
Cyto-Genotoxic and Behavioral Effects of Flubendiamide in Allium cepa Root Cells, Drosophila melanogaster and Molecular Docking Studies

 ,
,  and
and
Abstract
:1. Introduction
2. Results
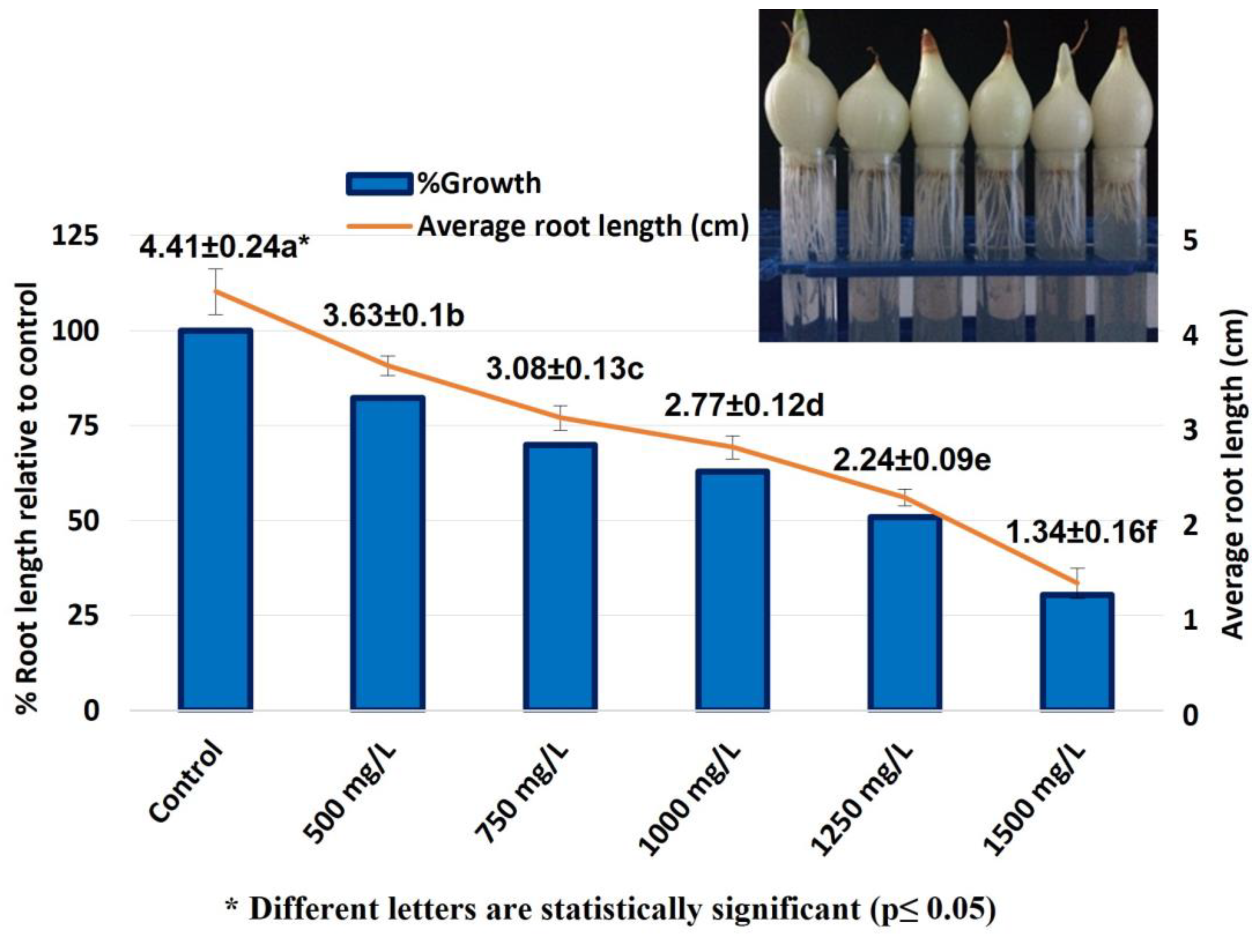
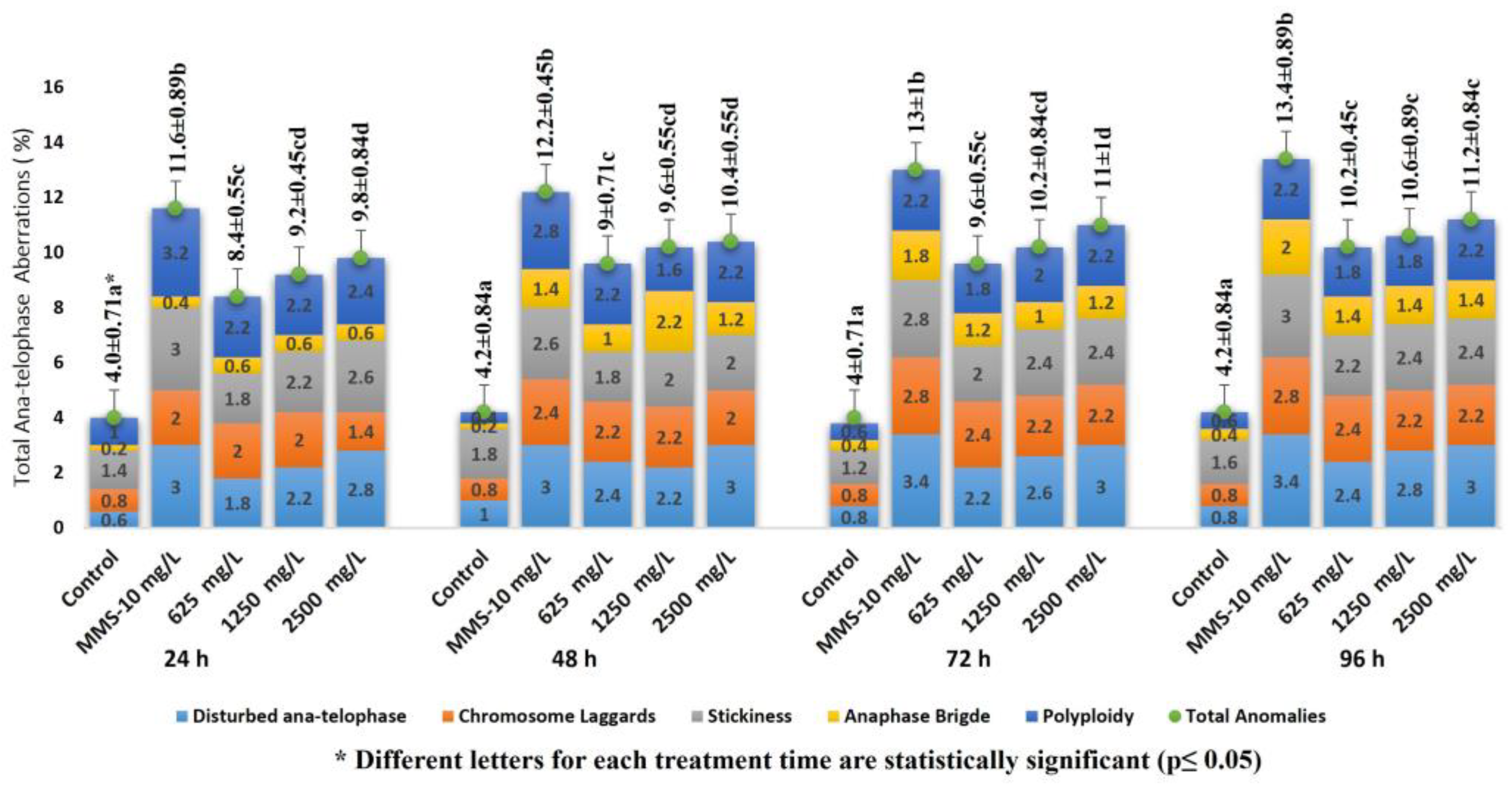
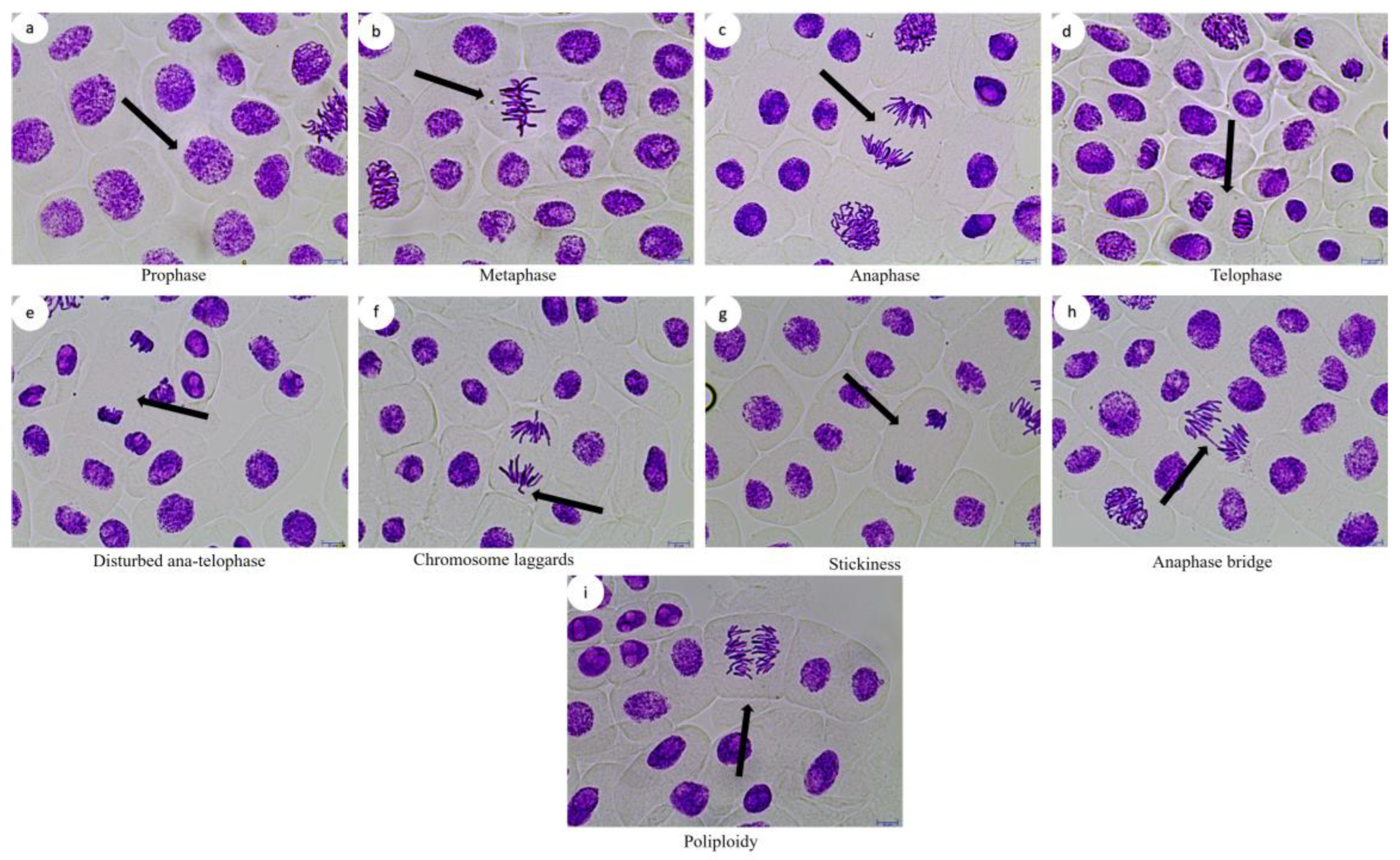
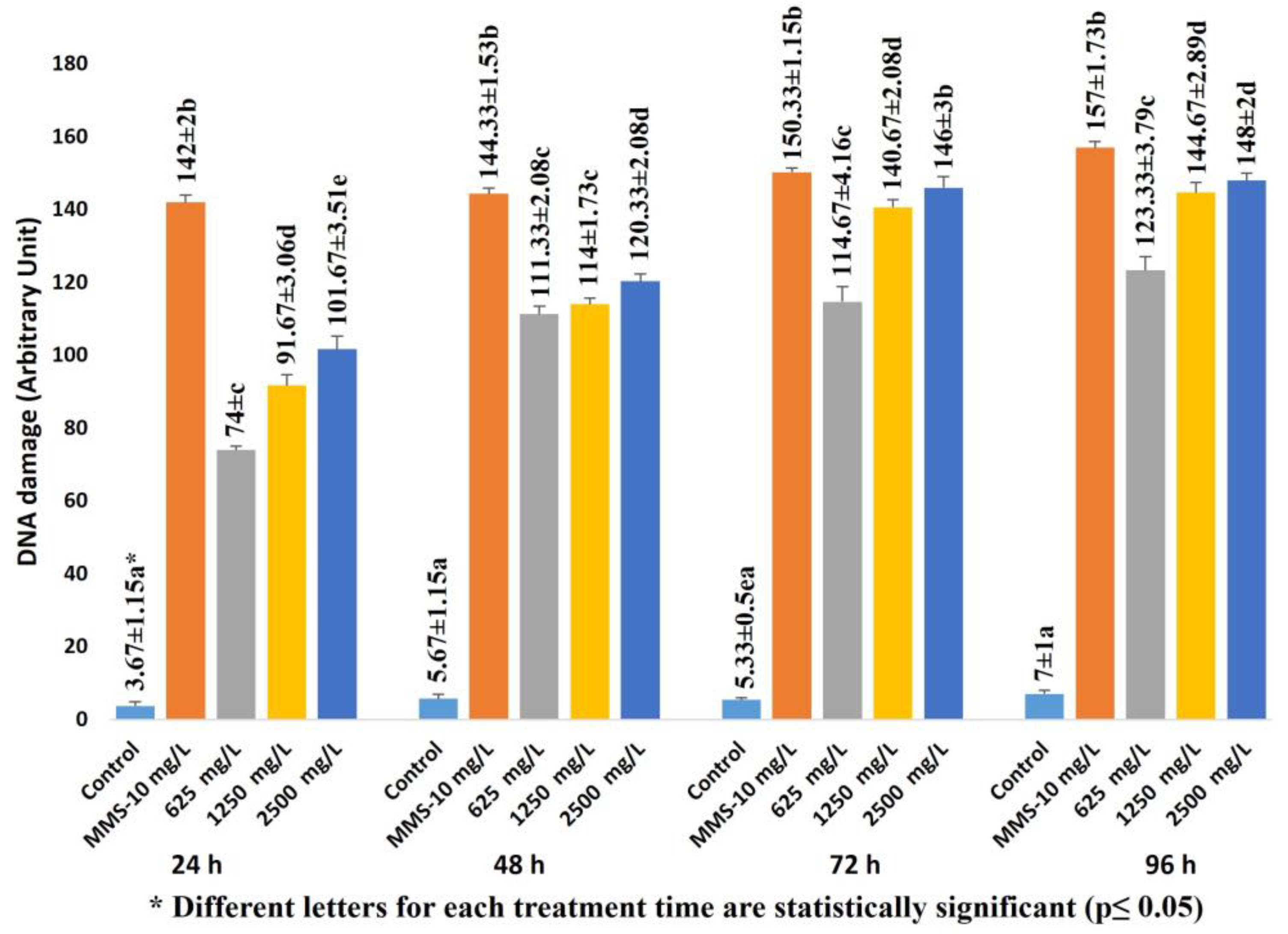
2.1. Allium Test, D. melanogaster Behavioral Assay and Comet Assay
