
Interleukin-31 Receptor A Expression in the Dorsal Root Ganglion of Mice with Atopic Dermatitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
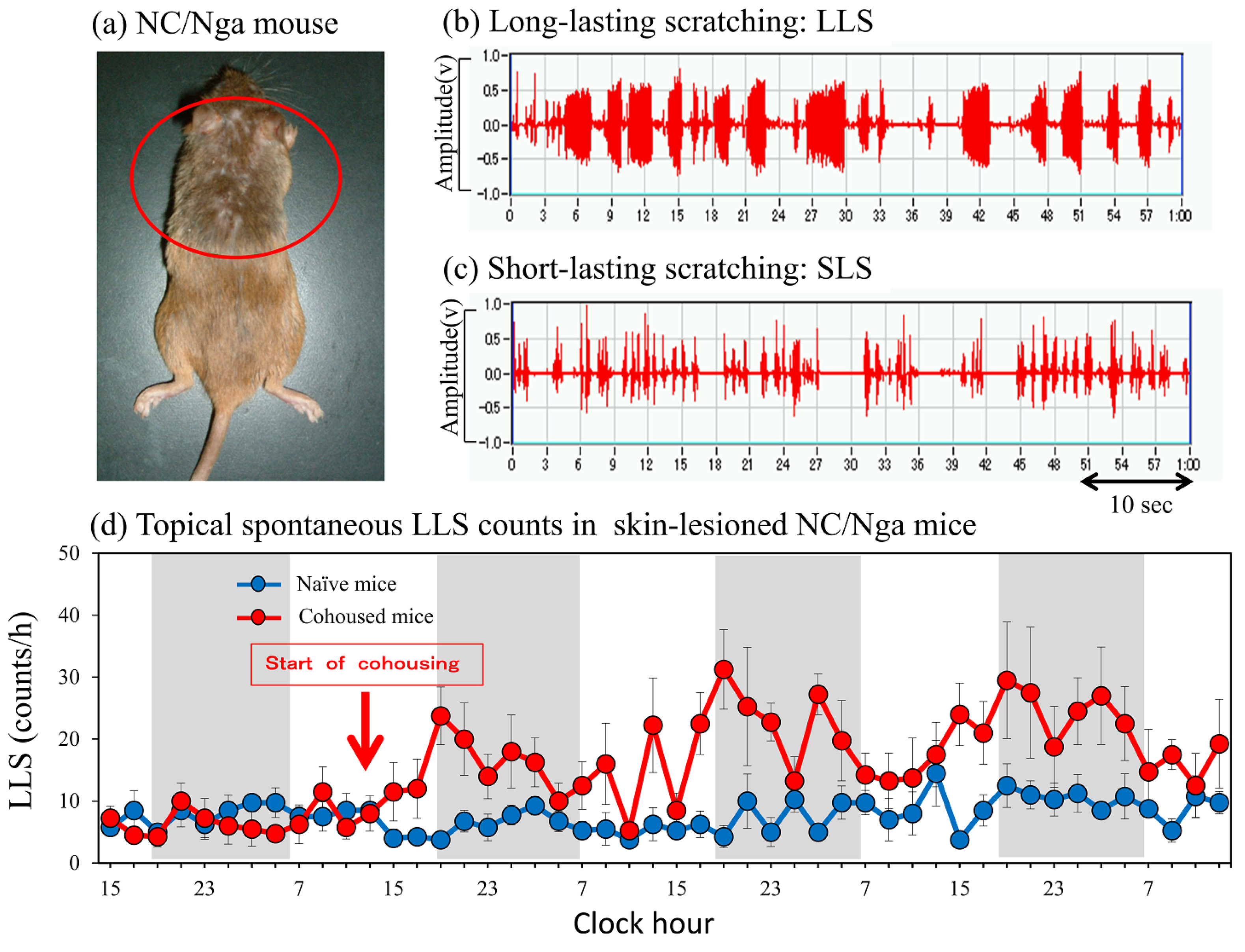
2.1. LLS Caused by Cohousing with Skin-Lesioned NC/Nga Mice
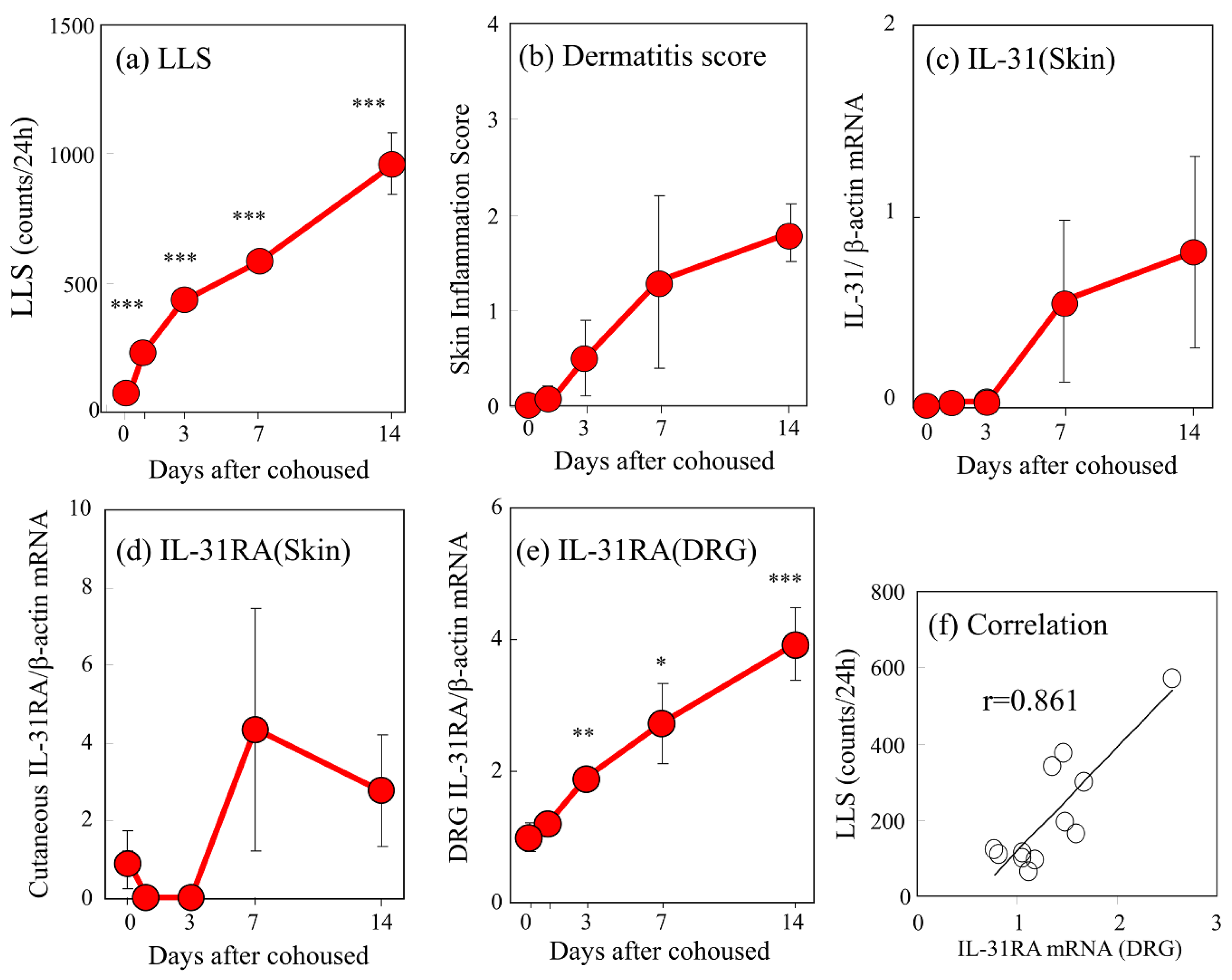
2.2. Time-Course Changes in Several Parameters of Dermatopathy in SPF-NC/Nga Mice Cohoused with Skin-Lesioned NC/Nga Mice
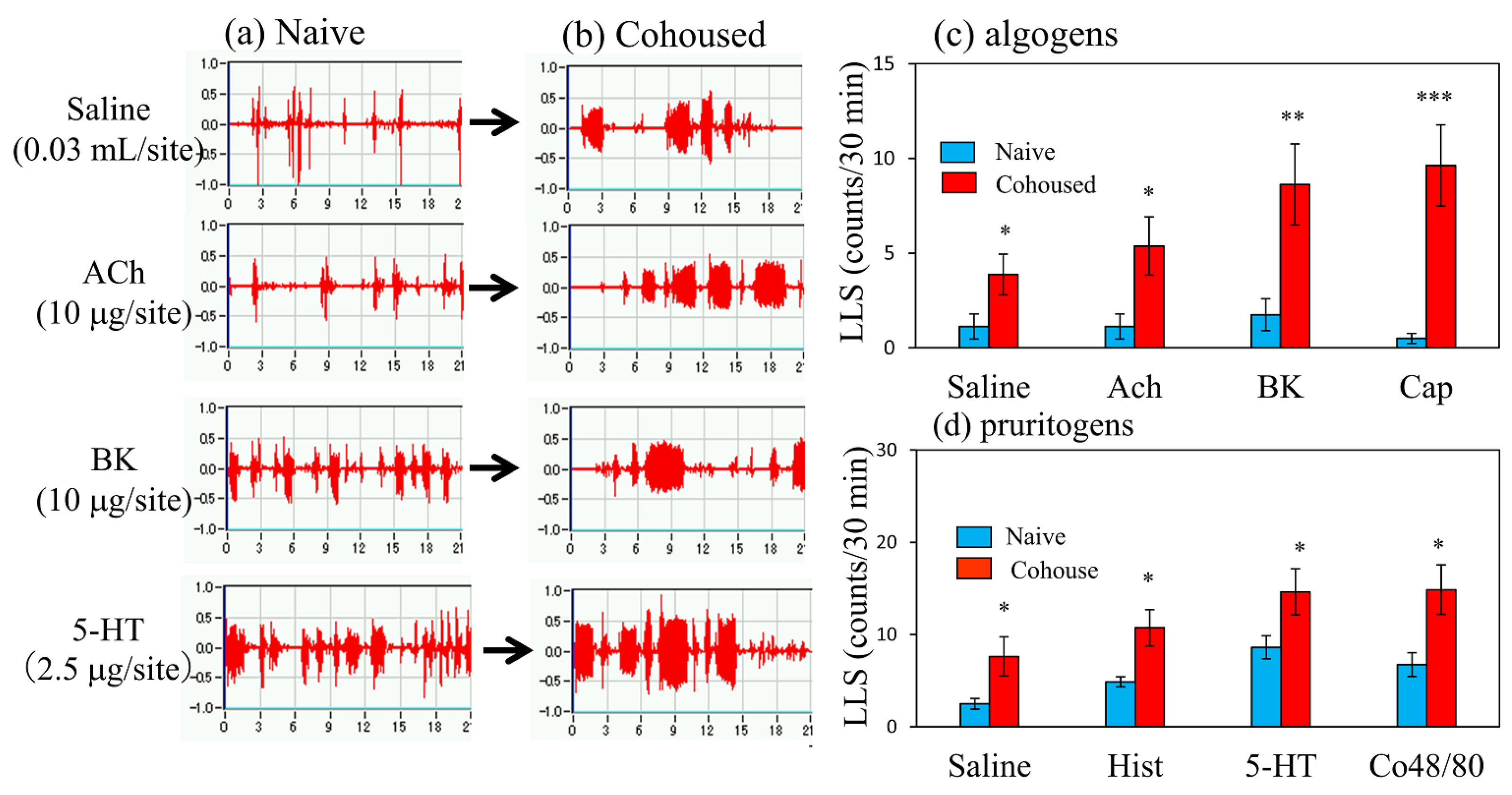
2.3. Effects of Pruritogens and Algogens on LLS Counts in BALB/c Mice Cohoused with Skin-Lesioned NC/Nga Mice
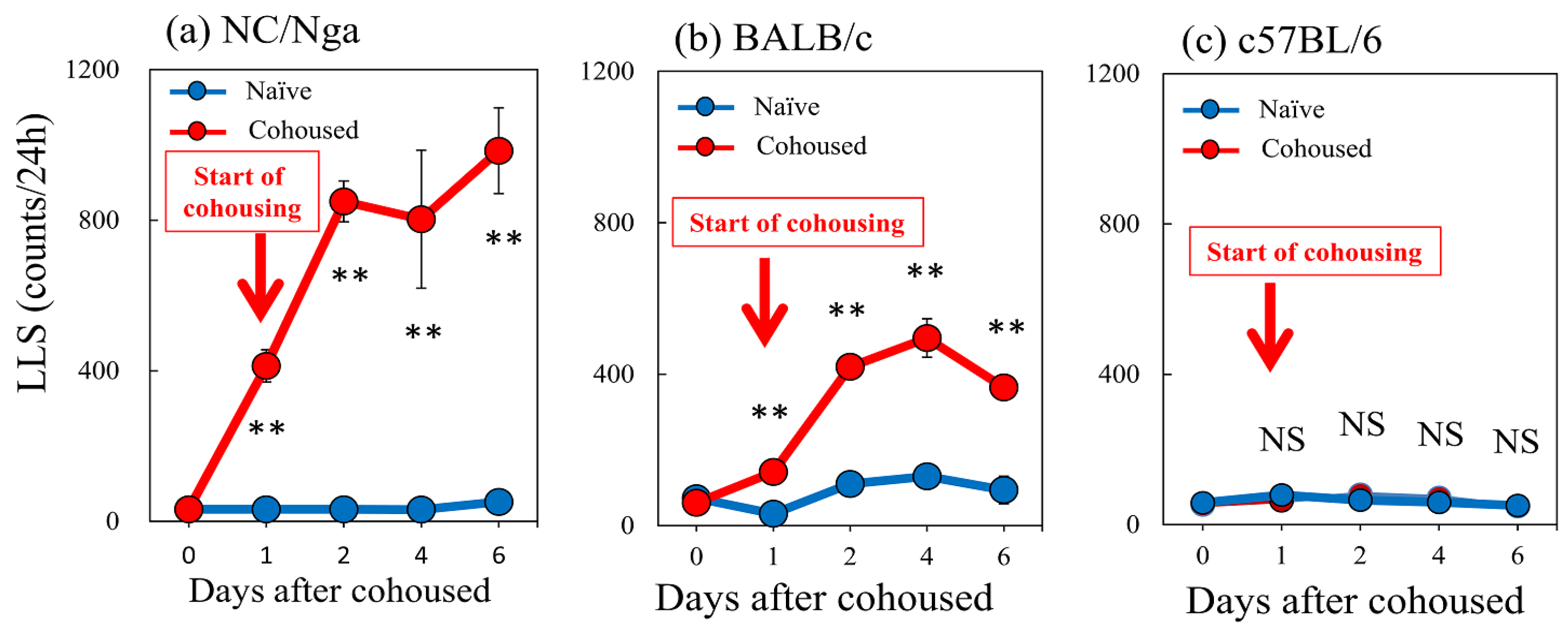
2.4. Effects of Cohousing with Skin-Lesioned NC/Nga Mice on LLS Counts in NC/Nga, BALB/c, and C57BL/6 Mice
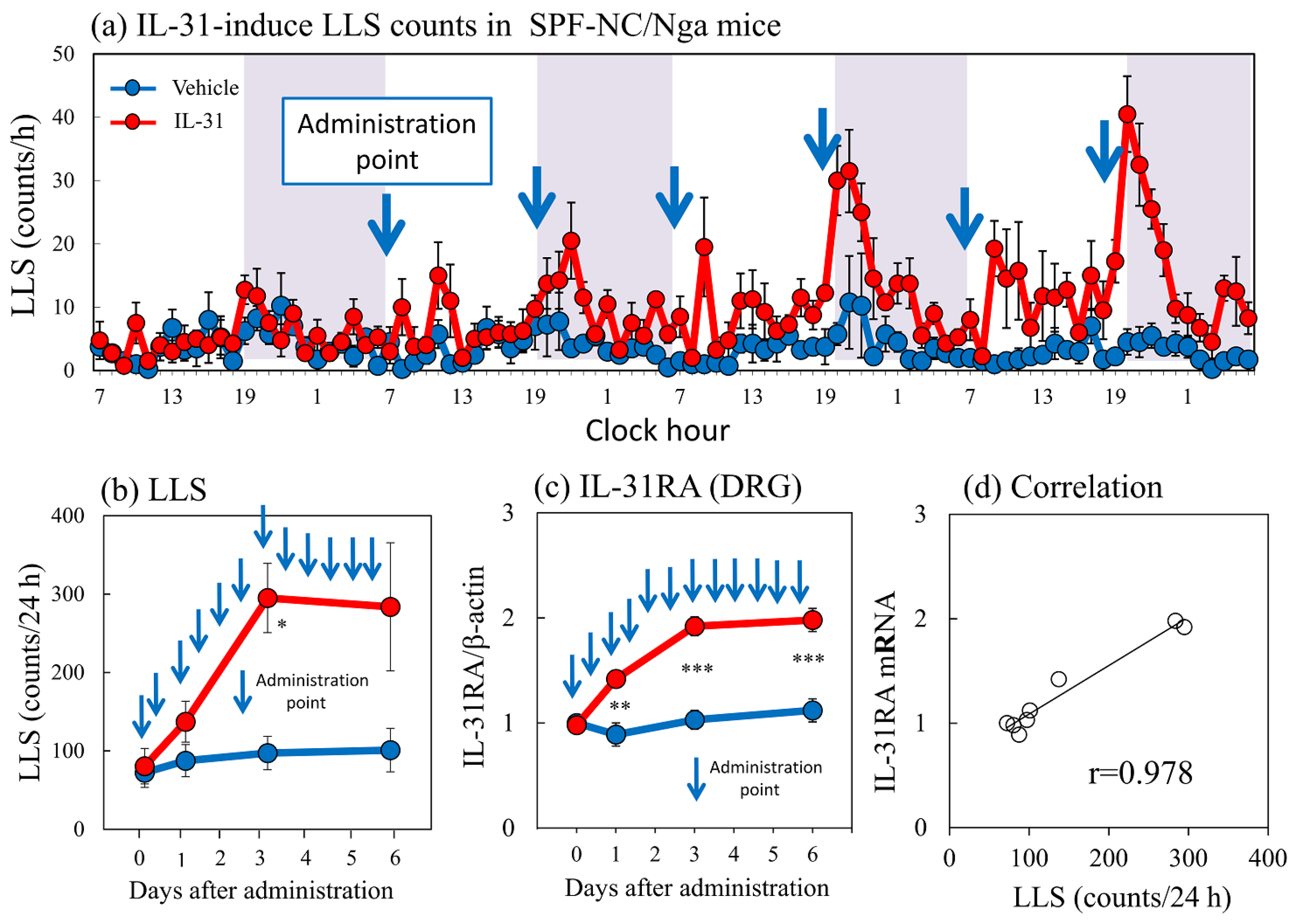
2.5. Effects of Repeated Administration of IL-31 on LLS Counts and DRG Neuronal IL-31RA mRNA Expression in SPF-NC/Nga Mice
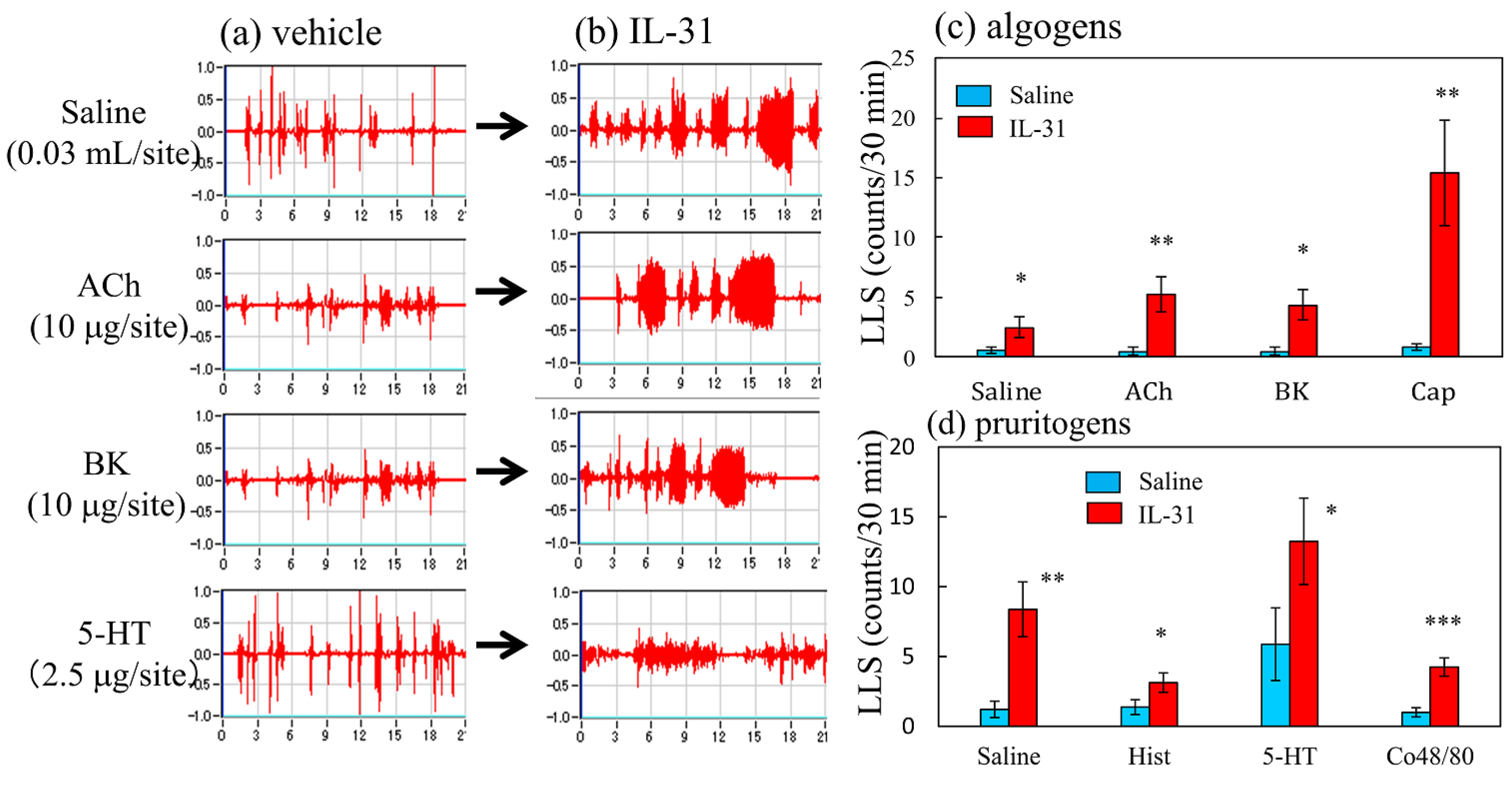
2.6. Effect of IL-31 Pretreatment on Pruritogen- or Algogen-Induced Scratching Behavior in BALB Mice
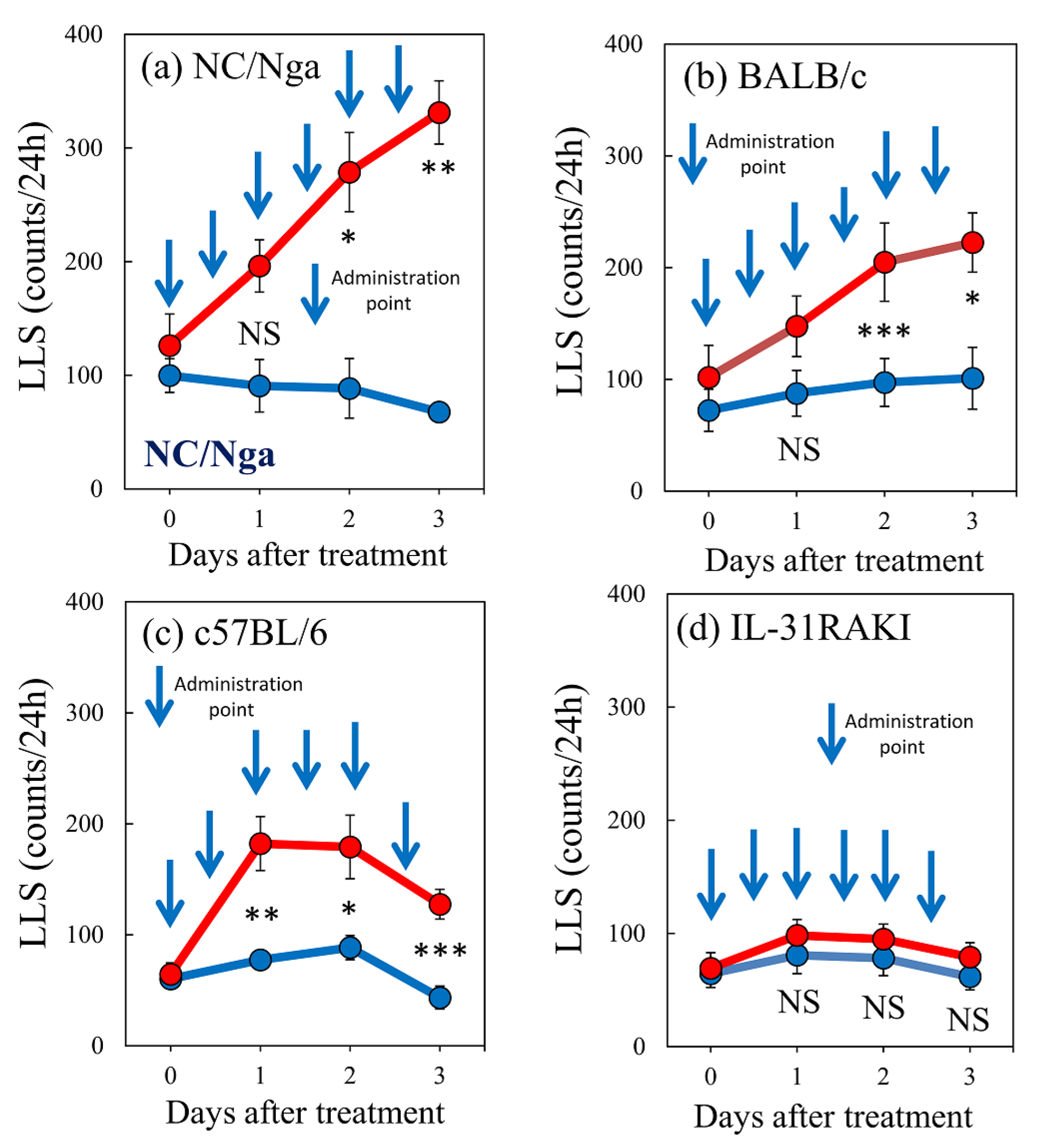
2.7. Effects of Repeated Administration of IL-31 on LLS Counts in NC/Nga, BALB/c, C57BL/6, and IL-31RAKI (IL-31RA-Deficient) Mice
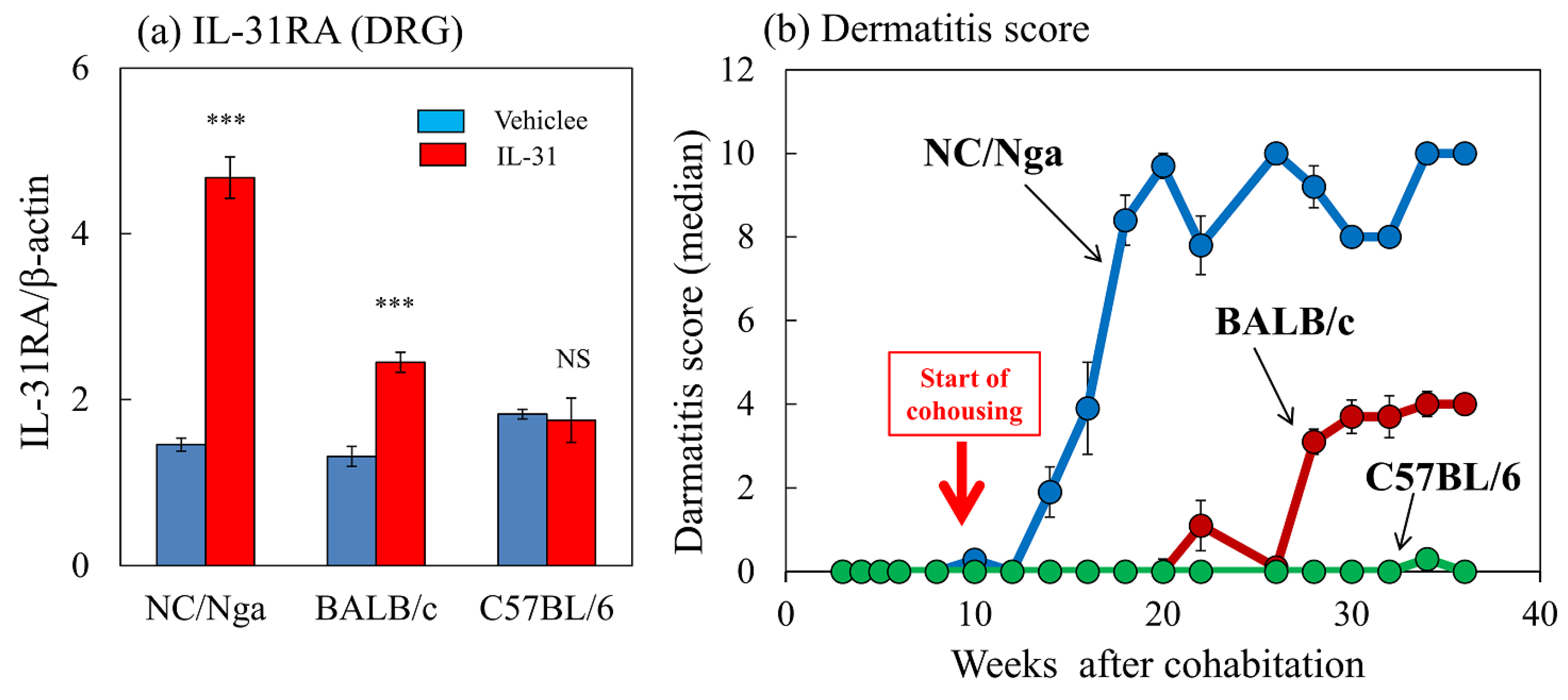
2.8. Effects of Repeated Administration of IL-31 on IL-31RA Expression and Cohousing with Skin-Lesioned NC/Nga Mice-Induced Dermatitis Score in NC/Nga, BALB/c, and C57BL/6 Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagent
4.3. Measurement of Scratching Counts
4.4. Evaluation of Dermatitis Scores
4.5. Induction of Itching and Dermatitis
4.6. Measurement of Compound-Induced Scratch Counts
