
TMT and PRM Based Quantitative Proteomics to Explore the Protective Role and Mechanism of Iristectorin B in Stroke

Abstract
:1. Introduction
2. Results
2.1. Cell Experimental Results
2.1.1. Morphological Changes in PC12 Cells
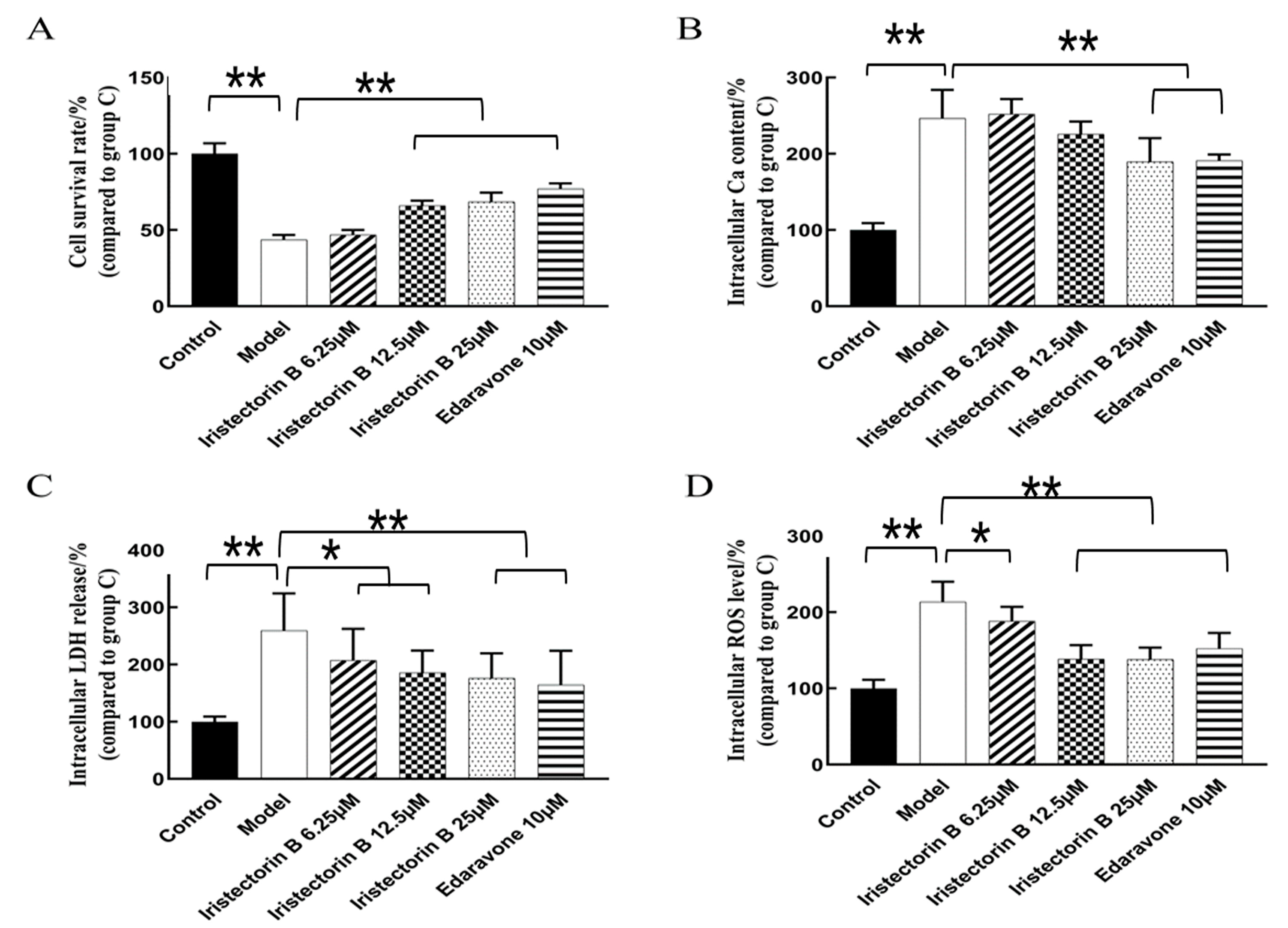
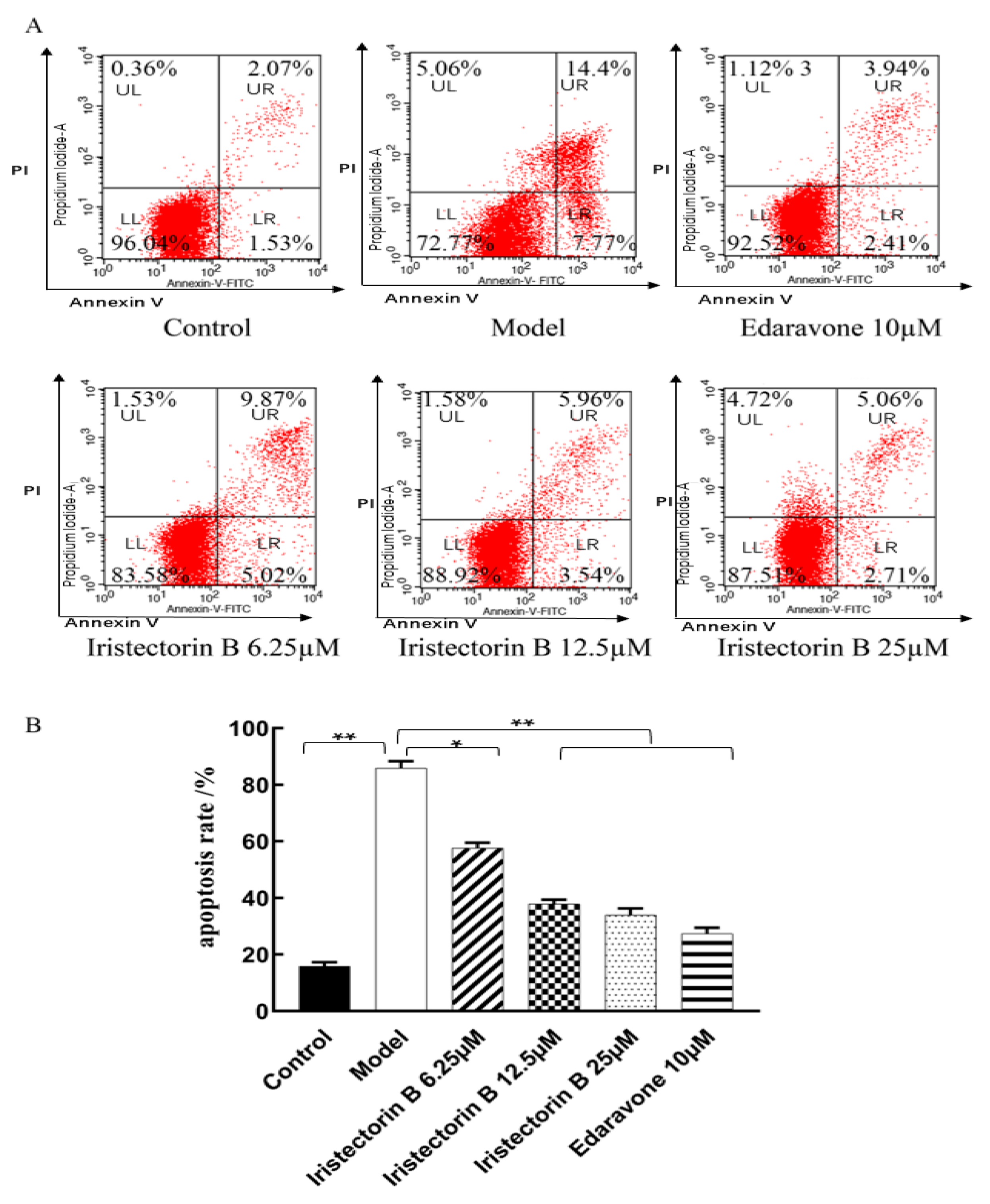
2.1.2. Effect of the Treatment of the PC12 Cells with Increasing Concentrations of Iristectorin B
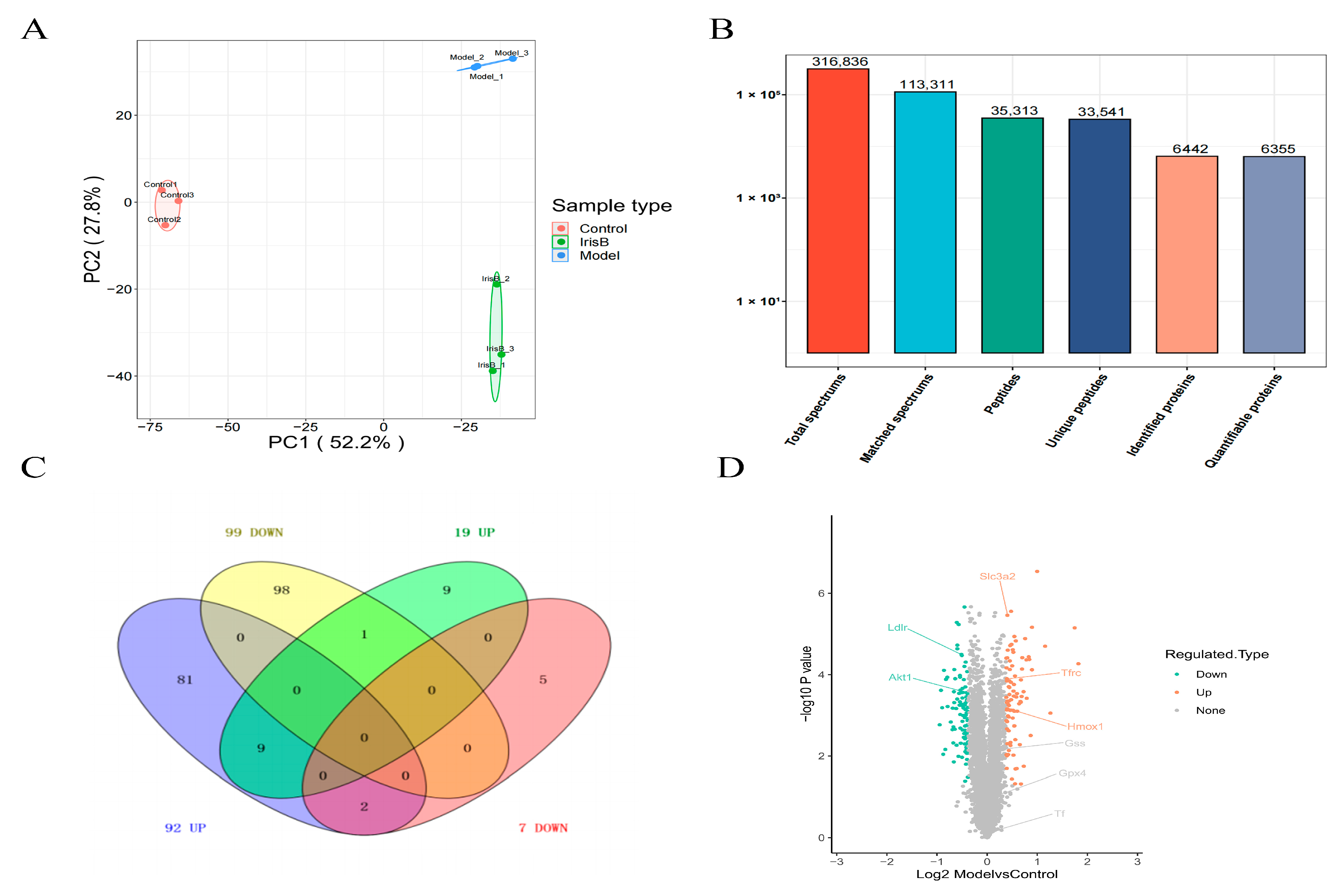
2.2. General Overview of Protein Identification
2.3. GO and KEGG Analysis of Differentially Expressed Proteins
2.4. Bioinformatics Analysis
2.4.1. Subcellular Structures
2.4.2. COG/KOG and Cluster Enrichment Analysis
2.5. PPI Analysis
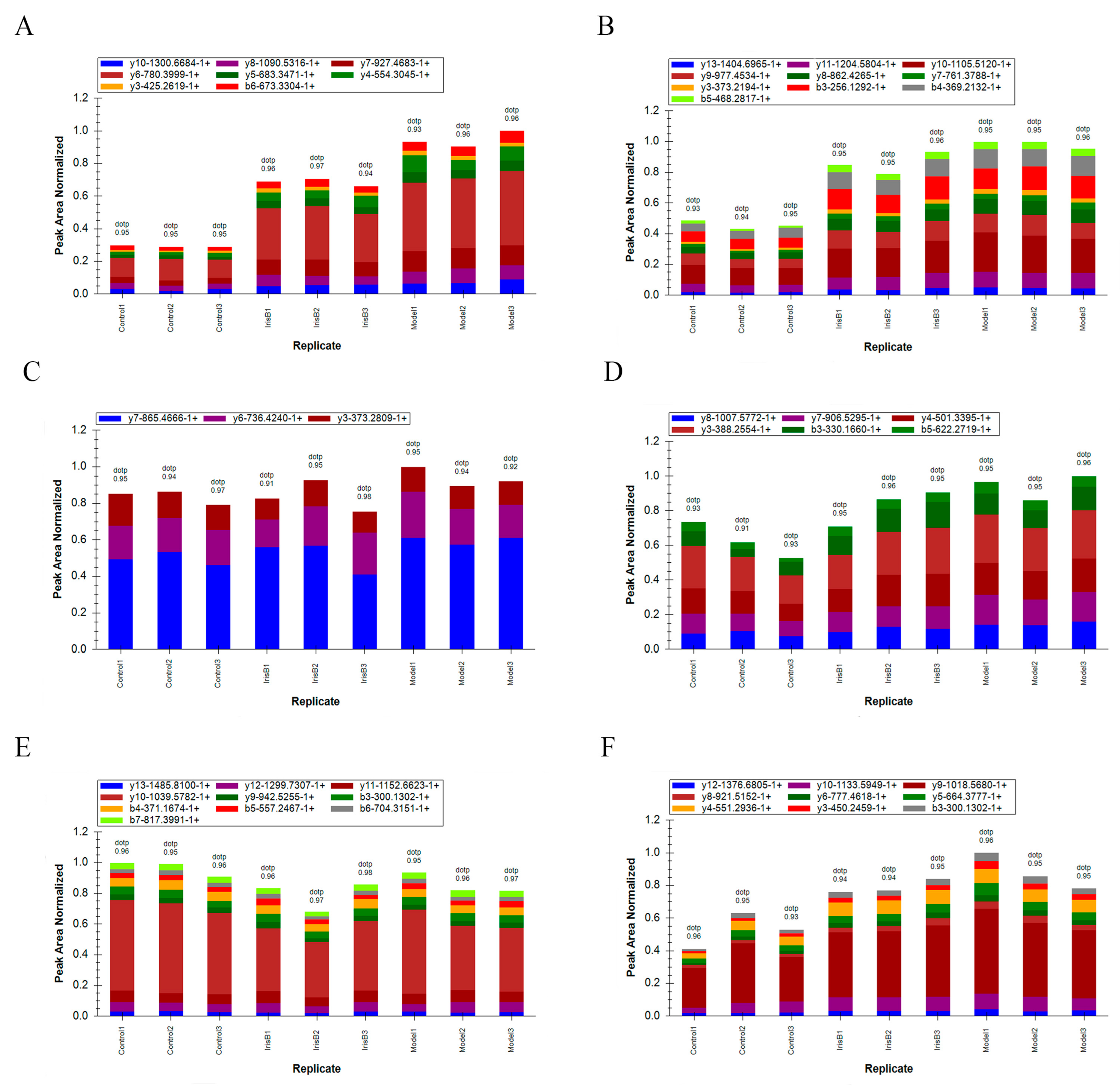
2.6. PRM Validation of Differential Proteins
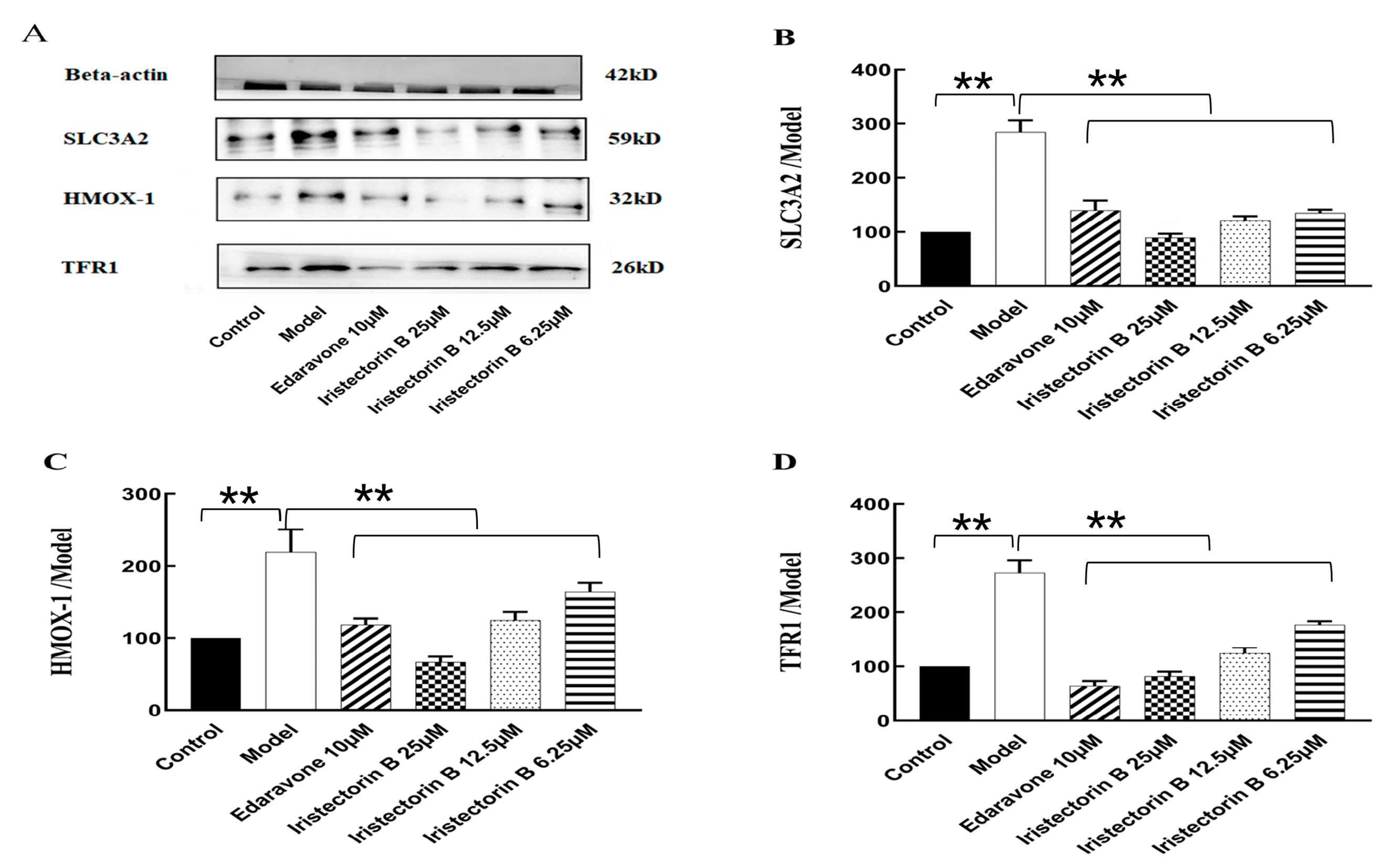
2.7. Western Blot Analysis
3. Discussion
4. Materials and Methods
4.1. PC12 Cell Culture
4.2. Establishment and Experimental Grouping of a Glucose Oxygen Deprivation/Reoxygenation (OGD/R) Ex Vivo Stroke Model
4.2.1. Morphological Observation
4.2.2. MTT Method to Measure Cellular Activity
4.2.3. Detection of Ca2+ Content in PC12 Cells
4.2.4. LDH Release Assay in PC12 Cells
4.2.5. Detection of ROS Levels in PC12 Cells
4.2.6. Apoptosis Assay
4.3. Proteomics Steps
4.3.1. Protein Extraction
4.3.2. Trypsin Digestion
4.3.3. Quantitative Protein Mass Spectrometry TMT Labeling
4.3.4. Classification
4.3.5. Liquid Chromatography-Mass Spectrometry (LC-MS/MS) Analysis
4.3.6. Proteomics Data Analysis
4.3.7. Bioinformatics Analysis
4.3.8. PRM Validation
4.3.9. Western Blot Analysis
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barthels, D.; Das, H. Current advances in ischemic stroke research and therapies. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165260. [Google Scholar] [CrossRef]
- Cordonnier, C.; Sprigg, N.; Sandset, E.C.; Pavlovic, A.; Sunnerhagen, K.S.; Caso, V.; Christensen, H.; Women Initiative for Stroke in Europe (WISE) Group. Stroke in women–from evidence to inequalities. Nat. Rev. Neurol. 2017, 13, 521–532. [Google Scholar] [CrossRef]