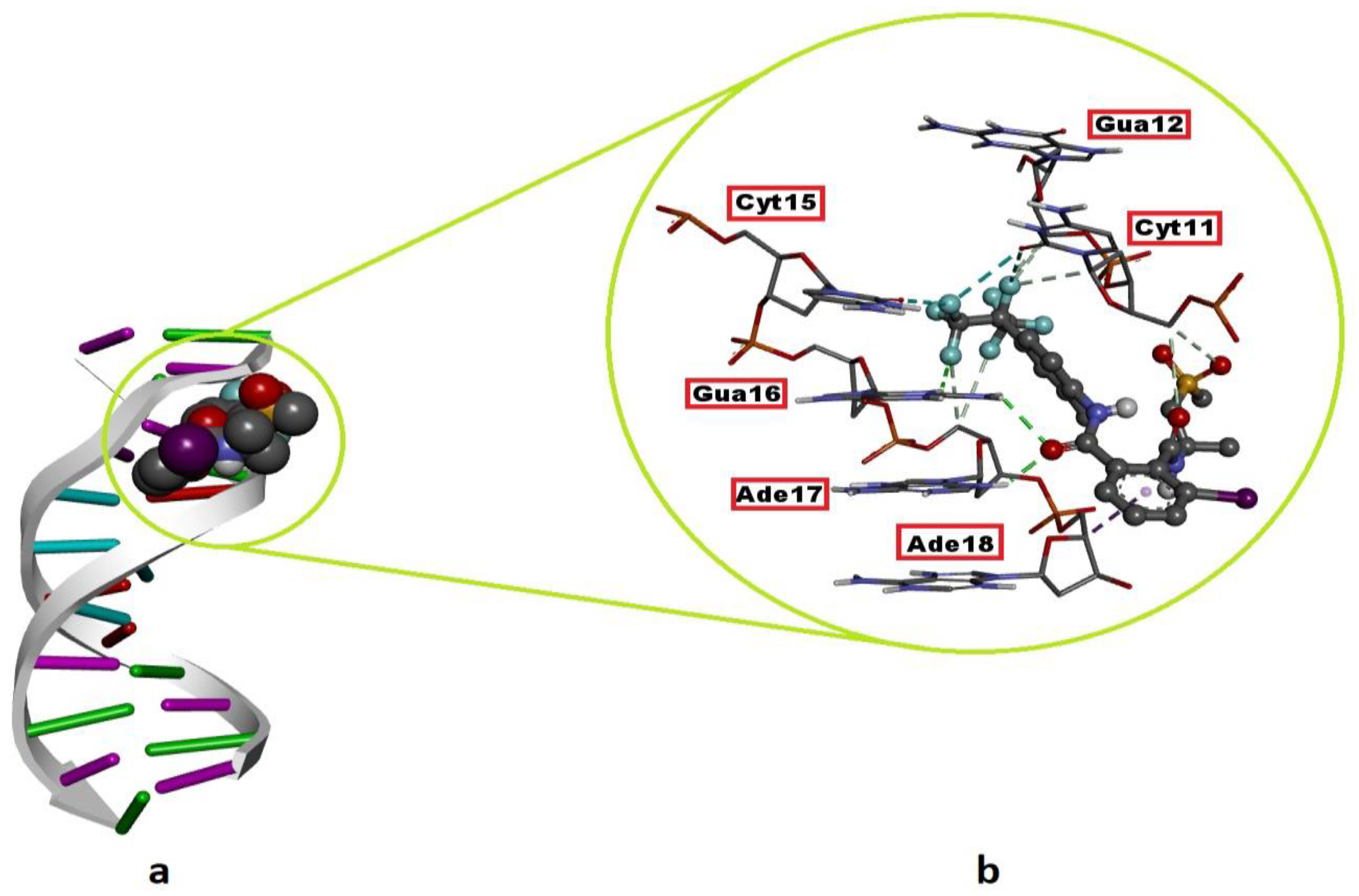
2.2. DNA–FLB Molecular Interactions
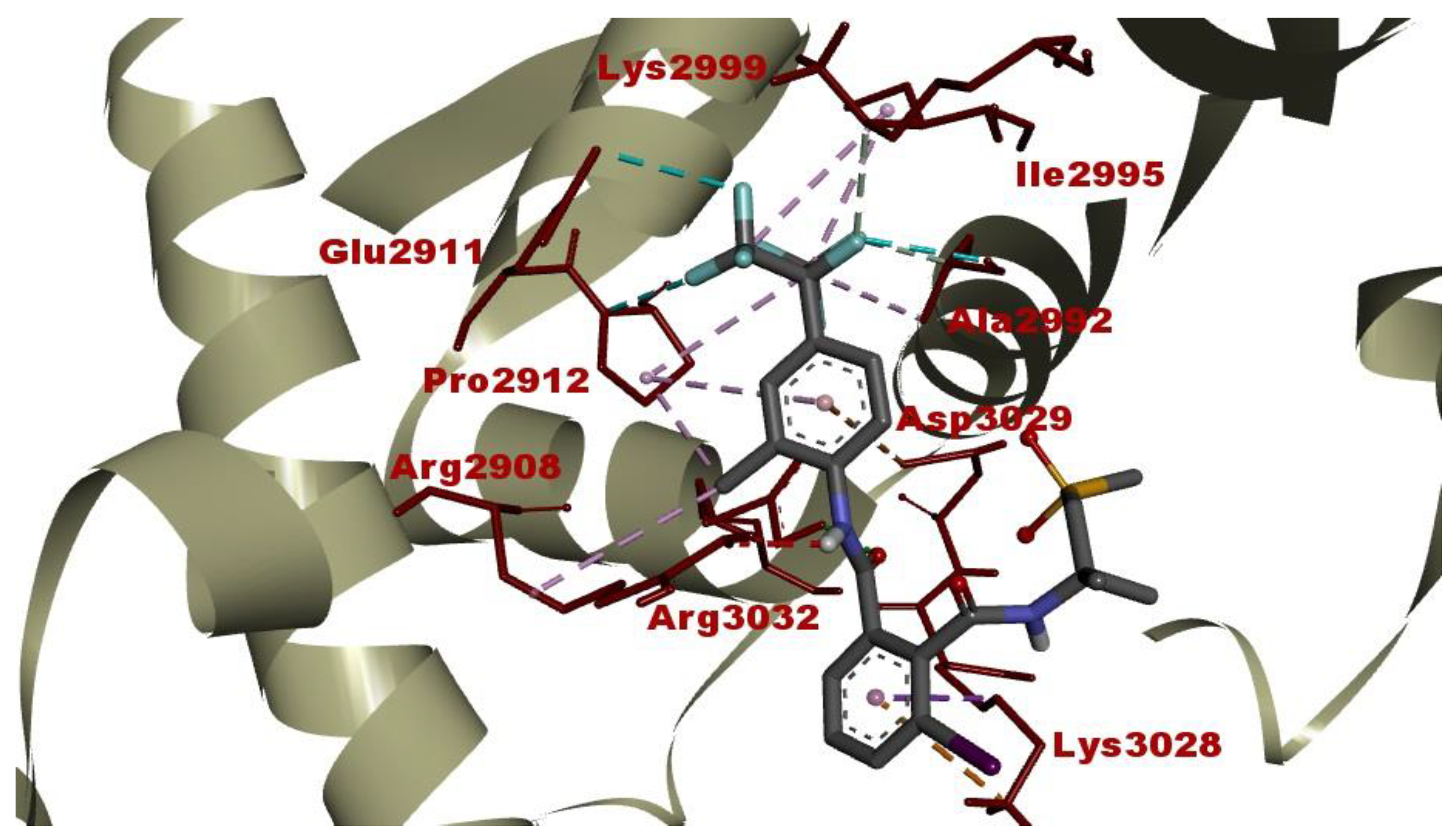
2.3. Intermolecular Interactions between FLB and Diamondback Moth (DBM) RyR Repeat34 Domain
3. Discussion
4. Materials and Methods
4.1. Allium Test
4.2. Drosophila Strain and Chemical Substances
4.2.1. Behavioral Experiments
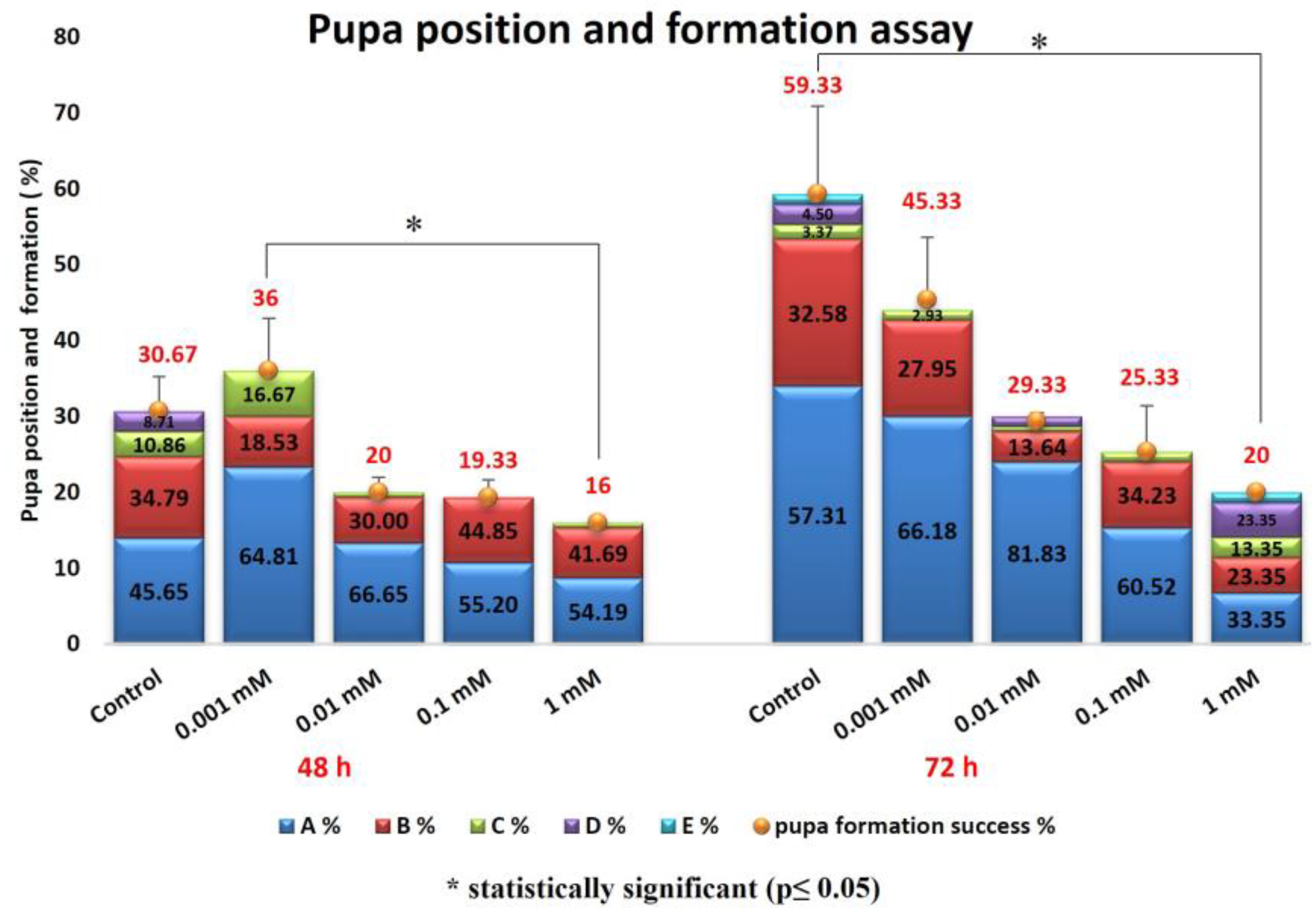
