
Overexpression of ApHIPP26 from the Hyperaccumulator Arabis paniculata Confers Enhanced Cadmium Tolerance and Accumulation to Arabidopsis thaliana
 and
and 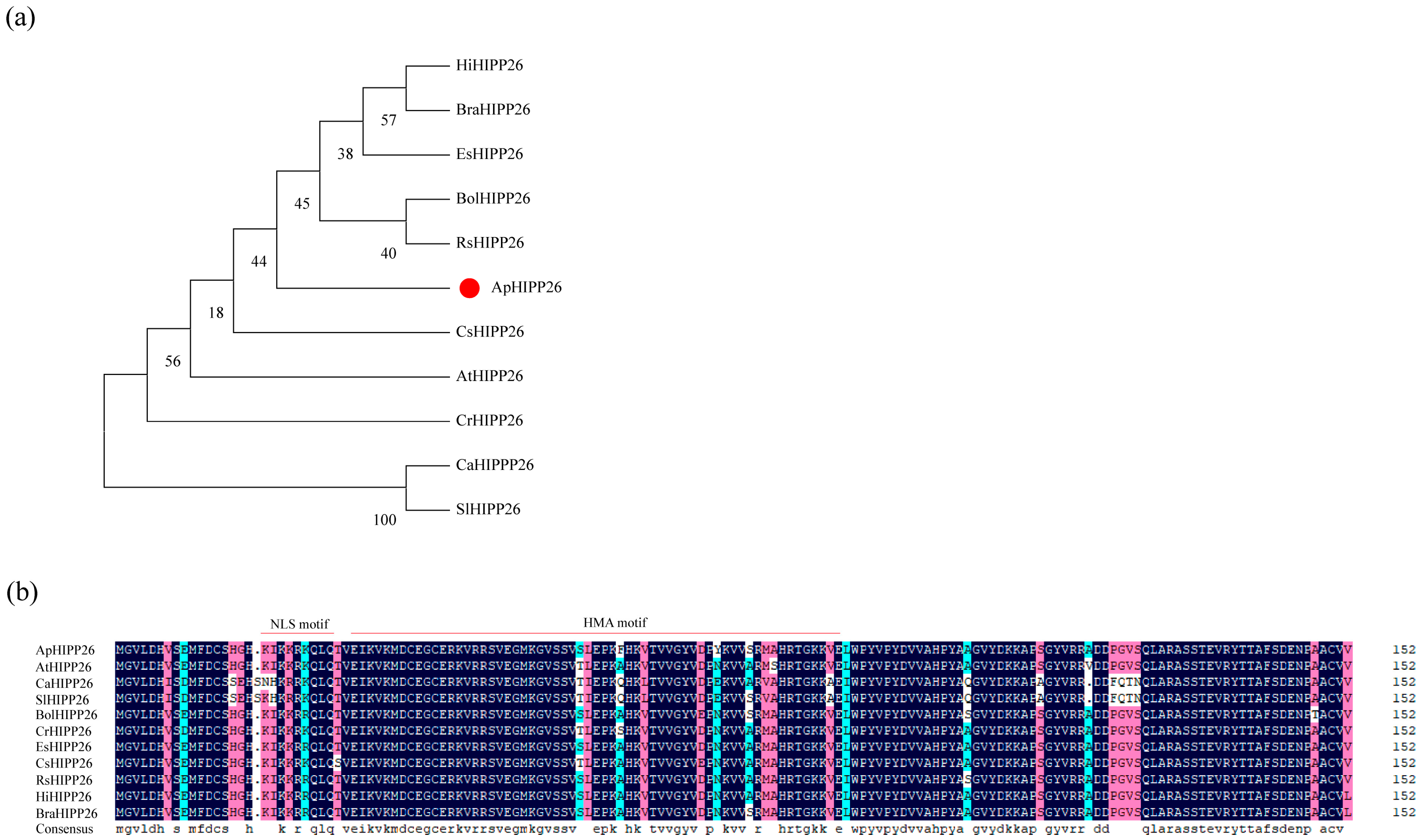{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ApHIPP26 Conserved Motif and Phylogenetic Analysis
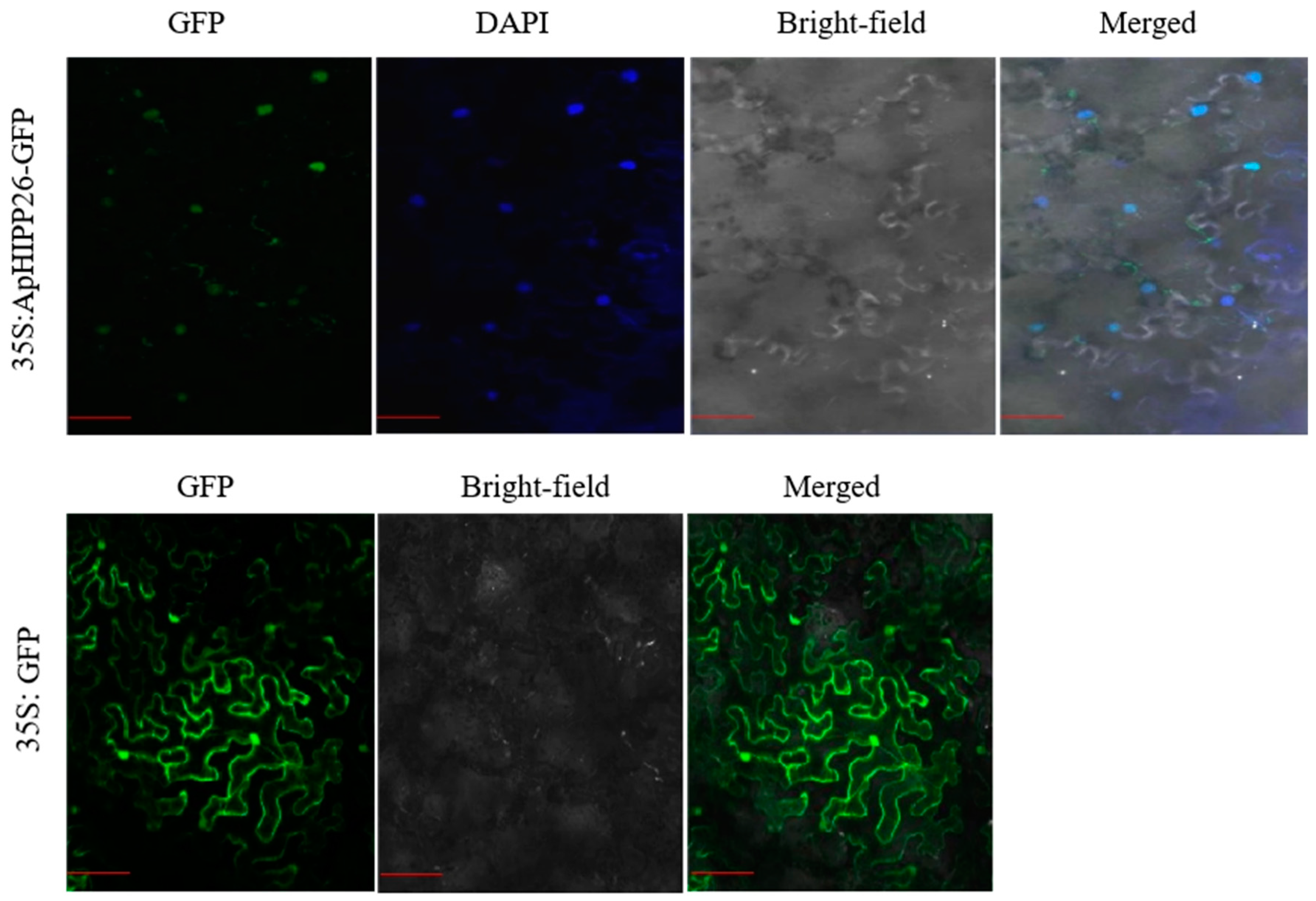
2.2. ApHIPP26 Is Localized in the Nucleus
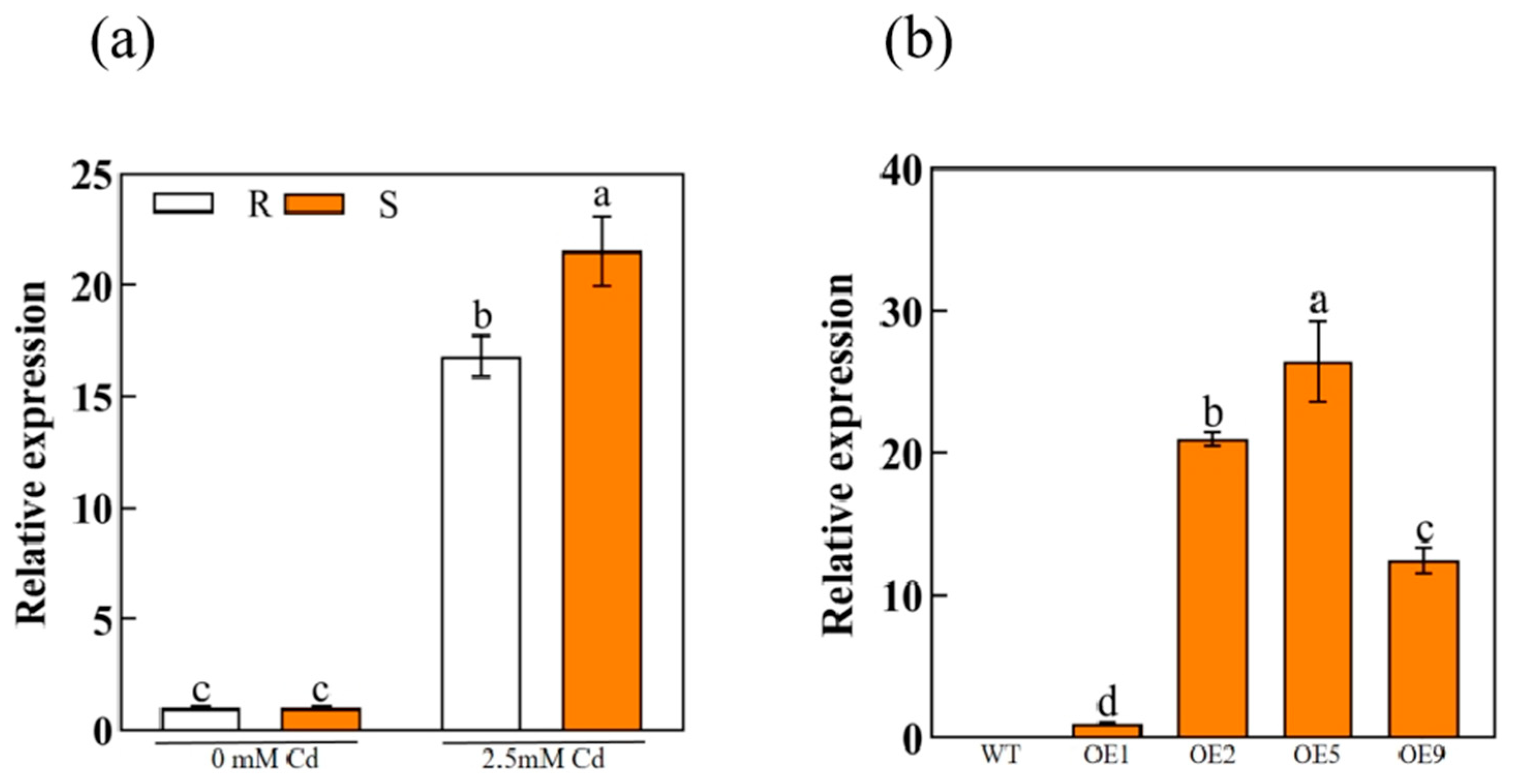
2.3. Cd Induced ApHIPP26 Expression
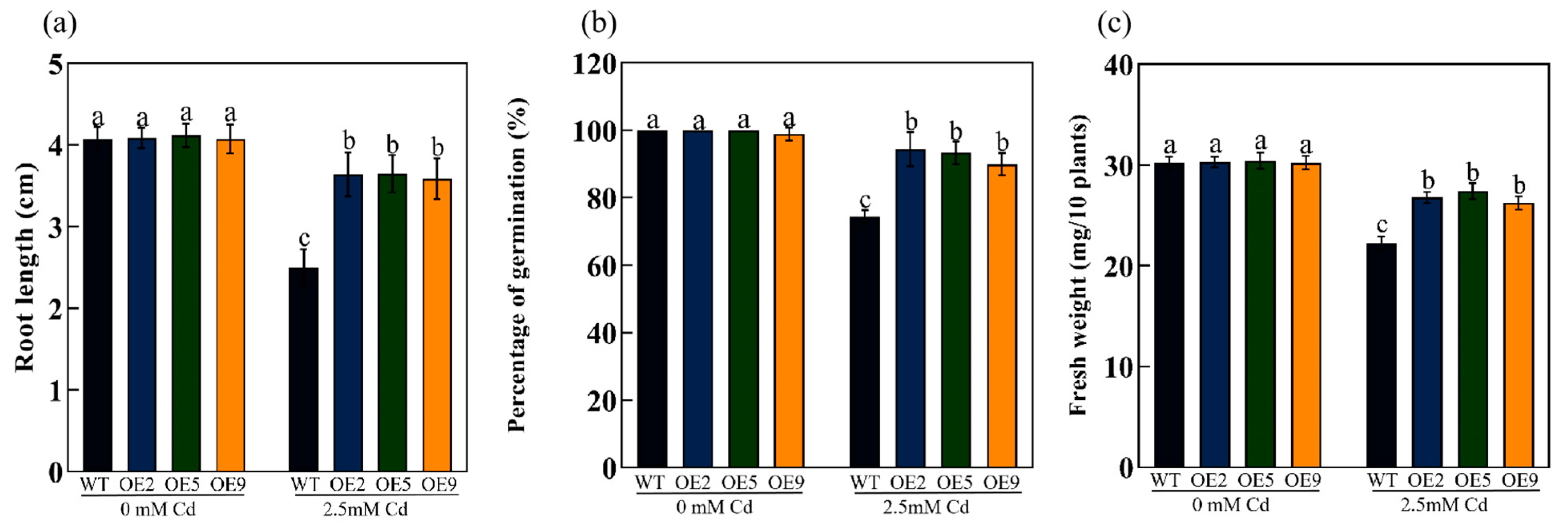
2.4. ApHIPP26 Overexpression Enhanced Cd Tolerance and Accumulation in Transgenic Arabidopsis
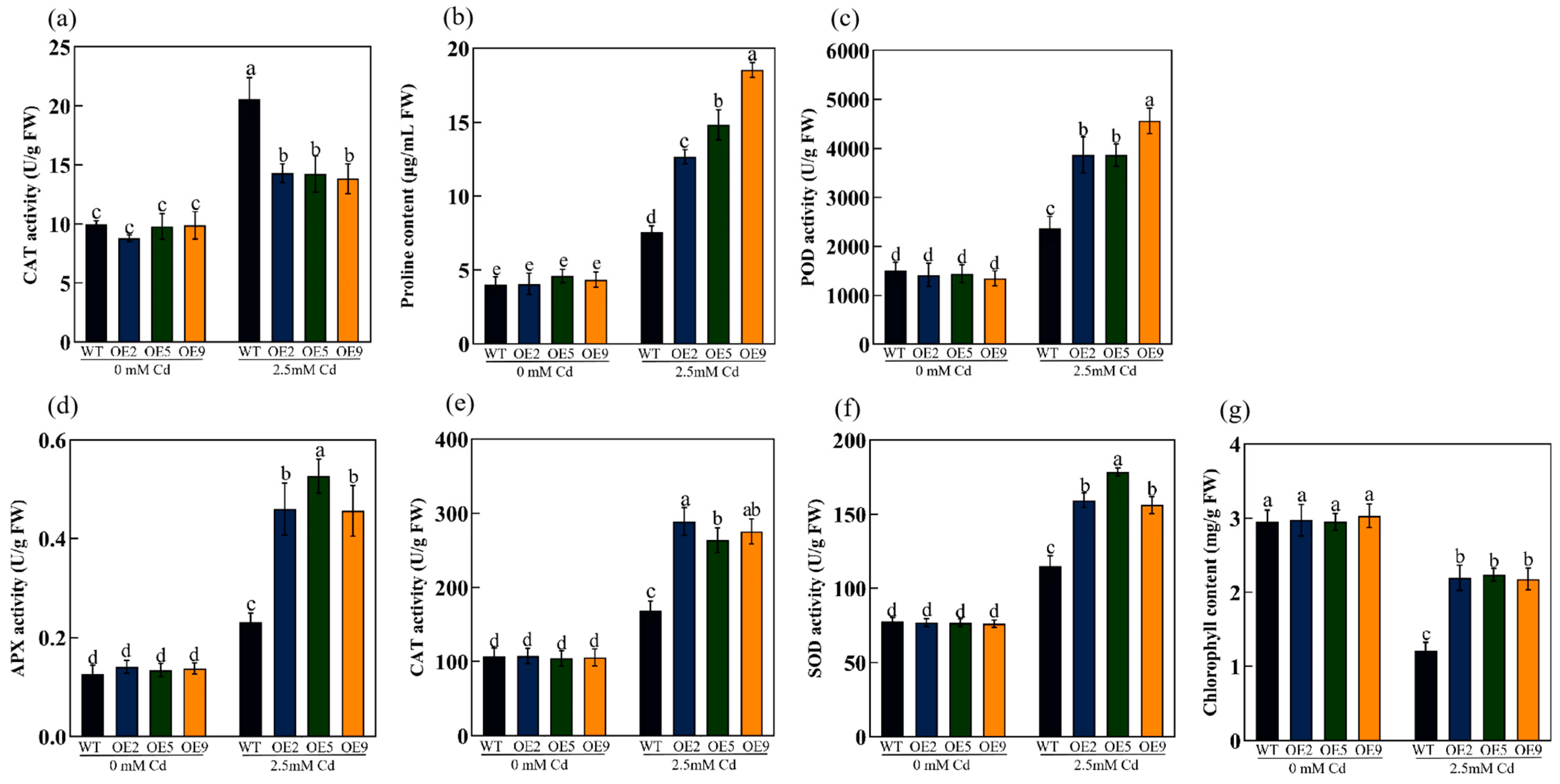
2.5. Overexpression of ApHIPP26 Enhanced the Antioxidant Capacity and Photosynthesis of Transgenic Arabidopsis
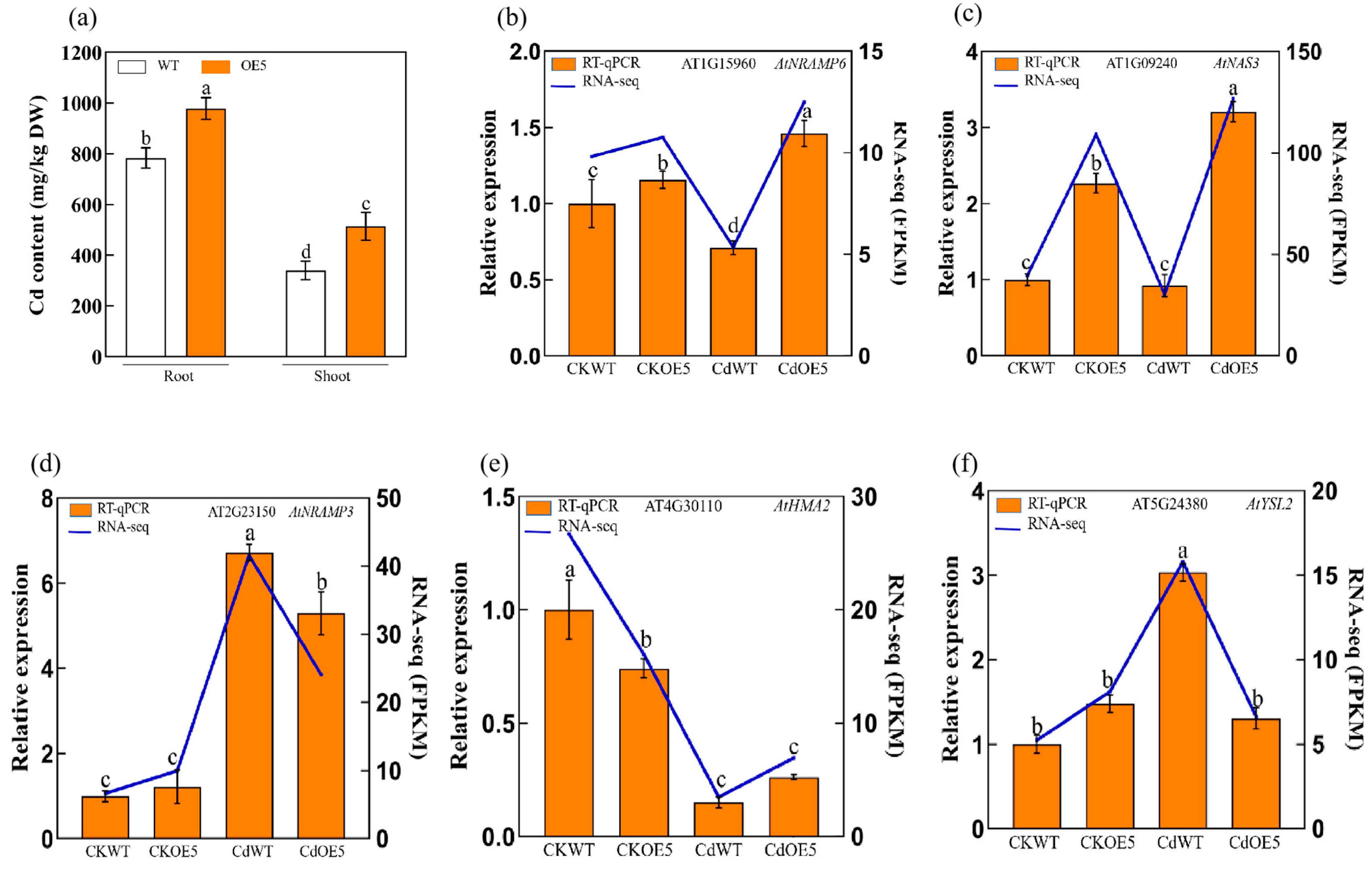
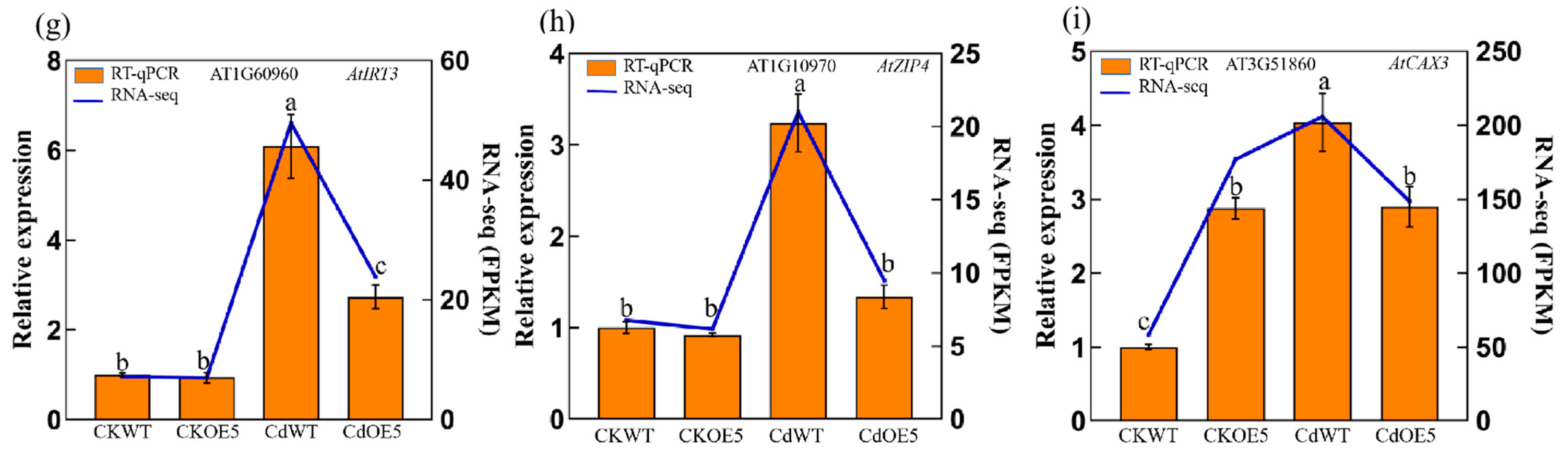
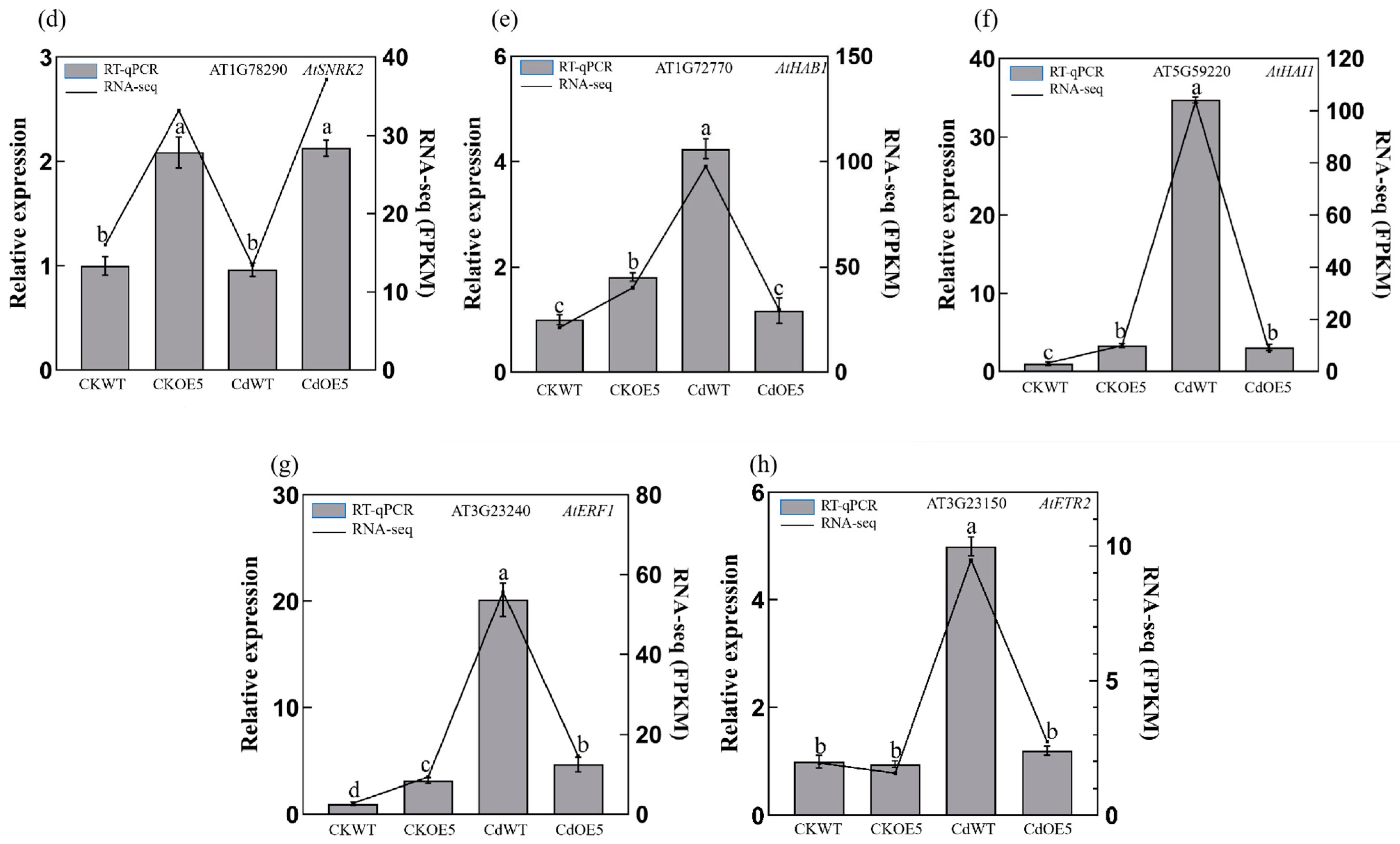
2.6. ApHIPP26 Increased Cd Uptake by Regulating the Expression of Heavy Metal Transporters
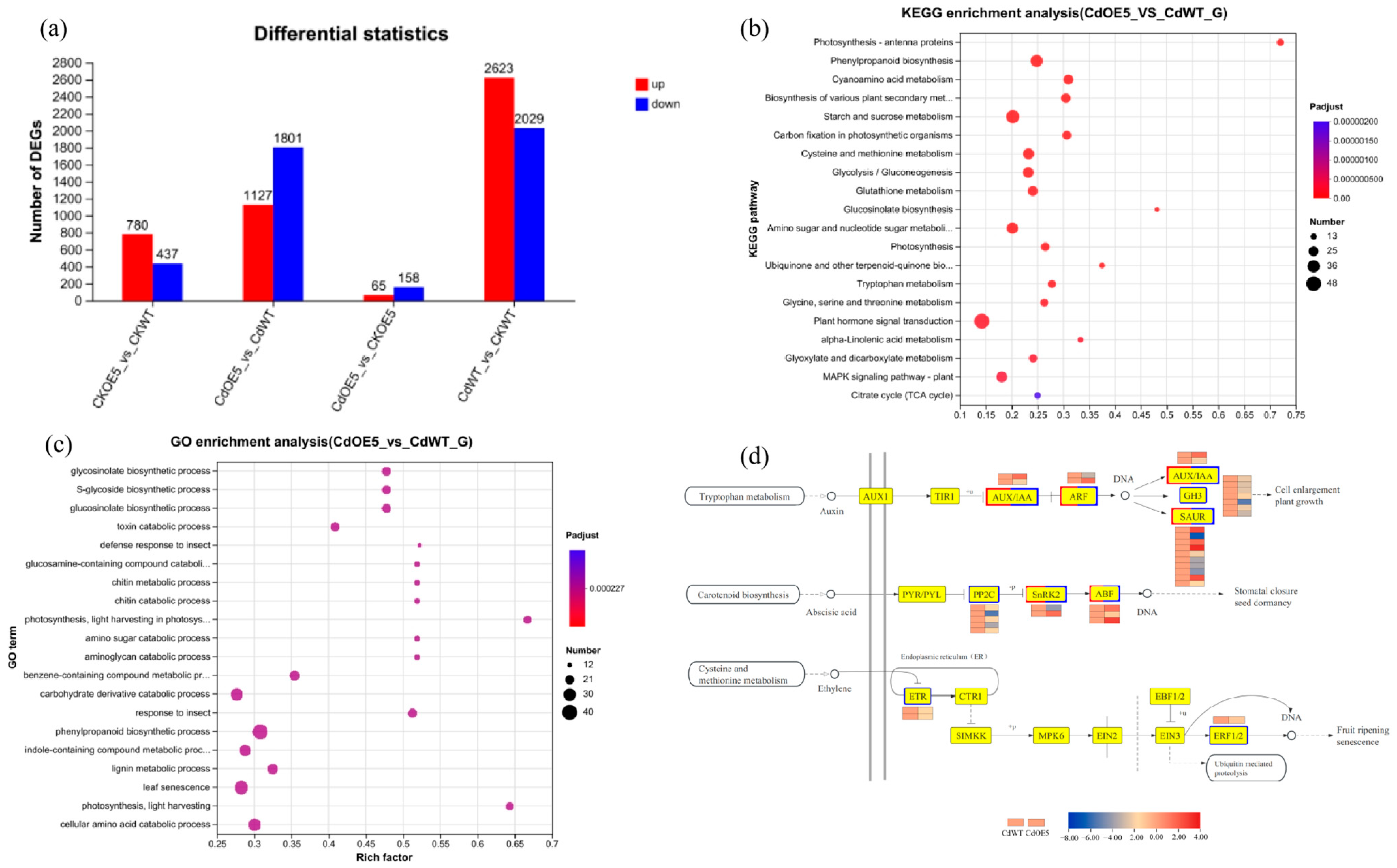
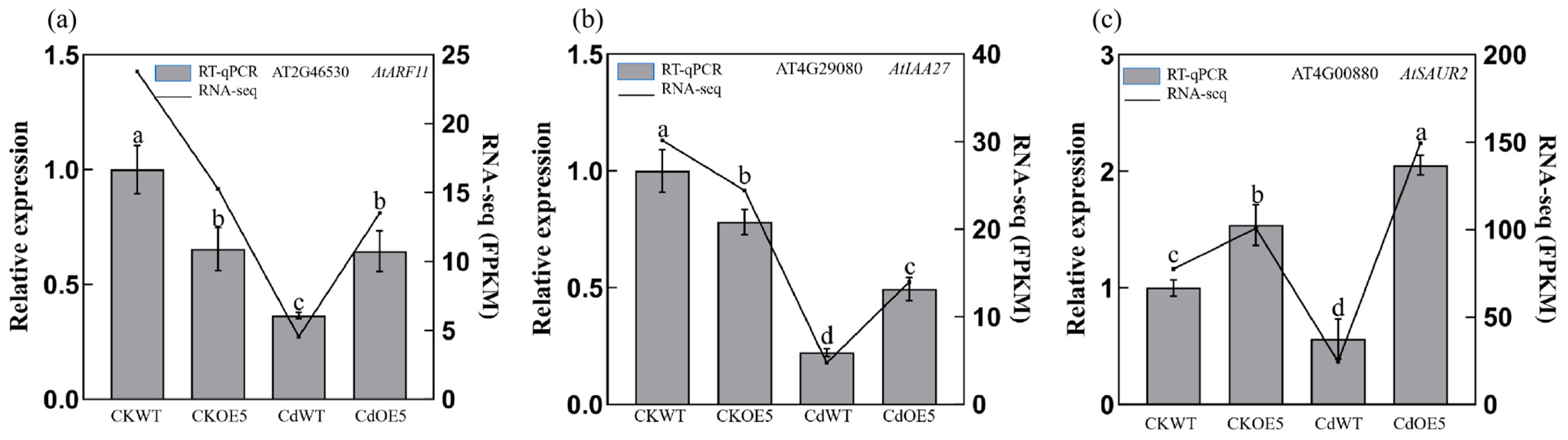
2.7. ApHIPP26 Enhanced the Synthesis of Plant Hormones in the Aerial Parts of Transgenic A. thaliana
3. Discussion
4. Materials and Methods
4.1. Plant Culture and Treatments
4.2. Phylogenetic Analysis
4.3. Subcellular Localization
4.4. RNA Isolation and qRT–PCR
4.5. Construction of the Expression Vector and Genetic Transformation of A. thaliana
4.6. Phenotypic Analysis and Determination of Physiological Indices
4.7. Determination of Cd Content
4.8. RNA-Seq Analysis
4.9. Statistical Analysis
4.10. Primers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- An, J.-P.; Wang, X.-N.; Yao, J.-F.; Ren, Y.-R.; You, C.-X.; Wang, X.-F.; Hao, Y.-J. Apple MdMYC2 Reduces Aluminum Stress Tolerance by Directly Regulating MdERF3 Gene. Plant Soil 2017, 418, 255–266. [Google Scholar] [CrossRef]
- Khoudi, H. Significance of Vacuolar Proton Pumps and Metal/H+ Antiporters in Plant Heavy Metal Tolerance. Physiol. Plant. 2021, 173, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative Roles of Nitric Oxide and Hydrogen Sulfide in Melatonin-induced Tolerance of Pepper (Capsicum annuum L.) Plants to Iron Deficiency and Salt Stress Alone or in Combination. Physiol. Plant. 2019, 168, 256–277. [Google Scholar] [CrossRef]
- Maharajan, T.; Chellasamy, G.; Tp, A.K.; Ceasar, S.A.; Yun, K. The Role of Metal Transporters in Phytoremediation: A Closer Look at Arabidopsis. Chemosphere 2023, 310, 136881. [Google Scholar] [CrossRef]
- Akindele, E.O.; Omisakin, O.D.; Oni, O.A.; Aliu, O.O.; Omoniyi, G.E.; Akinpelu, O.T. Heavy metal toxicity in the water column and benthic sediments of a degraded tropical stream. Ecotoxicol. Environ. Saf. 2020, 190, 110153. [Google Scholar] [CrossRef]