- Xu, Y.; Li, K.; Zhao, Y.; Zhou, L.; Liu, Y.; Zhao, J. Role of Ferroptosis in Stroke. Cell. Mol. Neurobiol. 2023, 43, 205–222. [Google Scholar] [CrossRef]
- Yuan, H.; Pratte, J.; Giardina, C. Ferroptosis and its potential as a therapeutic target. Biochem. Pharmacol. 2021, 186, 114486. [Google Scholar] [CrossRef]
- Zhou, Y.; Liao, J.; Mei, Z.; Liu, X.; Ge, J. Insight into Crosstalk between Ferroptosis and Necroptosis: Novel Therapeutics in Ischemic Stroke. Oxidative Med. Cell. Longev. 2021, 2021, 9991001. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef]
- Weiland, A.; Wang, Y.; Wu, W.; Lan, X.; Han, X.; Li, Q.; Wang, J. Ferroptosis and Its Role in Diverse Brain Diseases. Mol. Neurobiol. 2019, 56, 4880–4893. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.-L.; Ding, S.-Y.; Du, X.-Z.; Wang, J.-H.; Li, X.-L. Ferroptosis—A Novel Mechanism with Multifaceted Actions on Stroke. Front. Neurol. 2022, 13, 881809. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Chen, X.; Tan, Q.; Zhou, H.; Xu, J.; Gu, Q. Inhibiting Ferroptosis through Disrupting the NCOA4–FTH1 Interaction: A New Mechanism of Action. ACS Cent. Sci. 2021, 7, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhuang, Y.; Liu, M.; Che, J.; Dong, X. Inhibiting ferroptosis: A novel approach for stroke therapeutics. Drug Discov. Today 2021, 26, 916–930. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fan, H.; Wang, S.; Tang, G.; Zhai, C.; Shen, L. Ferroptosis: A Novel Therapeutic Target for Ischemia-Reperfusion Injury. Front. Cell Dev. Biol. 2021, 9, 688605. [Google Scholar] [CrossRef]
- Wu, X.; Li, Y.; Zhang, S.; Zhou, X. Ferroptosis as a novel therapeutic target for cardiovascular disease. Theranostics 2021, 11, 3052–3059. [Google Scholar] [CrossRef]
- Letunica, N.; Van Den Helm, S.; McCafferty, C.; Swaney, E.; Cai, T.; Attard, C.; Karlaftis, V.; Monagle, P.; Ignjatovic, V. Proteomics in Thrombosis and Hemostasis. Thromb. Haemost. 2022, 122, 1076–1084. [Google Scholar] [CrossRef]
- Kushner, J.S.; Liu, G.; Eisert, R.J.; Bradshaw, G.A.; Pitt, G.S.; Hinson, J.T.; Kalocsay, M.; Marx, S.O. Detecting Cardiovascular Protein-Protein Interactions by Proximity Proteomics. Circ. Res. 2022, 130, 273–287. [Google Scholar] [CrossRef]