Pupa Formation Success and Pupal Position
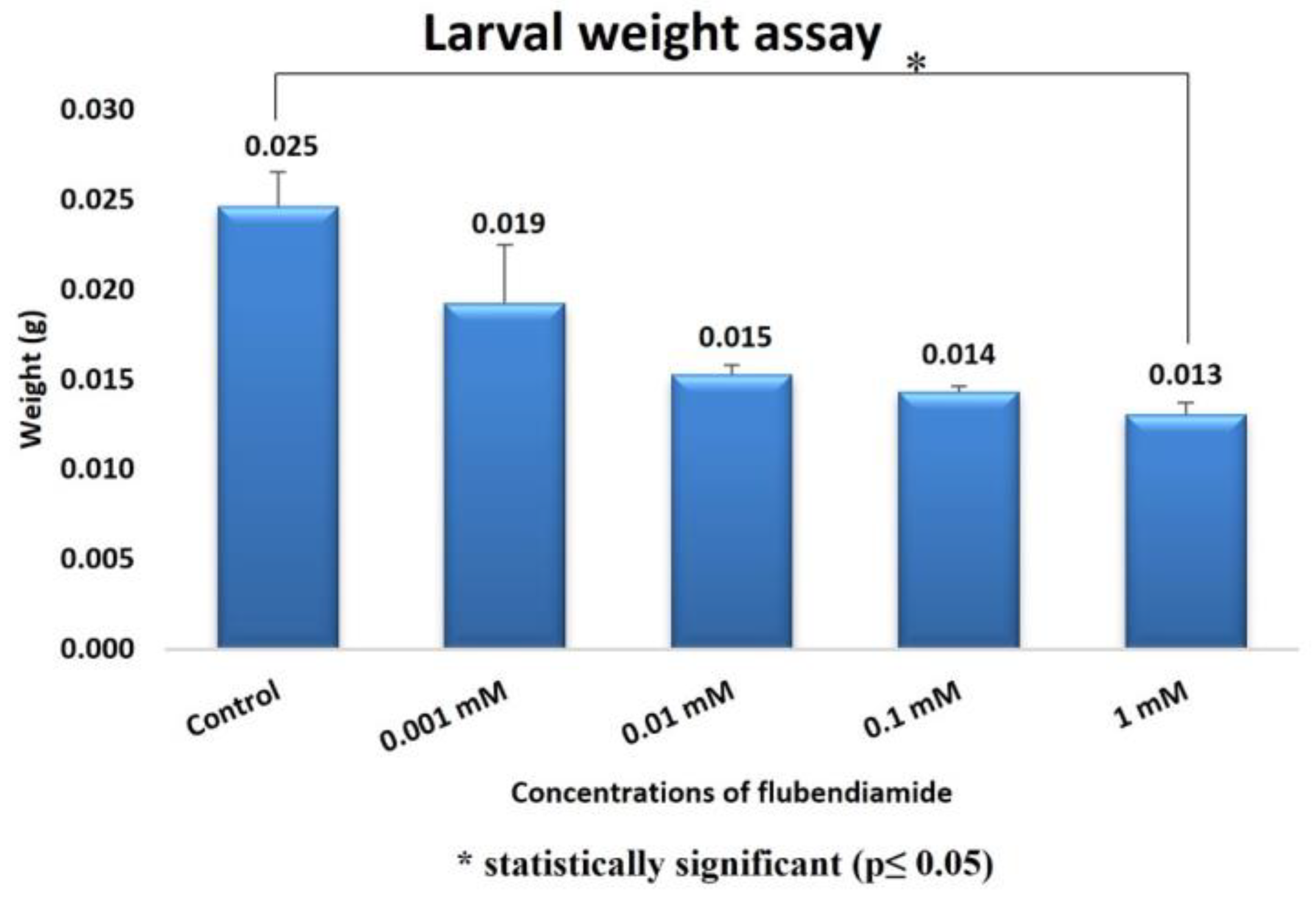
Larval Weight
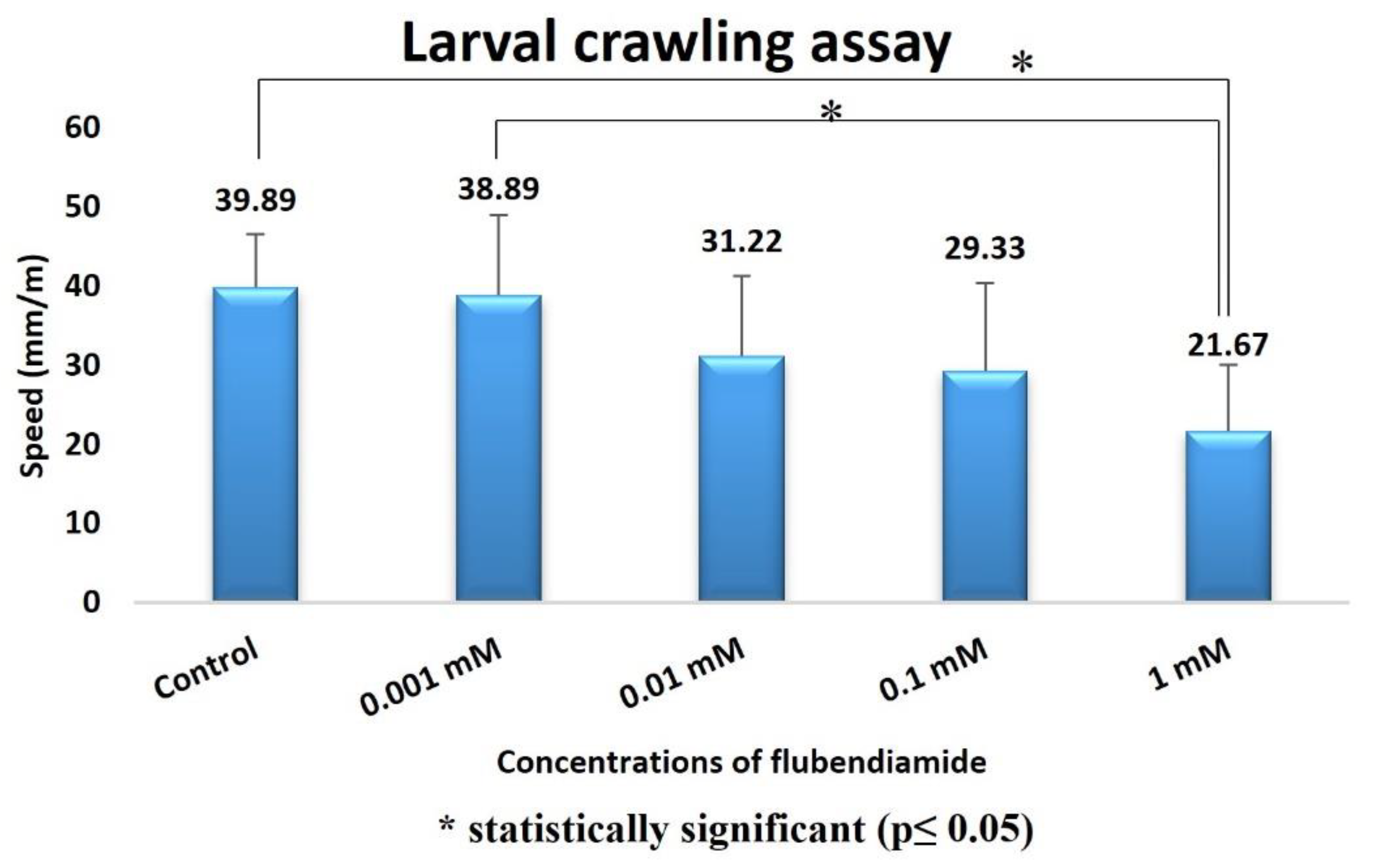
Crawling
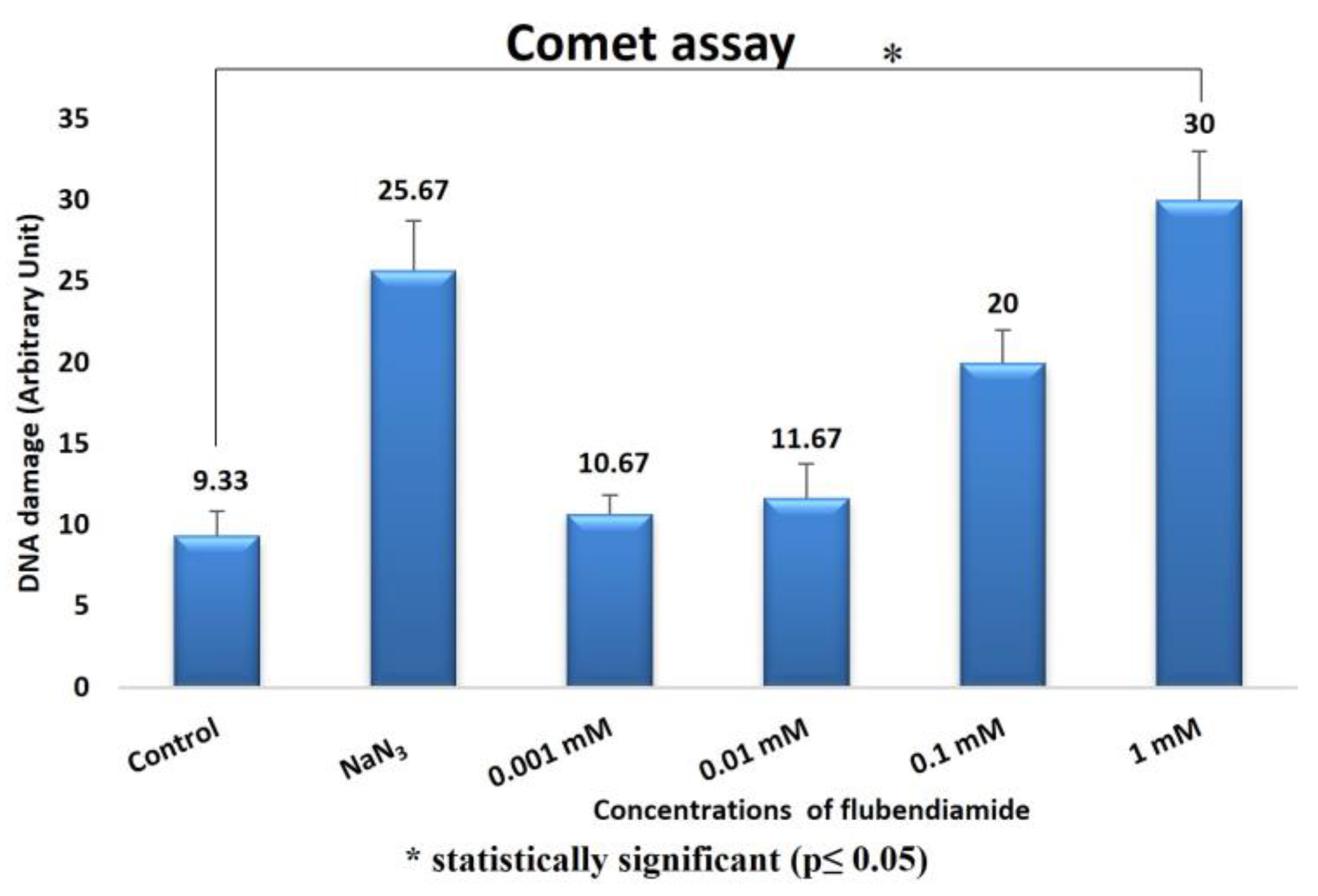
4.3. Comet Assay
4.4. Molecular Docking Studies
4.4.1. Protein and Ligand Preparation
4.4.2. Molecular Docking
4.5. Statistical Analyzes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Olszewski, K.L.; Mather, M.W.; Morrisey, J.M.; Garcia, B.A.; Vaidya, A.B.; Rabinowitz, J.D.; Llinás, M. Branched tricarboxylic acid metabolism in Plasmodium falciparum. Nature 2010, 466, 774–778. [Google Scholar] [CrossRef] [Green Version]
- Aghris, S.; Alaoui, O.T.; Laghrib, F.; Farahi, A.; Bakasse, M.; Saqrane, S.; Lahrich, S.; Mhammedi, E.L. Extraction and determination of flubendiamide insecticide in food samples: A review. Curr. Res. Food Sci. 2022, 5, 401–413. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 11193251, Flubendiamide. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Flubendiamide (accessed on 19 December 2022).
- EPA (Environmental Protection Agency). Available online: http://www.gpo.gov/fdsys/pkg/FR-2010-11-05/html/2010-27998.html (accessed on 19 September 2022).