- Ali, B.; Tao, Q.; Zhou, Y.; Gill, R.A.; Ali, S.; Rafiq, M.T.; Xu, L.; Zhou, W. 5-Aminolevolinic Acid Mitigates the Cadmium-Induced Changes in Brassica napus as Revealed by the Biochemical and Ultra-Structural Evaluation of Roots. Ecotoxicol. Environ. Saf. 2013, 92, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Qian, P.; Jin, R.; Ali, S.; Khan, M.; Aziz, R.; Tian, T.; Zhou, W. Physiological and Ultra-Structural Changes in Brassica napus Seedlings Induced by Cadmium Stress. Biol. Plant. 2014, 58, 131–138. [Google Scholar] [CrossRef]
- Ali, B.; Gill, R.A.; Yang, S.; Gill, M.B.; Ali, S.; Rafiq, M.T.; Zhou, W. Hydrogen Sulfide Alleviates Cadmium-Induced Morpho-Physiological and Ultrastructural Changes in Brassica napus. Ecotoxicol. Environ. Saf. 2014, 110, 197–207. [Google Scholar] [CrossRef]
- Zhang, W.; Song, J.; Yue, S.; Duan, K.; Yang, H. MhMAPK4 from Malus Hupehensis Rehd. Decreases Cell Death in Tobacco Roots by Controlling Cd2+ Uptake. Ecotoxicol. Environ. Saf. 2019, 168, 230–240. [Google Scholar] [CrossRef]
- Ankit; Saha, L.; Kumar, V.; Tiwari, J.; Sweta; Rawat, S.; Singh, J.; Bauddh, K. Electronic Waste and Their Leachates Impact on Human Health and Environment: Global Ecological Threat and Management. Environ. Technol. Innov. 2021, 24, 102049. [Google Scholar] [CrossRef]
- Liao, R.; Ratié, G.; Shi, Z.; Šípková, A.; Vaňková, Z.; Chrastný, V.; Zhang, J.; Komárek, M. Cadmium Isotope Systematics for Source Apportionment in an Urban–Rural Region. Appl. Geochem. 2022, 137, 105196. [Google Scholar] [CrossRef]
- Nocito, F.F.; Lancilli, C.; Dendena, B.; Lucchini, G.; Sacchi, G.A. Cadmium Retention in Rice Roots Is Influenced by Cadmium Availability, Chelation and Translocation: Cadmium Retention in Rice Roots. Plant Cell Environ. 2011, 34, 994–1008. [Google Scholar] [CrossRef]
- Yamaji, N.; Sasaki, A.; Xia, J.X.; Yokosho, K.; Ma, J.F. A Node-Based Switch for Preferential Distribution of Manganese in Rice. Nat. Commun. 2013, 4, 2442. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Sang, P.; Guo, Y.; Jin, P.; Cheng, Y.; Yu, H.; Xie, Y.; Yao, W.; Qian, H. Cadmium in Food: Source, Distribution and Removal. Food Chem. 2023, 405, 134666. [Google Scholar] [CrossRef] [PubMed]
- Castaings, L.; Caquot, A.; Loubet, S.; Curie, C. The High-Affinity Metal Transporters NRAMP1 and IRT1 Team up to Take up Iron under Sufficient Metal Provision. Sci. Rep. 2016, 6, 37222. [Google Scholar] [CrossRef] [PubMed]