- Hou, J.; Deng, Q.; Liu, S.; Qiu, X.; Deng, X.; Zhong, W.; Zhong, Z. Plasma Proteome Profiling of Patients With In-stent Restenosis by Tandem Mass Tag-Based Quantitative Proteomics Approach. Front. Cardiovasc. Med. 2022, 9, 793405. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Q.; Ge, Q.Q.; Lu, S.H.; Yang, M.S.; Zhuang, Y.; Zhang, B.; Niu, F.; Xu, X.J.; Liu, B.Y. Tandem Mass Tag-based proteomics analysis reveals the vital role of inflammation in traumatic brain injury in a mouse model. Neural Regen. Res. 2023, 18, 155–161. [Google Scholar] [CrossRef]
- Cao, H.; Zhu, X.; Chen, X.; Yang, Y.; Zhou, Q.; Xu, W.; Wang, D. Quantitative proteomic analysis to identify differentially expressed proteins in the persistent atrial fibrillation using TMT coupled with nano-LC-MS/MS. Am. J. Transl. Res. 2020, 12, 5032–5047. [Google Scholar]
- Kehm, R.; Baldensperger, T.; Raupbach, J.; Höhn, A. Protein oxidation—Formation mechanisms, detection and relevance as biomarkers in human diseases. Redox Biol. 2021, 42, 101901. [Google Scholar] [CrossRef]
- Zwirner, J.; Bohnert, S.; Franke, H.; Garland, J.; Hammer, N.; Möbius, D.; Tse, R.; Ondruschka, B. Assessing Protein Biomarkers to Detect Lethal Acute Traumatic Brain Injuries in Cerebrospinal Fluid. Biomolecules 2021, 11, 1577. [Google Scholar] [CrossRef]
- Yu, K.; Wang, Z.; Wu, Z.; Tan, H.; Mishra, A.; Peng, J. High-Throughput Profiling of Proteome and Posttranslational Modifications by 16-Plex TMT Labeling and Mass Spectrometry. Methods Mol. Biol. 2021, 2228, 205–224. [Google Scholar] [CrossRef]
- Andersen, L.-A.C.; Palstrøm, N.B.; Diederichsen, A.; Lindholt, J.S.; Rasmussen, L.M.; Beck, H.C. Determining Plasma Protein Variation Parameters as a Prerequisite for Biomarker Studies-A TMT-Based LC-MSMS Proteome Investigation. Proteomes 2021, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yang, S.; Kavdia, K.; Sifford, J.M.; Wu, Z.; Xie, B.; Wang, Z.; Pagala, V.R.; Wang, H.; Yu, K.; et al. Deep Profiling of Microgram-Scale Proteome by Tandem Mass Tag Mass Spectrometry. J. Proteome Res. 2021, 20, 337–345. [Google Scholar] [CrossRef]
