



Peptide Inhibitors of Insulin Fibrillation: Current and Future Challenges



Abstract
:1. Introduction
1.1. Pathological Amyloidoses
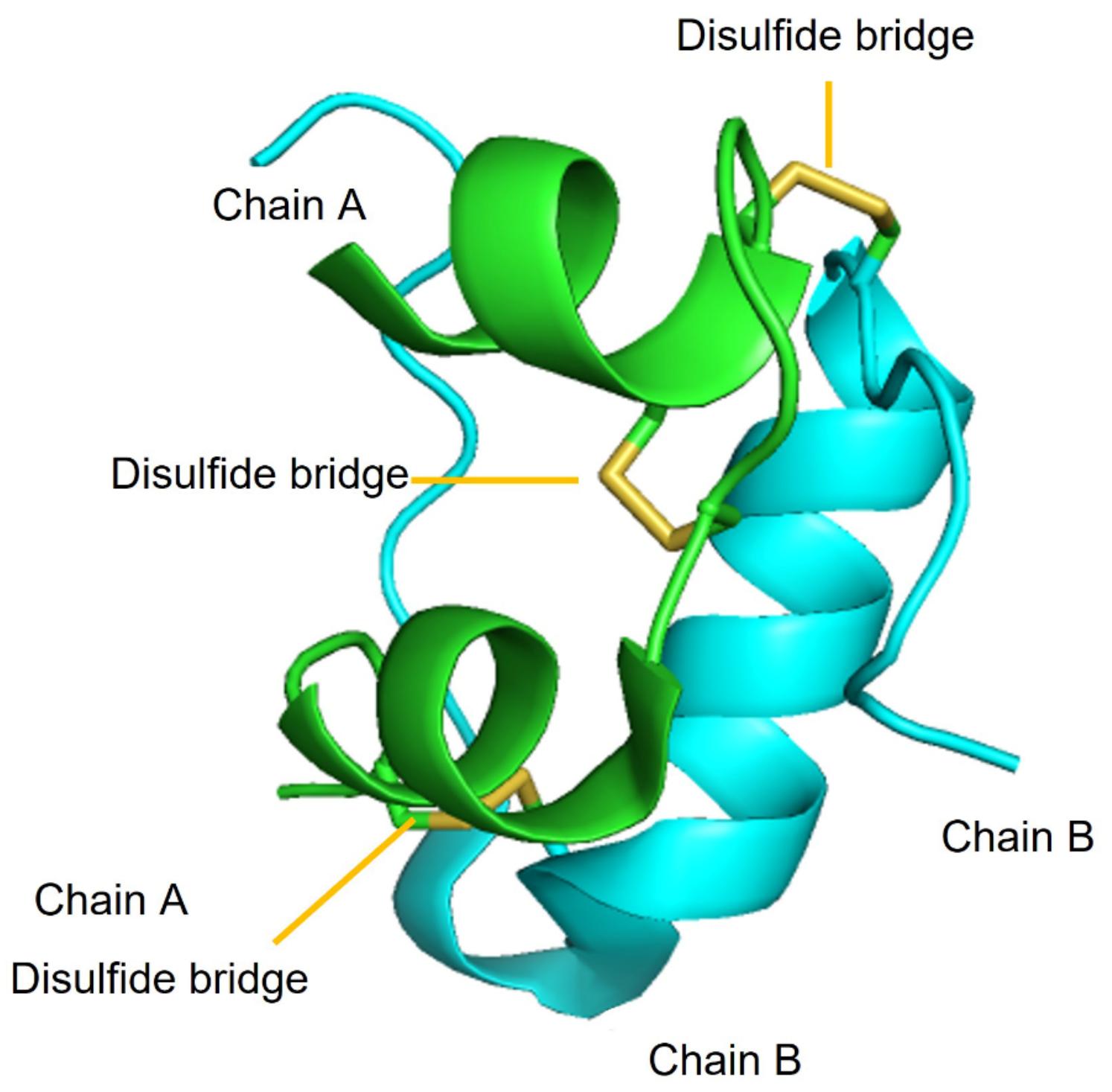
1.2. Insulin Structure
1.3. Insulin Biogenesis and Storage in Granules
1.4. Insulin Function
1.5. Insulin Therapy for Diabetes
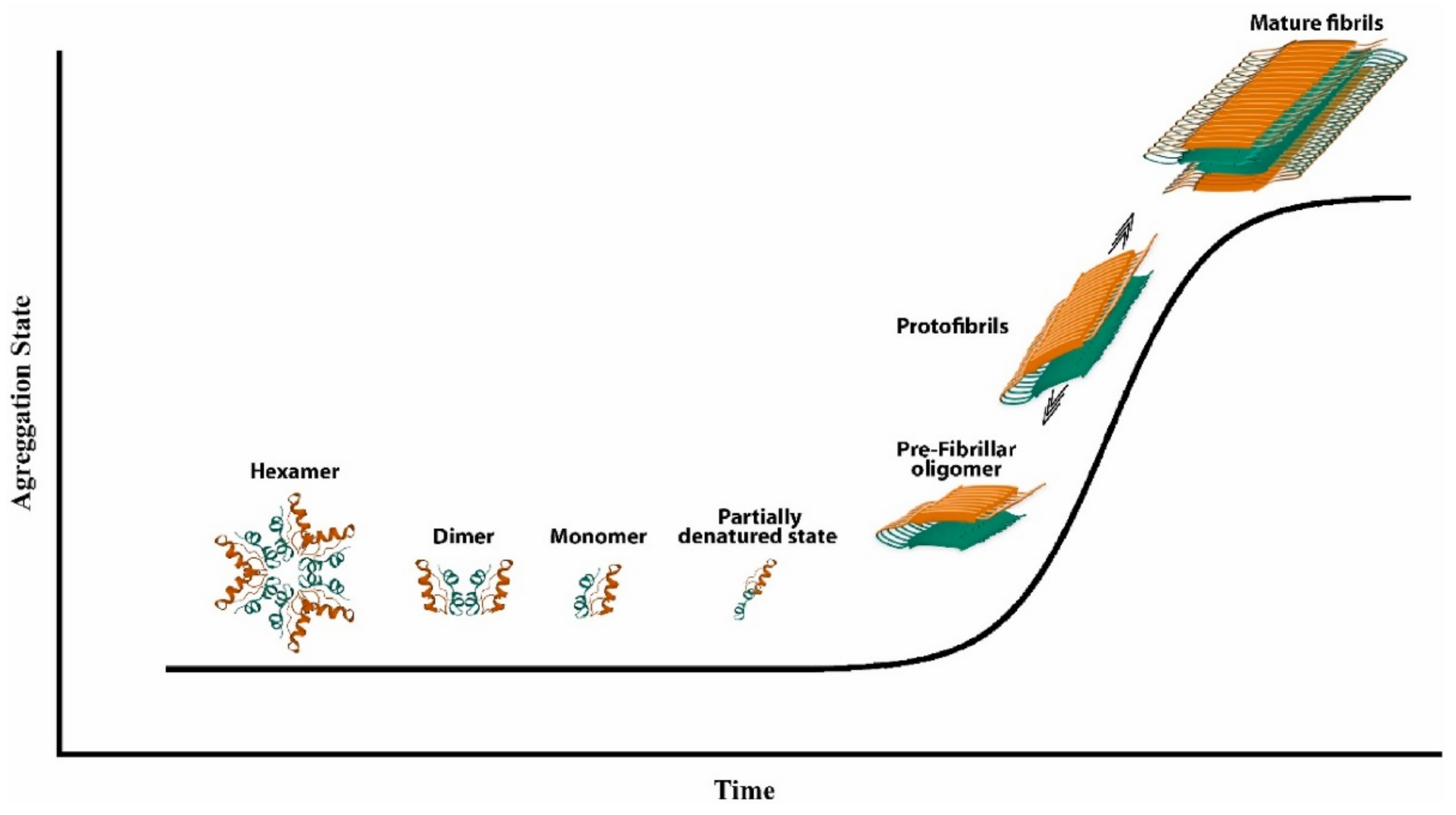
2. Insulin Fibrillation
2.1. Effect of Agitation on Insulin Fibrillation
2.2. Effect of Low pH on Insulin Fibrillation
2.3. Effect of Insulin Concentration on Fibrillation
2.4. Effect of High Ionic Strength on Insulin Fibrillation
3. Peptide Inhibitors of Insulin Fibrillation
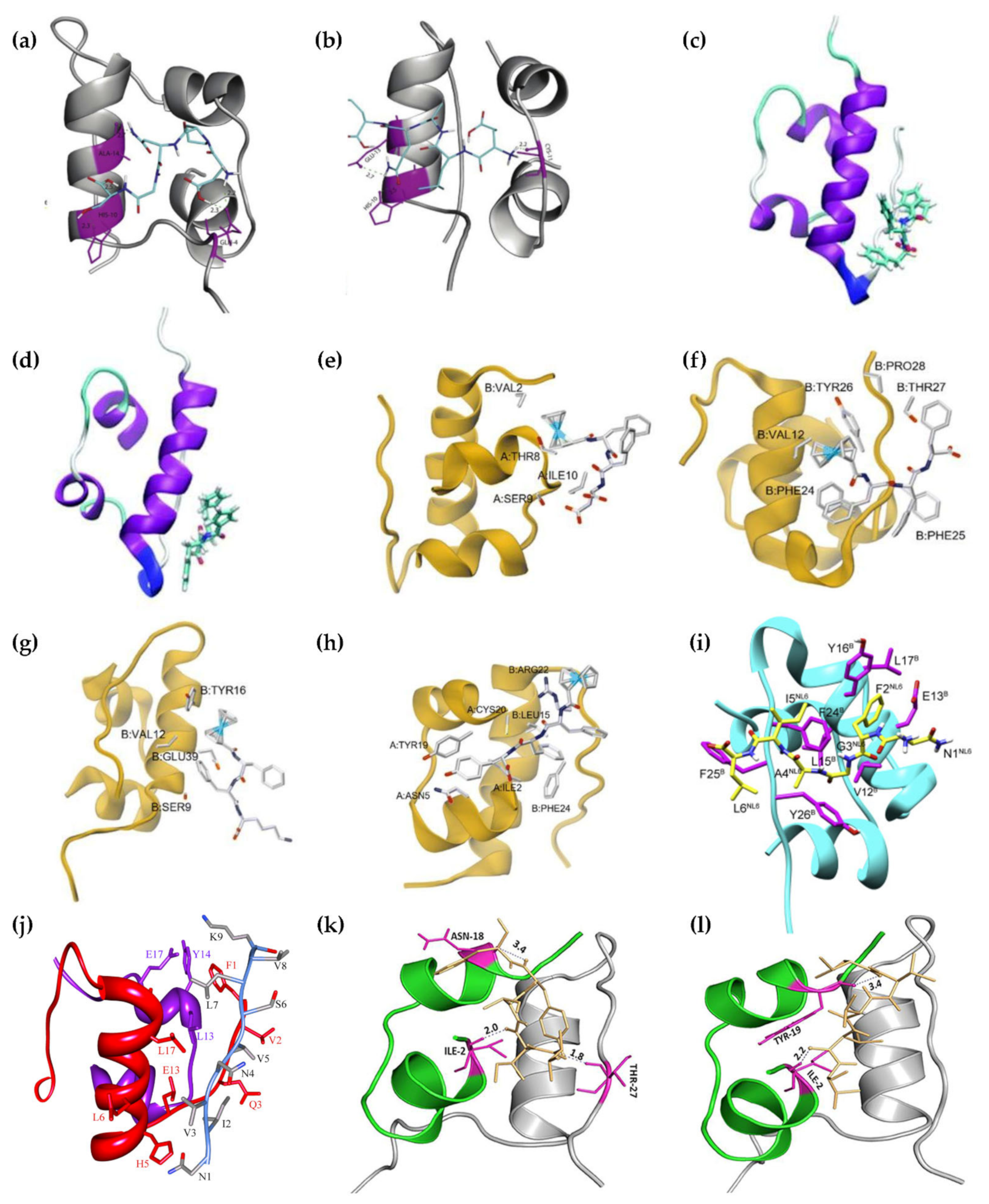
3.1. AF- and FA-Perylenebisimides
3.2. DPNGS and ELAQM
3.3. Ferrocenyl Peptides
3.4. FVPRK
3.5. KPWWPRR
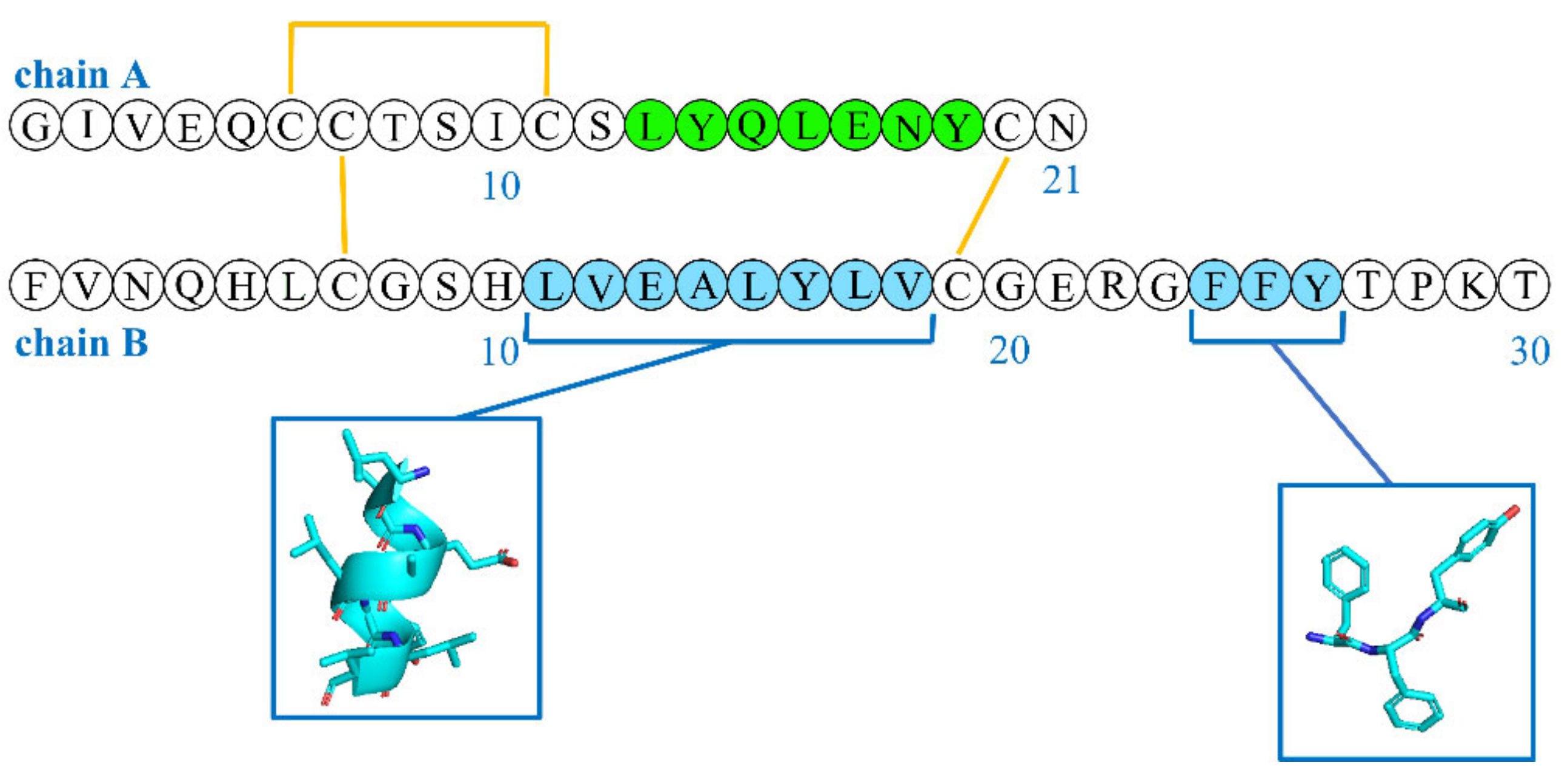
3.6. LVEALYL
3.7. NFGAIL and NFGAXL
3.8. NIVNVSLVK
3.9. VIFYW and VVVVV
3.10. VYYR
3.11. Wn-Taurine (n = 1–4) Conjugates
3.12. Techniques to Study Inhibition of Fibrillation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alraawi, Z.; Banerjee, N.; Mohanty, S.; Kumar, T.K.S. Amyloidogenesis: What do we know so far? Int. J. Mol. Sci. 2022, 23, 13970. [Google Scholar] [CrossRef] [PubMed]
- Muchtar, E.; Dispenzieri, A.; Magen, H.; Grogan, M.; Mauermann, M.; McPhail, E.D.; Kurtin, P.J.; Leung, N.; Buadi, F.K.; Dingli, D.; et al. Systemic amyloidosis from A (AA) to T (ATTR): A review. J. Intern. Med. 2021, 289, 268–292. [Google Scholar] [CrossRef] [PubMed]
- Konstantoulea, K.; Louros, N.; Rousseau, F.; Schymkowitz, J. Heterotypic interactions in amyloid function and disease. FEBS J. 2022, 289, 2025–2046. [Google Scholar] [CrossRef] [PubMed]
- Subedi, S.; Sasidharan, S.; Nag, N.; Saudagar, P.; Tripathi, T. Amyloid cross-seeding: Mechanism, implication, and inhibition. Molecules 2022, 27, 1776. [Google Scholar] [CrossRef]
- Hong, D.-P.; Ahmad, A.; Fink, A.L. Fibrillation of human insulin A and B chains. Biochemistry 2006, 45, 9342–9353. [Google Scholar] [CrossRef]
- Bhasikuttan, A.C.; Mohanty, J. Detection, inhibition and disintegration of amyloid fibrils: The role of optical probes and macrocyclic receptors. Chem. Commun. 2017, 53, 2789–2809. [Google Scholar] [CrossRef]
- Sinnige, T. Molecular mechanisms of amyloid formation in living systems. Chem. Sci. 2022, 13, 7080–7097. [Google Scholar] [CrossRef]
- Lewkowicz, E.; Gursky, O. Dynamic protùein structures in normal function and pathologic misfolding in systemic amyloidosis. Biophys. Chem. 2022, 280, 106699. [Google Scholar] [CrossRef] [PubMed]
- Stepanenko, O.V.; Sulatsky, M.I.; Mikhailova, E.V.; Stepanenko, O.V.; Kuznetsova, I.M.; Turoverov, K.K.; Sulatskaya, A.I. Trypsin induced degradation of amyloid fibrils. Int. J. Mol. Sci. 2021, 22, 4828. [Google Scholar]
- Schönfelder, J.; Pfeiffer, P.B.; Pradhan, T.; Bijzet, J.; Hazenberg, B.P.C.; Schönland, S.O.; Hegenbart, U.; Reif, B.; Haupt, C.; Fändrich, M. Protease resistance of ex vivo amyloid fibrils implies the proteolytic selection of disease-associated fibril morphologies. Amyloid 2021, 28, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, K.; Sturchio, A.; Espay, A.J. Proteins do not replicate, they precipitate: Phase transition and loss of function toxicity in amyloid pathologies. Biology 2022, 11, 535. [Google Scholar] [CrossRef]
- Buell, A.K. Stability matters, too—The thermodynamics of amyloid fibril formation. Chem. Sci. 2022, 13, 10177–10192. [Google Scholar] [CrossRef]
- Rubel, M.S.; Fedotov, S.A.; Grizel, A.V.; Sopova, J.V.; Malikova, O.A.; Chernoff, Y.O.; Rubel, A.A. Functional mammalian amyloids and amyloid-like proteins. Life 2020, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chattopadhyay, T.; Pal Singh, M.; Surolia, A. Supramolecular insulin assembly ii for a sustained treatment of type 1 diabetes mellitus. Appl. Biol. Sci. 2010, 107, 13246–13251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.E.; Mulka, A.; Mao, L.; Sharafieh, R.; Qiao, Y.; Kesserwan, S.; Wu, R.; Kreutzer, D.; Klueh, U. Insulin derived fibrils induce cytotoxicity in vitro and trigger inflammation in murine models. J. Diabetes Sci. Technol. 2023, 17, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Ma, J.W.; Lei, P.; Wu, W.H.; Yu, Y.P.; Xiang, Y.; Tong, A.J.; Zhao, Y.F.; Li, Y.M. Insulin is a kinetic but not a thermodynamic inhibitor of amylin aggregation. FEBS J. 2009, 276, 3365–3371. [Google Scholar] [CrossRef]