4.7. Quantitative Real-Time PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matsuda, H.; Watanabe, N.; Geba, G.P.; Sperl, J.; Tsudzuki, M.; Hiroi, J.; Matsumoto, M.; Ushio, H.; Saito, S.; Askenase, P.W.; et al. Development of Atopic Dermatitis-Like Skin Lesion with IgE Hyperproduction in NC/Nga Mice. Int. Immunol. 1997, 9, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Kuraishi, Y.; Nagasawa, T.; Hayashi, K.; Satoh, M. Scratching Behavior Induced by Pruritogenic but Not Algesiogenic Agents in Mice. Eur. J. Pharmacol. 1995, 275, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Takano, N.; Arai, I.; Kurachi, M. Analysis of the Spontaneous Scratching Behavior by NC/Nga Mice: A Possible Approach to Evaluate Antipruritics for Subjects with Atopic Dermatitis. Eur. J. Pharmacol. 2003, 471, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Takano, N.; Arai, I.; Hashimoto, Y.; Kurachi, M. Evaluation of Antipruritic Effects of Several Agents on Scratching Behavior by NC/Nga Mice. Eur. J. Pharmacol. 2004, 495, 159–165. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Arai, I.; Takano, N.; Tanaka, M.; Nakaike, S. Induction of Scratching Behavior and Dermatitis in Various Strains of Mice Cohabiting with NC/Nga Mice Which Had Chronic Dermatitis. Br. J. Dermatol. 2006, 154, 28–33. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Maekawa, T.; Nishikawa, Y.; Nojima, H.; Kaneko, M.; Kawakita, T.; Miyamoto, T.; Kuraishi, Y. Characterization of Itch-Associated Responses of NC/Nga Mice with Mite-Induced Chronic Dermatitis. J. Dermatol. Sci. 2001, 25, 20–28. [Google Scholar] [CrossRef]
- Takano, N.; Arai, I.; Kurachi, M. A Method to Induce Stable Atopic Dermatitis-Like Symptoms in NC/Nga Mice Housed with Skin-Lesioned Mice. Br. J. Dermatol. 2006, 154, 426–430. [Google Scholar] [CrossRef]
