
The Endothelial Centrosome: Specific Features and Functional Significance for Endothelial Cell Activity and Barrier Maintenance
 , and
, and 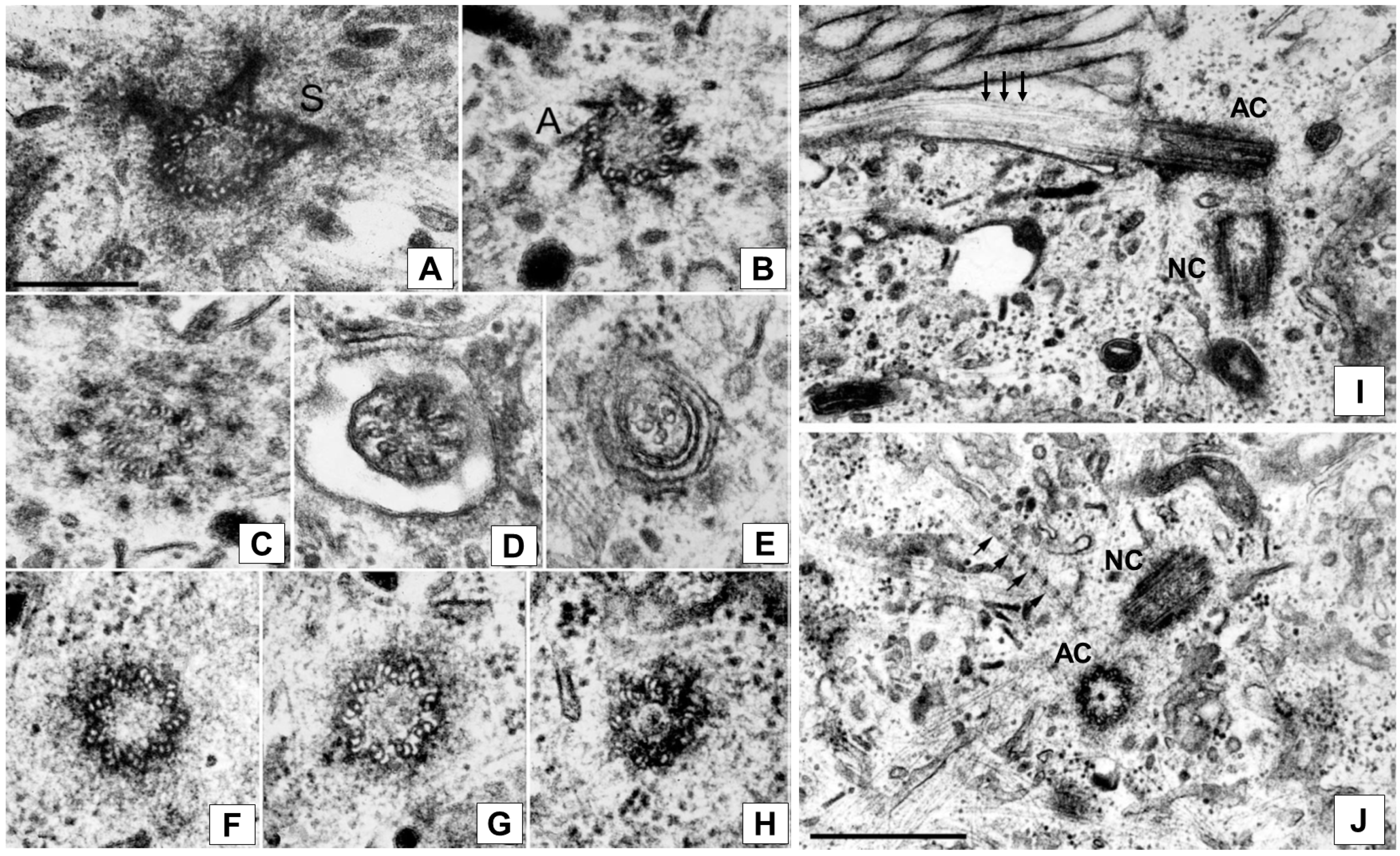{kind=link}
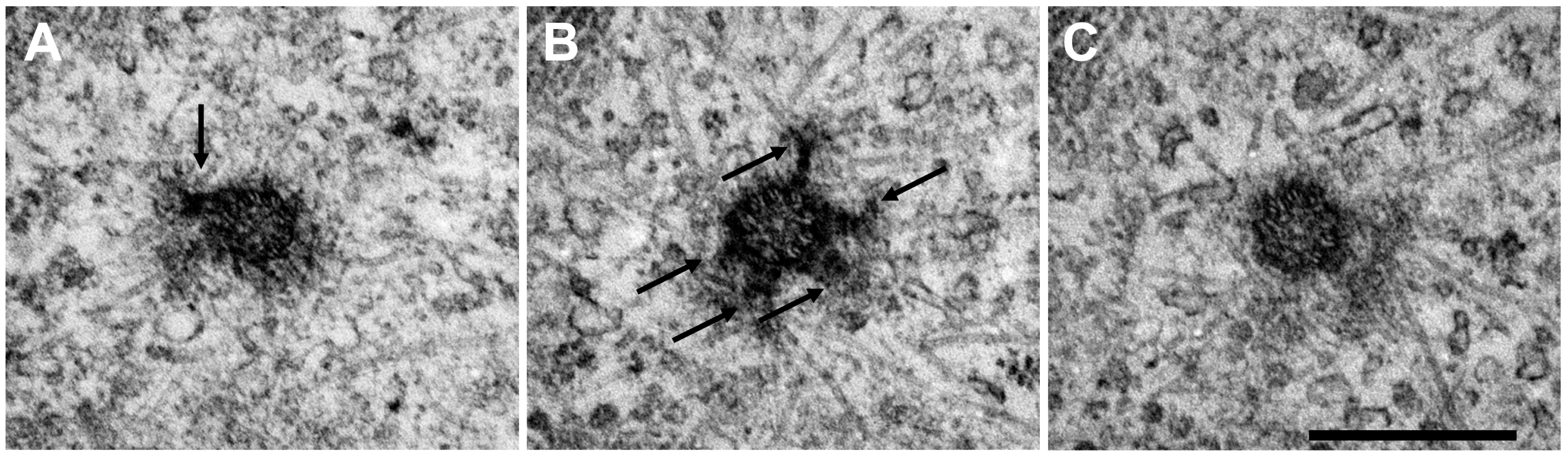{kind=link}
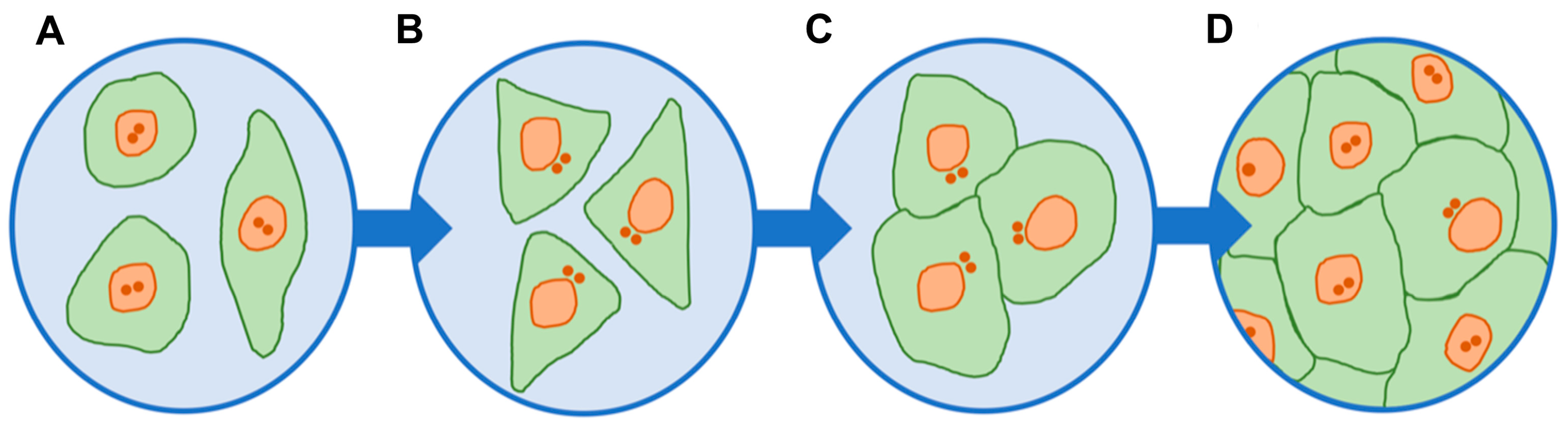{kind=link}