- Gong, Q.; Zhang, X.; Liang, A.; Huang, S.; Tian, G.; Yuan, M.; Ke, Q.; Cai, Y.; Yan, B.; Wang, J.; et al. Proteomic screening of potential N-glycoprotein biomarkers for colorectal cancer by TMT labeling combined with LC-MS/MS. Clin. Chim. Acta. 2021, 521, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Feng, S.; Wang, W.; Zhao, F.; Gong, Y. Biomarker and Drug Target Discovery Using Quantitative Proteomics Post-Intracerebral Hemorrhage Stroke in the Rat Brain. J. Mol. Neurosci. 2018, 66, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Cikic, S.; Chandra, P.K.; Harman, J.C.; Rutkai, I.; Katakam, P.V.; Guidry, J.J.; Gidday, J.M.; Busija, D.W. Sexual differences in mitochondrial and related proteins in rat cerebral microvessels: A proteomic approach. J. Cereb. Blood Flow Metab. 2021, 41, 397–412. [Google Scholar] [CrossRef]
- Mai, L.; He, G.; Chen, J.; Zhu, J.; Chen, S.; Hou, X.; Yang, H.; Zhang, M.; Wu, Y.; Lin, Q.; et al. Proteomic Analysis of Hypoxia-Induced Senescence of Human Bone Marrow Mesenchymal Stem Cells. Stem Cells Int. 2021, 2021, 5555590. [Google Scholar] [CrossRef]
- Chai, Y.-N.; Qin, J.; Li, Y.-L.; Tong, Y.-L.; Liu, G.-H.; Wang, X.-R.; Liu, C.-Y.; Peng, M.-H.; Qin, C.-Z.; Xing, Y.-R. TMT proteomics analysis of intestinal tissue from patients of irritable bowel syndrome with diarrhea: Implications for multiple nutrient ingestion abnormality. J. Proteom. 2021, 231, 103995. [Google Scholar] [CrossRef]
- Sun, X.; Li, Q.; Wang, J.; Ma, Y.; Wang, M.; Qin, W. Urinary Proteome Analysis of Global Cerebral Ischemia-Reperfusion Injury Rat Model via Data-Independent Acquisition and Parallel Reaction Monitoring Proteomics. J. Mol. Neurosci. 2022, 72, 2020–2029. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Y.; Bian, L.; Zhang, Y.; Zhang, Y.; Wang, C.; Tian, J.; Lu, T. The crosstalk signals of Sodium Tanshinone ⅡA Sulfonate in rats with cerebral ischemic stroke: Insights from proteomics. Biomed. Pharmacother. 2022, 151, 113059. [Google Scholar] [CrossRef]
- Azurmendi, L.; Lapierre-Fetaud, V.; Schneider, J.; Montaner, J.; Katan, M.; Sanchez, J.-C. Proteomic discovery and verification of serum amyloid A as a predictor marker of patients at risk of post-stroke infection: A pilot study. Clin. Proteom. 2017, 14, 27. [Google Scholar] [CrossRef]