- Dillon, S.R.; Sprecher, C.; Hammond, A.; Bilsborough, J.; Rosenfeld-Franklin, M.; Presnell, S.R.; Haugen, H.S.; Maurer, M.; Harder, B.; Johnston, J.; et al. Interleukin-31, a Cytokine Produced by Activated T Cells, Induces Dermatitis in Mice. Nat. Immunol. 2004, 5, 752–760. [Google Scholar] [CrossRef]
- Raap, U.; Wichmann, K.; Bruder, M.; Ständer, S.; Wedi, B.; Kapp, A.; Werfel, T. Correlation of IL-31 Serum Levels with Severity of Atopic Dermatitis. J. Allergy Clin. Immunol. 2008, 122, 421–423. [Google Scholar] [CrossRef]
- Scholzen, T.E.; Steinhoff, M.; Sindrilaru, A.; Schwarz, A.; Bunnett, N.W.; Luger, T.A.; Armstrong, C.A.; Ansel, J.C. Cutaneous Allergic Contact Dermatitis Responses Are Diminished in Mice Deficient in Neurokinin 1 Receptors and Augmented by Neurokinin 2 Receptor Blockage. FASEB J. 2004, 18, 1007–1009. [Google Scholar] [CrossRef]
- Takaoka, A.; Arai, I.; Sugimoto, M.; Yamaguchi, A.; Tanaka, M.; Nakaike, S. Expression of IL-31 Gene Transcripts in NC/Nga Mice with Atopic Dermatitis. Eur. J. Pharmacol. 2005, 516, 180–181. [Google Scholar] [CrossRef]
- Takaoka, A.; Arai, I.; Sugimoto, M.; Honma, Y.; Futaki, N.; Nakamura, A.; Nakaike, S. Involvement of IL-31 on Scratching Behavior in NC/Nga Mice with Atopic-Like Dermatitis. Exp. Dermatol. 2006, 15, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.J.; Yang, H.S.; Kim, E.; Huh, I.S.; Yang, J.M. IL-31 Serum Protein and Tissue mRNA Levels in Patients with Atopic Dermatitis. Ann. Dermatol. 2011, 23, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Ezzat, M.H.; Hasan, Z.E.; Shaheen, K.Y. Serum Measurement of Interleukin-31 (IL-31) in Paediatric Atopic Dermatitis: Elevated Levels Correlate with Severity Scoring. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 334–339. [Google Scholar] [CrossRef]