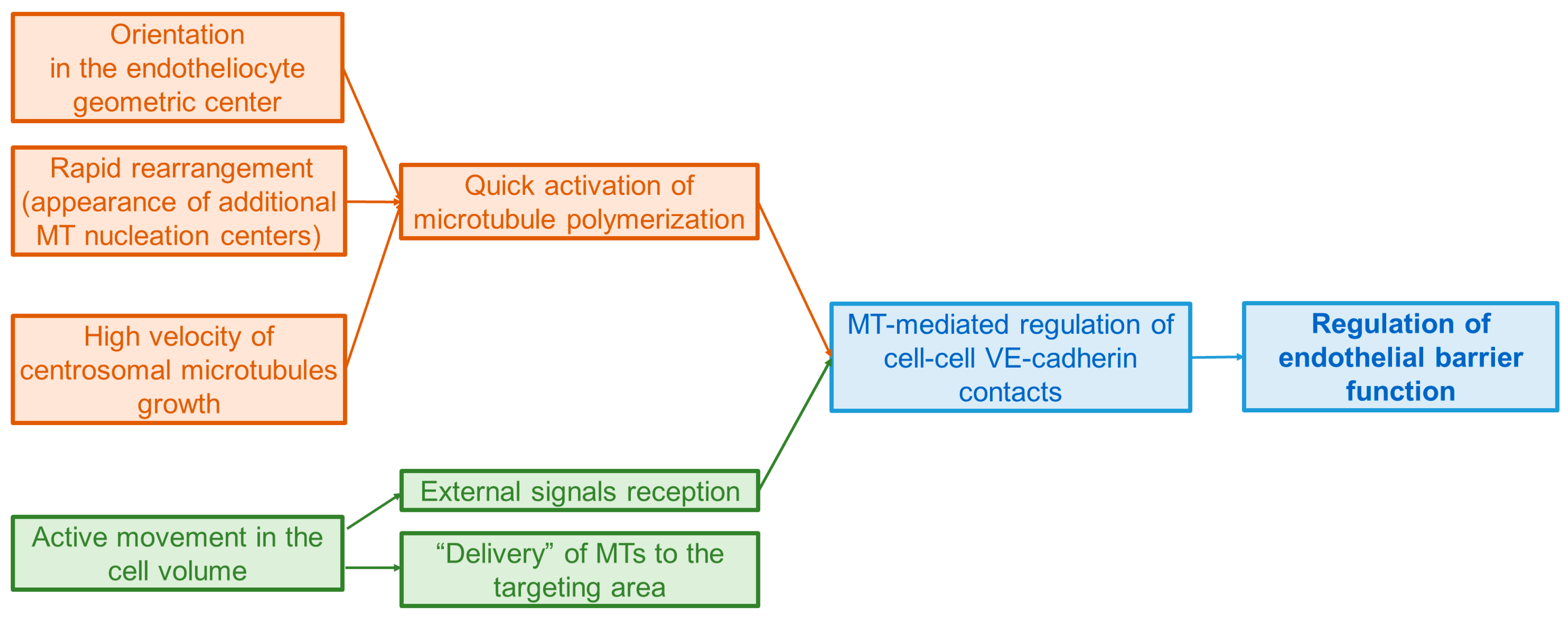{kind=link}
Abstract
:1. Introduction
2. A Quick View on the Mammalian Centrosome Architecture
3. Constancy of Architecture and Tissue-Specific Functioning Features: Can Multiple Centrosome Activities Be Selectively Modulated to Perform Specific Cellular Functions?