- Dykema, P.E.; Sipes, P.R.; Marie, A.; Biermann, B.J.; Crowell, D.N.; Randall, S.K. A New Class of Proteins Capable of Binding Transition Metals. Plant Mol. Biol. 1999, 41, 139–150. [Google Scholar] [CrossRef]
- Gao, W.; Xiao, S.; Li, H.; Tsao, S.; Chye, M. Arabidopsis thaliana acyl-CoA-binding Protein ACBP2 Interacts with Heavy-metal-binding Farnesylated Protein AtFP6. New Phytol. 2009, 181, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Parasyri, A.; Barth, O.; Zschiesche, W.; Humbeck, K. The Barley Heavy Metal Associated Isoprenylated Plant Protein HvFP1 Is Involved in a Crosstalk between the Leaf Development and Abscisic Acid-Related Drought Stress Responses. Plants 2022, 11, 2851. [Google Scholar] [CrossRef]
- Wei, Y.; Peng, X.; Wang, X.; Wang, C. The Heavy Metal-Associated Isoprenylated Plant Protein (HIPP) Gene Family Plays a Crucial Role in Cadmium Resistance and Accumulation in the Tea Plant (Camellia sinensis L.). Ecotoxicol. Environ. Saf. 2023, 260, 115077. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Rono, J.K.; Zhang, B.Q.; Liu, X.S.; Wang, M.Q.; Wang, L.L.; Wu, X.C.; Chen, X.; Cao, H.W.; Yang, Z.M. Identification of Novel Rice (Oryza Sativa) HPP and HIPP Genes Tolerant to Heavy Metal Toxicity. Ecotoxicol. Environ. Saf. 2019, 175, 8–18. [Google Scholar] [CrossRef]
- Ye, X.; Liu, C.; Yan, H.; Wan, Y.; Wu, Q.; Wu, X.; Zhao, G.; Zou, L.; Xiang, D. Genome-Wide Identification and Transcriptome Analysis of the Heavy Metal-Associated (HMA) Gene Family in Tartary Buckwheat and Their Regulatory Roles under Cadmium Stress. Gene 2022, 847, 146884. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Yamaguchi, Y.; Koizumi, N.; Sano, H. Functional Characterization of a Heavy Metal Binding Protein CdI19 from Arabidopsis. Plant J. 2002, 32, 165–173. [Google Scholar] [CrossRef]
- Zhao, Y.N.; Wang, M.Q.; Li, C.; Cao, H.W.; Rono, J.K.; Yang, Z.M. The Metallochaperone OsHIPP56 Gene Is Required for Cadmium Detoxification in Rice Crops. Environ. Exp. Bot. 2022, 193, 104680. [Google Scholar] [CrossRef]
- De Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; De Oliveira, L.F.V.; Bodanese Zanettini, M.H.; Margis-Pinheiro, M. Heavy Metal-Associated Isoprenylated Plant Protein (HIPP): Characterization of a Family of Proteins Exclusive to Plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.-S.P.; Xu, J. The R2R3-MYB Transcription Factor MYB49 Regulates Cadmium Accu mulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef]
- Tang, Y.-T.; Qiu, R.-L.; Zeng, X.-W.; Ying, R.-R.; Yu, F.-M.; Zhou, X.-Y. Lead, Zinc, Cadmium Hyperaccumulation and Growth Stimulation in Arabis paniculata Franch. Environ. Exp. Bot. 2009, 66, 126–134. [Google Scholar] [CrossRef]
