
Comprehensive Analysis of Metabolome and Transcriptome in Fruits and Roots of Kiwifruit


Abstract
:1. Introduction
2. Results
2.1. Difference in Flesh Color among Four Kiwifruit Varieties
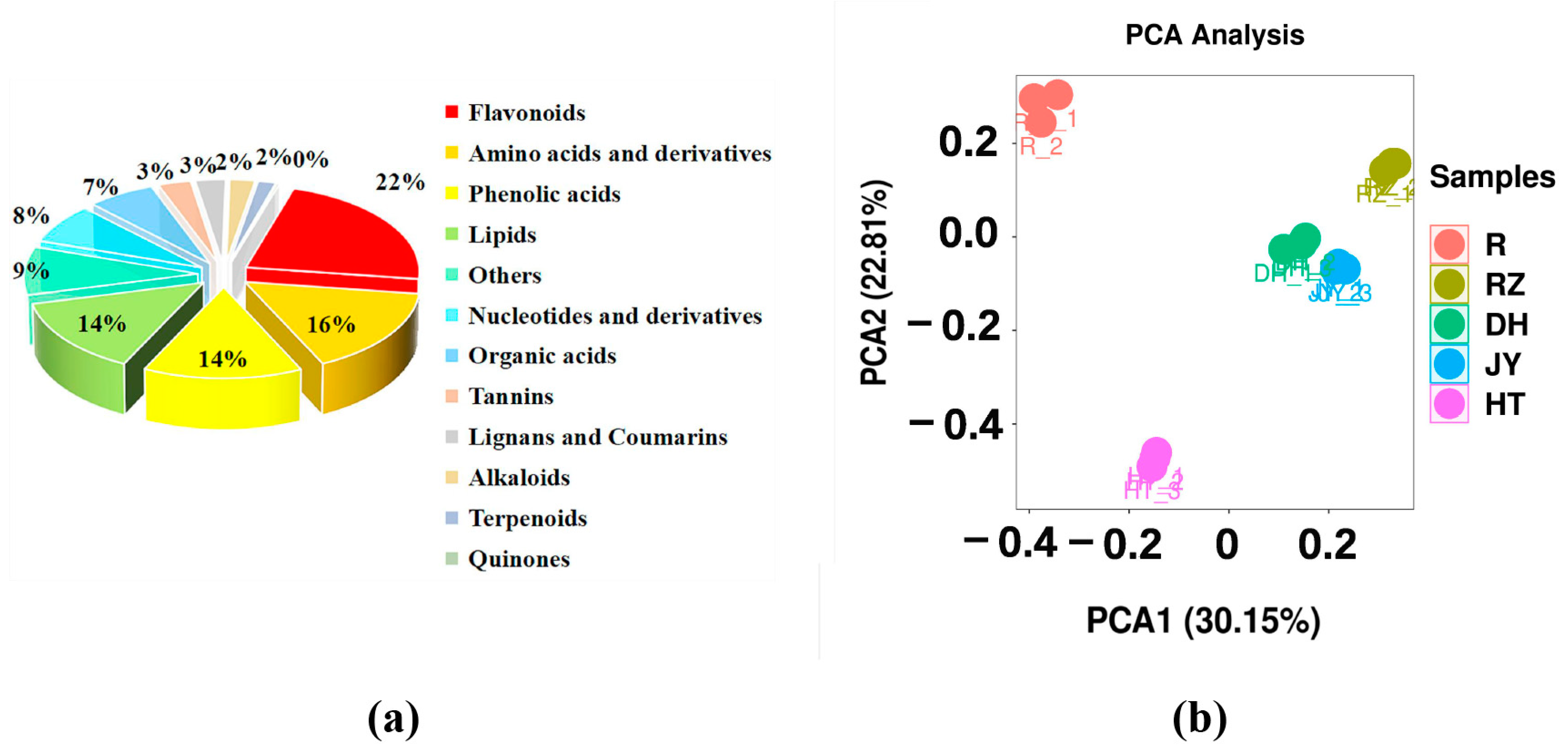
2.2. The Metabolome Profiling of Fruit and Root in Kiwifruit
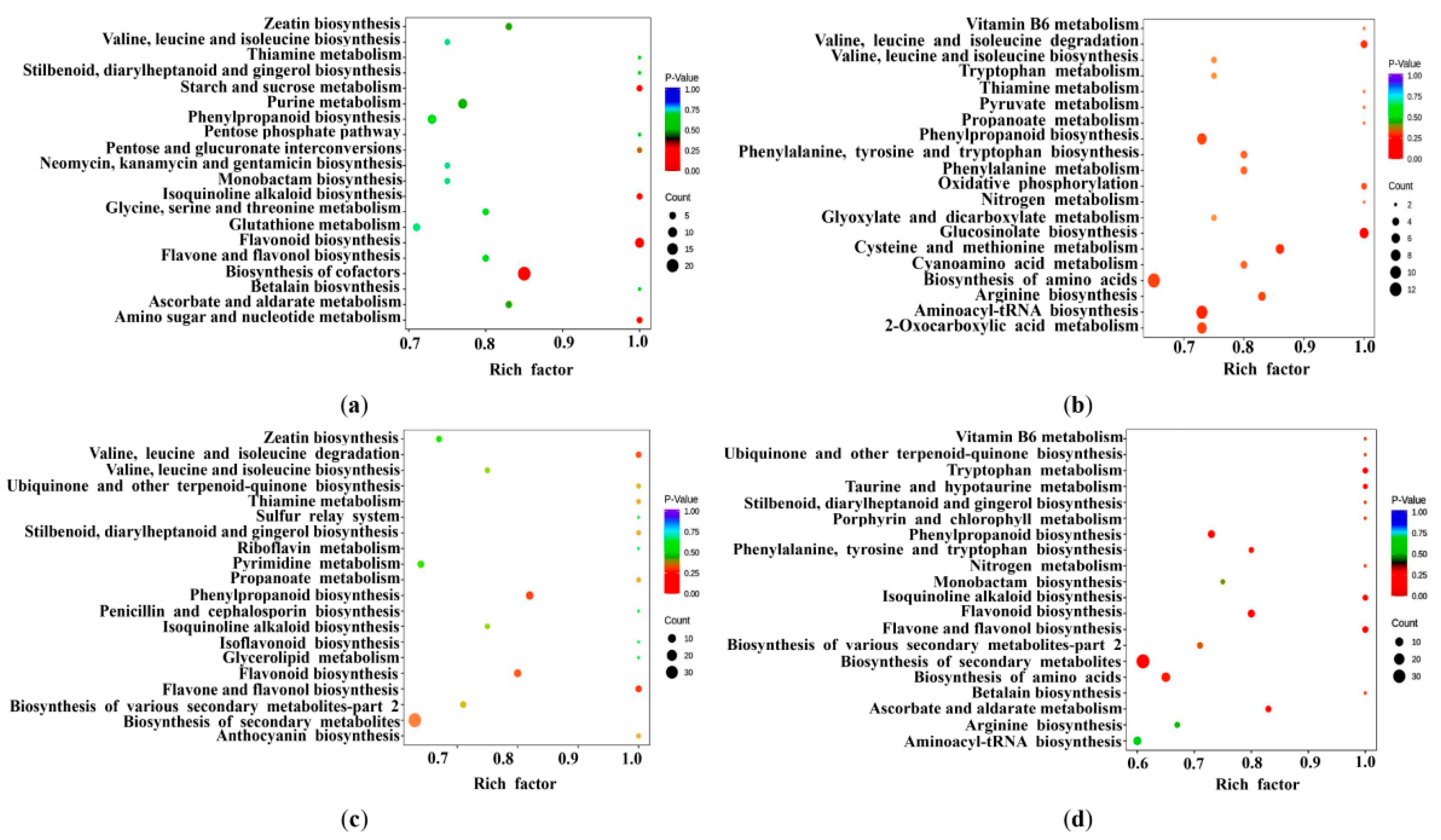
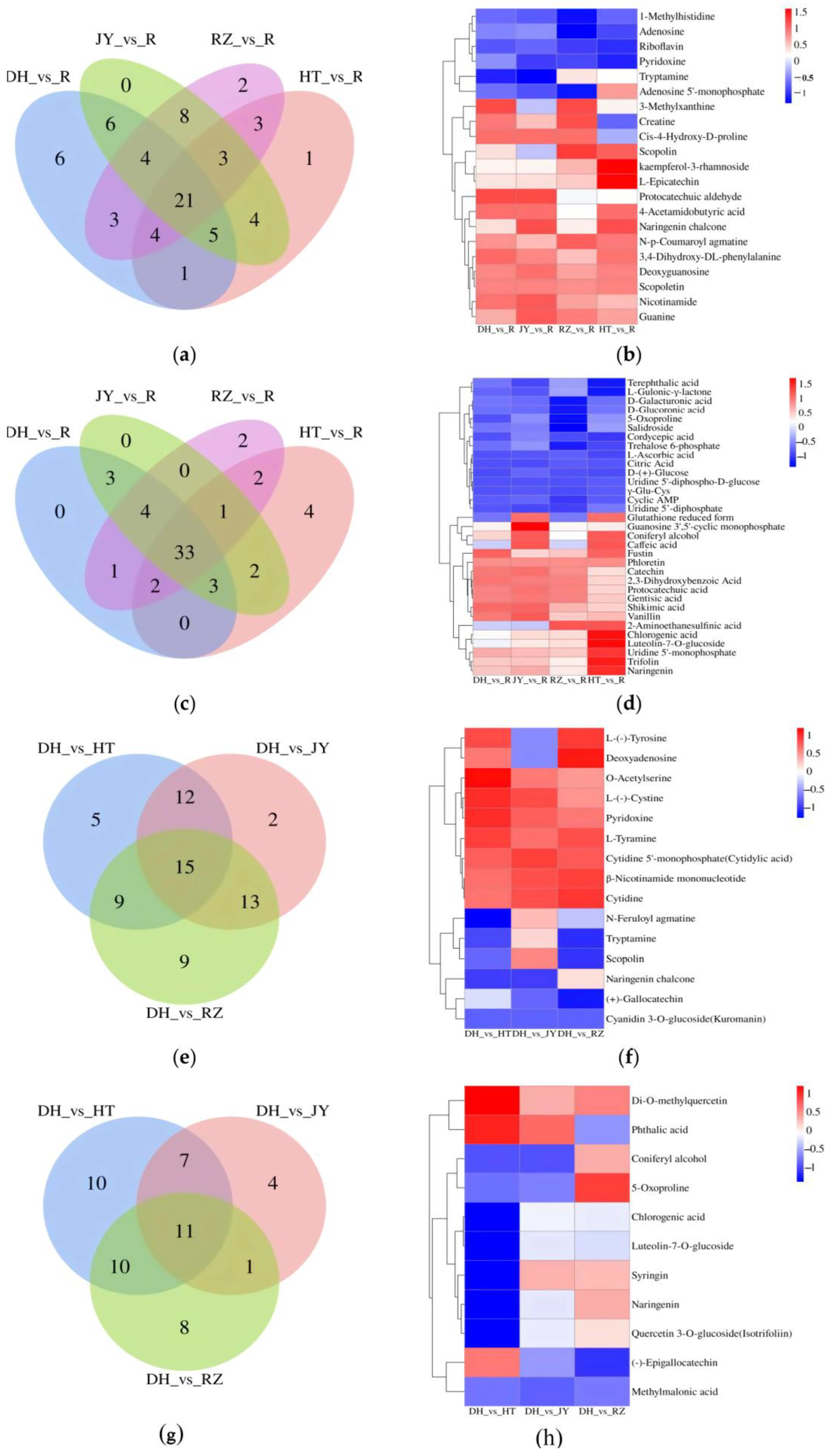
2.3. Identification of Differentially Expressed Metabolites (DEMs)
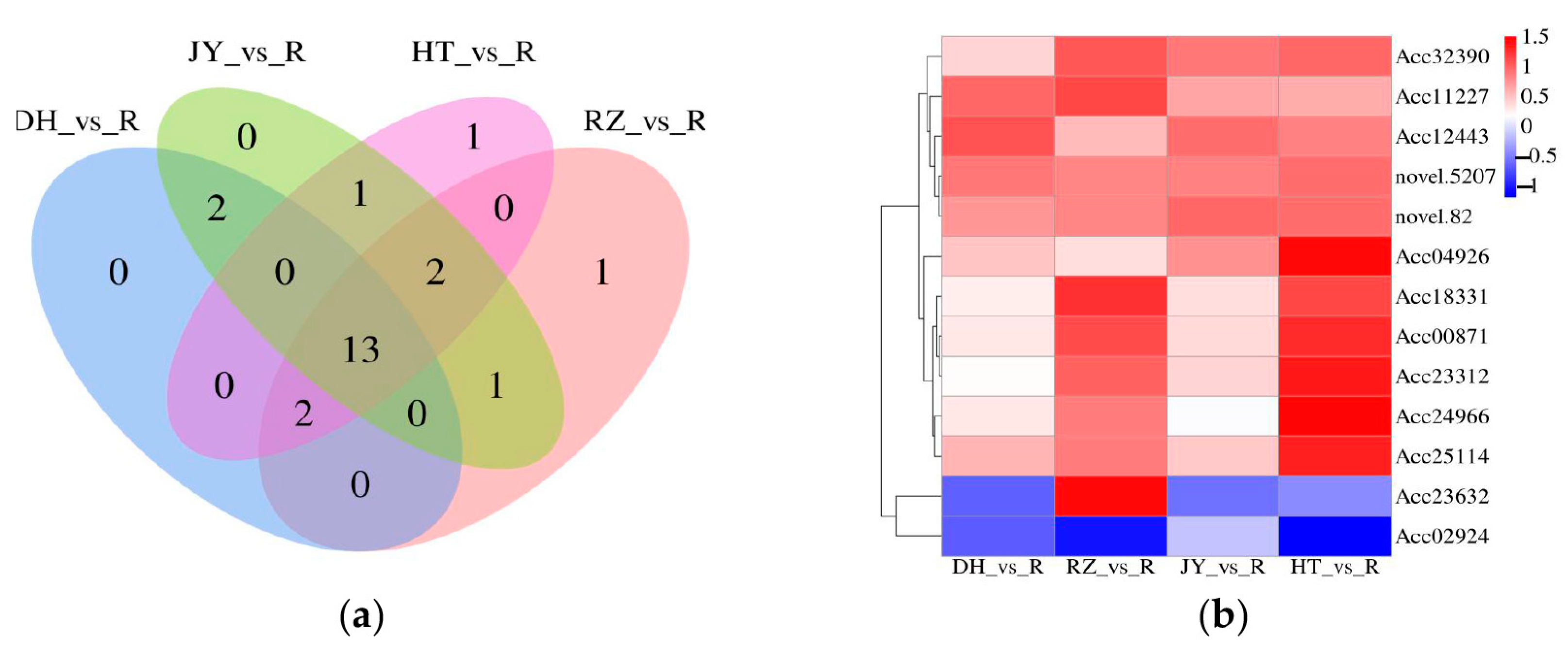
2.4. Identification of Differentially Expressed Genes (DEGs)
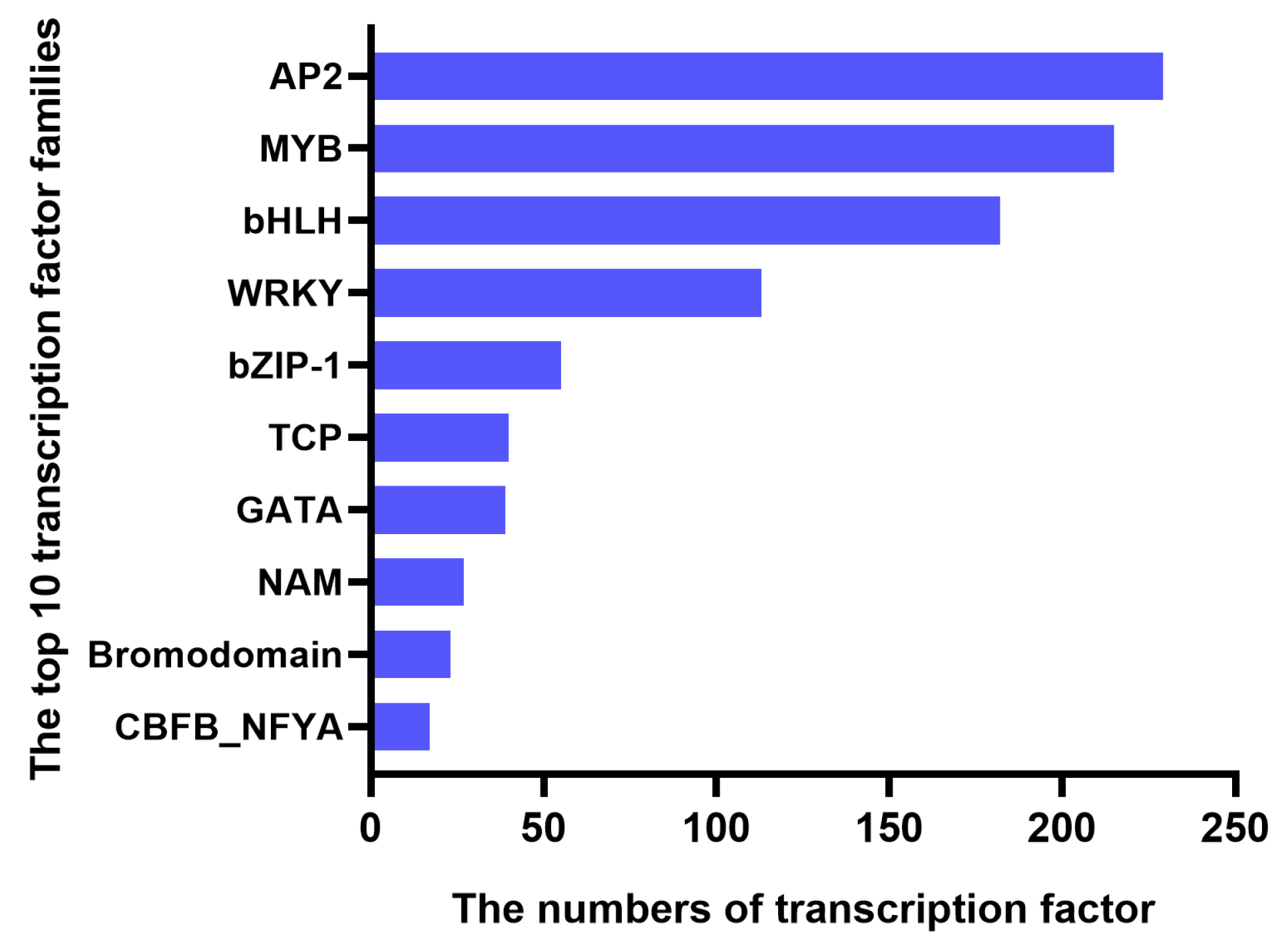
2.5. Functional Classification of DEGs
2.6. Analysis of Association between DEMs and DEGs
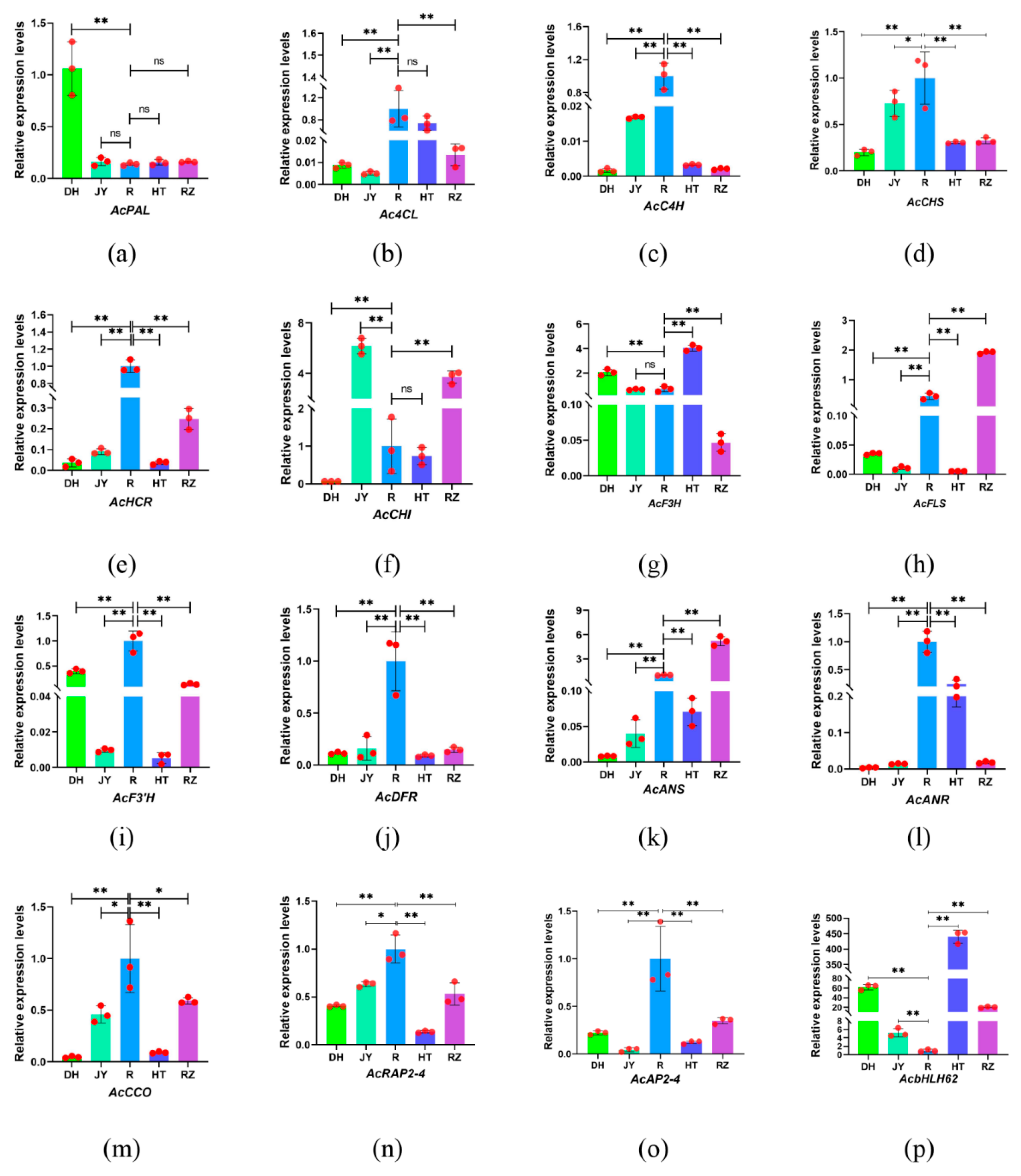
2.7. Validation of Genes Related to Flavonoid Biosynthesis Using RT-qPCR
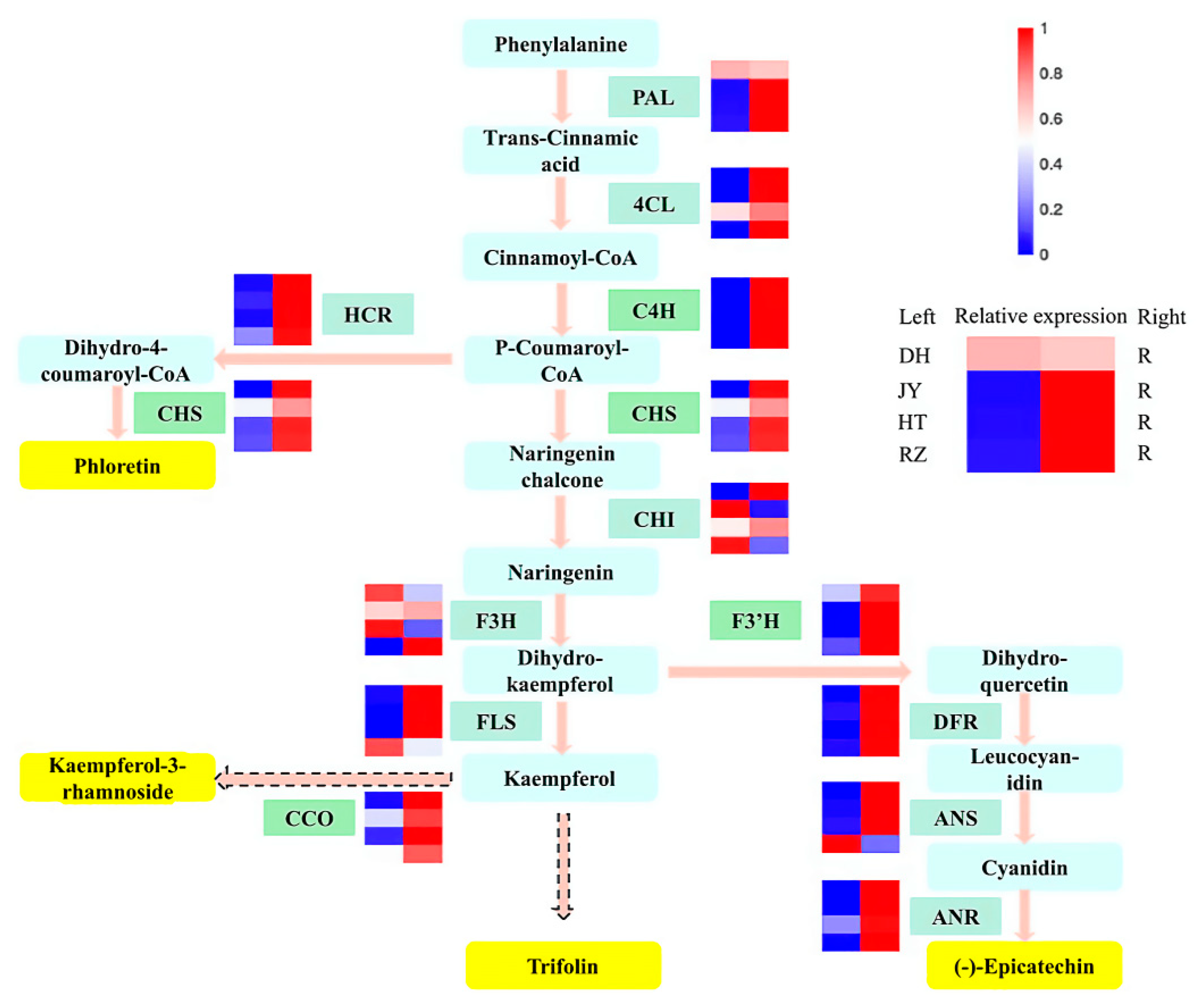
2.8. Genes Related to Biosynthesis of Dihydrochalcones and Flavonols Were Activated in Kiwifruit Roots
3. Discussion
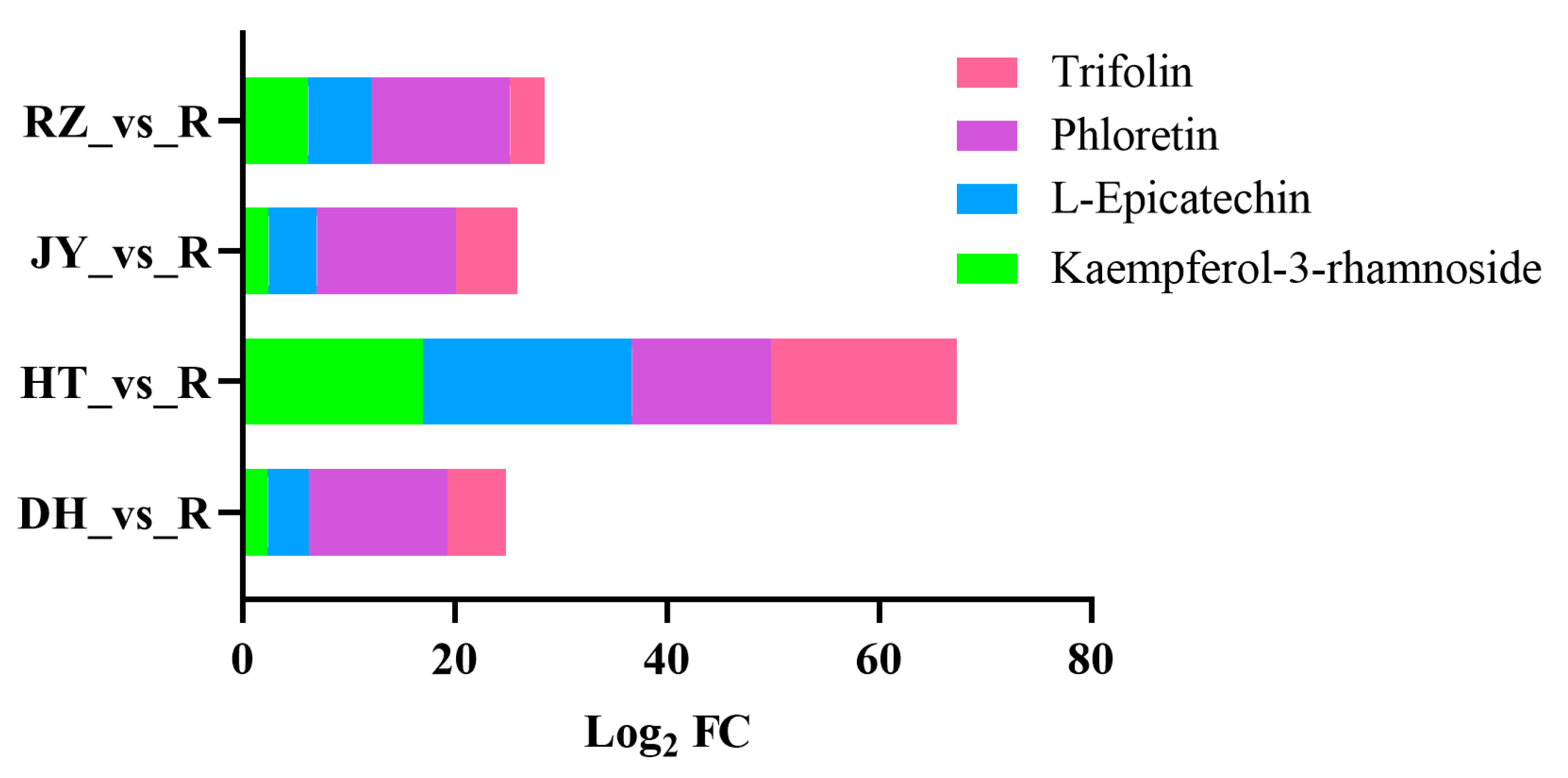
3.1. DEMs between Kiwifruit Roots and Fruits Were Enriched in Dihydrochalcones and Flavonols
3.2. Metabolome Profiling Is an Efficient Approach to Estimate the Genetic Relationship between Cultivars
3.3. The Biosynthesis of Dihydrochalcones and Flavonols in Kiwifruit Roots Is Regulated at the Transcriptional Level
4. Materials and Methods
4.1. Plant Materials
4.2. Metabolite Extraction and Parameter Setting
4.3. RNA Extraction and Sequencing
4.4. Analysis of TFs and DEGs
4.5. Real-Time Quantitative PCR (RT-qPCR)
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, H.W.; Ferguson, A. Actinidia in China: Natural diversity, phylogeographical evolution, interspecific gene flow and kiwifruit cultivar improvement. Acta Hortic. 2007, 753, 31–40. [Google Scholar] [CrossRef]
- Nishiyama, I.; Fukuda, T.; Shimohashi, A.; Oota, T. Sugar and organic acid composition in the fruit juice of different Actinidia varieties. Food Sci. Technol. Res. 2008, 14, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Fisk, C.L.; McDaniel, M.R.; Strik, B.C.; Zhao, Y. Physicochemical, sensory, and nutritive qualities of hardy kiwifruit (Actinidia arguta ‘Ananasnaya’) as affected by harvest maturity and storage. J. Food Sci. 2006, 71, S204–S210. [Google Scholar] [CrossRef]