4. Ultrastructural Features of Endothelial Cell Centrosome
5. Position and Intracellular Movement of Endothelial Centrosome
6. Endothelial Centrosomes: Over-Duplication in Tumor Vessels and in Angiogenesis
7. Structural Disorders of Centrosomes in Endotheliocytes Leading to Endothelial Barrier Dysfunction
8. The Centrosome of Endotheliocytes, Being an Active Center of Polymerization of Microtubules, Coordinates the Architecture of the Cytoskeleton through Dynamic Microtubules
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wheatley, D.N. The Centriole: A Central Enigma of Cell Biology; Elsevier Biomedical Press: Amsterdam, The Netherlands; New York, NY, USA, 1982; 232p. [Google Scholar]
- Flemming, W. Studien in der Entwicklungsgeschichte der Najaden. Sitzungsber Akad. Wissensch Wien 1875, 71, 81–147. [Google Scholar]
- Hertwig, O. Beitrage zur Kenntniss der Bildung, Befruchtung und Theilung des thierischen Eies. Morphol. Jb. 1875, 1, 347–434. [Google Scholar] [CrossRef]
- Van Beneden, E. Recherches sur les Dicyémides, survivants actuels d’un embranchement des mésozoaires. Bull. Acad. R. Belg. 1876, 41, 1160–1205. [Google Scholar]
- Van Beneden, E.; Neyt, A. Nouvelles recherches sur la fecondation et la division mitosique ches l’Ascaride megalocephale. Bull. Acad. R. Belg. 1887, 14, 1–110. [Google Scholar]
- Boveri, T. Die Bildung der Richtungskörper bei Ascaris Megalocephala und Ascaris Lumbricoides; Zellen-studien, 1; Verlag von Gustav Fisher: Jena, Germany, 1887; 93p. [Google Scholar]
- Boveri, T. Ueber das Verhalten der Centrosomen bei der Befruchtung des Seeigel-Eies, nebst allgemeinen Bemerkungen über Centrosomen und Verwandtes. Verhandl. Phys.-Med. Ges. Würzburg 1895, 29, 1–75. [Google Scholar]
- Bernhard, W.; De Harven, E. Electron microscopic study of the ultrastructure of centrioles in vertebra. Z. Fur Zellforsch. Und Mikrosk. Anat. 1956, 45, 378–398. [Google Scholar]
- Osborn, M.; Weber, K. Cytoplasmic Microtubules in Tissue Culture Cells Appear to Grow from an Organizing Structure towards the Plasma Membrane. Proc. Natl. Acad. Sci. USA 1976, 73, 867–871. [Google Scholar] [CrossRef]
- Jensen, C.G.; Jensen, L.C.; Rieder, C.L. The Occurrence and Structure of Primary Cilia in a Subline of Potorous tridactylus. Exp. Cell Res. 1979, 123, 444–449. [Google Scholar] [CrossRef]
- Fais, D.A.; Nadezhdina, E.S.; Chentsov, Y.S. The Centriolar Rim. The Structure That Maintains the Configuration of Centrioles and Basal Bodies in the Absence of Their Microtubules. Exp. Cell Res. 1986, 164, 27–34. [Google Scholar] [CrossRef]
- Alieva, I.B.; Vorobjev, I.A. Vertebrate Primary Cilia: A Sensory Part of Centrosomal Complex in Tissue Cells, but a “Sleeping Beauty” in Cultured Cells? Cell Biol. Int. 2004, 28, 139–150. [Google Scholar] [CrossRef]
- Vasquez-Limeta, A.; Loncarek, J. Human Centrosome Organization and Function in Interphase and Mitosis. Semin. Cell Dev. Biol. 2021, 117, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular Signalling by Primary Cilia in Development, Organ Function and Disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, H.M.; Valente, E.M. Motile and Non-motile Cilia in Human Pathology: From Function to Phenotypes. J. Pathol. 2017, 241, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Nachury, M.V. How Do Cilia Organize Signalling Cascades? Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130465. [Google Scholar] [CrossRef]
- Nishimura, Y.; Kasahara, K.; Shiromizu, T.; Watanabe, M.; Inagaki, M. Primary Cilia as Signaling Hubs in Health and Disease. Adv. Sci. 2019, 6, 1801138. [Google Scholar] [CrossRef]
- Avidor-Reiss, T.; Carr, A.; Fishman, E.L. The Sperm Centrioles. Mol. Cell. Endocrinol. 2020, 518, 110987. [Google Scholar] [CrossRef]
- Lindemann, C.B.; Lesich, K.A. Functional Anatomy of the Mammalian Sperm Flagellum: Mammalian Sperm Mechanics. Cytoskeleton 2016, 73, 652–669. [Google Scholar] [CrossRef]
- Legendre, M.; Zaragosi, L.-E.; Mitchison, H.M. Motile Cilia and Airway Disease. Semin. Cell Dev. Biol. 2021, 110, 19–33. [Google Scholar] [CrossRef]
- Lee, L.; Ostrowski, L.E. Motile Cilia Genetics and Cell Biology: Big Results from Little Mice. Cell. Mol. Life Sci. 2021, 78, 769–797. [Google Scholar] [CrossRef]