- Montalván Ayala, V.; Rojas Cheje, Z.; Aldave Salazar, R. Controversies in cerebrovascular disease: High or low doses of recombinant tissue plasminogen activator to treat acute stroke? A literature review. Neurologia 2022, 37, 130–135. [Google Scholar] [CrossRef]
- Yu, Y.; Yan, Y.; Niu, F.; Wang, Y.; Chen, X.; Su, G.; Liu, Y.; Zhao, X.; Qian, L.; Liu, P.; et al. Ferroptosis: A cell death connecting oxidative stress, inflammation and cardiovascular diseases. Cell Death Discov. 2021, 7, 193. [Google Scholar] [CrossRef]
- Davidson, S.M.; Adameová, A.; Barile, L.; Cabrera-Fuentes, H.A.; Lazou, A.; Pagliaro, P.; Stensløkken, K.O.; Garcia-Dorado, D. EU-CARDIOPROTECTION COST Action (CA16225) Mitochondrial and mitochondrial-independent pathways of myocardial cell death during ischaemia and reperfusion injury. J. Cell. Mol. Med. 2020, 24, 3795–3806. [Google Scholar] [CrossRef]
- Yan, H.-F.; Tuo, Q.-Z.; Yin, Q.-Z.; Lei, P. The pathological role of ferroptosis in ischemia/reperfusion-related injury. Zool. Res. 2020, 41, 220–230. [Google Scholar] [CrossRef]
- Tang, L.-J.; Zhou, Y.-J.; Xiong, X.-M.; Li, N.-S.; Zhang, J.-J.; Luo, X.-J.; Peng, J. Ubiquitin-specific protease 7 promotes ferroptosis via activation of the p53/TfR1 pathway in the rat hearts after ischemia/reperfusion. Free Radic. Biol. Med. 2021, 162, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Illés, E.; Patra, S.G.; Marks, V.; Mizrahi, A.; Meyerstein, D. The FeII(citrate) Fenton reaction under physiological conditions. J. Inorg. Biochem. 2020, 206, 111018. [Google Scholar] [CrossRef]
- Liu, C.X.; Guo, X.Y.; Zhang, L.L.; Chang, X. Research progress in ferroptosis and ischemic stroke and its medical intervention. J. Pract. Cardio-Cerebrovasc. Dis. 2022, 30, 135–140. [Google Scholar]
- Shi, X.H.; Mang, J.; Xu, Z.X. Research progress on cell death model of cerebral ischemia reperfusion injury. J. Jilin Univ. 2022, 48, 1635–1643. [Google Scholar]