- Huang, H.W. Chapter 2—Species. In Kiwifruit: The Genus Actinidia, 1st ed.; Huang, H.W., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 45–167. [Google Scholar]
- Zhong, C.H.; Wang, S.M.; Jiang, Z.W.; Huang, H.W. ‘Jinyan’, an interspecific hybrid kiwifruit with brilliant yellow flesh and good storage quality. HortScience 2012, 47, 1187–1190. [Google Scholar] [CrossRef] [Green Version]
- Drummond, L.; Gearry, R.B. Kiwifruit modulation of gastrointestinal motility. Adv. Food Nutr. Res. 2013, 68, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, A.R.; Ferguson, L.R. Are kiwifruit really good for you? Acta Hortic. 2003, 610, 131–138. [Google Scholar] [CrossRef]
- Monro, J.; Bentley-Hewitt, K.; Mishra, S. Kiwifruit Exchanges for Increased Nutrient Richness with Little Effect on Carbohydrate Intake, Glycaemic Impact, or Insulin Response. Nutrients 2018, 10, 1710. [Google Scholar] [CrossRef] [Green Version]
- Ward, C.; Courtney, D. Kiwifruit: Taking its place in the global fruit bowl. Adv. Food Nutr. Res. 2013, 68, 1–14. [Google Scholar] [CrossRef]
- Singletary, K. Kiwifruit: Overview of potential health benefits. Nutr. Today 2012, 47, 133–147. [Google Scholar] [CrossRef]
- Stonehouse, W.; Gammon, C.S.; Beck, K.L.; Conlon, C.A.; von Hurst, P.R.; Kruger, R. Kiwifruit: Our daily prescription for health. Can. J. Physiol. Pharmacol. 2013, 91, 442–447. [Google Scholar] [CrossRef]
- Wang, X.J.; Miao, L.P.; Wu, T.; Shen, L.H. Advances in studies on chemical constituents in roots of Actinidia chinensis and their pharmacological activities. Chin. Tradit. Herb. Drugs 2012, 42, 1233–1240. [Google Scholar]
