

Anxiety Modulation by Cannabinoids—The Role of Stress Responses and Coping

Abstract
:1. Introduction
1.1. The Endocannabinoid System
1.2. Functions
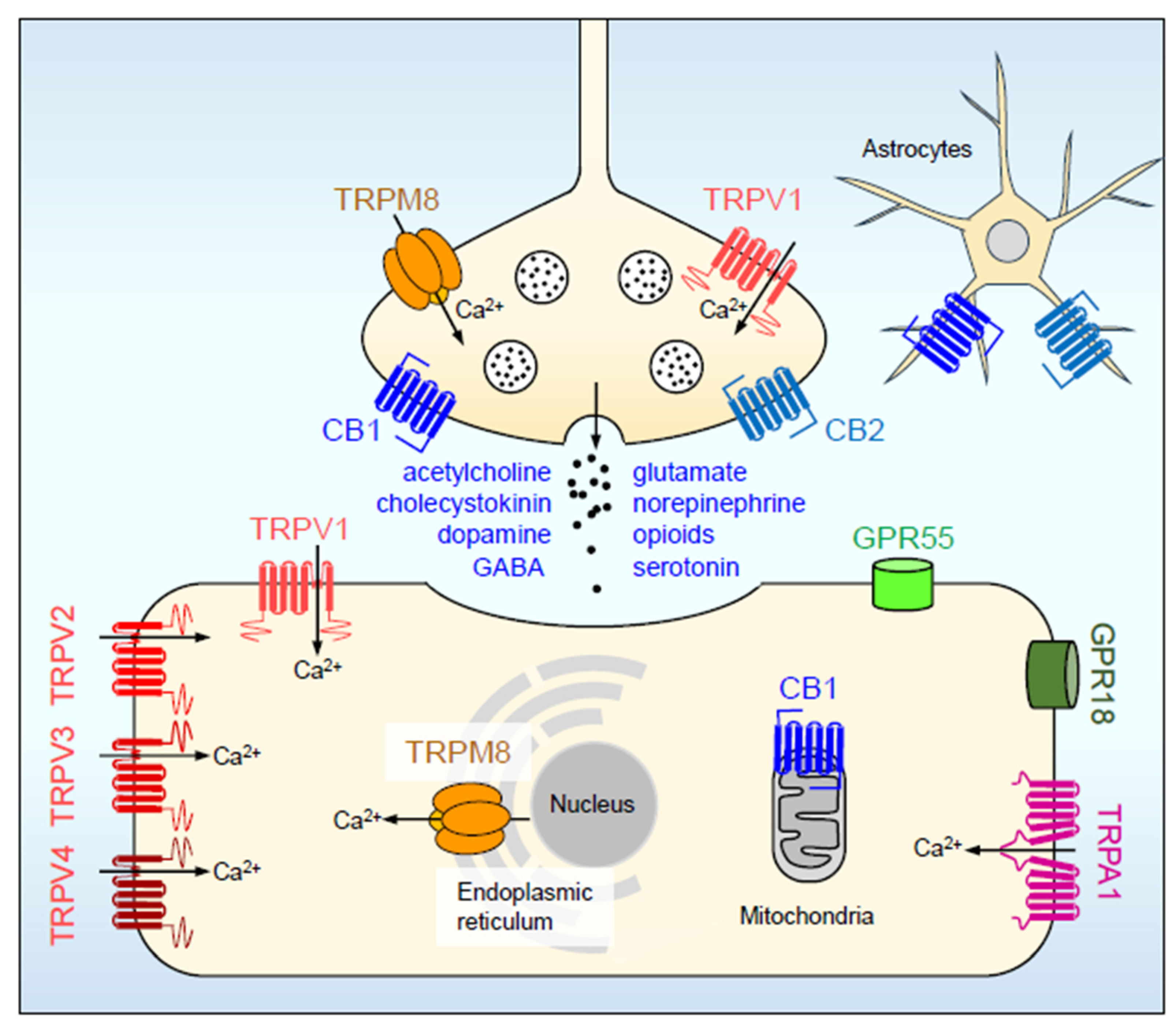
1.3. Mechanisms
2. Research Tools—What Questions Can They Answer?
2.1. Targeting Cannabinoid Receptors
2.2. Targeting the Degrading Enzymes
2.3. Targeting Other Mechanisms
3. Cannabinoids and Anxiety
3.1. Transgenic Animals
3.2. CB1 Receptor Pharmacology
3.3. Enzyme Inhibitors and Transporters
3.4. FAAH Inhibitors and Coping Styles
4. Overall Interpretation of the Findings
- The endocannabinoid system controls behavior by its interactions with the stress system (HPA-axis) and ensures normal functioning by eliminating excessive stress responses at all three levels: the hormonal, neural, and behavioral [202].
- The endocannabinoid system contributes to the integration of perception and execution, by allowing this adaptation to the environment. It buffers maladaptive responses and protects against psychiatric symptoms [203].
- The endocannabinoid system promotes an active coping with challenges, which confers the organism advantages in critical situations. This may be the common denominator of its anxiolytic- and antidepression-like effects [204].
Funding
Conflicts of Interest
References
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Rezende, B.; Alencar, A.K.N.; de Bem, G.F.; Fontes-Dantas, F.L.; Montes, G.C. Endocannabinoid System: Chemical Characteristics and Biological Activity. Pharmaceuticals 2023, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Dysarz, F.A., 3rd; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 1988, 34, 605–613. [Google Scholar] [PubMed]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol. Ther. 1997, 74, 129–180. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V. Endocannabinoids: Synthesis and degradation. In Reviews of Physiology, Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 2008; Volume 160, pp. 1–24. [Google Scholar] [CrossRef]
- Jain, T.; Wager-Miller, J.; Mackie, K.; Straiker, A. Diacylglycerol lipaseα (DAGLα) and DAGLβ cooperatively regulate the production of 2-arachidonoyl glycerol in autaptic hippocampal neurons. Mol. Pharmacol. 2013, 84, 296–302. [Google Scholar] [CrossRef]
- Lu, H.C.; Mackie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef]
- Pertwee, R.G. Cannabinoid pharmacology: The first 66 years. Br. J. Pharmacol. 2006, 147 (Suppl. S1), S163–S171. [Google Scholar] [CrossRef]
- Lowe, H.; Toyang, N.; Steele, B.; Bryant, J.; Ngwa, W. The Endocannabinoid System: A Potential Target for the Treatment of Various Diseases. Int. J. Mol. Sci. 2021, 22, 9472. [Google Scholar] [CrossRef]
- Rakotoarivelo, V.; Sihag, J.; Flamand, N. Role of the Endocannabinoid System in the Adipose Tissue with Focus on Energy Metabolism. Cells 2021, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- Fraga, D.; Zanoni, C.I.; Rae, G.A.; Parada, C.A.; Souza, G.E. Endogenous cannabinoids induce fever through the activation of CB1 receptors. Br. J. Pharmacol. 2009, 157, 1494–1501. [Google Scholar] [CrossRef]