- Diao, F.; Dang, Z.; Cui, X.; Xu, J.; Jia, B.; Ding, S.; Zhang, Z.; Guo, W. Transcriptomic Analysis Revealed Distinctive Modulations of Arbuscular Mycorrhizal Fungi Inoculation in Halophyte suaeda Salsa under Moderate Salt Conditions. Environ. Exp. Bot. 2021, 183, 104337. [Google Scholar] [CrossRef]
- Renu, K.; Chakraborty, R.; Myakala, H.; Koti, R.; Famurewa, A.C.; Madhyastha, H.; Vellingiri, B.; George, A.; Valsala Gopalakrishnan, A. Molecular Mechanism of Heavy Metals (Lead, Chromium, Arsenic, Mercury, Nickel and Cadmium)—Induced Hepatotoxicity—A Review. Chemosphere 2021, 271, 129735. [Google Scholar] [CrossRef]
- Gerhardt, K.E.; Gerwing, P.D.; Greenberg, B.M. Opinion: Taking Phytoremediation from Proven Technology to Accepted Practice. Plant Sci. 2017, 256, 170–185. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 402647. [Google Scholar] [CrossRef]
- DalCorso, G.; Fasani, E.; Manara, A.; Visioli, G.; Furini, A. Heavy Metal Pollutions: State of the Art and Innovation in Phytore mediation. IJMS 2019, 20, 3412. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of Heavy Metals—Concepts and Applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Sivarajasekar, N.; Baskar, R. Biosorption of Basic Violet 10 onto Activated Gossypium hirsutum Seeds: Batch and Fixed-Bed Column Studies. Chin. J. Chem. Eng. 2015, 23, 1610–1619. [Google Scholar] [CrossRef]
- Parmar, S.; Singh, V. Phytoremediation approaches for heavy metal pollution: A review. J. Plant Sci. Res. 2015, 2, 135–139. [Google Scholar]
- Li, S.; Zhuo, R.; Yu, M.; Lin, X.; Xu, J.; Qiu, W.; Li, H.; Han, X. A Novel Gene SpCTP3 from the Hyperaccumulator Sedum plumbizincicola Redistributes Cadmium and Increases Its Accumulation in Transgenic Populus × Canescens. Front. Plant Sci. 2023, 14, 1111789. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.; Xu, W. Heavy Metal ATPase 3 (HMA3) Confers Cadmium Hypertolerance on the Cadmium/Zinc Hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef]
- Lin, J.; Gao, X.; Zhao, J.; Zhang, J.; Chen, S.; Lu, L. Plant Cadmium Resistance 2 (SaPCR2) Facilitates Cadmium Efflux in the Roots of Hyperaccumulator Sedum alfredii Hance. Front. Plant Sci. 2020, 11, 568887. [Google Scholar] [CrossRef] [PubMed]