- Lupaescu, A.V.; Iavorschi, M.; Covasa, M. The use of bioactive compounds in hyperglycemia- and amyloid fibrils-induced toxicity in type 2 diabetes and alzheimer’s disease. Pharmaceutics 2022, 14, 235. [Google Scholar] [CrossRef]
- Hassan, M.N.; Nabi, F.; Khan, A.N.; Hussain, M.; Siddiqui, W.A.; Uversky, V.N.; Khan, R.H. The amyloid state of proteins: A boon or bane? Int. J. Biol. Macromol. 2022, 200, 593–617. [Google Scholar] [CrossRef]
- Ryle, A.P.; Sanger, F.; Smith, L.F.; Kitai, R. The disulphide bonds of insulin. Biochem. J. 1955, 60, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Mayer, J.P.; Zhang, F.; DiMarchi, R.D. Insulin structure and function. Biopolymers 2007, 88, 687–713. [Google Scholar] [CrossRef]
- Qafary, M.; Rashno, F.; Khajeh, K.; Khaledi, M.; Moosavi-Movahedi, A.A. Insulin fibrillation: Strategies for inhibition. Progress Biophys. Mol. Biol. 2022, 175, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Omar-Hmeadi, M.; Idevall-Hagren, O. Insulin granule biogenesis and exocytosis. Cell. Mol. Life Sci. 2021, 78, 1957–1970. [Google Scholar] [CrossRef]
- Guthrie, R.A.; Guthrie, D.W. Pathophysiology of diabetes mellitus. Crit. Care Nurs. Q. 2004, 27, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Fazakerley, D.J.; Koumanov, F.; Holman, G.D. Glut4 on the move. Biochem. J. 2022, 479, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Moraes-Vieira, P.M.; Saghatelian, A.; Kahn, B.B. Glut4 expression in adipocytes regulates de novo lipogenesis and levels of a novel class of lipids with antidiabetic and anti-inflammatory effects. Diabetes 2016, 65, 1808–1815. [Google Scholar] [CrossRef] [Green Version]
- LeRoith, D.; Holly, J.M.P.; Forbes, B.E. Insulin-like growth factors: Ligands, binding proteins, and receptors. Mol. Metab. 2021, 52, 101245. [Google Scholar] [CrossRef] [PubMed]
- Miller, Y. Advancements and future directions in research of the roles of insulin in amyloid diseases. Biophys. Chem. 2022, 281, 106720. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.diabetesatlas.org (accessed on 12 September 2022).
- American Diabetes Association. 2. Classification and diagnosis of diabetes. Diabetes Care 2014, 38, S8–S16. [Google Scholar]
- Wu, Y.P.; Graves, M.M.; Roberts, M.C.; Mitchell, A.C. Is insulin pump therapy better than injection for adolescents with diabetes? Diabetes Res. Clin. Pract. 2010, 89, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.M.; Osmani, L.; Matsangos, A.E.; Li, Q.K. Current insight in the localized insulin-derived amyloidosis (lida): Clinico-pathological characteristics and differential diagnosis. Pathol. Res. Pract. 2017, 213, 1237–1241. [Google Scholar] [CrossRef]
- Bußmann, A.B.; Grünerbel, L.M.; Durasiewicz, C.P.; Thalhofer, T.A.; Wille, A.; Richter, M. Microdosing for drug delivery application—A review. Sens. Actuator A Phys. 2021, 330, 112820. [Google Scholar] [CrossRef]
- Giugliano, D.; Scappaticcio, L.; Longo, M.; Caruso, P.; Maiorino, M.I.; Bellastella, G.; Esposito, K. Simplification of complex insulin therapy: A story of dogma and therapeutic resignation. Diabetes Res. Clin. Pract. 2021, 178, 108958. [Google Scholar] [CrossRef]