- Rahaman, O.; Ganguly, D. Endocannabinoids in immune regulation and immunopathologies. Immunology 2021, 164, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, B.M.; Rebelo, I. Cannabis and Cannabinoids in Reproduction and Fertility: Where We Stand. Reprod. Sci. 2022, 29, 2429–2439. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Scipioni, L.; Maccarrone, M. Endocannabinoid system and adult neurogenesis: A focused review. Curr. Opin. Pharmacol. 2020, 50, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kasatkina, L.A.; Rittchen, S.; Sturm, E.M. Neuroprotective and Immunomodulatory Action of the Endocannabinoid System under Neuroinflammation. Int. J. Mol. Sci. 2021, 22, 5431. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Moncada, I.; Rodrigues, R.S.; Fundazuri, U.B.; Bellocchio, L.; Marsicano, G. Type-1 cannabinoid receptors and their ever-expanding roles in brain energy processes. J. Neurochem. 2023; early view. [Google Scholar] [CrossRef] [PubMed]
- Solinas, M.; Goldberg, S.R.; Piomelli, D. The endocannabinoid system in brain reward processes. Br. J. Pharmacol. 2008, 154, 369–383. [Google Scholar] [CrossRef]
- Zanettini, C.; Panlilio, L.V.; Alicki, M.; Goldberg, S.R.; Haller, J.; Yasar, S. Effects of endocannabinoid system mod-ulation on cognitive and emotional behavior. Front. Behav. Neurosci. 2011, 5, 57. [Google Scholar] [CrossRef]
- Micale, V.; Mazzola, C.; Drago, F. Endocannabinoids and neurodegenerative diseases. Pharmacol. Res. 2007, 56, 382–392. [Google Scholar] [CrossRef]
- Moreno, E.; Cavic, M.; Krivokuca, A.; Canela, E.I. The Interplay between Cancer Biology and the Endocannabinoid System-Significance for Cancer Risk, Prognosis and Response to Treatment. Cancers 2020, 12, 3275. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Steffens, S. The emerging role of the endocannabinoid system in cardiovascular disease. Semin. Immunopathol. 2009, 31, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.K.; Chan, S.C.; Yung, W.H. Presynaptic inhibition of GABAergic inputs to rat substantia nigra pars re-ticulata neurones by a cannabinoid agonist. Neuroreport 1998, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I.; Nicoll, R.A. Endocannabinoid signaling in the brain. Science 2002, 296, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Katona, I.; Sperlágh, B.; Sík, A.; Käfalvi, A.; Vizi, E.S.; Mackie, K.; Freund, T.F. Presynaptically located CB1 can-nabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J. Neuro-Sci. Off. J. Soc. Neurosci. 1999, 19, 4544–4558. [Google Scholar] [CrossRef] [PubMed]
- Bowers, M.E.; Ressler, K.J. Interaction between the cholecystokinin and endogenous cannabinoid systems in cued fear expression and extinction retention. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2015, 40, 688–700. [Google Scholar] [CrossRef]
