


2,3-Diphosphoglycerate and the Protective Effect of Pyruvate Kinase Deficiency against Malaria Infection—Exploring the Role of the Red Blood Cell Membrane
 ,
,  ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
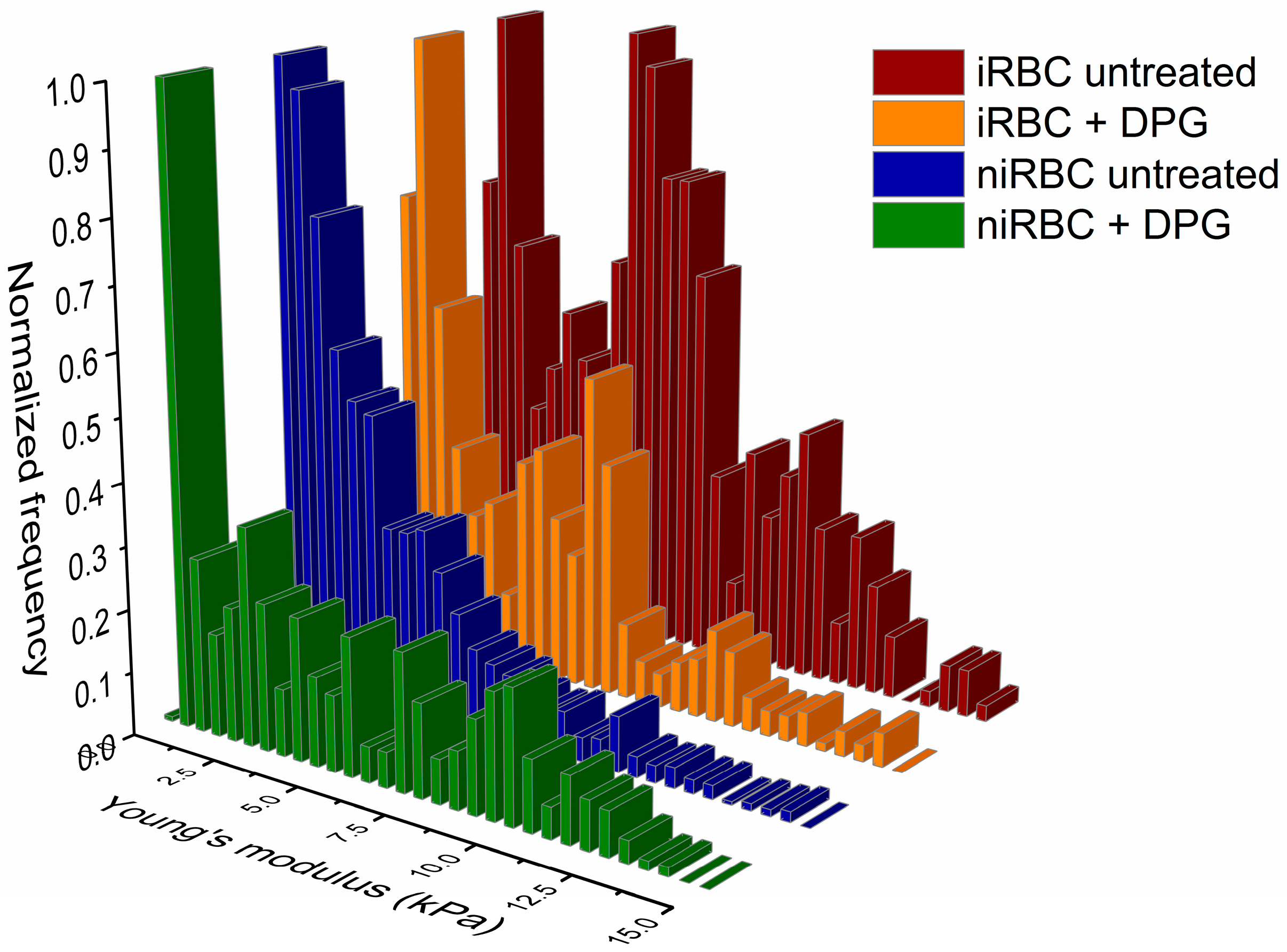
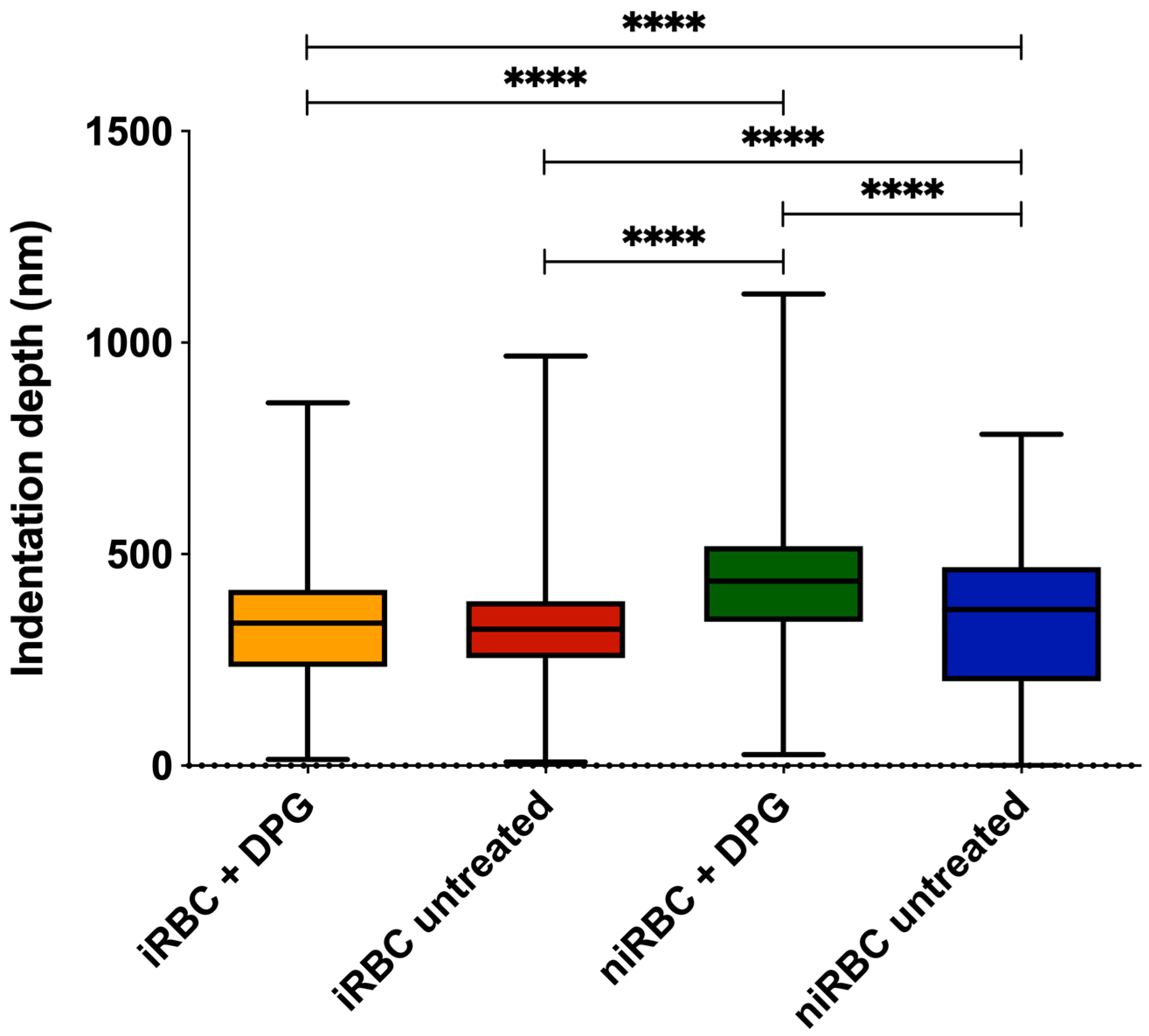
2.1. AFM Force Spectroscopy—Red Blood Cell Stiffness
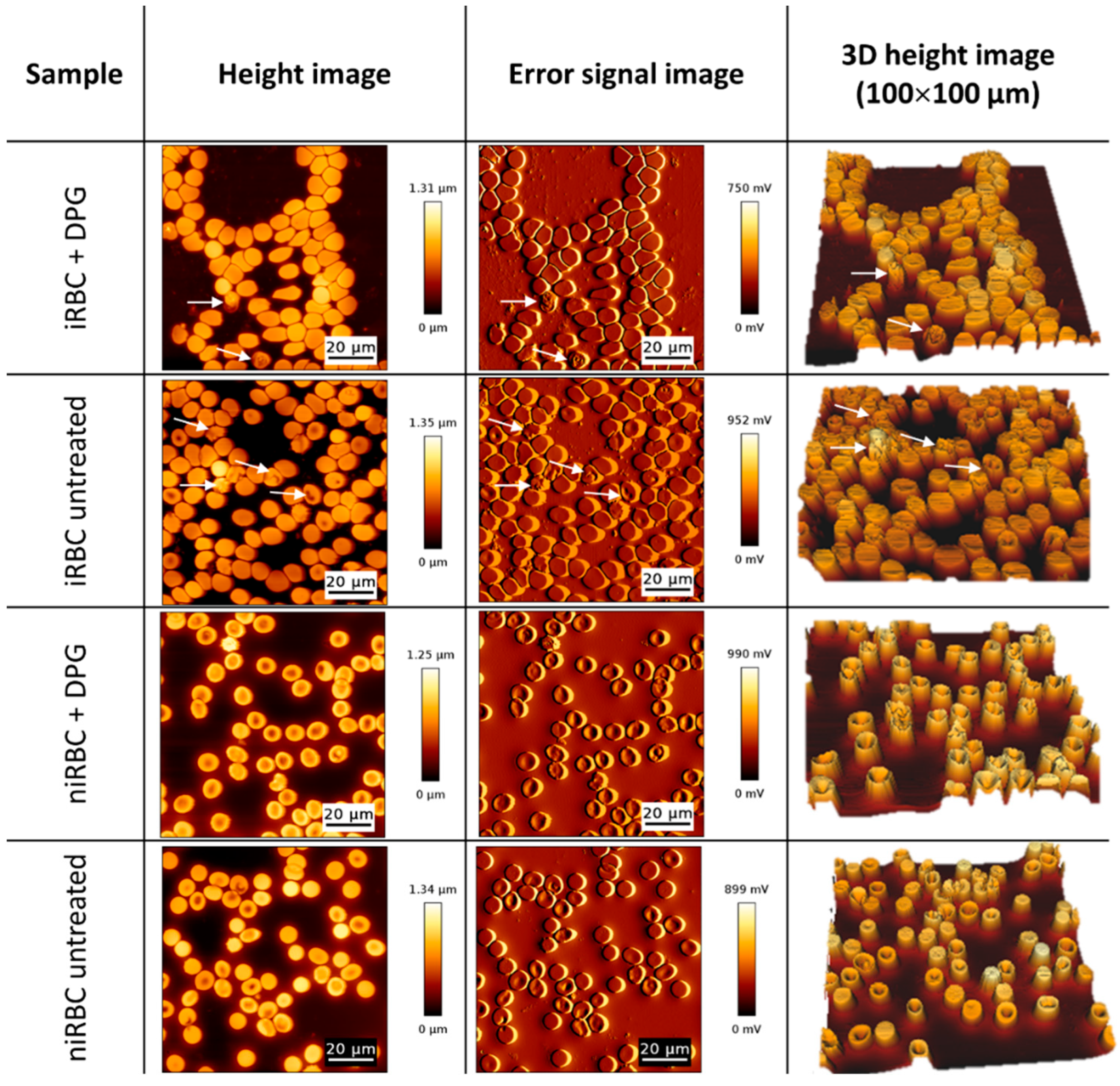
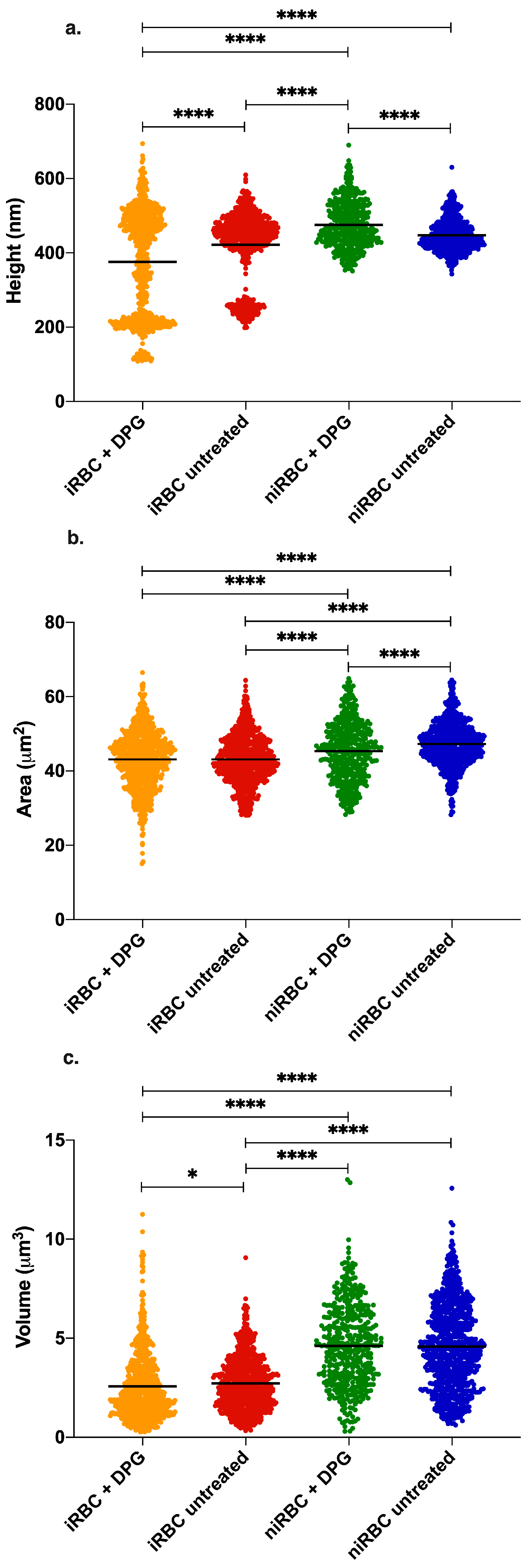