- Lyons, R.A.; Saridogan, E.; Djahanbakhch, O. The Reproductive Significance of Human Fallopian Tube Cilia. Hum. Reprod. Update 2006, 12, 363–372. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Porter, K.R. A Study of the Fine Structure of Ciliated Epithelia. J. Morphol. 1954, 94, 221–281. [Google Scholar] [CrossRef]
- Yamada, E. The fine structure of centriole in some animal cells. In Proceedings of the 1st Conference Asia Oceania, Tokyo, Japan, 12–15 October 1956; pp. 247–250. [Google Scholar]
- Vorobjev, I.A.; YuS, C. Centrioles in the Cell Cycle. I. Epithelial Cells. J. Cell Biol. 1982, 93, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Uzbekov, R.E.; Alieva, I.B. The centrosome—A riddle of the “cell processor”. Tsitologiia 2008, 50, 91–112. [Google Scholar] [PubMed]
- Azimzadeh, J. Exploring the Evolutionary History of Centrosomes. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130453. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Kitagawa, D. Ultrastructural Diversity between Centrioles of Eukaryotes. J. Biochem. 2018, 164, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Uzbekov, R.; Alieva, I. Who Are You, Subdistal Appendages of Centriole? Open Biol. 2018, 8, 180062. [Google Scholar] [CrossRef]
- Gaudin, N.; Martin Gil, P.; Boumendjel, M.; Ershov, D.; Pioche-Durieu, C.; Bouix, M.; Delobelle, Q.; Maniscalco, L.; Phan, T.B.N.; Heyer, V.; et al. Evolutionary Conservation of Centriole Rotational Asymmetry in the Human Centrosome. Elife 2022, 11, e72382. [Google Scholar] [CrossRef]
- Habermann, K.; Lange, B.M. New Insights into Subcomplex Assembly and Modifications of Centrosomal Proteins. Cell Div. 2012, 7, 17. [Google Scholar] [CrossRef]
- Bowler, M.; Kong, D.; Sun, S.; Nanjundappa, R.; Evans, L.; Farmer, V.; Holland, A.; Mahjoub, M.R.; Sui, H.; Loncarek, J. High-Resolution Characterization of Centriole Distal Appendage Morphology and Dynamics by Correlative STORM and Electron Microscopy. Nat. Commun. 2019, 10, 993. [Google Scholar] [CrossRef]
- Chong, W.M.; Wang, W.-J.; Lo, C.-H.; Chiu, T.-Y.; Chang, T.-J.; Liu, Y.-P.; Tanos, B.; Mazo, G.; Tsou, M.-F.B.; Jane, W.-N.; et al. Super-Resolution Microscopy Reveals Coupling between Mammalian Centriole Subdistal Appendages and Distal Appendages. eLife 2020, 9, e53580. [Google Scholar] [CrossRef]
- Tian, Y.; Wei, C.; He, J.; Yan, Y.; Pang, N.; Fang, X.; Liang, X.; Fu, J. Superresolution Characterization of Core Centriole Architecture. J. Cell Biol. 2021, 220, e202005103. [Google Scholar] [CrossRef]
- Li, S.; Fernandez, J.-J.; Marshall, W.F.; Agard, D.A. Three-Dimensional Structure of Basal Body Triplet Revealed by Electron Cryo-Tomography. EMBO J. 2012, 31, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Greenan, G.A.; Keszthelyi, B.; Vale, R.D.; Agard, D.A. Insights into Centriole Geometry Revealed by Cryotomography of Doublet and Triplet Centrioles. Elife 2018, 7, e36851. [Google Scholar] [CrossRef]
- Li, S.; Fernandez, J.-J.; Marshall, W.F.; Agard, D.A. Electron Cryo-Tomography Provides Insight into Procentriole Architecture and Assembly Mechanism. Elife 2019, 8, e43434. [Google Scholar] [CrossRef]
- Steib, E.; Laporte, M.H.; Gambarotto, D.; Olieric, N.; Zheng, C.; Borgers, S.; Olieric, V.; Le Guennec, M.; Koll, F.; Tassin, A.-M.; et al. WDR90 Is a Centriolar Microtubule Wall Protein Important for Centriole Architecture Integrity. Elife 2020, 9, e57205. [Google Scholar] [CrossRef]
- Nazarov, S.; Bezler, A.; Hatzopoulos, G.N.; Nemčíková Villímová, V.; Demurtas, D.; Le Guennec, M.; Guichard, P.; Gönczy, P. Novel Features of Centriole Polarity and Cartwheel Stacking Revealed by Cryo-Tomography. EMBO J. 2020, 39, e106249. [Google Scholar] [CrossRef]
- Klena, N.; Le Guennec, M.; Tassin, A.-M.; van den Hoek, H.; Erdmann, P.S.; Schaffer, M.; Geimer, S.; Aeschlimann, G.; Kovacik, L.; Sadian, Y.; et al. Architecture of the Centriole Cartwheel-Containing Region Revealed by Cryo-Electron Tomography. EMBO J. 2020, 39, e106246. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.J.; Schaeffer, A.; De Pascalis, C.; Letort, G.; Vianay, B.; Bornens, M.; Piel, M.; Blanchoin, L.; Théry, M. Acto-Myosin Network Geometry Defines Centrosome Position. Curr. Biol. 2021, 31, 1206–1220.e5. [Google Scholar] [CrossRef]
- Burakov, A.V.; Nadezhdina, E.S. Centering and Shifting of Centrosomes in Cells. Cells 2020, 9, 1351. [Google Scholar] [CrossRef]