- Bilsborough, J.; Leung, D.Y.; Maurer, M.; Howell, M.; Boguniewicz, M.; Yao, L.; Storey, H.; LeCiel, C.; Harder, B.; Gross, J.A. IL-31 Is Associated with Cutaneous Lymphocyte Antigen-Positive Skin Homing T Cells in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2006, 117, 418–425. [Google Scholar] [CrossRef]
- Szegedi, K.; Kremer, A.E.; Kezic, S.; Teunissen, M.B.; Bos, J.D.; Luiten, R.M.; Res, P.C.; Middelkamp-Hup, M.A. Increased Frequencies of IL-31-Producing T Cells Are Found in Chronic Atopic Dermatitis Skin. Exp. Dermatol. 2012, 21, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Arai, I.; Tsuji, M.; Miyagawa, K.; Takeda, H.; Akiyama, N.; Saito, S. Repeated Administration of IL-31 Upregulates IL-31 Receptor A (IL-31RA) in Dorsal Root Ganglia and Causes Severe Itch-Associated Scratching Behaviour in Mice. Exp. Dermatol. 2015, 24, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Ikoma, A.; Rukwied, R.; Ständer, S.; Steinhoff, M.; Miyachi, Y.; Schmelz, M. Neuronal Sensitization for Histamine-Induced Itch in Lesional Skin of Patients with Atopic Dermatitis. Arch. Dermatol. 2003, 139, 1455–1458. [Google Scholar] [CrossRef] [Green Version]
- Ikoma, A.; Fartasch, M.; Heyer, G.; Miyachi, Y.; Handwerker, H.; Schmelz, M. Painful Stimuli Evoke Itch in Patients with Chronic Pruritus: Central Sensitization for Itch. Neurology 2004, 62, 212–217. [Google Scholar] [CrossRef]
- Ikoma, A.; Steinhoff, M.; Ständer, S.; Yosipovitch, G.; Schmelz, M. The Neurobiology of Itch. Nat. Rev. Neurosci. 2006, 7, 535–547. [Google Scholar] [CrossRef]