- Arora, S.; Agrawal, N.K.; Shanthaiah, D.M.; Verma, A.; Singh, S.; Patne, S.C.U.; Kalra, S.; Singh, P.; Goyal, S. Early detection of cutaneous complications of insulin therapy in type 1 and type 2 diabetes mellitus. Prim. Care Diab. 2021, 15, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Binder, C.; Lauritzen, T.; Faber, O.; Pramming, S. Insulin pharmacokinetics. Diabetes Care 1984, 7, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Azarfar, K.; Yaghmaei, P.; Amoli, M.M.; Hayati-Roodbari, N.; Ebrahim-Habibi, A. Local insulin-derived amyloidosis modelconfronted with silymarin: Histological insights and gene expression of MMP, TNF-alpha; and IL-6. Int. J. Mol. Sci. 2022, 23, 4952. [Google Scholar] [CrossRef] [PubMed]
- Kranc, C.; Wagner, R.; Joy, N.M.; Feldman, J.; Reid, D.C. Cutaneous insulin-derived amyloidosis presenting as hyperkeratotic nodules. Cutis 2021, 107, E6–E9. [Google Scholar] [CrossRef] [PubMed]
- Kunitomi, R.; Pradipta, A.R.; Kawabe, H.; Lobsiger, N.; Tanaka, K.; Zako, T. Inhibition of amyloid formation of amyloid β (1-42), amylin and insulin by 1,5-diazacyclooctanes, a spermine-acrolein conjugate. Bioorg. Med. Chem. 2021, 46, 116391. [Google Scholar] [CrossRef]
- Ratha, B.N.; Ghosh, A.; Brender, J.R.; Gayen, N.; Ilyas, H.; Neeraja, C.; Das, K.P.; Mandal, A.K.; Bhunia, A. Inhibition of insulin amyloid fibrillation by a novel amphipathic heptapeptide. J. Biol. Chem. 2016, 291, 23545–23556. [Google Scholar] [CrossRef] [Green Version]
- Engudar, G.; Rodríguez-Rodríguez, C.; Mishra, N.K.; Bergamo, M.; Amouroux, G.; Jensen, K.J.; Saatchi, K.; Häfeli, U.O. Metal-ion coordinated self-assembly of human insulin directs kinetics of insulin release as determined by preclinical spect/ct imaging. J. Control. Release 2022, 343, 347–360. [Google Scholar] [CrossRef]
- Iannuzzi, C.; Borriello, M.; Portaccio, M.; Irace, G.; Sirangelo, I. Insights into insulin fibril assembly at physiological and acidic pH and related amyloid intrinsic fluorescence. Int. J. Mol. Sci. 2017, 18, 2551. [Google Scholar] [CrossRef] [Green Version]
- Fagihi, M.H.A.; Bhattacharjee, S. Amyloid fibrillation of insulin: Amelioration strategies and implications for translation. ACS Pharmacol. Transl. Sci. 2022, 5, 1050–1061. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Movafaghi, S.; Francino Urdániz, I.M.; Rowe, T.M.; Goodwin, A.; Randolph, T.W. Insulin fibril formation caused by mechanical shock and cavitation. J. Phys. Chem. B 2021, 125, 8021–8027. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.; Khurana, R.; Coats, A.; Frokjaer, S.; Brange, J.; Vyas, S.; Uversky, V.N.; Fink, A.L. Effect of environmental factors on the kinetics of insulin fibril formation: Elucidation of the molecular mechanism. Biochemistry 2001, 40, 6036–6046. [Google Scholar] [CrossRef] [PubMed]
- Kurouski, D.; Washington, J.; Ozbil, M.; Prabhakar, R.; Shekhtman, A.; Lednev, I.K. Disulfide bridges remain intact while native insulin converts into amyloid fibrils. PloS ONE 2012, 7, e36989. [Google Scholar] [CrossRef]
- Ivanova, M.I.; Sievers, S.A.; Sawaya, M.R.; Wall, J.S.; Eisenberg, D. Molecular basis for insulin fibril assembly. Proc. Natl. Acad. Sci. USA 2009, 106, 18990–18995. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Shah, M.; Saraogi, I. Molecular aspects of insulin aggregation and various therapeutic interventions. ACS Bio Med. Chem. Au 2022, 2, 205–221. [Google Scholar] [CrossRef]
- Mishra, N.K.; Krishna Deepak, R.N.V.; Sankararamakrishnan, R.; Verma, S. Controlling in vitro insulin amyloidosis with stable peptide conjugates: A combined experimental and computational study. J. Phys. Chem. B 2015, 119, 15395–15406. [Google Scholar] [CrossRef] [PubMed]
- Vashisth, H.; Abrams, C.F. All-atom structural models of insulin binding to the insulin receptor in the presence of a tandem hormone-binding element. Proteins 2013, 81, 1017–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blyth, A.; Ortiz, M.; Merriman, A.; Delaine, C.; Forbes, B. Determinants of igf-ii influencing stability, receptor binding and activation. Sci. Rep. 2022, 12, 4695. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, Z.; Feng, D.; Shang, Y.; Li, X.; Liu, J.; Li, C.; Yang, Z. An insulin-inspired supramolecular hydrogel for prevention of type 1 diabetes. Adv. Sci. 2021, 8, 2003599. [Google Scholar] [CrossRef]