- Kakhlon, O.; Cabantchik, Z.I. The labile iron pool: Characterization, measurement, and participation in cellular processes. Free Radic Biol. Med. 2002, 33, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Li, J.; Zhang, Y.; Chang, Y.-Z. Cellular Iron Metabolism and Regulation. Adv. Exp. Med. Biol. 2019, 1173, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Arosio, P.; Elia, L.; Poli, M. Ferritin, cellular iron storage and regulation. IUBMB Life 2017, 69, 414–422. [Google Scholar] [CrossRef]
- Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Regulators of Iron Homeostasis: New Players in Metabolism, Cell Death, and Disease. Trends Biochem. Sci. 2016, 41, 274–286. [Google Scholar] [CrossRef]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Redox functions of heme oxygenase-1 and biliverdin reductase in diabetes. Trends Endocrinol. Metab. 2018, 29, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.-C.; Chiang, S.-K.; Chen, S.-E.; Yu, Y.-L.; Chou, R.-H.; Chang, W.-C. Heme oxygenase-1 mediates BAY 11–7085 induced ferroptosis. Cancer Lett. 2018, 416, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Komatsu, M.; Yamaguchi-Iwai, Y.; Ishikawa, F.; Mizushima, N.; Iwai, K. Distinct mechanisms of ferritin delivery to lysosomes in iron-depleted and iron-replete cells. Mol. Cell. Biol. 2011, 31, 2040–2052. [Google Scholar] [CrossRef]
- Bauckman, K.A.; Mysorekar, I.U. Ferritinophagy drives uropathogenic Escherichia coli persistence in bladder epithelial cells. Autophagy 2016, 12, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Bellelli, R.; Federico, G.; Matte’, A.; Colecchia, D.; Iolascon, A.; Chiariello, M.; Santoro, M.; De Franceschi, L.; Carlomagno, F. NCOA4 Deficiency Impairs Systemic Iron Homeostasis. Cell Rep. 2016, 14, 411–421. [Google Scholar] [CrossRef]
- Zhou, B.; Liu, J.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Tang, D. Ferroptosis is a type of autophagy-dependent cell death. Semin. Cancer Biol. 2020, 66, 89–100. [Google Scholar] [CrossRef]
- Sato, H.; Tamba, M.; Ishii, T.; Bannai, S. Cloning and expression of a plasma membrane cystine/glutamate exchange transporter composed of two distinct proteins. J. Biol. Chem. 1999, 274, 11455–11458. [Google Scholar] [CrossRef]