- Riparbelli, M.G.; Persico, V.; Dallai, R.; Callaini, G. Centrioles and Ciliary Structures during Male Gametogenesis in Hexapoda: Discovery of New Models. Cells 2020, 9, 744. [Google Scholar] [CrossRef]
- Bornens, M. Centrosome Composition and Microtubule Anchoring Mechanisms. Curr. Opin. Cell Biol. 2002, 14, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Winey, M.; O’Toole, E. Centriole Structure. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130457. [Google Scholar] [CrossRef] [PubMed]
- Sullenberger, C.; Vasquez-Limeta, A.; Kong, D.; Loncarek, J. With Age Comes Maturity: Biochemical and Structural Transformation of a Human Centriole in the Making. Cells 2020, 9, 1429. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, A.; Bornens, M. Organisation and Functional Regulation of the Centrosome in Animal Cells. Prog. Cell Cycle Res. 1997, 3, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Vertii, A.; Hehnly, H.; Doxsey, S. The Centrosome, a Multitalented Renaissance Organelle. Cold Spring Harb. Perspect. Biol. 2016, 8, a025049. [Google Scholar] [CrossRef]
- Garanina, A.S.; Alieva, I.B.; Bragina, E.E.; Blanchard, E.; Arbeille, B.; Guerif, F.; Uzbekova, S.; Uzbekov, R.E. The Centriolar Adjunct–Appearance and Disassembly in Spermiogenesis and the Potential Impact on Fertility. Cells 2019, 8, 180. [Google Scholar] [CrossRef]
- Araújo, S.J. Centrosomes in Branching Morphogenesis. Results Probl. Cell Differ. 2019, 67, 323–336. [Google Scholar] [CrossRef]
- Ricolo, D.; Castro-Ribera, J.; Araújo, S.J. Cytoskeletal Players in Single-Cell Branching Morphogenesis. Dev. Biol. 2021, 477, 22–34. [Google Scholar] [CrossRef]
- Lüders, J. Nucleating Microtubules in Neurons: Challenges and Solutions. Dev. Neurobiol. 2021, 81, 273–283. [Google Scholar] [CrossRef]
- Kim, A.H.; Puram, S.V.; Bilimoria, P.M.; Ikeuchi, Y.; Keough, S.; Wong, M.; Rowitch, D.; Bonni, A. A Centrosomal Cdc20-APC Pathway Controls Dendrite Morphogenesis in Postmitotic Neurons. Cell 2009, 136, 322–336. [Google Scholar] [CrossRef]
- Yang, J.; Hu, X.; Ma, J.; Shi, S.-H. Centrosome Regulation and Function in Mammalian Cortical Neurogenesis. Curr. Opin. Neurobiol. 2021, 69, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Meka, D.P.; Scharrenberg, R.; Calderon de Anda, F. Emerging Roles of the Centrosome in Neuronal Development. Cytoskeleton 2020, 77, 84–96. [Google Scholar] [CrossRef]
- Tawa, H.; Rikitake, Y.; Takahashi, M.; Amano, H.; Miyata, M.; Satomi-Kobayashi, S.; Kinugasa, M.; Nagamatsu, Y.; Majima, T.; Ogita, H.; et al. Role of Afadin in Vascular Endothelial Growth Factor- and Sphingosine 1-Phosphate-Induced Angiogenesis. Circ. Res. 2010, 106, 1731–1742. [Google Scholar] [CrossRef] [PubMed]
- Bautch, V.L. Ninein Leads the Way in Vessel Sprouting. Arter. Thromb. Vasc. Biol. 2008, 28, 2094–2095. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Schiller, P.; Dieterich, L.C.; Bahram, F.; Iribe, Y.; Hellman, U.; Wikner, C.; Chan, G.; Claesson-Welsh, L.; Dimberg, A. Ninein Is Expressed in the Cytoplasm of Angiogenic Tip-Cells and Regulates Tubular Morphogenesis of Endothelial Cells. Arter. Thromb. Vasc. Biol. 2008, 28, 2123–2130. [Google Scholar] [CrossRef]
- Cao, Z.; Ge, S.; Xu, Z.; Ma, Y.-Q. Β3-Endonexin Interacts with Ninein in Vascular Endothelial Cells to Promote Angiogenesis. Biochem. Biophys. Res. Commun. 2021, 566, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Jalimarada, S.S.; Shivanna, M.; Kini, V.; Mehta, D.; Srinivas, S.P. Microtubule Disassembly Breaks down the Barrier Integrity of Corneal Endothelium. Exp. Eye Res. 2009, 89, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ware, L.B.; Matthay, M.A. The Acute Respiratory Distress Syndrome. N. Engl. J. Med. 2000, 342, 1344–1349. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.G.; Davis, H.W.; Patterson, C.E. Regulation of Endothelial Cell Gap Formation and Barrier Dysfunction: Role of Myosin Light Chain Phosphorylation. J. Cell. Physiol. 1995, 163, 510–522. [Google Scholar] [CrossRef]
- Lum, H.; Malik, A.B. Mechanisms of Increased Endothelial Permeability. Can. J. Physiol. Pharmacol. 1996, 74, 787–800. [Google Scholar] [CrossRef]