- Chouchane, K.; Frachon, T.; Marichal, L.; Nault, L.; Vendrely, C.; Maze, A.; Bruckert, F.; Weidenhaupt, M. Insulin aggregation starts at dynamic triple interfaces, originating from solution agitation. Coll. Surf. B 2022, 214, 112451. [Google Scholar] [CrossRef] [PubMed]
- Adam, J.A.; Middlestead, H.R.; Debono, N.E.; Hirsa, A.H. Effects of shear rate and protein concentration on amyloidogenesis via interfacial shear. J. Phys. Chem. B 2021, 125, 10355–10363. [Google Scholar] [CrossRef] [PubMed]
- Whittingham, J.L.; Scott, D.J.; Chance, K.; Wilson, A.; Finch, J.; Brange, J.; Guy Dodson, G. Insulin at pH 2: Structural analysis of the conditions promoting insulin fibre formation. J. Mol. Biol. 2002, 318, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Tománková, H.; Pinkasová, M. Determination of parabens and their degradation product p-hydroxy-benzoic acid in pharmaceutical dosage forms by hptlc densitometry. Anal. Lett. 1990, 23, 1319–1332. [Google Scholar] [CrossRef]
- Melberg, S.G.; Havelund, S.; Villumsen, J.; Brange, J. Insulin compatibility with polymer materials used in external pump infusion systems. Diabet. Med. 1988, 5, 243–247. [Google Scholar] [CrossRef]
- Goto, Y.; Noji, M.; Nakajima, K.; Yamaguchi, K. Supersaturation-dependent formation of amyloid fibrils. Molecules 2022, 27, 4588. [Google Scholar] [CrossRef]
- Owczarz, M.; Arosio, P. Sulfate anion delays the self-assembly of human insulin by modifying the aggregation pathway. Biophys. J. 2014, 107, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Kasai, T.; Wada, T.; Iijima, T.; Minami, Y.; Sakaguchi, T.; Koga, R.; Shiratori, T.; Otsuka, Y.; Shimada, Y.; Okayama, Y.; et al. Comparative study of the hydrophobic interaction effect of ph and ionic strength on aggregation/emulsification of congo red and amyloid fibrillation of insulin. Biochim. Biophys. Acta Adv. 2022, 2, 100036. [Google Scholar] [CrossRef]
- Ziaunys, M.; Sakalauskas, A.; Sneideris, T.; Smirnovas, V. Lysozyme fibrils alter the mechanism of insulin amyloid aggregation. Int. J. Mol. Sci. 2021, 22, 1775. [Google Scholar] [CrossRef]
- Marchesan, S.; Prato, M. Nanomaterials for (nano)medicine. ACS Med. Chem. Lett. 2013, 4, 147–149. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Huang, R.; Qi, W.; Su, R.; He, Z. Fluorescent silicon nanoparticles inhibit the amyloid fibrillation of insulin. J. Mater. Chem. B 2019, 7, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Shlapa, Y.; Solopan, S.; Sarnatskaya, V.; Siposova, K.; Garcarova, I.; Veltruska, K.; Timashkov, I.; Lykhova, O.; Kolesnik, D.; Musatov, A.; et al. Cerium dioxide nanoparticles synthesized via precipitation at constant ph: Synthesis, physical-chemical and antioxidant properties. Coll. Surf. B 2022, 220, 112960. [Google Scholar] [CrossRef] [PubMed]
- Antosova, A.; Gancar, M.; Bednarikova, Z.; Marek, J.; Zahn, D.; Dutz, S.; Gazova, Z. Surface-modified magnetite nanoparticles affect lysozyme amyloid fibrillization. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129941. [Google Scholar] [CrossRef]
- Meesaragandla, B.; Karanth, S.; Janke, U.; Delcea, M. Biopolymer-coated gold nanoparticles inhibit human insulin amyloid fibrillation. Sci. Rep. 2020, 10, 7862. [Google Scholar] [CrossRef]
- Vus, K.; Tarabara, U.; Danylenko, I.; Pirko, Y.; Krupodorova, T.; Yemets, A.; Blume, Y.; Turchenko, V.; Klymchuk, D.; Smertenko, P.; et al. Silver nanoparticles as inhibitors of insulin amyloid formation: A fluorescence study. J. Mol. Liq. 2021, 342, 117508. [Google Scholar] [CrossRef]