- Tang, D.; Chen, X.; Kang, R.; Kroemer, G. Ferroptosis: Molecular mechanisms and health implications. Cell Res. 2021, 31, 107–125. [Google Scholar] [CrossRef]
- Lin, C.-H.; Lin, P.-P.; Lin, C.-Y.; Lin, C.-H.; Huang, C.-H.; Huang, Y.-J.; Lane, H.-Y. Decreased mRNA expression for the two subunits of system Xc−, SLC3A2 and SLC7A11, in WBC in patients with schizophrenia: Evidence in support of the hypo-glutamatergic hypothesis of schizophrenia. J. Psychiatr. Res. 2016, 72, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Forcina, G.C.; Dixon, S.J. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics 2019, 19, e1800311. [Google Scholar] [CrossRef] [PubMed]
- Ursini, F.; Maiorino, M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef]
- Li, C.; Sun, G.; Chen, B.; Xu, L.; Ye, Y.; He, J.; Bao, Z.; Zhao, P.; Miao, Z.; Zhao, L.; et al. Nuclear receptor coactivator 4-mediated ferritinophagy contributes to cerebral ischemia-induced ferroptosis in ischemic stroke. Pharmacol. Res. 2021, 174, 105933. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Cao, X.; Zhang, X.; Wang, Y.; Huang, H.; Meng, Y.; Chang, J. Brozopine Inhibits 15-LOX-2 Metabolism Pathway After Transient Focal Cerebral Ischemia in Rats and OGD/R-Induced Hypoxia Injury in PC12 Cells. Front. Pharmacol. 2020, 11, 99. [Google Scholar] [CrossRef]
- Shi, Y.; Ding, Y.; Li, G.; Wang, L.; Osman, R.A.; Sun, J.; Qian, L.; Zheng, G.; Zhang, G. Discovery of Novel Biomarkers for Diagnosing and Predicting the Progression of Multiple Sclerosis Using TMT-Based Quantitative Proteomics. Front. Immunol. 2021, 12, 700031. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Post-Translational Modifications | Signaling Function Proteins | Metabolism-Related Proteins | Information Storage | Unknown Function Proteins |
|---|---|---|---|---|---|
| model/control | 26 | 71 | 31 | 36 | 24 |
| experimental/control | 3 | 8 | 2 | 12 | 2 |
| BP | Model/Control p < 0.0005 | BP | Experimental/Control p < 0.01 |
|---|---|---|---|
| negatively regulated for programmed cell death | 17–24 | peptide biosynthesis process | 6–8 |