2.2. AFM Imaging
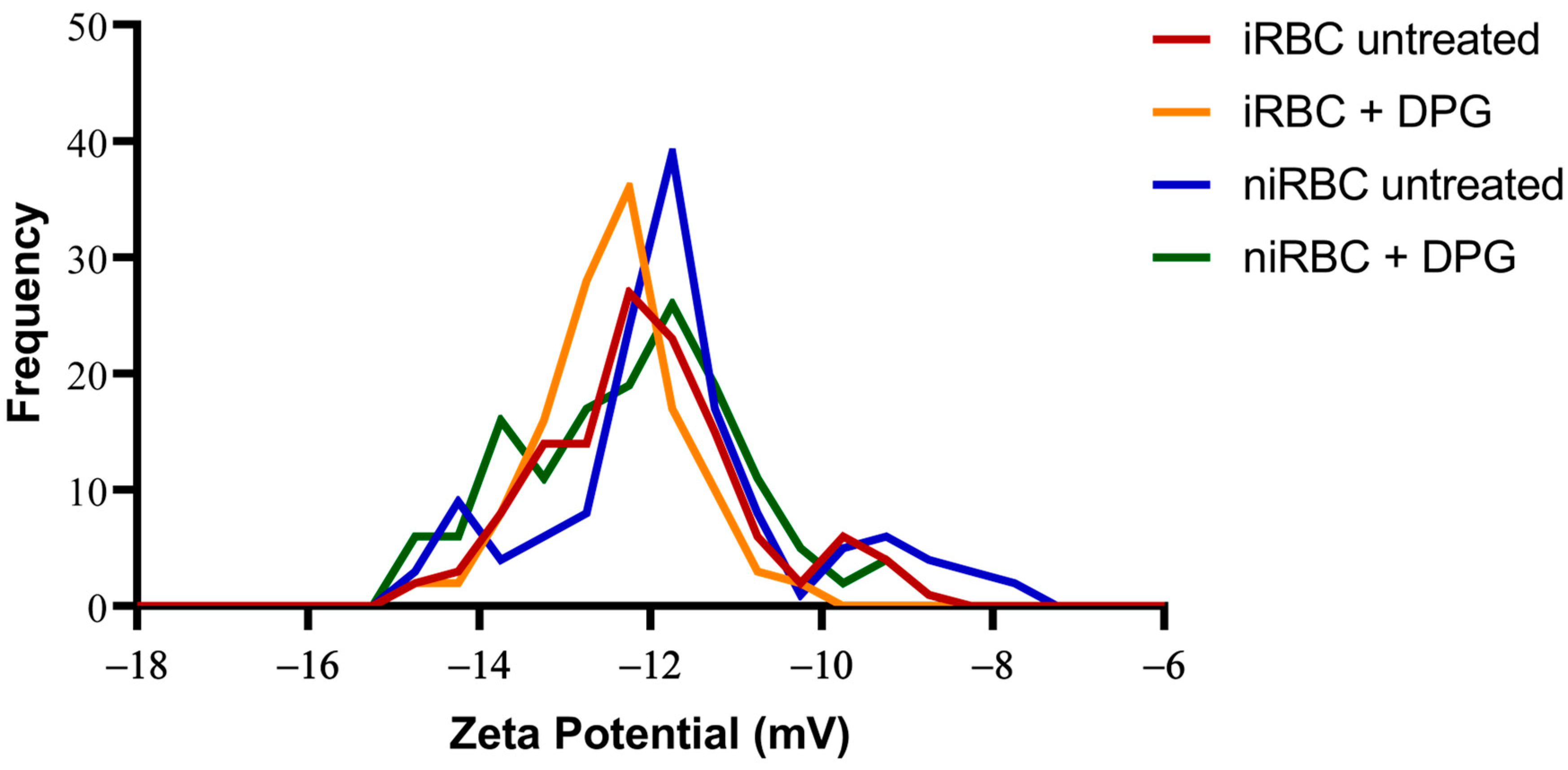
2.3. RBC Surface Charge Analysis
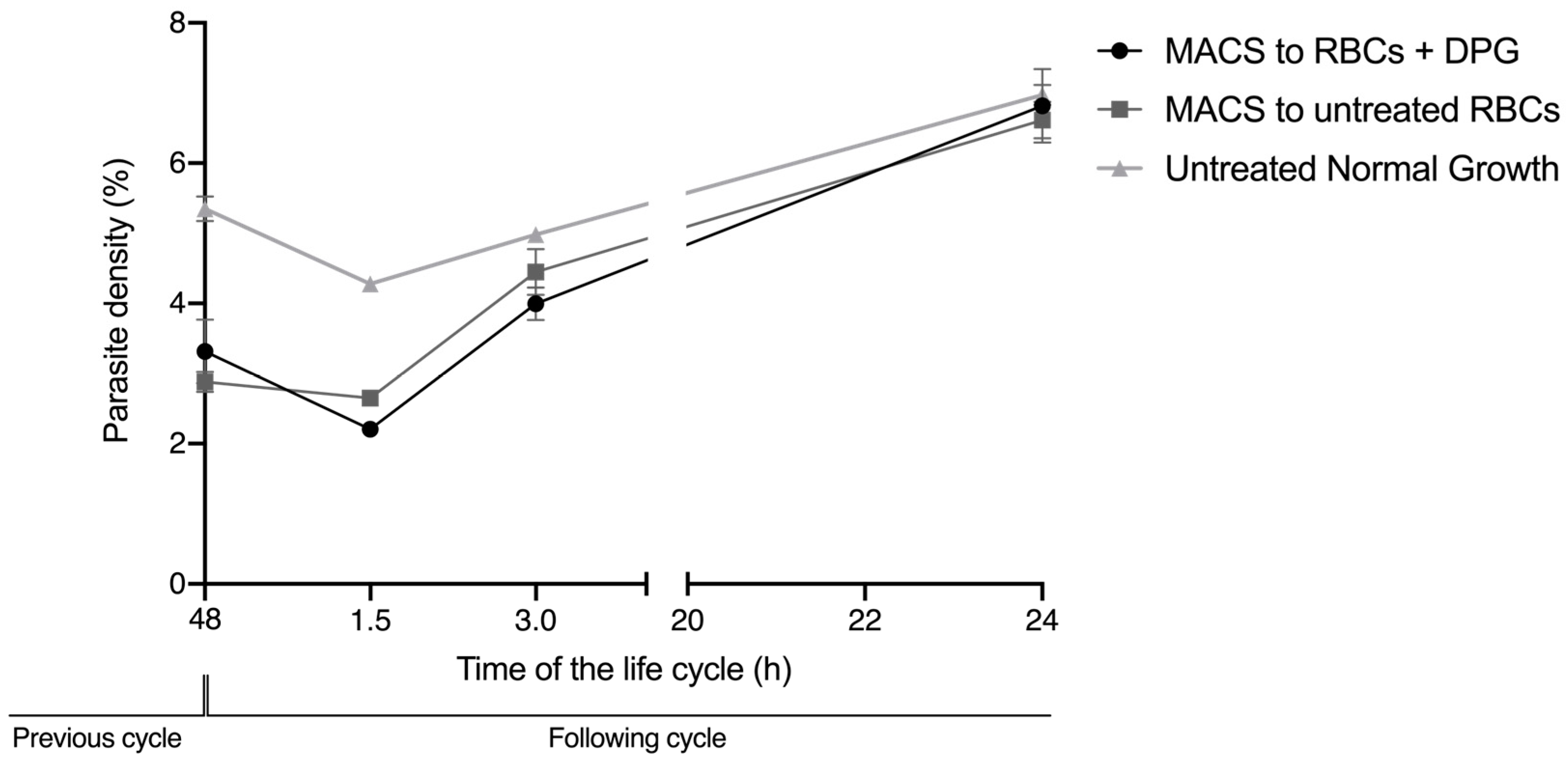
2.4. Cell Viability and Re-Invasion Assays
3. Discussion
4. Materials and Methods
4.1. Blood Donors
4.2. Plasmodium Falciparum In Vitro Cultures
4.3. Sample Treatment with 2,3-Diphosphoglycerate
4.4. Atomic Force Microscopy Assays
4.4.1. Force Spectroscopy for RBC Stiffness Analysis
4.4.2. AFM Scanning Images for Morphology Analysis
4.5. RBC Zeta Potential
4.6. Cell Viability Assay
4.7. Reinvasion Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2022; World Health Organization: Geneva, Switzerland, 2022; ISBN 978-92-4-006489-8.