- Alger, B.E. Retrograde signaling in the regulation of synaptic transmission: Focus on endocannabinoids. Prog. Neurobiol. 2002, 68, 247–286. [Google Scholar] [CrossRef]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef]
- Schlicker, E.; Kathmann, M. Modulation of transmitter release via presynaptic cannabinoid receptors. Trends Pharmacol. Sci. 2001, 22, 565–572. [Google Scholar] [CrossRef]
- Gong, J.P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 re-ceptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Di Marzo, V. Non-CB1, non-CB2 receptors for endocannabinoids, plant cannabinoids, and syn-thetic cannabimimetics: Focus on G-protein-coupled receptors and transient receptor potential channels. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2010, 5, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 2019, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Biringer, R.G. Endocannabinoid signaling pathways: Beyond CB1R and CB2R. J. Cell Commun. Signal. 2021, 15, 335–360. [Google Scholar] [CrossRef] [PubMed]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.Y.; Lu, H.C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that in-creases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Δ(9)-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [PubMed]
- Puighermanal, E.; Busquets-Garcia, A.; Maldonado, R.; Ozaita, A. Cellular and intracellular mechanisms involved in the cognitive impairment of cannabinoids. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2012, 367, 3254–3263. [Google Scholar] [CrossRef]
- Castillo, P.E.; Younts, T.J.; Chávez, A.E.; Hashimotodani, Y. Endocannabinoid signaling and synaptic function. Neuron 2012, 76, 70–81. [Google Scholar] [CrossRef]
- Eraso-Pichot, A.; Pouvreau, S.; Olivera-Pinto, A.; Gomez-Sotres, P.; Skupio, U.; Marsicano, G. Endocannabinoid signaling in astrocytes. Glia 2023, 71, 44–59. [Google Scholar] [CrossRef]
- Ledent, C.; Valverde, O.; Cossu, G.; Petitet, F.; Aubert, J.F.; Beslot, F.; Böhme, G.A.; Imperato, A.; Pedrazzini, T.; Roques, B.P.; et al. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science 1999, 283, 401–404. [Google Scholar] [CrossRef]
- Buckley, N.E.; McCoy, K.L.; Mezey, E.; Bonner, T.; Zimmer, A.; Felder, C.C.; Glass, M.; Zimmer, A. Immu-no-modulation by cannabinoids is absent in mice deficient for the cannabinoid CB(2) receptor. Eur. J. Pharmacol. 2000, 396, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Járai, Z.; Wagner, J.A.; Varga, K.; Lake, K.D.; Compton, D.R.; Martin, B.R.; Zimmer, A.M.; Bonner, T.I.; Buckley, N.E.; Mezey, E.; et al. Cannabinoid-induced mesenteric vasodilation through an endo-thelial site distinct from CB1 or CB2 receptors. Proc. Natl. Acad. Sci. USA 1999, 96, 14136–14141. [Google Scholar] [CrossRef] [PubMed]