- Sun, X.; Yang, C.; Liu, W.; Lu, K.; Yin, H. Charge modifications of graphene oxide enhance the inhibitory effect on insulin amyloid fibrillation based on electrostatic interactions. Int. J. Biol. Macromol. 2022, 225, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.-M.; Zhang, W.; Zhang, J.-D.; Li, X.-F.; Yu, F.-D.; Li, C.-Y.; Liu, G.-J.; Xing, G.-W. Dual functional amphiphilic sugar-coated aie-active fluorescent organic nanoparticles for the monitoring and inhibition of insulin amyloid fibrillation based oncarbohydrate–protein interactions. J. Mater. Chem. B 2022, 10, 5602–5611. [Google Scholar] [CrossRef]
- Camlik, G.; Ozakca, I.; Bilakaya, B.; Ozcelikay, A.T.; Velaro, A.J.; Wasnik, S.; Degim, I.T. Development of composite carbon quantum dots-insulin formulation for oral administration. J. Drug Deliv. Sci. Technol. 2022, 76, 103833. [Google Scholar] [CrossRef]
- Yang, Q.Q.; He, H.; Li, C.Q.; Luo, L.B.; Li, S.L.; Xu, Z.Q.; Jin, J.C.; Jiang, F.L.; Liu, Y.; Yang, M. Molecular mechanisms of the ultra-strong inhibition effect of oxidized carbon dots on human insulin fibrillation. ACS Appl. Bio Mater. 2020, 3, 217–226. [Google Scholar] [CrossRef]
- Wang, B.B.; Wang, Y.Y.; Zhang, X.Y.; Xu, Z.Q.; Jiang, P.; Jiang, F.L.; Liu, Y. Bifunctional carbon dots for cell imaging and inhibition of human insulin fibrillation in the whole aggregation process. Int. J. Biol. Macromol. 2020, 147, 453–462. [Google Scholar] [CrossRef]
- Li, S.-L.; Yang, Q.-Q.; Liu, X.-Y.; Jiang, F.-L.; Xiong, J.; Jiang, P.; Liu, Y. Zn-doped Cu2S quantum dots as new high-efficiency inhibitors against human insulin fibrillation based on specific electrostatic interaction with oligomers. Int. J. Biol. Macromol. 2021, 179, 161–169. [Google Scholar] [CrossRef]
- Milewska, M.; Milewski, A.; Wandzik, I.; Stenzel, M.H. Structurally analogous trehalose and sucrose glycopolymers—Comparative characterization and evaluation of their effects on insulin fibrillation. Polym. Chem. 2022, 13, 1831–1843. [Google Scholar] [CrossRef]
- Gelb, M.B.; Messina, K.M.M.; Vinciguerra, D.; Ko, J.H.; Collins, J.; Tamboline, M.; Xu, S.; Ibarrondo, F.J.; Maynard, H.D. Poly(trehalose methacrylate) as an excipient for insulin stabilization: Mechanism and safety. ACS Appl. Mater. Interfaces 2022, 14, 37410–37423. [Google Scholar] [CrossRef]
- Das, A.; Gangarde, Y.M.; Pariary, R.; Bhunia, A.; Saraogi, I. An amphiphilic small molecule drives insulin aggregation inhibition and amyloid disintegration. Int. J. Biol. Macromol. 2022, 218, 981–991. [Google Scholar] [CrossRef]
- Levy-Sakin, M.; Shreberk, M.; Daniel, Y.; Gazit, E. Targeting insulin amyloid assembly by small aromatic molecules: Toward rational design of aggregation inhibitors. Islets 2009, 1, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yin, H.; Zhang, L.; Liu, R.; Qi, W.; He, Z.; Su, R. Oligomeric procyanidins inhibit insulin fibrillation by forming unstructured and off-pathway aggregates. RSC Adv. 2021, 11, 37290–37298. [Google Scholar] [CrossRef] [PubMed]
- Channuwong, P.; Salae, K.; Chongruchiroj, S.; Cheng, H.; Suantawee, T.; Thilavech, T.; Adisakwattana, S. Dietary anthocyanins inhibit insulin fibril formation and cytotoxicity in 3T3-L1 preadipocytes. Int. J. Biol. Macromol. 2022, 223, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Alam, P.; Chaturvedi, S.K.; Siddiqi, M.K.; Rajpoot, R.K.; Ajmal, M.R.; Zaman, M.; Khan, R.H. Vitamin k3 inhibits protein aggregation: Implication in the treatment of amyloid diseases. Sci. Rep. 2016, 6, 26759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, P.N.; Parmar, K.; Patel, S.; Das, M. Orange g is a potential inhibitor of human insulin amyloid fibrillation and can be used as a probe to study mechanism of amyloid fibrillation and its inhibition. Int. J. Biol. Macromol. 2022, 220, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Nusrat, S.; Zaidi, N.; Siddiqi, M.K.; Zaman, M.; Siddique, I.A.; Ajmal, M.R.; Abdelhameed, A.S.; Khan, R.H. Anti-parkinsonian l-dopa can also act as anti-systemic amyloidosis—A mechanistic exploration. Int. J. Biol. Macromol. 2017, 99, 630–640. [Google Scholar] [CrossRef]