- Takaoka, A.; Arai, I.; Sugimoto, M.; Futaki, N.; Sakurai, T.; Honma, Y.; Nakaike, S. Role of Scratch-Induced Cutaneous Prostaglandin D Production on Atopic-Like Scratching Behaviour in Mice. Exp. Dermatol. 2007, 16, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Arai, I.; Tsuji, M.; Miyagawa, K.; Takeda, H.; Akiyama, N.; Saito, S. Interleukin-31 (L-31) Causes Alloknesis: Pain-Stimulation Becomes Itch-Stimulation in Mouse Skin. Itch Pain 2015, 2, e924. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, W.; Yourey, P.A.; Gohari, S.; Zukauskas, D.; Zhang, J.; Ruben, S.; Alderson, R.F. Computational EST Database Analysis Identifies A Novel Member of the Neuropoietic Cytokine Family. Biochem. Biophys. Res. Commun. 1999, 262, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Diveu, C.; Lelièvre, E.; Perret, D.; Lak-Hal, A.H.; Froger, J.; Guillet, C.; Chevalier, S.; Rousseau, F.; Wesa, A.; Preisser, L.; et al. GPL, a Novel Cytokine Receptor Related to GP130 and Leukemia Inhibitory Factor Receptor. J. Biol. Chem. 2003, 278, 49850–49859. [Google Scholar] [CrossRef] [Green Version]
- Taga, T.; Kishimoto, T. Gp130 and the Interleukin-6 Family of Cytokines. Annu. Rev. Immunol. 1997, 15, 797–819. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Muller, A.; Lauerma, A.I.; Pivarcsi, A.; Soto, H.; Kemeny, L.; Alenius, H.; Dieu-Nosjean, M.C.; Meller, S.; Rieker, J.; et al. IL-31: A New Link Between T Cells and Pruritus in Atopic Skin Inflammation. J. Allergy Clin. Immunol. 2006, 117, 411–417. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Greaves, M.W.; Schmelz, M. Itch. Lancet 2003, 361, 690–694. [Google Scholar] [CrossRef]
- Hanifin, J.M.; Rajka, G. Diagnostic Features of Atopic Dermatitis. Acta Derm. Venereol. 1980, 92, 44–47. [Google Scholar]