- Wang, M.X.; Wang, T.Y.; Bai, N.S. Research progress in chemical components and anticancer pharmacological effects of root of Actinidia arguta (Sieb. et Zucc) planch. ex Miq. Chin. J. Inf. Tradit. Chin. Med. 2019, 26, 137–140. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wu, J.Z.; Wu, J.G. Studies on the purification process of triterpenoid from the root of Actinidia eriantha Benth. Strait Pharm. J. 2017, 29, 29–32. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Du, J.P.; Ji, C.S.; Jiang, F.S.; Wang, G. Study in Yangtao Actinidia Root combineing chemotherapy in treatment of advanced gastric cancer. Nei Mongol J. Tradit. Chin. Med. 2010, 29, 2. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Li, G.; Dai, B.; Tan, W. Actinidia chinensis Planch. Improves the Indices of Antioxidant and Anti-Inflammation Status of Type 2 Diabetes Mellitus by Activating Keap1 and Nrf2 via the Upregulation of MicroRNA-424. Oxid. Med. Cell. Longev. 2017, 2017, 7038789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.; Chen, M.; Niu, J.; Qu, N.; Ji, B.; Duan, X.; Liu, Z.; Liu, X.; Wang, Y.; Zhao, B. The Manufacturing Process of Kiwifruit Fruit Powder with High Dietary Fiber and Its Laxative Effect. Molecules 2019, 24, 3813. [Google Scholar] [CrossRef] [Green Version]
- Eady, S.L.; Wallace, A.J.; Butts, C.A.; Hedderley, D.; Drummond, L.; Ansell, J.; Gearry, R.B. The effect of ‘Zesy002’ kiwifruit (Actinidia chinensis var. chinensis) on gut health function: A randomised cross-over clinical trial. J. Nutr. Sci. 2019, 8, e18. [Google Scholar] [CrossRef] [Green Version]
- Bayer, S.B.; Gearry, R.B.; Drummond, L.N. Putative mechanisms of kiwifruit on maintenance of normal gastrointestinal function. Crit. Rev. Food Sci. Nutr. 2018, 58, 2432–2452. [Google Scholar] [CrossRef] [Green Version]
- Svendsen, M.; Tonstad, S.; Heggen, E.; Pedersen, T.R.; Seljeflot, I.; Bøhn, S.K.; Bastani, N.E.; Blomhoff, R.; Holme, I.M.; Klemsdal, T.O. The effect of kiwifruit consumption on blood pressure in subjects with moderately elevated blood pressure: A randomized, controlled study. Blood Press. 2015, 24, 48–54. [Google Scholar] [CrossRef]