- Guggenhuber, S.; Monory, K.; Lutz, B.; Klugmann, M. AAV vector-mediated overexpression of CB1 cannabinoid receptor in pyramidal neurons of the hippocampus protects against seizure-induced excitoxicity. PLoS ONE 2010, 5, e15707. [Google Scholar] [CrossRef] [PubMed]
- Klugmann, M.; Goepfrich, A.; Friemel, C.M.; Schneider, M. AAV-Mediated Overexpression of the CB1 Receptor in the mPFC of Adult Rats Alters Cognitive Flexibility, Social Behavior, and Emotional Reactivity. Front. Behav. Neurosci. 2011, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Di Maria, V.; Moindrot, M.; Ryde, M.; Bono, A.; Quintino, L.; Ledri, M. Development and Validation of CRISPR Activator Systems for Overexpression of CB1 Receptors in Neurons. Front. Mol. Neurosci. 2020, 13, 168. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Ledent, C.; Parmentier, M.; Maldonado, R.; Valverde, O. Involvement of CB1 cannabinoid receptors in emotional behaviour. Psychopharmacology 2002, 159, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Bakos, N.; Szirmay, M.; Ledent, C.; Freund, T.F. The effects of genetic and pharmacological blockade of the CB1 cannabinoid receptor on anxiety. Eur. J. Neurosci. 2002, 16, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Varga, B.; Ledent, C.; Freund, T.F. CB1 cannabinoid receptors mediate anxiolytic effects: Convergent genetic and pharmacological evidence with CB1-specific agents. Behav. Pharmacol. 2004, 15, 299–304. [Google Scholar] [CrossRef]
- Urigüen, L.; Pérez-Rial, S.; Ledent, C.; Palomo, T.; Manzanares, J. Impaired action of anxiolytic drugs in mice deficient in cannabinoid CB1 receptors. Neuropharmacology 2004, 46, 966–973. [Google Scholar] [CrossRef]
- Haller, J.; Varga, B.; Ledent, C.; Barna, I.; Freund, T.F. Context-dependent effects of CB1 cannabinoid gene disruption on anxiety-like and social behaviour in mice. Eur. J. Neurosci. 2004, 19, 1906–1912. [Google Scholar] [CrossRef]
- Jacob, W.; Yassouridis, A.; Marsicano, G.; Monory, K.; Lutz, B.; Wotjak, C.T. Endocannabinoids render exploratory behaviour largely independent of the test aversiveness: Role of glutamatergic transmission. Genes Brain Behav. 2009, 8, 685–698. [Google Scholar] [CrossRef]
- Kamprath, K.; Plendl, W.; Marsicano, G.; Deussing, J.M.; Wurst, W.; Lutz, B.; Wotjak, C.T. Endocannabinoids mediate acute fear adaptation via glutamatergic neurons independently of corticotropin-releasing hormone signaling. Genes Brain Behav. 2009, 8, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.N.; Hillard, C.J.; McEwen, B.S. Alterations in corticolimbic dendritic morphology and emotional behavior in cannabinoid CB1 receptor-deficient mice parallel the effects of chronic stress. Cereb. Cortex 2011, 21, 2056–2064. [Google Scholar] [CrossRef]
- Derks, N.M.; Pintér, O.; Zuure, W.; Ledent, C.; Watanabe, M.; Molnár, C.S.; Wei, Y.; Roubos, E.W.; Wu, S.; Hrabovszky, E.; et al. Cannabinoid modulation of midbrain urocortin 1 neurones during acute and chronic stress. J. Neuroendocrinol. 2012, 24, 1447–1461. [Google Scholar] [CrossRef] [PubMed]