- de Koning-Ward, T.F.; Dixon, M.W.A.; Tilley, L.; Gilson, P.R. Plasmodium Species: Master Renovators of Their Host Cells. Nat. Rev. Microbiol. 2016, 14, 494–507. [Google Scholar] [CrossRef]
- Lelliott, P.M.; McMorran, B.J.; Foote, S.J.; Burgio, G. The Influence of Host Genetics on Erythrocytes and Malaria Infection: Is There Therapeutic Potential? Malar. J. 2015, 14, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litwack, G. Glycolysis and Gluconeogenesis. In Human Biochemistry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 183–198. ISBN 978-0-12-383864-3. [Google Scholar]
- MacRae, J.I.; Dixon, M.W.; Dearnley, M.K.; Chua, H.H.; Chambers, J.M.; Kenny, S.; Bottova, I.; Tilley, L.; McConville, M.J. Mitochondrial Metabolism of Sexual and Asexual Blood Stages of the Malaria Parasite Plasmodium Falciparum. BMC Biol. 2013, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, M.; Sonawat, H.M.; Sharma, S. Glycolysis in Plasmodium Falciparum Results in Modulation of Host Enzyme Activities. J. Vector. Borne Dis. 2006, 43, 95–103. [Google Scholar] [PubMed]
- Durand, P.M.; Coetzer, T.L. Pyruvate Kinase Deficiency Protects against Malaria in Humans. Haematologica 2008, 93, 939–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayi, K.; Min-Oo, G.; Serghides, L.; Crockett, M.; Kirby-Allen, M.; Quirt, I.; Gros, P.; Kain, K.C. Pyruvate Kinase Deficiency and Malaria. N. Engl. J. Med. 2008, 358, 1805–1810. [Google Scholar] [CrossRef] [PubMed]
- Machado, P.; Pereira, R.; Rocha, A.M.; Manco, L.; Fernandes, N.; Miranda, J.; Ribeiro, L.; Do Rosário, V.E.; Amorim, A.; Gusmão, L.; et al. Malaria: Looking for Selection Signatures in the Human PKLR Gene Region. Br. J. Haematol. 2010, 149, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Machado, P.; Manco, L.; Gomes, C.; Mendes, C.; Fernandes, N.; Salomé, G.; Sitoe, L.; Chibute, S.; Langa, J.; Ribeiro, L.; et al. Pyruvate Kinase Deficiency in Sub-Saharan Africa: Identification of a Highly Frequent Missense Mutation (G829A;Glu277Lys) and Association with Malaria. PLoS ONE 2012, 7, e47071. [Google Scholar] [CrossRef] [Green Version]
- van Bruggen, R.; Gualtieri, C.; Iliescu, A.; Louicharoen Cheepsunthorn, C.; Mungkalasut, P.; Trape, J.-F.; Modiano, D.; Sodiomon Sirima, B.; Singhasivanon, P.; Lathrop, M.; et al. Modulation of Malaria Phenotypes by Pyruvate Kinase (PKLR) Variants in a Thai Population. PLoS ONE 2015, 10, e0144555. [Google Scholar] [CrossRef] [Green Version]
- Juel, R.; Milam, J.O. 2,3-Diphosphoglycerate: Its Role in Health and Disease. CRC Crit. Rev. Clin. Lab. Sci. 1979, 10, 113–146. [Google Scholar] [CrossRef]
- Sheetz, M.P.; Casaly, J. 2,3-Diphosphoglycerate and ATP Dissociate Erythrocyte Membrane Skeletons. J. Biol. Chem. 1980, 255, 9955–9960. [Google Scholar] [CrossRef]
- Roth, E. Plasmodium Falciparum Carbohydrate Metabolism: A Connection between Host Cell and Parasite. Blood Cells 1990, 16, 453–460; discussion 461–466. [Google Scholar]
- Morais, I.; Medeiros, M.M.; Carvalho, M.; Morello, J.; Teixeira, S.M.; Maciel, S.; Nhantumbo, J.; Balau, A.; Rosa, M.T.G.; Nogueira, F.; et al. Synthetic Red Blood Cell-Specific Glycolytic Intermediate 2,3-Diphosphoglycerate (2,3-DPG) Inhibits Plasmodium Falciparum Development In Vitro. Front. Cell. Infect. Microbiol. 2022, 12, 840968. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.R.S.; Takeuschi, M.; Yoshioka, K.; Miyamoto, H. ImagingPlasmodium Falciparum-Infected Ghost and Parasite by Atomic Force Microscopy. J. Struct. Biol. 1997, 119, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Lee, D.; Han, S.; Yang, W.; Quan, F.; Kim, K.S. Biomechanical Properties of Red Blood Cells Infected by Plasmodium berghei ANKA. J. Cell. Physiol. 2019, 234, 20546–20553. [Google Scholar] [CrossRef]
- Mauritz, J.M.A.; Esposito, A.; Tiffert, T.; Skepper, J.N.; Warley, A.; Yoon, Y.-Z.; Cicuta, P.; Lew, V.L.; Guck, J.R.; Kaminski, C.F. Biophotonic Techniques for the Study of Malaria-Infected Red Blood Cells. Med. Biol. Eng. Comput. 2010, 48, 1055–1063. [Google Scholar] [CrossRef]