- Xia, H.; Zhou, Y.; Lin, Z.; Guo, Y.; Liu, X.; Wang, T.; Wang, J.; Deng, H.; Lin, L.; Deng, Q.; et al. Characterization and functional validation of β-carotene hydroxylase AcBCH genes in Actinidia chinensis. Hortic. Res. 2022, 9, uhac063. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fang, J.; Qi, X.; Lin, M.; Zhong, Y.; Sun, L.; Cui, W. Combined Analysis of the Fruit Metabolome and Transcriptome Reveals Candidate Genes Involved in Flavonoid Biosynthesis in Actinidia arguta. Int. J. Mol. Sci. 2018, 19, 1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, G.; Liu, Q.; Xu, X.; He, Y.; Li, Y.; Wang, H.; Ye, B.; Huang, C.; Zhong, M.; Jia, D. Metabolome and Transcriptome Reveal Novel Formation Mechanism of Early Mature Trait in Kiwifruit (Actinidia eriantha). Front. Plant Sci. 2021, 12, 760496. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, M.; McGhie, T.K.; Costa, G.; Ferguson, A.R. Pigments in the fruit of red-fleshed kiwifruit (Actinidia chinensis and Actinidia deliciosa). J. Agric. Food Chem. 2005, 53, 9526–9530. [Google Scholar] [CrossRef]
- McGhie, T.K. Secondary metabolite components of kiwifruit. Adv. Food Nutr. Res. 2013, 68, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Wolber, F.M.; Beck, K.L.; Conlon, C.A.; Kruger, M.C. Kiwifruit and mineral nutrition. Adv. Food Nutr. Res. 2013, 68, 233–256. [Google Scholar] [CrossRef]
- Wang, R.; Shu, P.; Zhang, C.; Zhang, J.; Chen, Y.; Zhang, Y.; Du, K.; Xie, Y.; Li, M.; Ma, T.; et al. Integrative analyses of metabolome and genome-wide transcriptome reveal the regulatory network governing flavor formation in kiwifruit (Actinidia chinensis). New Phytol. 2022, 233, 373–389. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, S.; Dai, M.; Wang, M.; Feng, X.Y. Research process in biosynthesis pathway and metabolic regulation of phenolic acid. Food Sci. 2017, 39, 286–293. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Oliveras, M.J.; Quesada, J.; Rufián-Henares, J.A.; Pastoriza, S. Relationship between composition and bioactivity of persimmon and kiwifruit. Food Res. Int. 2018, 105, 461–472. [Google Scholar] [CrossRef]
