qin, R.; Zhang, F.; Yu, C.; Zhang, Q.; Qi, J.; Li, F.M. Contributions made by rain-fed potato with mulching to food security in China. Eur. J. Agron. 2022, 133, 126435. [Google Scholar] [CrossRef]
- Zhang, H.; Fen, X.; Yu, W.; Hu, H.h.; Dai, X.F. Progress of potato staple food research and industry development in China. J. Integr. Agric. 2017, 16, 2924–2932. [Google Scholar] [CrossRef]
- Waterer, D.; Benning, N.T.; Wu, G.; Luo, X.; Liu, X.; Gusta, M.; McHughen, A.; Gusta, L.V. Evaluation of abiotic stress tolerance of genetically modified potatoes (Solanum tuberosum cv. Desiree). Mol. Breed. 2010, 25, 527–540. [Google Scholar] [CrossRef]
- Shi, J.; Wang, J.; Wang, N.; Zhou, H.; Xu, Q.; Yan, G. Overexpression of StGA2ox1 gene increases the tolerance to abiotic stress in transgenic potato (Solanum tuberosum L.) plants. Appl. Biochem. Biotech. 2019, 187, 1204–1219. [Google Scholar] [CrossRef]
- Sun, C.; Liang, W.; Yan, K.; Xu, D.; Qin, T.; Fiaz, S.; Kear, P.; Bi, Z.; Liu, Y.; Liu, Z. Expression of potato StDRO1 in Arabidopsis alters root architecture and drought tolerance. Front. Plant Sci. 2022, 13, 836063. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Feng, S.; Yang, J.; Li, D.; Zhang, J. Roles of plasmalemma aquaporin gene StPIP1 in enhancing drought tolerance in potato. Front. Plant Sci. 2017, 8, 616. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Tang, X.; Liu, W.; Fu, X.; Luo, H.; Ghimire, S.; Zhang, N.; Si, H. A potato RING-finger protein gene StRFP2 is involved in drought tolerance. Plant Physiol. Bioch. 2020, 146, 438–446. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, N.; Liu, X.; Li, S.; Yang, J.; Hong, X.; Wang, F.; Si, H. Mitogen-activated protein kinase 11 (MAPK11) maintains growth and photosynthesis of potato plant under drought condition. Plant Cell Rep. 2021, 40, 491–506. [Google Scholar] [CrossRef]
- Li, S.; Zhang, N.; Zhu, X.; Ma, R.; Yang, J.; Tang, X.; Si, H. Enhanced drought tolerance with artificial microRNA-mediated StProDH1 gene silencing in potato. Crop Sci. 2020, 60, 1462–1471. [Google Scholar] [CrossRef]
- Bai, J.; Mao, J.; Yang, H.; Khan, A.; Fan, A.; Liu, S.; Zhang, J.; Wang, D.; Gao, H.; Zhang, J. Sucrose non-ferment 1 related protein kinase 2 (SnRK2) genes could mediate the stress responses in potato (Solanum tuberosum L.). BMC Genet. 2017, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef]
- Liu, L.; Xu, W.; Hu, X.; Liu, H.; Lin, Y. W-box and G-box elements play important roles in early senescence of rice flag leaf. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yu, Z.; Gao, L.; Zheng, C. SnRK2s at the crossroads of growth and stress responses. Trends Plant Sci. 2019, 24, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Weiner, J.J.; Peterson, F.C.; Volkman, B.F.; Cutler, S.R. Structural and functional insights into core ABA signaling. Curr. Opin. Plant Biol. 2010, 13, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.B.; Zhang, Y.R.; Liu, K.C.; Ni, Z.F.; Fang, Z.J.; Sun, Q.X.; Gao, J.W. Identification and bioinformatics analysis of SnRK2 and CIPK family genes in sorghum. Agric. Sci. China 2010, 9, 19–30. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Xu, C.; Wang, P.; Sun, W.; Yin, T.; Zhuge, Q. Heterologous overexpression of the Arabidopsis SnRK2.8 gene enhances drought and salt tolerance in Populus × euramericana cv ‘Nanlin895’. Plant Biotechnol. Rep. 2019, 13, 245–261. [Google Scholar] [CrossRef]