- Kimura, T.; Miyazawa, H. The ‘Butterfly’ Sign in Patients with Atopic Dermatitis: Evidence for the Role of Scratching in the Development of Skin Manifestations. J. Am. Acad. Dermatol. 1989, 21, 579–580. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Arai, I.; Nakanishi, Y.; Sakurai, T.; Nakamura, A.; Nakaike, S. Scratching of Their Skin by NC/Nga Mice Leads to Development of Dermatitis. Life Sci. 2004, 76, 783–794. [Google Scholar] [CrossRef]
- Arai, I.; Tsuji, M.; Takeda, H.; Akiyama, N.; Saito, S. Increased Itching Sensation Depends on an Increase in the Neuronal IL-31 Receptor A (IL-31RA) Expression in Mice with Atopic-Like Dermatitis. Itch Pain 2014, 1, 1–10. [Google Scholar] [CrossRef]
- Arai, I.; Tsuji, M.; Takeda, H.; Akiyama, N.; Saito, S. A Single Dose of Interleukin-31 (IL-31) Causes Continuous Itch-Associated scratching Behavior in Mice. Exp. Dermatol. 2013, 22, 669–671. [Google Scholar] [CrossRef]
- Akiyama, T.; Merrill, A.W.; Carstens, M.I.; Carstens, E. Activation of Superficial Dorsal Horn Neurons in the Mouse by a PAR-2 Agonist and 5-HT: Potential Role in Itch. J. Neurosci. 2009, 29, 6691–6699. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, N.; Igeta, K.; Kim, J.F.; Nagao, M.; Shiraishi, N.; Nakamura, N.; Nagai, H. Involvement of Unique Mechanisms in the Induction of Scratching Behavior in BALB/C Mice by Compound 48/80. Eur. J. Pharmacol. 2002, 448, 175–183. [Google Scholar] [CrossRef]
- Wahlgren, C.F. Itch and Atopic Dermatitis: An Overview. J. Dermatol. 1999, 26, 770–779. [Google Scholar] [CrossRef]
- Shimada, S.G.; LaMotte, R.H. Behavioral Differentiation Between Itch and Pain in Mice. Pain 2008, 139, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Hosogi, M.; Schmelz, M.; Miyachi, Y.; Ikoma, A. Bradykinin Is a Potent Pruritogen in Atopic Dermatitis: A Switch from Pain to Itch. Pain 2006, 126, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Vogelsang, M.; Heyer, G.; Hornstein, O.P. Acetylcholine Induced Different Cutaneous Sensations in Atopic and Non-atopic Subjects. Acta Derm. Venereol. 1995, 75, 434–436. [Google Scholar] [CrossRef]