- Capitani, D.; Mannina, L.; Proietti, N.; Sobolev, A.P.; Tomassini, A.; Miccheli, A.; Di Cocco, M.E.; Capuani, G.; De Salvador, F.R.; Delfini, M. Metabolic profiling and outer pericarp water state in Zespri, CI.GI, and Hayward kiwifruits. J. Agric. Food Chem. 2013, 61, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.P.; Ansell, J.; Drummond, L.N. The nutritional and health attributes of kiwifruit: A review. Eur. J. Nutr. 2018, 57, 2659–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Yan, P.; Du, K.; Li, M.; Xie, Y.; Gao, P. Nutritional component analyses of kiwifruit in different development stages by metabolomic and transcriptomic approaches. J. Sci. Food Agric. 2020, 100, 2399–2409. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fang, J.; Qi, X.; Lin, M.; Zhong, Y.; Sun, L. A key structural gene, AaLDOX, is involved in anthocyanin biosynthesis in all red-fleshed kiwifruit (Actinidia arguta) based on transcriptome analysis. Gene 2018, 648, 31–41. [Google Scholar] [CrossRef]
- Wang, S.H. The Research on Extraction, Purification and Liver-Protective Function of Active Ingredient from Kiwifruit Roots. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2008. [Google Scholar] [CrossRef]
- Miller, K.D.; Strommer, J.; Taylor, L.P. Conservation in divergent solanaceous species of the unique gene structure and enzyme activity of a gametophytically-expressed flavonol 3-O-galactosyltransferase. Plant Mol. Biol. 2002, 48, 233–242. [Google Scholar] [CrossRef]
- Rezk, B.M.; Haenen, G.R.; van der Vijgh, W.J.; Bast, A. The antioxidant activity of phloretin: The disclosure of a new antioxidant pharmacophore in flavonoids. Biochem. Biophys. Res. Commun. 2002, 295, 9–13. [Google Scholar] [CrossRef]
- Mei, Q.G.; Yuan, W.C.; Wang, C. Progress in the synthesis of 3-Hydroxyflavones. Chin. J. Org. Chem. 2015, 35, 70–84. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Meng, L.C. Research progress of the naturally occurring dihydrochalcones compounds. Ningxia Eng. Technol. 2007, 6, 5. [Google Scholar] [CrossRef]