- Dubreucq, S.; Matias, I.; Cardinal, P.; Häring, M.; Lutz, B.; Marsicano, G.; Chaouloff, F. Genetic dissection of the role of cannabinoid type-1 receptors in the emotional consequences of repeated social stress in mice. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2012, 37, 1885–1900. [Google Scholar] [CrossRef] [PubMed]
- Litvin, Y.; Phan, A.; Hill, M.N.; Pfaff, D.W.; McEwen, B.S. CB1 receptor signaling regulates social anxiety and memory. Genes Brain Behav. 2013, 12, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Ruehle, S.; Remmers, F.; Romo-Parra, H.; Massa, F.; Wickert, M.; Wörtge, S.; Häring, M.; Kaiser, N.; Marsicano, G.; Pape, H.C.; et al. Cannabinoid CB1 receptor in dorsal telencephalic glutamatergic neurons: Distinctive sufficiency for hippo-campus-dependent and amygdala-dependent synaptic and behavioral functions. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 10264–10277. [Google Scholar] [CrossRef] [PubMed]
- Remmers, F.; Lange, M.D.; Hamann, M.; Ruehle, S.; Pape, H.C.; Lutz, B. Addressing sufficiency of the CB1 receptor for endocannabinoid-mediated functions through conditional genetic rescue in forebrain GABAergic neurons. Brain Struct. Funct. 2017, 222, 3431–3452. [Google Scholar] [CrossRef]
- Fyke, W.; Premoli, M.; Echeverry Alzate, V.; López-Moreno, J.A.; Lemaire-Mayo, V.; Crusio, W.E.; Marsicano, G.; Wöhr, M.; Pietropaolo, S. Communication and social interaction in the cannabinoid-type 1 receptor null mouse: Im-plications for autism spectrum disorder. Autism Res. Off. J. Int. Soc. Autism Res. 2021, 14, 1854–1872. [Google Scholar] [CrossRef]
- Maccarrone, M.; Valverde, O.; Barbaccia, M.L.; Castañé, A.; Maldonado, R.; Ledent, C.; Parmentier, M.; Finazzi-Agrò, A. Age-related changes of anandamide metabolism in CB1 cannabinoid receptor knockout mice: Correlation with behaviour. Eur. J. Neurosci. 2002, 15, 1178–1186. [Google Scholar] [CrossRef]
- Rácz, I.; Nent, E.; Erxlebe, E.; Zimmer, A. CB1 receptors modulate affective behaviour induced by neuropathic pain. Brain Res. Bull. 2015, 114, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Bowers, M.E.; Ressler, K.J. Sex-dependence of anxiety-like behavior in cannabinoid receptor 1 (Cnr1) knockout mice. Behav. Brain Res. 2016, 300, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Thiemann, G.; Watt, C.A.; Ledent, C.; Molleman, A.; Hasenöhrl, R.U. Modulation of anxiety by acute blockade and genetic deletion of the CB(1) cannabinoid receptor in mice together with biogenic amine changes in the forebrain. Behav. Brain Res. 2009, 200, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Terzian, A.L.; Micale, V.; Wotjak, C.T. Cannabinoid receptor type 1 receptors on GABAergic vs. glutamatergic neurons differentially gate sex-dependent social interest in mice. Eur. J. Neurosci. 2014, 40, 2293–2298. [Google Scholar] [CrossRef] [PubMed]