- Deng, X.; Duffy, S.P.; Myrand-Lapierre, M.-E.; Matthews, K.; Santoso, A.T.; Du, Y.-L.; Ryan, K.S.; Ma, H. Reduced Deformability of Parasitized Red Blood Cells as a Biomarker for Anti-Malarial Drug Efficacy. Malar. J. 2015, 14, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillman, N.J.; Beck, J.R.; Goldberg, D.E. Protein Export into Malaria Parasite–Infected Erythrocytes: Mechanisms and Functional Consequences. Annu. Rev. Biochem. 2015, 84, 813–841. [Google Scholar] [CrossRef]
- Lavazec, C. Molecular Mechanisms of Deformability of Plasmodium -Infected Erythrocytes. Curr. Opin. Microbiol. 2017, 40, 138–144. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Feng, J.J. How Malaria Parasites Reduce the Deformability of Infected Red Blood Cells. Biophys. J. 2012, 103, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Krüger-Genge, A.; Jung, F.; Küpper, J.H.; Lehmann, C.; Franke, R.P. Actin Type and Distribution in Erythrocytes. J. Cell. Biotechnol. 2018, 3, 81–83. [Google Scholar] [CrossRef]
- Shi, H.; Liu, Z.; Li, A.; Yin, J.; Chong, A.G.L.; Tan, K.S.W.; Zhang, Y.; Lim, C.T. Life Cycle-Dependent Cytoskeletal Modifications in Plasmodium Falciparum Infected Erythrocytes. PLoS ONE 2013, 8, e61170. [Google Scholar] [CrossRef] [PubMed]
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Counihan, N.A.; Modak, J.K.; de Koning-Ward, T.F. How Malaria Parasites Acquire Nutrients from Their Host. Front. Cell Dev. Biol. 2021, 9, 649184. [Google Scholar] [CrossRef]
- Sherling, E.S.; van Ooij, C. Host Cell Remodeling by Pathogens: The Exomembrane System in Plasmodium -Infected Erythrocytes. FEMS Microbiol. Rev. 2016, 40, 701–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauritz, J.M.A.; Esposito, A.; Ginsburg, H.; Kaminski, C.F.; Tiffert, T.; Lew, V.L. The Homeostasis of Plasmodium Falciparum-Infected Red Blood Cells. PLoS Comput. Biol. 2009, 5, e1000339. [Google Scholar] [CrossRef] [Green Version]
- Zarchin, S.; Krugliak, M.; Ginsburg, H. Digestion of the Host Erythrocyte by Malaria Parasites Is the Primary Target for Quinolinecontaining Antimalarials. Biochem. Pharmacol. 1986, 35, 2435–2442. [Google Scholar] [CrossRef]
- Boulet, C.; Doerig, C.D.; Carvalho, T.G. Manipulating Eryptosis of Human Red Blood Cells: A Novel Antimalarial Strategy? Front. Cell. Infect. Microbiol. 2018, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.P.; Vestergaard, A.L.; Mikkelsen, S.A.; Mogensen, L.S.; Chalat, M.; Molday, R.S. P4-ATPases as Phospholipid Flippases—Structure, Function, and Enigmas. Front. Physiol. 2016, 7, 00275. [Google Scholar] [CrossRef] [Green Version]
- Fraser, M.; Matuschewski, K.; Maier, A.G. Of Membranes and Malaria: Phospholipid Asymmetry in Plasmodium Falciparum-Infected Red Blood Cells. Cell. Mol. Life Sci. 2021, 78, 4545–4561. [Google Scholar] [CrossRef] [PubMed]
- Zavodnik, I.B.; Piasecka, A.; Szosland, K.; Bryszewska, M. Human Red Blood Cell Membrane Potential and Fluidity in Glucose Solutions. Scand. J. Clin. Lab. Investig. 1997, 57, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Dearnley, M.; Chu, T.; Zhang, Y.; Looker, O.; Huang, C.; Klonis, N.; Yeoman, J.; Kenny, S.; Arora, M.; Osborne, J.M.; et al. Reversible Host Cell Remodeling Underpins Deformability Changes in Malaria Parasite Sexual Blood Stages. Proc. Natl. Acad. Sci. USA 2016, 113, 4800–4805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, G.J.; Piel, F.B.; Reid, C.D.; Gaston, M.H.; Ohene-Frempong, K.; Krishnamurti, L.; Smith, W.R.; Panepinto, J.A.; Weatherall, D.J.; Costa, F.F.; et al. Sickle Cell Disease. Nat. Rev. Dis. Primer 2018, 4, 18010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzounakas, V.L.; Anastasiadi, A.T.; Stefanoni, D.; Cendali, F.; Bertolone, L.; Gamboni, F.; Dzieciatkowska, M.; Rousakis, P.; Vergaki, A.; Soulakis, V.; et al. Beta Thalassemia Minor Is a Beneficial Determinant of Red Blood Cell Storage Lesion. Haematologica 2021, 107, 112–125. [Google Scholar] [CrossRef]