- Fasani, E.; DalCorso, G.; Zorzi, G.; Agrimonti, C.; Fragni, R.; Visioli, G.; Furini, A. Overexpression of ZNT1 and NRAMP4 from the Ni Hyperaccumulator Noccaea Caerulescens Population Monte Prinzera in Arabidopsis thaliana Perturbs Fe, Mn, and Ni Accumulation. IJMS 2021, 22, 11896. [Google Scholar] [CrossRef]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and Characterization of a Novel Cadmium-Regu lated Yellow Stripe-Like Transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, C.; Guo, H.; Hu, Y.; He, Y.; Jiang, D. Overexpression of a Miscanthus Sacchariflorus Yellow Stripe-like Trans porter MsYSL1 Enhances Resistance of Arabidopsis to Cadmium by Mediating Metal Ion Reallocation. Plant Growth Regul. 2018, 85, 101–111. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, L.; Gan, C.; Hu, L.; Pang, B.; Zuo, D.; Wang, G.; Wang, H.; Liu, Y. Transcriptomic Analysis Reveals Key Genes and Pathways Corresponding to Cd and Pb in the Hyperaccumulator Arabis paniculata. Ecotoxicol. Environ. Saf. 2023, 254, 114757. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Kandhol, N.; Sharma, S.; Singh, V.P.; Chauhan, D.K.; Ludwig-Müller, J.; Corpas, F.J.; Tripathi, D.K. Auxin Cross talk with Reactive Oxygen and Nitrogen Species in Plant Development and Abiotic Stress. Plant Cell Physiol. 2023, 63, 1814–1825. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, S.; Ali Wani, O.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive Oxygen Species in Plants: From Source to Sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Sedaghat, M.; Tahmasebi-Sarvestani, Z.; Emam, Y.; Mokhtassi-Bidgoli, A. Physiological and Antioxidant Responses of Winter Wheat Cultivars to Strigolactone and Salicylic Acid in Drought. Plant Physiol. Biochem. 2017, 119, 59–69. [Google Scholar] [CrossRef]
- Cui, D.; Yin, Y.; Wang, J.; Wang, Z.; Ding, H.; Ma, R.; Jiao, Z. Research on the Physio-Biochemical Mechanism of Non-Thermal Plasma-Regulated Seed Germination and Early Seedling Development in Arabidopsis. Front. Plant Sci. 2019, 10, 1322. [Google Scholar] [CrossRef]
- Liu, Z.; He, X.; Chen, W.; Yuan, F.; Yan, K.; Tao, D. Accumulation and Tolerance Characteristics of Cadmium in a Potential Hyperaccumulator—Lonicera Japonica Thunb. J. Hazard. Mater. 2009, 169, 170–175. [Google Scholar] [CrossRef]
- Lu, Z.; Chen, S.; Han, X.; Zhang, J.; Qiao, G.; Jiang, Y.; Zhuo, R.; Qiu, W. A Single Amino Acid Change in Nramp6 from Sedum alfredii Hance Affects Cadmium Accumulation. IJMS 2020, 21, 3169. [Google Scholar] [CrossRef]
- Jia, H.; Yin, Z.; Xuan, D.; Lian, W.; Han, D.; Zhu, Z.; Li, C.; Li, C.; Song, Z. Mutation of NtNRAMP3 Improves Cadmium Tolerance and Its Accumulation in Tobacco Leaves by Regulating the Subcellular Distribution of Cadmium. J. Hazard. Mater. 2022, 432, 128701. [Google Scholar] [CrossRef]