- Dudash, J., Jr.; Zhang, X.; Zeck, R.E.; Johnson, S.G.; Cox, G.G.; Conway, B.R.; Rybczynski, P.J.; Demarest, K.T. Glycosylated dihydrochalcones as potent and selective sodium glucose co-transporter 2 (SGLT2) inhibitors. Bioorg. Med. Chem. Lett. 2004, 14, 5121–5125. [Google Scholar] [CrossRef]
- Anunciato Casarini, T.P.; Frank, L.A.; Pohlmann, A.R.; Guterres, S.S. Dermatological applications of the flavonoid phloretin. Eur. J. Pharmacol. 2020, 889, 173593. [Google Scholar] [CrossRef]
- Choi, B.Y. Biochemical Basis of Anti-Cancer-Effects of Phloretin-A Natural Dihydrochalcone. Molecules 2019, 24, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.J.; Xu, S.K.; Lin, M.M.; Li, Y.K.; Sun, L.M.; Fang, J.B. Research advances on fruit coloring mechanism in red-fleshed kiwifruit. J. Fruit Sci. 2015, 32, 1232–1240. [Google Scholar] [CrossRef]
- Li, W.; Liu, Y.; Zeng, S.; Xiao, G.; Wang, G.; Wang, Y.; Peng, M.; Huang, H. Gene Expression Profiling of Development and Anthocyanin Accumulation in Kiwifruit (Actinidia chinensis) Based on Transcriptome Sequencing. PLoS ONE 2015, 10, e0136439. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, B.; Qi, Y.; Chen, X.; Liu, C.; Liu, Z.; Ren, X. Expression Differences of Pigment Structural Genes and Transcription Factors Explain Flesh Coloration in Three Contrasting Kiwifruit Cultivars. Front. Plant Sci. 2017, 8, 1507. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Desgagné-Penix, I. Transcriptome and metabolome profiling of Narcissus pseudonarcissus ‘King Alfred’ reveal components of Amaryllidaceae alkaloid metabolism. Sci. Rep. 2017, 7, 17356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.P.; An, L.J.; Hou, H.S. The Structure and Function of AP2/EREBP Transcription Factors. Chin. Agric. Sci. Bull. 2006, 22, 33–38. [Google Scholar] [CrossRef]
- Liu, W.W.; Li, W.X. Progress of plant bHLH transcription factor. Curr. Biotech. 2013, 1, 7–11. [Google Scholar]
- Burdon, J.N. Kiwifruit Biology: The Commercial Implications of Fruit Maturation. Hortic. Rev. 2019, 46, 46. [Google Scholar] [CrossRef]