- Pandya, A.K.; Petravale, V.B. Computational avenues in oral protein and peptide therapeutics. Drug Discov. Today 2021, 26, 1510–1520. [Google Scholar] [CrossRef]
- Fernandez, L.; Bustos, R.H.; Zapata, C.; Garcia, J.; Jauregui, E.; Ashraf, G.M. Immunogenicity in Protein and Peptide Based-Therapeutics: An Overview. Curr. Protein Pept. Sci. 2018, 19, 958–971. [Google Scholar] [CrossRef]
- Yamamoto, N.; Chatani, E. Multistep growth of amyloid intermediates and its inhibition toward exploring therapeutic way: A case study using insulin b chain and fibrinogen. Biophys. Physicobiol. 2022, 19, e190017. [Google Scholar] [CrossRef] [PubMed]
- Sedov, I.; Khaibrakhmanova, D. Molecular mechanisms of inhibition of protein amyloid fibril formation: Evidence and perspectives based on kinetic models. Int. J. Mol. Sci. 2022, 23, 13428. [Google Scholar] [PubMed]
- Roy Chowdhury, S.; Mondal, S.; Iyer, P.K. Blocking oligomeric insulin amyloid fibrillation via perylenebisimides containing dipeptide tentacles. ACS Biomater. Sci. Eng. 2018, 4, 4076–4083. [Google Scholar] [CrossRef] [PubMed]
- Siddiqi, M.K.; Majid, N.; Alam, P.; Malik, S.; Alam, A.; Rajan, S.; Ajmal, M.R.; Khan, R.H. Both beta sheet breaker and alpha helix forming pentapeptide inhibits protein fibrillation: Implication for the treatment of amyloid disorders. Int. J. Biol. Macromol. 2020, 143, 102–111. [Google Scholar] [CrossRef]
- Yao, P.; Zhang, J.; You, S.; Qi, W.; Su, R.; He, Z. Ferrocene-modified peptides as inhibitors against insulin amyloid aggregation based on molecular simulation. J. Mater. Chem. B 2020, 8, 3076–3086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yao, P.; You, S.; Qi, W.; Su, R.; He, Z. Study of the kinetics and mechanism of ferrocene-tripeptide inhibiting insulin aggregation. J. Mater. Chem. B 2022, 10, 7780–7788. [Google Scholar] [CrossRef]
- Zhou, A.; Xie, J.; Han, H.; Chen, Y.; Zhao, C.; Li, J. Supramolecular nanoparticles of insulin and pentapeptide for inhibition of fibrillation and controlled release. J. Biomed. Nanotechnol. 2018, 14, 959–967. [Google Scholar] [CrossRef]
- Ratha, B.N.; Kar, R.K.; Kalita, S.; Kalita, S.; Raha, S.; Singha, A.; Garai, K.; Mandal, B.; Bhunia, A. Sequence specificity of amylin-insulin interaction: A fragment-based insulin fibrillation inhibition study. Biochim. Biophys. Acta Prot. Proteom. 2019, 1867, 405–415. [Google Scholar] [CrossRef]
- Banerjee, V.; Kar, R.K.; Datta, A.; Parthasarathi, K.; Chatterjee, S.; Das, K.P.; Bhunia, A. Use of a small peptide fragment as an inhibitor of insulin fibrillation process: A study by high and low resolution spectroscopy. PLoS ONE 2013, 8, e72318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqi, M.K.; Alam, P.; Iqbal, T.; Majid, N.; Malik, S.; Nusrat, S.; Alam, A.; Ajmal, M.R.; Uversky, V.N.; Khan, R.H. Elucidating the inhibitory potential of designed peptides against amyloid fibrillation and amyloid associated cytotoxicity. Front. Chem. 2018, 6, 311. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Das, D.; Sarkar, J.; Banerjee, N.; Jana, J.; Bhat, J.; Reddy, G.J.; Bharatam, J.; Chattopadhyay, S.; Chatterjee, S.; et al. Prion-derived tetrapeptide stabilizes thermolabile insulin via conformational trapping. iScience 2021, 24, 102573. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.K.; Joshi, K.B.; Verma, S. Inhibition of human and bovine insulin fibril formation by designed peptide conjugates. Mol. Pharm. 2013, 10, 3903–3912. [Google Scholar] [CrossRef]
- Marchesan, S.; Vargiu, A.V.; Styan, K.E. The phe-phe motif for peptide self-assembly in nanomedicine. Molecules 2015, 20, 19775–19788. [Google Scholar] [CrossRef] [Green Version]