- Grimstad, O.; Sawanobori, Y.; Vestergaard, C.; Bilsborough, J.; Olsen, U.B.; Grønhøj-Larsen, C.; Matsushima, K. Anti-Interleukin-31-Antibodies Ameliorate Scratching Behaviour in NC/Nga Mice: A Model of Atopic Dermatitis. Exp. Dermatol. 2009, 18, 35–43. [Google Scholar] [CrossRef]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A Sensory Neuron-Expressed IL-31 Receptor Mediates T Helper Cell-Dependent Itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2014, 133, 448–460. [Google Scholar] [CrossRef] [Green Version]
- Arai, I.; Tsuji, M.; Takeda, H.; Akiyama, N.; Saito, S. Capsaicin Suppresses Interleukin-31-Induced Itching Partially Involved in Inhibiting the Expression of Dorsal Root Ganglion Interleukin-31 Receptor A in Male Mice. Neurobiol. Pain 2022, 29, 100088. [Google Scholar] [CrossRef] [PubMed]
- Pereira, U.; Boulais, N.; Lebonvallet, N.; Pennec, J.P.; Dorange, G.; Misery, L. Mechanisms of the Sensory Effects of Tacrolimus on the Skin. Br. J. Dermatol. 2010, 163, 70–77. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The Capsaicin Receptor: A Heat-Activated Ion Channel in the Pain Pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Saito, S.; Aoki, A.; Arai, I.; Takaishi, S.; Ito, H.; Akiyama, N.; Kiyonari, H. Regulation of Th2 Responses by Different Cell Types Expressing the Interleukin-31 Receptor. Allergy Asthma Clin. Immunol. 2017, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, N.; Ohno, Y.; Fukuda, T.; Manome, Y.; Saito, S. Enhancing Activity of N-Glycosylation for Constitutive Proteins Secretions in Non-polarized Cells. Biochem. Biophys. Res. Commun. 2009, 381, 612–618. [Google Scholar] [CrossRef]