- Sarkar, S.; Dutta, M.; Roy, S. Potential toxicity of flubendiamide in Drosophila melanogaster and associated structural alterations of its compound eye. Toxicol. Environ. Chem. 2014, 96, 1075–1087. [Google Scholar] [CrossRef]
- Iqbal, M. Vicia faba bioassay for environmental toxicity monitoring: A review. Chemosphere 2016, 144, 785–802. [Google Scholar] [CrossRef]
- Soodan, R.K.; Sharma, A.; Kaur, M.; Katnoria, J.K.; Nagpal, A.K. Allium cepa root chromosomal aberration assay: An application in assessing anti-genotoxic. Potential of ashwagandha. In Science of Ashwagandha: Preventive and Therapeutic Potentials; Kaul, S.C., Wadhwa, R., Eds.; Springer: Cham, Switzerland, 2017; pp. 69–79. [Google Scholar]
- Iqbal, M.; Abbas, M.; Nisar, J.; Nazir, A.; Qamar, A.Z. Bioassays based on higher plants as excellent dosimeters for ecotoxicity monitoring: A review. Chem. Int. 2019, 5, 1–80. [Google Scholar]
- Bakadir, K.; Kassale, A.; Barouni, K.; Lakhmiri, R.; Albourine, A. Retention of a compound of herbicides, 2,4-dichlorophenoxy acetic acid, to a soil in the absence and in the presence of Cu (II) and Zn (II) cations. J. Mater. Environ. Sci. 2016, 7, 1056–1063. [Google Scholar]
- Bonciu, E.; Firbas, P.; Fontanetti, C.S.; Wusheng, J.; Karaismailoğlu, M.C.; Liu, D.; Menicucci, F.; Pesnya, D.S.; Popescu, A.; Romanovsky, A.V.; et al. An evaluation for the standardization of the Allium cepa test as cytotoxicity assay. Caryologia 2018, 71, 191–209. [Google Scholar] [CrossRef] [Green Version]
- Özkara, A.; Akyıl, D.; Erdoğmuş, S.F.; Eren, Y.; Konuk, M. Mutagenic and cytotoxic activity of Alloxydim sodium with Allium and Ames tests. Res. J. Biothech. 2019, 14, 7–13. [Google Scholar]
- Liman, R.; Ali, M.M.; Ciğerci, İ.H.; İstifli, E.S.; Sarıkurkcu, C. Cytotoxic and genotoxic evaluation of copper oxychloride through Allium test and molecular docking studies. Environ. Sci. Pollut. Res. 2021, 28, 44998–45008. [Google Scholar] [CrossRef]
- Khallef, M.; Liman, R.; Konuk, M.; Ciğerci, İ.H.; Benouareth, D.; Tabet, M.; Abda, A. Genotoxicity of drinking water disinfection by-products (bromoform and chloroform) by using both Allium anaphase-telophase and comet tests. Cytotechnology 2015, 67, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Kassa, B.A. Cytotoxicity and Genotoxicity evaluation of municipal wastewater discharged into the head of Blue Nile River using the Allium cepa test. Sci. Afr. 2021, 13, 911. [Google Scholar] [CrossRef]
- Kurşun, A.Y.; Yalçın, B.; Güneş, M.; Tagorti, G.; Kaya, B. MgO nanopartiküllerinin Drosophila melanogaster üzerindeki davranışsal toksisitesinin değerlendirilmesi. AKU J. Sci. Eng. 2021, 21, 1283–1294. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef]
- Liman, R.; Açıkbaş, Y.; Ciğerci, İ.H. Cytotoxicity and genotoxicity of cerium oxide micro and nanoparticles by Allium and Comet tests. Ecotoxicol. Environ. Safety. 2019, 168, 408–414. [Google Scholar] [CrossRef]
- Graves, A.P.; Brenk, R.; Shoichet, B.K. Decoys for docking. J. Med. Chem. 2005, 48, 3714–3728. [Google Scholar] [CrossRef]
- Ricci, C.G.; Netz, P.A. Docking studies on DNA-ligand interactions: Building and application of a protocol to identify the binding mode. J. Chem. Inf. Model. 2009, 49, 1925–1935. [Google Scholar] [CrossRef]
- Gilad, Y.; Senderowitz, H. Docking studies on DNA intercalators. J. Chem. Inf. Model. 2014, 54, 96–107. [Google Scholar] [CrossRef]
- Wang, G.; Zhu, W. Molecular docking for drug discovery and development: A widely used approach but far from perfect. Future Med. Chem. 2016, 8, 1707–1710. [Google Scholar] [CrossRef] [Green Version]
- Torres, P.H.; Sodero, A.C.; Jofily, P.; Silva, F.P., Jr. Key topics in molecular docking for drug design. Int. J. Mol. Sci. 2019, 20, 4574. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Roy, A.; Roy, S. Flubendiamide affects visual and locomotory activities of Drosophila melanogaster for three successive generations (P, F1 and F2). Invert. Neurosci. 2018, 18, 6. [Google Scholar] [CrossRef]
- Kadala, A.; Charreton, M.; Collet, C. Flubendiamide, the first phthalic acid diamide insecticide, impairs neuronal calcium signalling in the honey bee’s antennae. J. Ins. Physiol. 2020, 125, 104086. [Google Scholar] [CrossRef]
- Mandil, R.; Prakash, A.; Rahal, A.; Singh, S.P.; Sharma, D.; Kumar, R.; Garg, S.K. In vitro and in vivo effects of flubendiamide and copper on cyto-genotoxicity, oxidative stress and spleen histology of rats and its modulation by resveratrol, catechin, curcumin and α-tocopherol. BMC Pharmacol. Toxicol. 2020, 21, 29. [Google Scholar] [CrossRef]
- Webster, P.L.; Macleod, R.D. The root apical meristem and its margin. In Plant Roots. The Hidden Half, 2nd ed.; Waishel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker: New York, NY, USA, 1996; pp. 51–76. [Google Scholar]
- Fusconi, A.; Repetto, O.; Bona, E.; Massa, N.; Gallo, C.; Dumas-Gaudot, E.; Berta, G. Effect of cadmium on meristem activity and nucleus ploidy in roots of Pisum sativum L. cv. Frisson seedlings. Environ. Exp. Bot. 2006, 58, 253–260. [Google Scholar] [CrossRef]
- Sudhakar, R.; Gowda, N.; Venu, G. Mitotic abnormalities induced by silk dyeing industry effluents in the cells of Allium cepa. Cytologia 2001, 66, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.; Mishra, K.; Srivastava, S.; Kumar, A. Cytotoxic assessment of chromium and arsenic using chromosomal behavior of root meristem in Allium cepa L. Bull. Environ. Contam. Toxicol. 2018, 100, 803–808. [Google Scholar] [CrossRef]
- El-Ghamery, A.A.; El-Nahas, A.L.; Mansour, M.M. The action of atrazine herbicide as an indicator of cell division on chromosomes and nucleic acid content in root meristems of Allium cepa and Vicia faba. Cytologia 2000, 65, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Evseeva, T.I.; Stanislav, A.; Geras’kin, I.; Shuktomova, I. Genotoxicity and toxicity assay of water sampled from a radium production industry storage cell territory by means of Allium-Test. J. Environ. Rad. 2005, 68, 235–248. [Google Scholar] [CrossRef]
- Kumari, M.; Mukherjee, A.; Chandrasekaran, N. Genotoxicity of silver nanoparticles in Allium cepa. Sci. Total Environ. 2009, 407, 5243–5246. [Google Scholar] [CrossRef]
- Singh, D.; Roy, B.K. Evaluation of malathion-induced cytogenetical effects and oxidative stress in plants using Allium test. Acta Physiol. Plant. 2017, 39, 92–102. [Google Scholar] [CrossRef]
- Fiskesjö, G.; Levan, A. Evaluation of the first ten MEIC chemicals in the Allium test. Animals 1993, 21, 139–149. [Google Scholar] [CrossRef]
- Türkoğlu, Ş. Evaluation of genotoxic effects of five flavour enhancers (glutamates) on the root meristem cells of Allium cepa. Toxicol. Indust. Health 2015, 31, 792–801. [Google Scholar] [CrossRef]
- El-Ghamery, A.A.; Mousa, M.A. Investigation on the effect of benzyladenine on the germination, radicle growth and meristematic cells of Nigella sativa L. and Allium cepa L. Ann. Agric. Sci. 2017, 62, 11–21. [Google Scholar] [CrossRef]