- Reches, M.; Gazit, E. Casting metal nanowires within discrete self-assembled peptide nanotubes. Science 2003, 300, 625–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, C.; Sigurdsson, E.M.; Morelli, L.; Asok Kumar, R.; Castaño, E.M.; Frangione, B. Β-sheet breaker peptides inhibit fibrillogenesis in a rat brain model of amyloidosis: Implications for alzheimer’s therapy. Nat. Med. 1998, 4, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.M.; Melchionna, M.; Bellotto, O.; Kralj, S.; Semeraro, S.; Parisi, E.; Iglesias, D.; D’Andrea, P.; De Zorzi, R.; Vargiu, A.V.; et al. Nanoscale assembly of functional peptides with divergent programming elements. ACS Nano 2021, 15, 3015–3025. [Google Scholar] [CrossRef]
- Sawaya, M.R.; Sambashivan, S.; Nelson, R.; Ivanova, M.I.; Sievers, S.A.; Apostol, M.I.; Thompson, M.J.; Balbirnie, M.; Wiltzius, J.J.W.; McFarlane, H.T.; et al. Atomic structures of amyloid cross-β spines reveal varied steric zippers. Nature 2007, 447, 453–457. [Google Scholar] [CrossRef]
- Marchesan, S.; Waddington, L.; Easton, C.D.; Winkler, D.A.; Goodall, L.; Forsythe, J.; Hartley, P.G. Unzipping the role of chirality in nanoscale self-assembly of tripeptide hydrogels. Nanoscale 2012, 4, 6752–6760. [Google Scholar] [CrossRef]
- Makin, O.S.; Atkins, E.; Sikorski, P.; Johansson, J.; Serpell, L.C. Molecular basis for amyloid fibril formation and stability. Proc. Natl. Acad. Sci. USA 2005, 102, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Gobeaux, F.; Wien, F. Reversible Assembly of a Drug Peptide into Amyloid Fibrils: A Dynamic Circular Dichroism Study. Langmuir 2018, 34, 7180–7191. [Google Scholar] [CrossRef]
- Zhou, Y.; Hua, J.; Ding, D.; Tang, Y. Interrogating amyloid aggregation with aggregation-induced emission fluorescence probes. Biomaterials 2022, 286, 121605. [Google Scholar]
- Ageeva, A.A.; Doktorov, A.B.; Polyakov, N.E.; Leshina, T.V. Chiral linked systems as a model for understanding D-amino acids influence on the structure and properties of amyloid peptides. Int. J. Mol. Sci. 2022, 23, 3060. [Google Scholar] [CrossRef] [PubMed]
- Ragonis-Bachar, P.; Landau, M. Functional and pathological amyloid structures in the eyes of 2020 cryo-em. Curr. Opin. Struct. Biol. 2021, 68, 184–193. [Google Scholar] [PubMed]
- Visser, M.J.E.; Pretorius, E. Atomic force microscopy: The characterisation of amyloid protein structure in pathology. Curr. Top. Med. Chem. 2019, 19, 2958–2973. [Google Scholar] [PubMed]
- Sasahara, K.; Goto, Y. Application and use of differential scanning calorimetry in studies of thermal fluctuation associated with amyloid fibril formation. Biophys. Rev. 2013, 5, 259–269. [Google Scholar]
- Lin, K.; Wu, G. Isothermal Titration Calorimetry Assays to Measure Binding Affinities In Vitro. Methods Mol. Biol. 2019, 1893, 257–272. [Google Scholar] [PubMed]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: Current applications and future directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar]
- La Manna, S.; Di Natale, C.; Florio, D.; Marasco, D. Peptides as therapeutic agents for inflammatory-related diseases. Int. J. Mol. Sci. 2018, 19, 2714. [Google Scholar]
- Russo, A.; Aiello, C.; Grieco, P.; Marasco, D. Targeting “undruggable” proteins: Design of synthetic cyclopeptides. Curr. Med. Chem. 2016, 23, 748–762. [Google Scholar] [CrossRef]
- Tesauro, D.; Accardo, A.; Diaferia, C.; Milano, V.; Guillon, J.; Ronga, L.; Rossi, F. Peptide-based drug-delivery systems in biotechnological applications: Recent advances and perspectives. Molecules 2019, 24, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, E.; Diaferia, C.; Gallo, E.; Morelli, G.; Accardo, A. Stable formulations of peptide-based nanogels. Molecules 2020, 25, 3455. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Ali, R.; Onkar, A.; Ganesh, S.; Verma, S. Strategies for interference of insulin fibrillogenesis: Challenges and advances. ChemBioChem 2022, 23, e202100678. [Google Scholar] [CrossRef]
- Windheim, J.; Colombo, L.; Battajni, N.C.; Russo, L.; Cagnotto, A.; Diomede, L.; Bigini, P.; Vismara, E.; Fiumara, F.; Gabbrielli, S.; et al. Micro- and nanoplastics’ effects on protein folding and amyloidosis. Int. J. Mol. Sci. 2022, 23, 10329. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Data Type | Interaction | Ref. |