- Kushner, E.J.; Ferro, L.S.; Liu, J.-Y.; Durrant, J.R.; Rogers, S.L.; Dudley, A.C.; Bautch, V.L. Excess Centrosomes Disrupt Endothelial Cell Migration via Centrosome Scattering. J. Cell Biol. 2014, 206, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.M.; Nevis, K.R.; Park, H.L.; Rogers, G.C.; Rogers, S.L.; Cook, J.G.; Bautch, V.L. Angiogenic Factor Signaling Regulates Centrosome Duplication in Endothelial Cells of Developing Blood Vessels. Blood 2010, 116, 3108–3117. [Google Scholar] [CrossRef] [PubMed]
- Shakhov, A.S.; Alieva, I.B. The Centrosome as the Main Integrator of Endothelial Cell Functional Activity. Biochemistry 2017, 82, 663–677. [Google Scholar] [CrossRef] [PubMed]
- Bystrevskaya, V.B.; Lichkun, V.V.; Antonov, A.S.; Perov, N.A. An Ultrastructural Study of Centriolar Complexes in Adult and Embryonic Human Aortic Endothelial Cells. Tissue Cell 1988, 20, 493–503. [Google Scholar] [CrossRef]
- Bystrevskaya, V.B.; Lichkun, V.V.; Krushinsky, A.V.; Smirnov, V.N. Centriole Modification in Human Aortic Endothelial Cells. J. Struct. Biol. 1992, 109, 1–12. [Google Scholar] [CrossRef]
- Smurova, K.M.; Birukova, A.A.; Verin, A.D.; Alieva, I.B. Microtubule System in Endothelial Barrier Dysfunction: Disassembly of Peripheral Microtubules and Microtubule Reorganization in Internal Cytoplasm. Cell Tissue Biol. 2008, 2, 45–52. [Google Scholar] [CrossRef]
- Alieva, I.B.; Zemskov, E.A.; Kireev, I.I.; Gorshkov, B.A.; Wiseman, D.A.; Black, S.M.; Verin, A.D. Microtubules Growth Rate Alteration in Human Endothelial Cells. J. Biomed. Biotechnol. 2010, 2010, 1–10. [Google Scholar] [CrossRef]
- Vinogradova, T.M.; Balashova, E.E.; Smirnov, V.N.; Bystrevskaya, V.B. Detection of the Centriole Tyr- or Acet-Tubulin Changes in Endothelial Cells Treated with Thrombin Using Microscopic Immunocytochemistry. Cell Motil. Cytoskelet. 2005, 62, 1–12. [Google Scholar] [CrossRef]
- Shakhov, A.S.; Uzbekov, R.E.; Alieva, I.B. Ultrastructural features of the endotheliocyte centrosome and its possible involvement into the functional activity of endothelium. In Proceedings of the EMBO Conference on Centrosome and Spindle Pole Bodies, Heidelberg, Germany, 24–27 September 2017; p. 80. [Google Scholar]
- Alieva, I.B.; Zemskov, E.A.; Smurova, K.M.; Kaverina, I.N.; Verin, A.D. The Leading Role of Microtubules in Endothelial Barrier Dysfunction: Disassembly of Peripheral Microtubules Leaves behind the Cytoskeletal Reorganization. J. Cell. Biochem. 2013, 114, 2258–2272. [Google Scholar] [CrossRef]
- Gotlieb, A.I.; May, L.M.; Subrahmanyan, L.; Kalnins, V.I. Distribution of Microtubule Organizing Centers in Migrating Sheets of Endothelial Cells. J. Cell Biol. 1981, 91, 589–594. [Google Scholar] [CrossRef]
- Gotlieb, A.I.; Subrahmanyan, L.; Kalnins, V.I. Microtubule-Organizing Centers and Cell Mtgration: Effect of Inhibition of Migration and Microtubule Disruption in Endothelial Cells. J. Cell Biol. 1983, 96, 1266–1272. [Google Scholar] [CrossRef]
- Rogers, K.A.; McKee, N.H.; Kalnins, V.I. Preferential Orientation of Centrioles toward the Heart in Endothelial Cells of Major Blood Vessels Is Reestablished after Reversal of a Segment. Proc. Natl. Acad. Sci. USA 1985, 82, 3272–3276. [Google Scholar] [CrossRef] [PubMed]
- Gotlieb, A.I.; Boden, P. Porcine Aortic Organ Culture: A Model to Study the Cellular Response to Vascular Injury. Vitr. Cell. Dev. Biology. Anim. 1984, 20, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Rogers, K.A.; Boden, P.; Kalnins, V.I.; Gotlieb, A.I. The Distribution of Centrosomes in Endothelial Cells of Non-Wounded and Wounded Aortic Organ Cultures. Cell Tissue Res. 1986, 243, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Coomber, B.L. Centrosome Reorientation in Regenerating Endothelial Monolayers Requires bFCF. J. Cell. Biochem. 1993, 52, 289–296. [Google Scholar] [CrossRef]
- Hotchkiss, K.A.; Ashton, A.W.; Mahmood, R.; Russell, R.G.; Sparano, J.A.; Schwartz, E.L. Inhibition of Endothelial Cell Function in Vitro and Angiogenesis In Vivo by Docetaxel (Taxotere): Association with Impaired Repositioning of the Microtubule Organizing Center. Mol. Cancer Ther. 2002, 1, 1191–1200. [Google Scholar]
- Kushner, E.; Ferro, L.; Yu, Z.; Bautch, V. Excess Centrosomes Perturb Dynamic Endothelial Cell Repolarization During Blood Vessel Formation. Mol. Biol. Cell 2016, 27, 1911–1920. [Google Scholar] [CrossRef]