- Holland, P.W. (Ed.) Gene duplication: Past, present and future. Semin. Cell Dev. Biol. 1999, 10, 541–547. [Google Scholar] [CrossRef]
- Copley, S.D. Evolution of new enzymes by gene duplication and divergence. FEBS J. 2020, 287, 1262–1283. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.; Xu, H.; Xiao, S.; Qin, Y.; Li, Y.; Yan, Y.; Hu, Y. The large soybean (Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups. BMC Plant Biol. 2013, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Li, Y.; Zhang, Z.; Liu, X.; Hsu, C.C.; Du, Y.; Sang, T.; Zhu, C.; Wang, Y.; Satheesh, V. A RAF-SnRK2 kinase cascade mediates early osmotic stress signaling in higher plants. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Jia, X.; Xue, M.; Gao, Y.; Yue, H.; Ma, F.; Gong, X. MpSnRK2.10 confers salt stress tolerance in apple via the ABA signaling pathway. Sci. Hortic. 2022, 298, 110998. [Google Scholar] [CrossRef]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 confers abscisic acid sensitivity and tolerance to drought stress in rice. Front. Plant Sci. 2017, 8, 993. [Google Scholar] [CrossRef]
- Shinozawa, A.; Otake, R.; Takezawa, D.; Umezawa, T.; Komatsu, K.; Tanaka, K.; Amagai, A.; Ishikawa, S.; Hara, Y.; Kamisugi, Y. SnRK2 protein kinases represent an ancient system in plants for adaptation to a terrestrial environment. Commun. Biol. 2019, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Toriyama, T.; Shinozawa, A.; Yasumura, Y.; Saruhashi, M.; Hiraide, M.; Ito, S.; Matsuura, H.; Kuwata, K.; Yoshida, M.; Baba, T. Sensor histidine kinases mediate ABA and osmostress signaling in the moss Physcomitrium patens. Curr. Biol. 2022, 32, 164–175. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Vang, S.; Yu, J.; Wong, G.K.S.; Wang, J. Correlation between Ka/Ks and Ks is related to substitution model and evolutionary lineage. J. Mol. Evol. 2009, 68, 414–423. [Google Scholar] [CrossRef]
- Chen, L.Y.; Xin, Y.; Wai, C.M.; Liu, J.; Ming, R. The role of cis-elements in the evolution of crassulacean acid metabolism photosynthesis. Hortic Res. 2020, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.M.; Zhao, J.; Scandalios, J.G. Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J. 2000, 22, 87–95. [Google Scholar] [CrossRef]
- Huang, J.; Chen, F.; Guo, Y.; Gan, X.; Yang, M.; Zeng, W.; Persson, S.; Li, J.; Xu, W. GhMYB7 promotes secondary wall cellulose deposition in cotton fibres by regulating GhCesA gene expression through three distinct cis-elements. New Phytol. 2021, 232, 1718–1737. [Google Scholar] [CrossRef]
- Yang, F.; Dong, F.S.; Hu, F.H.; Liu, Y.W.; Chai, J.F.; Zhao, H.; Lv, M.Y.; Zhou, S. Genome-wide identification and expression analysis of the calmodulin-binding transcription activator (CAMTA) gene family in wheat (Triticum aestivum L.). BMC Genet. 2020, 21, 1–10. [Google Scholar] [CrossRef]
- Miao, L.L.; Li, Y.Y.; Zhang, H.J.; Zhang, H.J.; Liu, X.L.; Wang, J.Y.; Chang, X.P.; Mao, X.G.; Jing, R.L. TaSnRK2.4 is a vital regulator in control of thousand-kernel weight and response to abiotic stress in wheat. J. Integr. Agric. 2021, 20, 46–54. [Google Scholar] [CrossRef]