| negatively regulated related proteins for protein metabolic processes | 17–24 | amide biosynthesis process | 6–8 |
| related proteins | 10–17 | peptide metabolism process | 6–8 |
| cell death | 17–24 | ribosome synthesis ranged | 4–6 |
| CC | Model/Control p < 0.0001 | CC | Experimental/Control p < 0.05 |
|---|---|---|---|
| cellular subunit-associated | 9–16 | ribosome-associated | 4–6 |
| endoplasmic reticulum lumen-associated | <9 | ribosomal subunit-associated | 6–8 |
| large ribosomal subunit-associated | <9 | ribosomal protein complex-associated | 6–8 |
| extracellular region-associated | 16–23 |
| MF | Model/Control p < 0.0001 | MF | Experimental/Control p < 0.05 |
|---|---|---|---|
| transforming growth factor-β-binding proteins | 4–6 | nucleic acid-binding proteins | 8–11 |
| growth factor-binding proteins | 6–8 | structural molecular activity | 5–8 |
| cytokine-binding proteins | <4 | chromatin DNA-binding proteins | 2–5 |
| ATP activity-regulating proteins | 4–6 | organic cyclic compound-binding proteins | 8–11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, M.; Zhou, M.; Lu, T.; Lu, Y.; Qin, P.; Liu, C. TMT and PRM Based Quantitative Proteomics to Explore the Protective Role and Mechanism of Iristectorin B in Stroke. Int. J. Mol. Sci. 2023, 24, 15195. https://doi.org/10.3390/ijms242015195
Zheng M, Zhou M, Lu T, Lu Y, Qin P, Liu C. TMT and PRM Based Quantitative Proteomics to Explore the Protective Role and Mechanism of Iristectorin B in Stroke. International Journal of Molecular Sciences. 2023; 24(20):15195. https://doi.org/10.3390/ijms242015195
Chicago/Turabian StyleZheng, Meizhu, Mi Zhou, Tingting Lu, Yao Lu, Peng Qin, and Chunming Liu. 2023. "TMT and PRM Based Quantitative Proteomics to Explore the Protective Role and Mechanism of Iristectorin B in Stroke" International Journal of Molecular Sciences 24, no. 20: 15195. https://doi.org/10.3390/ijms242015195
APA StyleZheng, M., Zhou, M., Lu, T., Lu, Y., Qin, P., & Liu, C. (2023). TMT and PRM Based Quantitative Proteomics to Explore the Protective Role and Mechanism of Iristectorin B in Stroke. International Journal of Molecular Sciences, 24(20), 15195. https://doi.org/10.3390/ijms242015195