- Wang, D.; Repo, E.; He, F.; Zhang, X.; Xiang, H.; Yang, W.; Min, X.; Zhao, F. Dual Functional Sites Strategies toward Enhanced Heavy Metal Remediation: Interlayer Expanded Mg-Al Layered Double Hydroxide by Intercalation with L-Cysteine. J. Hazard. Mater. 2022, 439, 129693. [Google Scholar] [CrossRef]
- Tao, Y.; Wu, Q.; Huang, J.; Fang Shen, R.; Zhu, X.F. The Upstream Regulation of the Root Cell Wall When Arabidopsis Thaliana in Response to Toxic Metal Ions Focusing on Al. Plant Signal. Behav. 2023, 18, 2178085. [Google Scholar] [CrossRef]
- Kumar, S.; Yadav, A.; Verma, R.; Dubey, A.K.; Narayan, S.; Pandey, A.; Sahu, A.; Srivastava, S.; Sanyal, I. Metallothionein (MT1): A Molecular Stress Marker in Chickpea Enhances Drought and Heavy Metal Stress Adaptive Efficacy in Transgenic Plants. Environ. Exp. Bot. 2022, 199, 104871. [Google Scholar] [CrossRef]
- Li, R.; Yang, Y.; Cao, H.; Peng, X.; Yu, Q.; He, L.; Chen, J.; Xiang, L.; Liu, W. Heterologous Expression of the Tobacco Metal lothionein Gene NtMT2F Confers Enhanced Tolerance to Cd Stress in Escherichia Coli and Arabidopsis thaliana. Plant Physiol. Biochem. 2023, 195, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-O.; Gwon, Y.; Kim, J. Exogenous Cysteine Improves Mercury Uptake and Tolerance in Arabidopsis by Regulating the Expression of Heavy Metal Chelators and Antioxidative Enzymes. Front. Plant Sci. 2022, 13, 898247. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Song, T.; Yu, J.; Zhang, W.; Hou, X.; Kong Ling, Z.; Cui, G. Genome-Wide Investigation of the Cysteine Synthase Gene Family Shows That Overexpression of CSase Confers Alkali Tolerance to Alfalfa (Medicago sativa L.). Front. Plant Sci. 2022, 12, 792862. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Li, X.; VandenLangenberg, K.M.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Overexpression of S-Adenosyl- l -Methionine Synthetase Increased Tomato Tolerance to Alkali Stress through Polyamine Metabolism. Plant Biotechnol. J. 2014, 12, 694–708. [Google Scholar] [CrossRef]
- Yang, S.; Wang, J.; Tang, Z.; Li, Y.; Zhang, J.; Guo, F.; Meng, J.; Cui, F.; Li, X.; Wan, S. Calcium/Calmodulin Modulates Salt Responses by Binding a Novel Interacting Protein SAMS1 in Peanut (Arachis hypogaea L.). Crop J. 2023, 11, 21–32. [Google Scholar] [CrossRef]
- Jiang, K.; Asami, T. Chemical Regulators of Plant Hormones and Their Applications in Basic Research and Agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and Their Metabolic Engineering for Abiotic Stress Tolerance in Crop Plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and Molecular Mechanisms of Metal Accumulation in Hyperaccumulator Plants. Physiol. Plant. 2020, 173, 148–166. [Google Scholar] [CrossRef]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between Plant Hormones and Heavy Metals Responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef]