- Bray, N.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal RNA-Seq quantification. arXiv 2015, arXiv:1505.02710. [Google Scholar]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Ampomah-Dwamena, C.; McGhie, T.; Wibisono, R.; Montefiori, M.; Hellens, R.P.; Allan, A.C. The kiwifruit lycopene beta-cyclase plays a significant role in carotenoid accumulation in fruit. J. Exp. Bot. 2009, 60, 3765–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Y.; Luo, L.; Chen, Y.; Westerberg, L.S.; Zhou, P.; Xu, Z.; Herrada, A.A.; Park, C.S.; Kubo, M.; Mei, H.; et al. SARS-CoV-2 infection causes immunodeficiency in recovered patients by downregulating CD19 expression in B cells via enhancing B-cell metabolism. Signal Transduct. Target. Ther. 2021, 6, 345. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | All Metabolites | MS1 KEGG |
|---|---|---|
| POS | 201 | 73 |
| NEG | 209 | 62 |
| Total | 410 | 135 |
| Metabolite Name | Comparison Group | Log2 FC | KEGG Pathway |
|---|---|---|---|
| Kaempferol-3-rhamnoside | DH_vs_R | 2.39 | Flavone and flavonol biosynthesis |
| HT_vs_R | 17.01 | ||
| JY_vs_R | 2.47 | ||
| RZ_vs_R | 6.18 | ||
| L-Epicatechin | DH_vs_R | 3.84 | Flavonoid biosynthesis |
| HT_vs_R | 19.68 | ||
| JY_vs_R | 4.55 | ||
| RZ_vs_R | 5.94 | ||
| Phloretin | DH_vs_R | 13.10 | Flavonoid biosynthesis |
| HT_vs_R | 13.10 | ||
| JY_vs_R | 13.10 | ||
| RZ_vs_R | 13.10 | ||
| Trifolin | DH_vs_R | 5.53 | Flavone and flavonol biosynthesis |
| HT_vs_R | 17.50 | ||
| JY_vs_R | 5.83 | ||
| RZ_vs_R | 3.25 |
| Groups | All DEGs | MS1 KEGG | ||
|---|---|---|---|---|
| Up | Down | Up | Down | |
| DH_vs_R | 12,191 | 9238 | 1817 | 2248 |
| HT_vs_R | 14,838 | 9624 | 2331 | 2349 |
| JY_vs_R | 11,895 | 9243 | 1788 | 2263 |
| RZ_vs_R | 15,814 | 8454 | 2573 | 2058 |
| Gene_ID | Gene_Description | TF_Family |
|---|---|---|
| Acc24966 | Chalcone synthase | Chal_sti_synt_N |
| Acc18331 | Flavonoid-3’-hydroxylase | p450 |
| Acc11227 | Caffeoyl-CoA-O-methyltransferase | – |
| Acc12443 | BAHD acyltransferase | – |
| Acc23632 | Dihydroflavonol 4-reductase | Epimerase |