- Li, J.; Sun, X.; Wang, Z.; Chen, L.; Li, D.; Zhou, J.; Liu, M. Regulation of Vascular Endothelial Cell Polarization and Migration by Hsp70/Hsp90-Organizing Protein. PLoS ONE 2012, 7, e36389. [Google Scholar] [CrossRef]
- Martin, M.; Veloso, A.; Wu, J.; Katrukha, E.A.; Akhmanova, A. Control of Endothelial Cell Polarity and Sprouting Angiogenesis by Non-Centrosomal Microtubules. eLife 2018, 7, e33864. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Angiogenesis in Cancer and Other Diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Akino, T.; Hida, K.; Hida, Y.; Tsuchiya, K.; Freedman, D.; Muraki, C.; Ohga, N.; Matsuda, K.; Akiyama, K.; Harabayashi, T.; et al. Cytogenetic Abnormalities of Tumor-Associated Endothelial Cells in Human Malignant Tumors. Am. J. Pathol. 2009, 175, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Hida, K.; Hida, Y.; Amin, D.N.; Flint, A.F.; Panigrahy, D.; Morton, C.C.; Klagsbrun, M. Tumor-Associated Endothelial Cells with Cytogenetic Abnormalities. Cancer Res. 2004, 64, 8249–8255. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Mouillesseaux, K.P.; Kushner, E.J.; Bautch, V.L. Tumor-Derived Factors and Reduced P53 Promote Endothelial Cell Centrosome Over-Duplication. PLoS ONE 2016, 11, e0168334. [Google Scholar] [CrossRef] [PubMed]
- Kearney, J.B.; Ambler, C.A.; Monaco, K.-A.; Johnson, N.; Rapoport, R.G.; Bautch, V.L. Vascular Endothelial Growth Factor Receptor Flt-1 Negatively Regulates Developmental Blood Vessel Formation by Modulating Endothelial Cell Division. Blood 2002, 99, 2397–2407. [Google Scholar] [CrossRef]
- Buglak, D.B.; Kushner, E.J.; Marvin, A.P.; Davis, K.L.; Bautch, V.L. Excess Centrosomes Disrupt Vascular Lumenization and Endothelial Cell Adherens Junctions. Angiogenesis 2020, 23, 567–575. [Google Scholar] [CrossRef]
- Park, A.Y.J.; Shen, T.-L.; Chien, S.; Guan, J.-L. Role of Focal Adhesion Kinase Ser-732 Phosphorylation in Centrosome Function during Mitosis. J. Biol. Chem. 2009, 284, 9418–9425. [Google Scholar] [CrossRef]
- Shi, X.; Liu, M.; Li, D.; Wang, J.; Aneja, R.; Zhou, J. Cep70 Contributes to Angiogenesis by Modulating Microtubule Rearrangement and Stimulating Cell Polarization and Migration. Cell Cycle 2012, 11, 1554–1563. [Google Scholar] [CrossRef]
- Sun, L.; Xie, S.; Peng, G.; Wang, J.; Li, Y.; Qin, J.; Zhong, D. Lonafarnib Is a Potential Inhibitor for Neovascularization. PLoS ONE 2015, 10, e0122830. [Google Scholar] [CrossRef]
- Karna, P.; Rida, P.C.G.; Turaga, R.C.; Gao, J.; Gupta, M.; Fritz, A.; Werner, E.; Yates, C.; Zhou, J.; Aneja, R. A Novel Microtubule-Modulating Agent EM011 Inhibits Angiogenesis by Repressing the HIF-1α Axis and Disrupting Cell Polarity and Migration. Carcinogenesis 2012, 33, 1769–1781. [Google Scholar] [CrossRef]
- Akhmanova, A.; Steinmetz, M.O. Control of Microtubule Organization and Dynamics: Two Ends in the Limelight. Nat. Rev. Mol. Cell Biol. 2015, 16, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Birukova, A.A.; Birukov, K.G.; Smurova, K.; Adyshev, D.; Kaibuchi, K.; Alieva, I.; Garcia, J.G.N.; Verin, A.D. Novel Role of Microtubules in Thrombin-induced Endothelial Barrier Dysfunction. FASEB J. 2004, 18, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Birukova, A.A.; Smurova, K.; Birukov, K.G.; Usatyuk, P.; Liu, F.; Kaibuchi, K.; Ricks-Cord, A.; Natarajan, V.; Alieva, I.; Garcia, J.G.N.; et al. Microtubule Disassembly Induces Cytoskeletal Remodeling and Lung Vascular Barrier Dysfunction: Role of Rho-Dependent Mechanisms. J. Cell. Physiol. 2004, 201, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Akhmanova, A.; Kapitein, L.C. Mechanisms of Microtubule Organization in Differentiated Animal Cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 541–558. [Google Scholar] [CrossRef] [PubMed]
- Hubert, T.; Vandekerckhove, J.; Gettemans, J. Actin and Arp2/3 Localize at the Centrosome of Interphase Cells. Biochem. Biophys. Res. Commun. 2011, 404, 153–158. [Google Scholar] [CrossRef]
- Hubert, T.; Impe, K.V.; Vandekerckhove, J.; Gettemans, J. The Actin-Capping Protein CapG Localizes to Microtubule-Dependent Organelles during the Cell Cycle. Biochem. Biophys. Res. Commun. 2009, 380, 166–170. [Google Scholar] [CrossRef]