- Huang, T.-L.; Nguyen, Q.T.T.; Fu, S.-F.; Lin, C.-Y.; Chen, Y.-C.; Huang, H.-J. Transcriptomic Changes and Signalling Pathways Induced by Arsenic Stress in Rice Roots. Plant Mol. Biol. 2012, 80, 587–608. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, H.; Fang, Y.; Guo, W.; Chen, H.; Zhang, X.; Dai, W.; Chen, S.; Hao, Q.; Yuan, S.; et al. Overexpression of GmMYB14 Improves High-density Yield and Drought Tolerance of Soybean through Regulating Plant Architecture Mediated by the Brassinosteroid Pathway. Plant Biotechnol. J. 2021, 19, 702–716. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, U.; Khan, S.M.; Khalid, N.; Ahmad, Z.; Jehangir, S.; Fatima Rizvi, Z.; Lho, L.H.; Han, H.; Raposo, A. Detoxifying the Heavy Metals: A Multipronged Study of Tolerance Strategies against Heavy Metals Toxicity in Plants. Front. Plant Sci. 2023, 14, 1154571. [Google Scholar] [CrossRef] [PubMed]
- Guillotin, B.; Etemadi, M.; Audran, C.; Bouzayen, M.; Bécard, G.; Combier, J. Sl-IAA27 Regulates Strigolactone Biosynthesis and Mycorrhization in Tomato (Var. MicroTom). New Phytol. 2017, 213, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Zuo, D.; Hu, M.; Zhou, W.; Lei, F.; Zhao, J.; Gu, L. EcAGL Enhances Cadmium Tolerance in Transgenic Arabidopsis Thaliana through Inhibits Cadmium Transport and Ethylene Synthesis Pathway. Plant Physiol. Biochem. 2023, 201, 107900. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zuo, D.; Zhu, B.; Du, X.; Gu, L. TaMYC8 Regulates TaERF6 and Inhibits Ethylene Synthesis to Confer Cd Tolerance in Wheat. Environ. Exp. Bot. 2022, 198, 104854. [Google Scholar] [CrossRef]
- Ng, L.M.; Melcher, K.; Teh, B.T.; Xu, H.E. Abscisic Acid Perception and Signaling: Structural Mechanisms and Applications. Acta Pharmacol. Sin. 2014, 35, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The Role and Regulation of ABI5 (ABA-Insensitive 5) in Plant Development, Abiotic Stress Responses and Phytohormone Crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef]
- Meng, L.; Yang, Y.; Ma, Z.; Jiang, J.; Zhang, X.; Chen, Z.; Cui, G.; Yin, X. Integrated Physiological, Transcriptomic and Metab olomic Analysis of the Response of Trifolium pratense L. to Pb Toxicity. J. Hazard. Mater. 2022, 436, 129128. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Z.; Song, J.; Yue, S.; Yang, H. Cd2+ Uptake Inhibited by MhNCED3 from Malus Hupehensis Alleviates Cd-Induced Cell Death. Environ. Exp. Bot. 2019, 166, 103802. [Google Scholar] [CrossRef]