|---|---|---|---|
| AF-perylenebisimide | Atomic force microscopy (AFM) Circular dichroism (CD) Isothermal titration calorimetry (ITC) Thioflavin T fluorescence Zeta (ξ) potential measurement | Not known | [86] |
| DPNGS | ANS fluorescence Circular dichroism Molecular modeling Thioflavin T fluorescence Transmission electron microscopy (TEM) | Q4B, H10B, A14B | [87] |
| ELAQM | ANS fluorescence Circular dichroism Molecular modeling Thioflavin T fluorescence Transmission electron microscopy | H10B and E13B C11A | [87] |
| FA-perylenebisimide | Atomic force microscopy (AFM) Circular dichroism Isothermal titration calorimetry Thioflavin T fluorescence Zeta (ξ) potential measurement | Not known | [86] |
| Ferrocenyl-FX (X = F, Y) | Circular dichroism Dynamic light scattering (DLS) Molecular modeling Thioflavin T fluorescence Transmission electron microscopy | Chain B | [88] |
| Ferrocenyl-FFX (X = D, F, K, Y) | Atomic force microscopy Circular dichroism Dynamic light scattering Molecular modeling Thioflavin T fluorescence | I2A, N5A, YCA, RGFFY | [89] |
| FVPRK | Dynamic light scattering Molecular modeling Thioflavin T fluorescence Transmission electron microscopy | FFY | [90] |
| KPWWPRR | Atomic force microscopy Circular dichroism Confocal microscopy Molecular modeling Nuclear magnetic resonance (NMR) Thioflavin T fluorescence Transmission electron microscopy Tryptophan fluorescence anisotropy | E13B, YL, GEFFY | [39] |
| LVEALYL | Electron microscopy (TEM/STEM) Molecular modeling Thioflavin T fluorescence Single-crystal X-ray diffraction (XRD) | LVEALYL LYQLENY | [46] |
| NFGAIL NFGAXL (X = 2-aminobenzoic acid) | Atomic force microscopy Circular dichroism Isothermal titration calorimetry Molecular modeling Nuclear magnetic resonance Raman spectroscopy Thioflavin T fluorescence Transmission electron microscopy | EAL, L17B, E21B, G23B, F25B; G8B, V12B, Y16B, V18B, R22B, Y26B; IVA, C7A, A8A, L16A | [91] |
| NIVNVSLVK | Circular dichroism Dynamic light scattering Immunoblotting Infrared spectroscopy Scanning electron microscopy (SEM) Size exclusion chromatography (SEC) Thioflavin T fluorescence Transmission electron microscopy Turbidity | LYA, and E17A; FVNB, L6B, H10B and L17B | [92] |
| VIFYW | Acrylamide quenching fluorescence ANS fluorescence Circular dichroism Dynamic light scattering Molecular modeling Thioflavin T fluorescence Transmission electron microscopy | GIVEQ, NY, FYTPK | [93] |
| VVVVV | Acrylamide quenching fluorescence ANS fluorescence Circular dichroism Dynamic light scattering Molecular modeling Thioflavin T fluorescence Transmission electron microscopy | GIVE, YCN, L15B, YTP | [93] |
| VYYR | Atomic force microscopy Circular dichroism Dynamic light scattering Immunoblotting Isothermal titration calorimetry Molecular modeling Nuclear magnetic resonance Thioflavin T fluorescence | NQH | [94] |
| Wn-taurine (n = 1–4) | Atomic force microscopy Circular dichroism Molecular modeling Nuclear magnetic resonance Thioflavin T fluorescence | FFY | [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosetti, B.; Marchesan, S. Peptide Inhibitors of Insulin Fibrillation: Current and Future Challenges. Int. J. Mol. Sci. 2023, 24, 1306. https://doi.org/10.3390/ijms24021306
Rosetti B, Marchesan S. Peptide Inhibitors of Insulin Fibrillation: Current and Future Challenges. International Journal of Molecular Sciences. 2023; 24(2):1306. https://doi.org/10.3390/ijms24021306
Chicago/Turabian StyleRosetti, Beatrice, and Silvia Marchesan. 2023. "Peptide Inhibitors of Insulin Fibrillation: Current and Future Challenges" International Journal of Molecular Sciences 24, no. 2: 1306. https://doi.org/10.3390/ijms24021306
APA StyleRosetti, B., & Marchesan, S. (2023). Peptide Inhibitors of Insulin Fibrillation: Current and Future Challenges. International Journal of Molecular Sciences, 24(2), 1306. https://doi.org/10.3390/ijms24021306