- Kim, H.; Hwang, H.; Hong, J.W.; Lee, Y.N.; Ahn, I.P.; Yoon, I.S.; Yoo, S.D.; Lee, S.; Lee, S.C.; Kim, B.G. A rice orthologue of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction pathway in seed germination and early seedling growth. J. Exp. Bot. 2012, 63, 1013–1024. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Z.; Hou, Y.; Wang, Y.; Wang, H.; Tong, X.; Ao, H.; Zhang, J. Protein interactomic analysis of SAPKs and ABA-inducible bZIPs revealed key roles of SAPK10 in rice flowering. Int. J. Mol. Sci. 2019, 20, 1427. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pattern | Species 1 | Species 2 | Nonsynonymous Substitution Rate (Ka) | Synonymous Substitution Rate (Ks) | Selection Strength (Ka/Ks) | Evolution Relationship | Divergence Time (Mya) |
|---|---|---|---|---|---|---|---|
| Monocots and monocots | Oryza sativa | Zea mays | 0.0526 | 0.6623 | 0.0798 | Purifying selection | 22.0782 |
| Hordeum vulgare | Zea mays | 0.0670 | 0.6602 | 0.1006 | Purifying selection | 22.0071 | |
| Hordeum vulgare | Oryza sativa | 0.0687 | 0.6601 | 0.0988 | Purifying selection | 22.0039 | |
| Monocots and dicots | Camellia sinensis | Oryza sativa | 0.1086 | 2.7496 | 0.0396 | Purifying selection | 91.6526 |
| Camellia sinensis | Hordeum vulgare | 0.1096 | 2.2413 | 0.0489 | Purifying selection | 74.7091 | |
| Camellia sinensis | Zea mays | 0.1468 | 3.1925 | 0.0481 | Purifying selection | 106.4167 | |
| Glycine max | Oryza sativa | 0.1783 | 3.0397 | 0.0588 | Purifying selection | 101.3221 | |
| Glycine max | Hordeum vulgare | 0.1392 | 2.7527 | 0.0505 | Purifying selection | 91.7571 | |
| Glycine max | Zea mays | 0.1419 | 2.3420 | 0.0607 | Purifying selection | 78.0659 | |
| Vitis vinifera | Hordeum vulgare | 0.1593 | 2.5584 | 0.0622 | Purifying selection | 85.2823 | |
| Vitis vinifera | Zea mays | 0.1292 | 6.7659 | 0.0190 | Purifying selection | 225.5309 | |
| Dicots and dicots | Arabidopsis thaliana | Brassica napus | 0.0502 | 0.5086 | 0.0992 | Purifying selection | 16.9549 |
| Arabidopsis thaliana | Glycine max | 0.1121 | 1.9277 | 0.0646 | Purifying selection | 64.2576 | |
| Arabidopsis thaliana | Vitis vinifera | 0.1268 | 1.3427 | 0.0944 | Purifying selection | 44.7592 | |
| Arabidopsis thaliana | Solanum tuberosum | 0.1268 | 1.7212 | 0.1455 | Purifying selection | 57.3766 | |
| Arabidopsis thaliana | Camellia sinensis | 0.1160 | 2.5458 | 0.0492 | Purifying selection | 84.8618 | |
| Brassica napus | Glycine max | 0.1191 | 1.7633 | 0.0682 | Purifying selection | 58.7779 | |
| Brassica napus | Camellia sinensis | 0.1694 | 1.4868 | 0.1192 | Purifying selection | 49.5612 | |
| Brassica napus | Solanum tuberosum | 0.1824 | 2.4539 | 0.0750 | Purifying selection | 81.7989 | |
| Brassica napus | Vitis vinifera | 0.1346 | 1.9996 | 0.0674 | Purifying selection | 66.6560 | |
| Camellia sinensis | Glycine max | 0.1028 | 1.4346 | 0.0762 | Purifying selection | 47.8206 | |
| Camellia sinensis | Solanum tuberosum | 0.0885 | 2.0827 | 0.0428 | Purifying selection | 69.4264 | |
| Camellia sinensis | Vitis vinifera | 0.0885 | 1.0203 | 0.0979 | Purifying selection | 34.0108 | |
| Glycine max | Solanum tuberosum | 0.1254 | 1.6182 | 0.0885 | Purifying selection | 53.9403 | |
| Glycine max | Vitis vinifera | 0.0893 | 1.2701 | 0.0722 | Purifying selection | 42.3375 | |
| Solanum tuberosum | Vitis vinifera | 0.1327 | 1.4690 | 0.0981 | Purifying selection | 48.9677 |
| Species | T3s | C3s | A3s | G3s | GC3s | CAI | CBI | FOP | Nc |
|---|---|---|---|---|---|---|---|---|---|
| Arabidopsis thaliana | 0.373 ± 0.096 | 0.245 ± 0.028 | 0.359 ± 0.043 | 0.301 ± 0.071 | 0.416 ± 0.068 | 0.177 ± 0.041 | −0.069 ± 0.059 | 0.374 ± 0.038 | 52.902 ± 3.802 |