- Lua, A.C.; Yang, T.; Guo, J. Effects of pyrolysis conditions on the properties of activated carbons prepared from pistachio-nut shells. J. Anal. Appl. Pyrol. 2004, 72, 279–287. [Google Scholar] [CrossRef]
- Fernandes, T.C.C.; Mazzeo, D.E.C.; Marin-Morales, M.A. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pest. Biochem. Physiol. 2007, 88, 252–259. [Google Scholar] [CrossRef]
- Isaacs, A.K.; Qi, S.; Sarpong, R.; Casida, J.E. Insect ryanodine receptor: Distinct but coupled insecticide binding sites for [N-C3H3] chlorantraniliprole, flubendiamide, and [3H] ryanodine. Chem. Res. Toxicol. 2012, 25, 1571–1573. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Khatun, S.; Dutta, M.; Roy, S. Trans-generational transmission of altered phenotype resulting from flubendiamide-induced changes in apoptosis in larval imaginal discs of Drosophila melanogaster. Environ. Toxicol. Pharmacol. 2017, 56, 350–360. [Google Scholar] [CrossRef]
- Liu, L.; MacKenzie, K.R.; Putluri, N.; Maletić-Savatić, M.; Bellen, H.J. The glia-neuron lactate shuttle and elevated ros promote lipid synthesis in neurons and lipid droplet accumulation in glia via APOE/D. Cell Metab. 2017, 26, 719–737. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Dumka, V.K.; Ranjan, R. Chronic flubendiamide exposure induces oxidative stress in water buffalo (Bubalus bubalis) calves. Curr. Sci. 2018, 114, 1610–1612. [Google Scholar] [CrossRef]
- Truong, L.; Saili, K.S.; Miller, J.M.; Hutchison, J.E.; Tanguay, R.L. Persistent adult zebrafish behavioral deficits results from acute embryonic exposure to gold nanoparticles. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Fauzi, A.; Zubaidah, S.; Susanto, H. The study of larva and adult behavior of Drosophila melanogaster: Do strains affect behavior? In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2020; Volume 2231, pp. 1–7. [Google Scholar]
- Sisodia, S.; Singh, B.N. Behaviour genetics of Drosophila: Non-sexual behaviour. J. Genetics 2005, 84, 195–216. [Google Scholar] [CrossRef]
- Chen, T.H.; Lin, C.C.; Meng, P.J. Zinc oxide nanoparticles alter hatching and larval activity in zebrafih (Danio rerio). J. Hazard. Mat. 2014, 277, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Preston, R.J.; Ross, J.A. DNA-Reactive Agents. In Comprehensive Toxicology, 2nd ed.; McQueen, C.A., Bond, J., Eds.; Elsevier: Kidlington, UK, 2010; Volume 1, pp. 349–360. [Google Scholar]
- Vijendravarma, R.K.; Narasimha, S.; Steinfath, E.; Clemens, J.; Leopold, P. Data for Drosophila females have an acoustic preference for symmetric males. Proc. Natl. Acad. Sci. USA 2022, 119, e2116136119. [Google Scholar] [CrossRef] [PubMed]
- Silva Bustos, B.A.; Hidalgo Sotelo, S.I.; Campusano, J.M. Dop1R1, a type 1 dopaminergic receptor expressed in Mushroom Bodies, modulates Drosophila larval locomotion. PLoS ONE 2020, 15, e0229671. [Google Scholar]
- Collins, A.R. The comet assay for DNA damage and repair: Principles, applications and limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, H.; Chen, Z.; Lin, Y.; Lin, W.; Liu, T.; Wang, X.; Yang, L. Biochemical toxicity and cytotoxicity of flubendiamide on earthworms (Eisenia fetida). Asian J. Ecotoxicol. 2017, 12, 293–301. [Google Scholar]
- Rucińska, R.; Sobkowiak, R.; Gwóźdź, E.A. Genotoxicity of lead in lupin root cells as evaluated by the comet assay. Cell Mol. Biol. Lett. 2004, 9, 519–528. [Google Scholar]
- Yıldız, M.; Ciğerci, İ.H.; Konuk, M.; Fidan, A.F.; Terzi, H. Determination of genotoxic effects of copper sulphate and cobalt chloride in Allium cepa root cells by chromosome aberration and comet assays. Chemosphere 2009, 75, 934–938. [Google Scholar] [CrossRef]
- Rebbeck, R.T.; Essawy, M.M.; Nitu, F.R.; Grant, B.D.; Gillispie, G.D.; Thomas, D.D.; Bers, D.M.; Cornea, R.L. High-throughput screens to discover small-molecule modulators of ryanodine receptor calcium release channels. J. Biomol. Screen. 2017, 22, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Hao, Z.; Cao, P.; Yuchi, Z. Homology modeling and docking study of diamondback moth ryanodine receptor reveals the mechanisms for channel activation, insecticide binding and resistance. Pest. Manag. Sci. 2020, 76, 1291–1303. [Google Scholar] [CrossRef]
- Xu, T.; Yuchi, Z. Crystal structure of diamondback moth ryanodine receptor Repeat34 domain reveals insect-specific phosphorylation sites. BMC Biol. 2019, 17, 77. [Google Scholar] [CrossRef] [Green Version]
- Nareshkumar, B.; Akbar, S.M.; Sharma, H.C.; Jayalakshmi, S.K.; Sreeramulu, K. Evaluation of flubendiamide-induced mitochondrial dysfunction and metabolic changes in Helicoverpa armigera (Hubner). Arch. Insect Biochem. Physiol. 2017, 96, 21401. [Google Scholar] [CrossRef] [PubMed]
- Fiskesjö, G. The Allium test as a standard in environmental monitoring. Hereditas 1985, 102, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Ciğerci, İ.H.; Liman, R.; Özgül, E.; Konuk, M. Genotoxicity of indium tin oxide by Allium and Comet tests. Cytotechnology 2015, 67, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Shang, J.; Yan, L.; Wei, T.; Xiang, L.; Wang, H.; Cheng, J.; Xiao, G. Oxidative stress caused by Lead (Pb) induces iron deficiency in Drosophila melanogaster. Chemosphere 2020, 243, 125428. [Google Scholar] [CrossRef]
- Dhar, G.; Mukherjee, S.; Nayak, N.; Sahu, S.; Bag, J.; Rout, R.; Mishra, M. Various Behavioural Assays to Detect the Neuronal Abnormality in Flies. In Fundamental Approaches to Screen Abnormalities in Drosophila; Mishra, M., Ed.; Springer: New York, NY, USA, 2020; pp. 223–251. [Google Scholar]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.-C.; Sasaki, Y.F. Single Cell Gel/Comet Assay: Guidelines for in vitro and in vivo genetic toxicology T-testing. Environ. Molecul. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Irving, P.; Ubeda, J.M.; Doucet, D.; Troxler, L.; Lagueux, M.; Zachary, D.; Hoffmann, J.A.; Hetru, C.; Meister, M. New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell Microbiol. 2005, 7, 335–350. [Google Scholar] [CrossRef]
- Drew, H.R.; Wing, R.M.; Takano, T.; Broka, C.; Tanaka, S.; Itakura, K.; Dickerson, R.E. Structure of a B-DNA dodecamer: Conformation and dynamics. Proc. Nat. Acad. Sci. USA 1981, 78, 2179–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine receptors: Structure, expression, molecular details, and function in calcium release. Cold Spring Harb. Perspect. Biol. 2010, 2, a003996. [Google Scholar] [CrossRef] [Green Version]
- Crystal Structure of Diamondback Moth Ryanodine Receptor Phosphorylation Domain (2836–3050) Mutant S2946D. Available online: https://www.rcsb.org/structure/6J6P (accessed on 15 October 2022).
- Biovia, D.S. Discovery studio. Dassault Systèmes BIOVIA. 2016. Available online: https://discover.3ds.com/discovery-studio-visualizer-download (accessed on 10 October 2022).