- Farina, F.; Gaillard, J.; Guérin, C.; Sillibourne, J.; Blanchoin, L.; Théry, M. The Centrosome Is an Actin-Organizing Center. Under Revis. Nat. Cell Biol. 2015, 18, 65–75. [Google Scholar] [CrossRef]
- van de Willige, D.; Hummel, J.J.; Alkemade, C.; Kahn, O.I.; Au, F.K.; Qi, R.Z.; Dogterom, M.; Koenderink, G.H.; Hoogenraad, C.C.; Akhmanova, A. Cytolinker Gas2L1 Regulates Axon Morphology through Microtubule-Modulated Actin Stabilization. EMBO Rep. 2019, 20, e47732. [Google Scholar] [CrossRef]
- Alkemade, C.; Wierenga, H.; Volkov, V.A.; Preciado López, M.; Akhmanova, A.; Ten Wolde, P.R.; Dogterom, M.; Koenderink, G.H. Cross-Linkers at Growing Microtubule Ends Generate Forces That Drive Actin Transport. Proc. Natl. Acad. Sci. USA 2022, 119, e2112799119. [Google Scholar] [CrossRef]
- Shum, M.S.Y.; Pasquier, E.; Po’uha, S.T.; O’Neill, G.M.; Chaponnier, C.; Gunning, P.W.; Kavallaris, M. γ-Actin Regulates Cell Migration and Modulates the ROCK Signaling Pathway. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 4423–4433. [Google Scholar] [CrossRef]
- Po‘uha, S.T.; Kavallaris, M. Gamma-Actin Is Involved in Regulating Centrosome Function and Mitotic Progression in Cancer Cells. Cell Cycle 2015, 14, 3908–3919. [Google Scholar] [CrossRef] [PubMed]
- Wordeman, L.; Decarreau, J. Revisiting Actin’s Role in Early Centrosome Separation. Cell Cycle 2016, 15, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.Y.; Gotlieb, A.I. Microtubules Regulate Aortic Endothelial Cell Actin Microfilament Reorganization in Intact and Repairing Monolayers. Histol. Histopathol. 2005, 20, 455–465. [Google Scholar] [PubMed]
- Goldman, R.D.; Hill, B.F.; Steinert, P.; Whitman, M.A.; Zackaroff, R.V. Microtubules and Microtubule Inhibitors; De Brabander, M., De Mey, J., Eds.; Elsevier: New York, NY, USA, 1980; pp. 91–102. [Google Scholar]
- Alieva, I.B.; Nadezhdina, E.S.; Vaisberg, E.A.; Vorobjev, A. Microtubule and Intermediate Filament Patterns around the Centrosome in Interphase Cells. Centrosome 1992, 15, 103–129. [Google Scholar]
- Robert, A.; Herrmann, H.; Davidson, M.W.; Gelfand, V.I. Microtubule-Dependent Transport of Vimentin Filament Precursors Is Regulated by Actin and by the Concerted Action of Rho- and P21-Activated Kinases. FASEB J. 2014, 28, 2879–2890. [Google Scholar] [CrossRef]
- Castañón, M.J.; Walko, G.; Winter, L.; Wiche, G. Plectin-Intermediate Filament Partnership in Skin, Skeletal Muscle, and Peripheral Nerve. Histochem. Cell Biol. 2013, 140, 33–53. [Google Scholar] [CrossRef]
- Jiu, Y.; Lehtimäki, J.; Tojkander, S.; Cheng, F.; Jäälinoja, H.; Liu, X.; Varjosalo, M.; Eriksson, J.E.; Lappalainen, P. Bidirectional Interplay between Vimentin Intermediate Filaments and Contractile Actin Stress Fibers. Cell Rep. 2015, 11, 1511–1518. [Google Scholar] [CrossRef]
- Osmanagic-Myers, S.; Rus, S.; Wolfram, M.; Brunner, D.; Goldmann, W.H.; Bonakdar, N.; Fischer, I.; Reipert, S.; Zuzuarregui, A.; Walko, G.; et al. Plectin Reinforces Vascular Integrity by Mediating Vimentin-Actin Network Crosstalk. J. Cell Sci. 2015, 44, 4138–4150. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakhov, A.S.; Churkina, A.S.; Kotlobay, A.A.; Alieva, I.B. The Endothelial Centrosome: Specific Features and Functional Significance for Endothelial Cell Activity and Barrier Maintenance. Int. J. Mol. Sci. 2023, 24, 15392. https://doi.org/10.3390/ijms242015392
Shakhov AS, Churkina AS, Kotlobay AA, Alieva IB. The Endothelial Centrosome: Specific Features and Functional Significance for Endothelial Cell Activity and Barrier Maintenance. International Journal of Molecular Sciences. 2023; 24(20):15392. https://doi.org/10.3390/ijms242015392
Chicago/Turabian StyleShakhov, Anton Sergeevich, Aleksandra Sergeevna Churkina, Anatoly Alekseevich Kotlobay, and Irina Borisovna Alieva. 2023. "The Endothelial Centrosome: Specific Features and Functional Significance for Endothelial Cell Activity and Barrier Maintenance" International Journal of Molecular Sciences 24, no. 20: 15392. https://doi.org/10.3390/ijms242015392
APA StyleShakhov, A. S., Churkina, A. S., Kotlobay, A. A., & Alieva, I. B. (2023). The Endothelial Centrosome: Specific Features and Functional Significance for Endothelial Cell Activity and Barrier Maintenance. International Journal of Molecular Sciences, 24(20), 15392. https://doi.org/10.3390/ijms242015392