- Arai, I.; Tsuji, M.; Miyagawa, K.; Takeda, H.; Akiyama, N.; Saito, S. Prostaglandin D2 Improves IL-31-Induced Alloknesis: Itch-Stimulation Becomes Pain-Stimulation in Mouse Skin. Itch Pain 2016, 3, e1138. [Google Scholar] [CrossRef]
- Inagaki, N.; Igeta, K.; Shiraishi, N.; Kim, J.F.; Nagao, M.; Nakamura, N.; Nagai, H. Evaluation and Characterization of Mouse Scratching Behavior by a New Apparatus, MicroAct. Skin Pharmacol. Appl. Skin Physiol. 2003, 16, 165–175. [Google Scholar] [CrossRef]
- Nagai, H.; Matsuo, A.; Hiyama, H.; Inagaki, N.; Kawada, K. Immunoglobulin E Production in Mice by Means of Contact Sensitization with a Simple Chemical, Hapten. J. Allergy Clin. Immunol. 1997, 100, S39–S44. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arai, I.; Saito, S. Interleukin-31 Receptor A Expression in the Dorsal Root Ganglion of Mice with Atopic Dermatitis. Int. J. Mol. Sci. 2023, 24, 1047. https://doi.org/10.3390/ijms24021047
Arai I, Saito S. Interleukin-31 Receptor A Expression in the Dorsal Root Ganglion of Mice with Atopic Dermatitis. International Journal of Molecular Sciences. 2023; 24(2):1047. https://doi.org/10.3390/ijms24021047
Chicago/Turabian StyleArai, Iwao, and Saburo Saito. 2023. "Interleukin-31 Receptor A Expression in the Dorsal Root Ganglion of Mice with Atopic Dermatitis" International Journal of Molecular Sciences 24, no. 2: 1047. https://doi.org/10.3390/ijms24021047
APA StyleArai, I., & Saito, S. (2023). Interleukin-31 Receptor A Expression in the Dorsal Root Ganglion of Mice with Atopic Dermatitis. International Journal of Molecular Sciences, 24(2), 1047. https://doi.org/10.3390/ijms24021047