- Tzounakas, V.L.; Kriebardis, A.G.; Georgatzakou, H.T.; Foudoulaki-Paparizos, L.E.; Dzieciatkowska, M.; Wither, M.J.; Nemkov, T.; Hansen, K.C.; Papassideri, I.S.; D’Alessandro, A.; et al. Glucose 6-Phosphate Dehydrogenase Deficient Subjects May Be Better “Storers” than Donors of Red Blood Cells. Free Radic. Biol. Med. 2016, 96, 152–165. [Google Scholar] [CrossRef]
- Kariuki, S.N.; Williams, T.N. Human Genetics and Malaria Resistance. Hum. Genet. 2020, 139, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Trager, W.; Jensen, J.B. Human Malaria Parasites in Continuous Culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Radfar, A.; Méndez, D.; Moneriz, C.; Linares, M.; Marín-García, P.; Puyet, A.; Diez, A.; Bautista, J.M. Synchronous Culture of Plasmodium Falciparum at High Parasitemia Levels. Nat. Protoc. 2009, 4, 1899–1915. [Google Scholar] [CrossRef]
- Chen, X.; Feng, L.; Jin, H.; Feng, S.; Yu, Y. Quantification of the Erythrocyte Deformability Using Atomic Force Microscopy: Correlation Study of the Erythrocyte Deformability with Atomic Force Microscopy and Hemorheology. Clin. Hemorheol. Microcirc. 2009, 43, 241–249. [Google Scholar] [CrossRef]
- Liu, B.; Ma, J.; Gao, E.; He, Y.; Cui, F.; Xu, Q. Development of an Artificial Neuronal Network with Post-Mitotic Rat Fetal Hippocampal Cells by Polyethylenimine. Biosens. Bioelectron. 2008, 23, 1221–1228. [Google Scholar] [CrossRef]
- Lin, D.C.; Dimitriadis, E.K.; Horkay, F. Robust Strategies for Automated AFM Force Curve Analysis—I. Non-Adhesive Indentation of Soft, Inhomogeneous Materials. J. Biomech. Eng. 2007, 129, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.A.; de Oliveira, S.; Freitas, T.; Gonçalves, S.; Santos, N.C. Variations on Fibrinogen-Erythrocyte Interactions during Cell Aging. PLoS ONE 2011, 6, e18167. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. In Current Protocols in Immunology; Coligan, J.E., Bierer, B.E., Margulies, D.H., Shevach, E.M., Strober, W., Eds.; Supplement 21; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; pp. A.3B.1–A.3B.2. ISBN 978-0-471-14273-7. [Google Scholar]
- World Health Organization. Global Technical Strategy for Malaria 2016–2030; 2021 Update; World Health Organization: Geneva, Switzerland, 2021; ISBN 978-92-4-003135-7.
- Alonso, P.L. Responding to Recent Changes in the Malaria Elimination Map 2021. Available online: https://learning.edx.org/course/course-v1:HarvardX+PH425x+2T2021/block-v1:HarvardX+PH425x+2T2021+type@sequential+block@0627ebfbd4494927b2fddf815ab734bd/block-v1:HarvardX+PH425x+2T2021+type@vertical+block@9dcbb0323f07411f9258b57c1dc8a4a4 (accessed on 10 February 2022).






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, M.; Medeiros, M.M.; Morais, I.; Lopes, C.S.; Balau, A.; Santos, N.C.; Carvalho, F.A.; Arez, A.P. 2,3-Diphosphoglycerate and the Protective Effect of Pyruvate Kinase Deficiency against Malaria Infection—Exploring the Role of the Red Blood Cell Membrane. Int. J. Mol. Sci. 2023, 24, 1336. https://doi.org/10.3390/ijms24021336
Carvalho M, Medeiros MM, Morais I, Lopes CS, Balau A, Santos NC, Carvalho FA, Arez AP. 2,3-Diphosphoglycerate and the Protective Effect of Pyruvate Kinase Deficiency against Malaria Infection—Exploring the Role of the Red Blood Cell Membrane. International Journal of Molecular Sciences. 2023; 24(2):1336. https://doi.org/10.3390/ijms24021336
Chicago/Turabian StyleCarvalho, Maria, Márcia M. Medeiros, Inês Morais, Catarina S. Lopes, Ana Balau, Nuno C. Santos, Filomena A. Carvalho, and Ana Paula Arez. 2023. "2,3-Diphosphoglycerate and the Protective Effect of Pyruvate Kinase Deficiency against Malaria Infection—Exploring the Role of the Red Blood Cell Membrane" International Journal of Molecular Sciences 24, no. 2: 1336. https://doi.org/10.3390/ijms24021336
APA StyleCarvalho, M., Medeiros, M. M., Morais, I., Lopes, C. S., Balau, A., Santos, N. C., Carvalho, F. A., & Arez, A. P. (2023). 2,3-Diphosphoglycerate and the Protective Effect of Pyruvate Kinase Deficiency against Malaria Infection—Exploring the Role of the Red Blood Cell Membrane. International Journal of Molecular Sciences, 24(2), 1336. https://doi.org/10.3390/ijms24021336