| novel.5207 | Caffeoyl-CoA-O-methyltransferase | – |
| novel.82 | Trans-cinnamate-4-monooxygenase | p450 |
| Acc00871 | Cytochrome P450 CYP73A100 like | p450 |
| Acc23312 | Caffeoyl-CoA-O-methyltransferase | – |
| Acc32390 | Flavonoid-3’,5’-hydroxylase | p450 |
| Acc04926 | Ethylene-responsive transcription factor RAP2–4 like | AP2 |
| Acc25114 | Ethylene-responsive transcription factor 4 like | AP2 |
| Acc02924 | Transcription factor bHLH62 like | bHLH |
| Kaempferol-3-Rhamnoside | L-Epicatechin | Phloretin | Trifolin | |
|---|---|---|---|---|
| Acc24966 | 0.878 ** | 0.838 ** | NA | NA |
| Acc18331 | 0.812 ** | NA | NA | NA |
| Acc11227 | NA | NA | NA | NA |
| Acc12443 | NA | NA | NA | NA |
| Acc23632 | NA | NA | NA | NA |
| novel.5207 | NA | NA | NA | NA |
| novel.82 | NA | NA | 1** | NA |
| Acc00871 | 0.82 | NA | NA | NA |
| Acc23312 | 0.819 ** | NA | NA | NA |
| Acc32390 | NA | NA | NA | NA |
| Acc04926 | 0.848 ** | 0.909 ** | NA | 0.889 ** |
| Acc25114 | 0.934 ** | 0.923 ** | NA | NA |
| Acc02924 | −0.849 ** | NA | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Tang, Z.; Zheng, H.; Zhong, C.; Zhang, Q. Comprehensive Analysis of Metabolome and Transcriptome in Fruits and Roots of Kiwifruit. Int. J. Mol. Sci. 2023, 24, 1299. https://doi.org/10.3390/ijms24021299
Zhang L, Tang Z, Zheng H, Zhong C, Zhang Q. Comprehensive Analysis of Metabolome and Transcriptome in Fruits and Roots of Kiwifruit. International Journal of Molecular Sciences. 2023; 24(2):1299. https://doi.org/10.3390/ijms24021299
Chicago/Turabian StyleZhang, Long, Zhengmin Tang, Hao Zheng, Caihong Zhong, and Qiong Zhang. 2023. "Comprehensive Analysis of Metabolome and Transcriptome in Fruits and Roots of Kiwifruit" International Journal of Molecular Sciences 24, no. 2: 1299. https://doi.org/10.3390/ijms24021299
APA StyleZhang, L., Tang, Z., Zheng, H., Zhong, C., & Zhang, Q. (2023). Comprehensive Analysis of Metabolome and Transcriptome in Fruits and Roots of Kiwifruit. International Journal of Molecular Sciences, 24(2), 1299. https://doi.org/10.3390/ijms24021299