- Lu, Q.; Chen, S.; Li, Y.; Zheng, F.; He, B.; Gu, M. Exogenous Abscisic Acid (ABA) Promotes Cadmium (Cd) Accumulation in Sedum Alfredii Hance by Regulating the Expression of Cd Stress Response Genes. Environ. Sci. Pollut. Res. 2020, 27, 8719–8731. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Fang, L.; Li, J.; Chen, T.; Cheng, Z.; Zhu, B.; Gu, L.; Wang, H. The TabHLH094–TaMYC8 Complex Mediates the Cad mium Response in Wheat. Mol. Breed. 2023, 43, 57. [Google Scholar] [CrossRef] [PubMed]
- Kamar, V.; Dağalp, R.; Taştekin, M. Determination of Heavy Metals in Almonds and Mistletoe as a Parasite Growing on the Almond Tree Using ICP-OES or ICP-MS. Biol. Trace Elem. Res. 2018, 185, 226–235. [Google Scholar] [CrossRef]
- Perrino, E.V.; Brunetti, G.; Farrag, K. Plant communities of multi-metal contaminated soils: A case study in National Park of Alta Murgia (Apulia Region—Southern Italy). Int. J. Phytoremediat. 2014, 16, 871–888. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P.; Medagli, P. The genus Aegilops L. (Poaceae) in Italy: Taxonomy, geographical distribution, ecology, vulnerability and conservation. Syst. Biodivers. 2014, 12, 331–349. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Ye, L.; Pang, B.; Hou, Y.; Yu, J.; Du, X.; Gu, L.; Wang, H.; Zhu, B. Overexpression of ApHIPP26 from the Hyperaccumulator Arabis paniculata Confers Enhanced Cadmium Tolerance and Accumulation to Arabidopsis thaliana. Int. J. Mol. Sci. 2023, 24, 15052. https://doi.org/10.3390/ijms242015052
Zhou L, Ye L, Pang B, Hou Y, Yu J, Du X, Gu L, Wang H, Zhu B. Overexpression of ApHIPP26 from the Hyperaccumulator Arabis paniculata Confers Enhanced Cadmium Tolerance and Accumulation to Arabidopsis thaliana. International Journal of Molecular Sciences. 2023; 24(20):15052. https://doi.org/10.3390/ijms242015052
Chicago/Turabian StyleZhou, Lizhou, Lvlan Ye, Biao Pang, Yunyan Hou, Junxing Yu, Xuye Du, Lei Gu, Hongcheng Wang, and Bin Zhu. 2023. "Overexpression of ApHIPP26 from the Hyperaccumulator Arabis paniculata Confers Enhanced Cadmium Tolerance and Accumulation to Arabidopsis thaliana" International Journal of Molecular Sciences 24, no. 20: 15052. https://doi.org/10.3390/ijms242015052
APA StyleZhou, L., Ye, L., Pang, B., Hou, Y., Yu, J., Du, X., Gu, L., Wang, H., & Zhu, B. (2023). Overexpression of ApHIPP26 from the Hyperaccumulator Arabis paniculata Confers Enhanced Cadmium Tolerance and Accumulation to Arabidopsis thaliana. International Journal of Molecular Sciences, 24(20), 15052. https://doi.org/10.3390/ijms242015052