| Brassica napus | 0.407 ± 0.037 | 0.277 ± 0.028 | 0.317 ± 0.043 | 0.297 ± 0.036 | 0.429 ± 0.040 | 0.223 ± 0.021 | −0.006 ± 0.050 | 0.418 ± 0.030 | 53.141 ± 2.886 |
| Camellia sinensis | 0.395 ± 0.032 | 0.256 ± 0.039 | 0.350 ± 0.048 | 0.288 ± 0.033 | 0.409 ± 0.050 | 0.196 ± 0.016 | −0.070 ± 0.069 | 0.377 ± 0.039 | 55.359 ± 2.575 |
| Glycine max | 0.434 ± 0.025 | 0.220 ± 0.023 | 0.341 ± 0.039 | 0.304 ± 0.036 | 0.386 ± 0.032 | 0.187 ± 0.016 | −0.116 ± 0.040 | 0.351 ± 0.025 | 52.645 ± 2.653 |
| Hordeum vulgare | 0.229 ± 0.076 | 0.292 ± 0.069 | 0.284 ± 0.054 | 0.367 ± 0.067 | 0.556 ± 0.081 | 0.142 ± 0.042 | −0.063 ± 0.051 | 0.384 ± 0.029 | 54.339 ± 4.296 |
| Oryza sativa | 0.276 ± 0.066 | 0.303 ± 0.092 | 0.317 ± 0.100 | 0.316 ± 0.097 | 0.502 ± 0.121 | 0.168 ± 0.056 | −0.011 ± 0.086 | 0.406 ± 0.051 | 53.879 ± 2.220 |
| Solanum tuberosum | 0.382 ± 0.036 | 0.247 ± 0.019 | 0.415 ± 0.040 | 0.241 ± 0.077 | 0.372 ± 0.045 | 0.172 ± 0.020 | −0.106 ± 0.031 | 0.362 ± 0.018 | 51.548 ± 1.442 |
| Vitis vinifera | 0.398 ± 0.028 | 0.231 ± 0.026 | 0.348 ± 0.031 | 0.320 ± 0.040 | 0.408 ± 0.043 | 0.176 ± 0.018 | −0.137 ± 0.063 | 0.338 ± 0.038 | 55.103 ± 3.300 |
| Zea mays | 0.318 ± 0.085 | 0.297 ± 0.108 | 0.354 ± 0.115 | 0.277 ± 0.128 | 0.452 ± 0.162 | 0.178 ± 0.043 | −0.042 ± 0.082 | 0.393 ± 0.046 | 52.848 ± 5.337 |
| T3s | C3s | A3s | G3s | GC3s | |
|---|---|---|---|---|---|
| CAI | 0.2666 * | 0.4727 * | −0.5666 * | 0.2097 | 0.3364 * |
| CBI | −0.1442 | 0.6568 * | −0.4286 * | 0.2097 | 0.5018 * |
| FOP | −0.0598 | 0.6568 * | −0.3592 * | 0.0543 | 0.4095 * |
| Nc | −0.4185 * | 0.4242 * | −0.3592 * | 0.0543 * | 0.4840 * |
| T3s | C3s | A3s | G3s | GC3s | |
|---|---|---|---|---|---|
| CAI | −0.0119 | 0.8613 * | −0.5995 * | 0.4265 * | 0.4933 * |
| CBI | −0.4263 * | 0.8336 * | −0.7127 * | 0.5654 * | 0.6814 * |
| FOP | −0.4431 * | 0.8667 * | −0.7600 * | 0.6128 * | 0.6814 * |
| Nc | 0.5374 * | −0.3702 * | 0.3275 | 0.6128 * | 0.6814 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, P.; Sun, L.; Dekomah, S.; Bi, Z.; Sun, C.; Mao, J.; Zhang, C.; Qin, T.; Wang, Y.; Liu, Y.; et al. Evolutionary Analysis of StSnRK2 Family Genes and Their Overexpression in Transgenic Tobacco Improve Drought Tolerance. Int. J. Mol. Sci. 2023, 24, 1000. https://doi.org/10.3390/ijms24021000
Yao P, Sun L, Dekomah S, Bi Z, Sun C, Mao J, Zhang C, Qin T, Wang Y, Liu Y, et al. Evolutionary Analysis of StSnRK2 Family Genes and Their Overexpression in Transgenic Tobacco Improve Drought Tolerance. International Journal of Molecular Sciences. 2023; 24(2):1000. https://doi.org/10.3390/ijms24021000
Chicago/Turabian StyleYao, Panfeng, Lei Sun, Simon Dekomah, Zhenzhen Bi, Chao Sun, Juan Mao, Chunli Zhang, Tianyuan Qin, Yihao Wang, Yuhui Liu, and et al. 2023. "Evolutionary Analysis of StSnRK2 Family Genes and Their Overexpression in Transgenic Tobacco Improve Drought Tolerance" International Journal of Molecular Sciences 24, no. 2: 1000. https://doi.org/10.3390/ijms24021000
APA StyleYao, P., Sun, L., Dekomah, S., Bi, Z., Sun, C., Mao, J., Zhang, C., Qin, T., Wang, Y., Liu, Y., Liu, Z., Ali, K., & Bai, J. (2023). Evolutionary Analysis of StSnRK2 Family Genes and Their Overexpression in Transgenic Tobacco Improve Drought Tolerance. International Journal of Molecular Sciences, 24(2), 1000. https://doi.org/10.3390/ijms24021000