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedretti, A.; Villa, L.; Vistoli, G. VEGA–an open platform to develop chemo-bio-informatics applications, using plug-in architecture and script programming. J. Comput.-Aided Mol. Des. 2004, 18, 167–173. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Chem. Inf. Model. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Chem. Inf. Model. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2. 0: New docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/L) | CCN | MI ± SD * | Phase Index (%) ± SD * | |||
|---|---|---|---|---|---|---|
| Prophase | Metaphase | Anaphase | Telophase | |||
| Control: 24 h | 5097 | 66.82 ± 0.74 a | 90.58 ± 0.67 a | 1.97 ± 0.26 a | 2.11 ± 0.28 a | 5.34 ± 0.43 a |
| MMS-10 | 5102 | 57.19 ± 0.61 b | 88.76 ± 0.66 b | 1.88 ± 0.26 ab | 2.92 ± 0.17 b | 6.44 ± 0.73 b |
| 625 | 5080 | 63.86 ± 0.52 c | 89.18 ± 0.66 b | 1.88 ± 0.13 ab | 2.56 ± 0.17 c | 6.38 ± 0.65 b |
| 1250 | 5102 | 61.64 ± 0.45 d | 89.32 ± 0.98 b | 1.65 ± 0.17 bc | 2.35 ± 0.26 ac | 6.67 ± 0.78 b |
| 2500 | 5100 | 60.63 ± 0.57 e | 88.9 ± 0.82 b | 1.59 ± 0.14 c | 2.17 ± 0.17 a | 7.35 ± 0.85 b |
| Control: 48 h | 5092 | 68.56 ± 0.59 a | 89.86 ± 0.84 a | 1.86 ± 0.21 a | 2.29 ± 0.32 a | 5.99 ± 0.46 a |
| MMS-10 | 5105 | 55.38 ± 0.52 b | 88.36 ± 0.43 b | 1.74 ± 0.25 ab | 2.94 ± 0.21 b | 6.97 ± 0.64 b |
| 625 | 5086 | 62.27 ± 0.49 c | 89.77 ± 0.29 a | 1.61 ± 0.2 ab | 1.99 ± 0.18 c | 6.63 ± 0.32 ab |
| 1250 | 5079 | 60.86 ± 0.35 d | 89.07 ± 0.61 ab | 1.81 ± 0.26 ab | 1.85 ± 0.2 c | 7.28 ± 0.68 b |
| 2500 | 5087 | 58.92 ± 0.81 e | 89.22 ± 0.63 a | 1.53 ± 0.12 b | 1.8 ± 0.12 c | 7.44 ± 0.69 b |
| Control: 72 h | 5110 | 68.3 ± 0.66 a | 90.06 ± 0.62 a | 1.89 ± 0.18 a | 2.32 ± 0.28 a | 5.73 ± 0.42 a |
| MMS-10 | 5092 | 53.91 ± 0.79 b | 87.8 ± 0.93 b | 1.89 ± 0.27 a | 2.99 ± 0.2 b | 7.32 ± 0.74 bc |
| 625 | 5086 | 59.97 ± 0.5 c | 89.34 ± 0.78 ac | 1.71 ± 0.18 ab | 2.26 ± 0.22 a | 6.7 ± 0.76 b |
| 1250 | 5093 | 58.97 ± 0.71 d | 89.31 ± 0.31 ac | 1.56 ± 0.18 b | 2.06 ± 0.26 a | 7.06 ± 0.32 bc |
| 2500 | 5118 | 56.94 ± 0.7 e | 89.05 ± 0.29 c | 1.82 ± 0.15 ab | 1.61 ± 0.09 c | 7.51 ± 0.39 c |
| Control: 96 h | 5106 | 68.12 ± 0.5 a | 90.14 ± 0.65 a | 1.72 ± 0.14 a | 2.21 ± 0.22 a | 5.92 ± 0.53 a |
| MMS-10 | 5081 | 52.88 ± 0.57 b | 87.72 ± 0.3 b | 1.97 ± 0.22 ab | 2.64 ± 0.27 b | 7.66 ± 0.49 b |
| 625 | 5072 | 58.4 ± 0.73 c | 89.26 ± 0.45 | 2.06 ± 0.15 b | 2.09 ± 0.1 ac | 6.59 ± 0.42 cb |
| 1250 | 5116 | 57.43 ± 0.31 d | 88.8 ± 0.34 | 1.87 ± 0.16 ab | 1.9 ± 0.22 c | 7.43 ± 0.48 b |
| 2500 | 5072 | 55.09 ± 0.61 e | 88.84 ± 0.22 | 1.79 ± 0.19 a | 1.93 ± 0.21 ac | 7.44 ± 0.3 b |
| Compound | Receptor | ΔGbest (kcal/mol) | Binding Mode | Classical H Bond | Non-Classical H Bond | π-Sigma Interaction | Halogen (Fluorine) |
|---|---|---|---|---|---|---|---|
| MMS (positive control) | B-DNA | −4.07 | Minor groove | Gua10 (2.12 Å, 2.33 Å), Gua16 (2.28 Å, 2.21 Å) | - | - | Gua10 (5.85 Å), Gua16 (5.45 Å) |
| NaN3 (positive control) | B-DNA | −4.13 | Minor groove | Cyt9 (2.42 Å), Gua10 (2.45 Å), Gua16 (2.14 Å), 2.77 Å) | - | - | - |
| Flubendiamide | B-DNA | −7.25 | Minor groove | Gua16 (2.50 Å, 2.70 Å), Ade17 (2.02 Å) | Cyt11 (3.45 Å, 3.55 Å, 3.70 Å), Gua12 (3.30 Å, 3.44 Å), Ade17 (3.43 Å, 3.46 Å) | Ade18 (3.56 Å) | Cyt11(3.02 Å, 3.59 Å), Cyt15 (3.25 Å) |
| Compound | Molecular Weight (g/mol) | Receptor | ΔGbest (kcal/mol) | Classical H Bond | Non-Classical H Bond | Alkyl, π-Sigma/ π-Alkyl Interaction | Electrostatic Interaction | Halogen (Fluorine) |
|---|---|---|---|---|---|---|---|---|
| Glycerol (native ligand) | 92.09 | RyR Repeat34 domain * | −3.32 | Asp3029 (2.10 Å) | - | - | - | - |
| Flubendiamide | 682.40 | RyR Repeat34 domain | −8.23 | Arg3032 (2.35 Å) | Ala2992 (3.02 Å), Lys2999 (3.11 Å) | Lys3028 (3.49 Å), Ala2992 (4.07 Å), Arg2908 (4.79 Å), Pro2912 (3.95 Å, 4.49 Å), Ile2995 (4.13 Å, 4.97 Å), Pro2912 (5.02 Å) | Lys3028 (4.12 Å), Asp3029 (4.84 Å) | Glu2911 (3.61 Å), Pro2912 (3.22 Å), Ala2992 (2.95 Å) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciğerci, İ.H.; Liman, R.; İstifli, E.S.; Akyıl, D.; Özkara, A.; Bonciu, E.; Colă, F. Cyto-Genotoxic and Behavioral Effects of Flubendiamide in Allium cepa Root Cells, Drosophila melanogaster and Molecular Docking Studies. Int. J. Mol. Sci. 2023, 24, 1565. https://doi.org/10.3390/ijms24021565
Ciğerci İH, Liman R, İstifli ES, Akyıl D, Özkara A, Bonciu E, Colă F. Cyto-Genotoxic and Behavioral Effects of Flubendiamide in Allium cepa Root Cells, Drosophila melanogaster and Molecular Docking Studies. International Journal of Molecular Sciences. 2023; 24(2):1565. https://doi.org/10.3390/ijms24021565
Chicago/Turabian StyleCiğerci, İbrahim Hakkı, Recep Liman, Erman Salih İstifli, Dilek Akyıl, Arzu Özkara, Elena Bonciu, and Florica Colă. 2023. "Cyto-Genotoxic and Behavioral Effects of Flubendiamide in Allium cepa Root Cells, Drosophila melanogaster and Molecular Docking Studies" International Journal of Molecular Sciences 24, no. 2: 1565. https://doi.org/10.3390/ijms24021565
APA StyleCiğerci, İ. H., Liman, R., İstifli, E. S., Akyıl, D., Özkara, A., Bonciu, E., & Colă, F. (2023). Cyto-Genotoxic and Behavioral Effects of Flubendiamide in Allium cepa Root Cells, Drosophila melanogaster and Molecular Docking Studies. International Journal of Molecular Sciences, 24(2), 1565. https://doi.org/10.3390/ijms24021565