- Degroot, A.; Nomikos, G.G. Genetic deletion and pharmacological blockade of CB1 receptors modulates anxiety in the shock-probe burying test. Eur. J. Neurosci. 2004, 20, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Alvaro, A.; Aracil-Fernández, A.; García-Gutiérrez, M.S.; Navarrete, F.; Manzanares, J. Deletion of CB2 cannabinoid receptor induces schizophrenia-related behaviors in mice. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2011, 36, 1489–1504. [Google Scholar] [CrossRef] [PubMed]
- Flake, N.M.; Zweifel, L.S. Behavioral effects of pulp exposure in mice lacking cannabinoid receptor 2. J. Endod. 2012, 38, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kim, J. CB2 Cannabinoid Receptor Knockout in Mice Impairs Contextual Long-Term Memory and En-hances Spatial Working Memory. Neural Plast. 2016, 2016, 9817089. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef]
- Rinaldi-Carmona, M.; Barth, F.; Héaulme, M.; Shire, D.; Calandra, B.; Congy, C.; Martinez, S.; Maruani, J.; Néliat, G.; Caput, D. SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Lett. 1994, 350, 240–244. [Google Scholar] [CrossRef]
- Mechoulam, R.; Gaoni, Y. A total synthesis of dl-delta-1-tetrahydrocannabinol, the active constituent of hashish. J. Am. Chem. Soc. 1965, 87, 3273–3275. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, J.; Saady, M.; Cardounel, A.; Kalimi, M. Identification and characterization of a novel synthetic can-nabinoid CP 55,940 binder in rat brain cytosol. Mol. Cell. Biochem. 1998, 181, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.R.; Tang, J.Q.; Zhang, W.N.; Zhuang, C.L.; Shi, Y. Rational drug design of CB2 receptor ligands: From 2012 to 2021. RSC Adv. 2022, 12, 35242–35259. [Google Scholar] [CrossRef] [PubMed]
- Griebel, G.; Stemmelin, J.; Scatton, B. Effects of the cannabinoid CB1 receptor antagonist rimonabant in models of emotional reactivity in rodents. Biol. Psychiatry 2005, 57, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.; Hernández, E.; Muñoz, R.M.; del Arco, I.; Villanúa, M.A.; Carrera, M.R.; Rodríguez de Fonseca, F. Acute administration of the CB1 cannabinoid receptor antagonist SR 141716A induces anxiety-like responses in the rat. Neuroreport 1997, 8, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, M.; Jamontt, J.; Gliniak, H. Effects of cannabinoids on the anxiety-like response in mice. Pharmacol. Rep. PR 2006, 58, 200–206. [Google Scholar] [PubMed]
- Sink, K.S.; Segovia, K.N.; Sink, J.; Randall, P.A.; Collins, L.E.; Correa, M.; Markus, E.J.; Vemuri, V.K.; Makriyannis, A.; Salamone, J.D. Potential anxiogenic effects of cannabinoid CB1 receptor antagonists/inverse agonists in rats: Compar-isons between AM4113, AM251, and the benzodiazepine inverse agonist FG-7142. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2010, 20, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Black, M.D.; Stevens, R.J.; Rogacki, N.; Featherstone, R.E.; Senyah, Y.; Giardino, O.; Borowsky, B.; Stemmelin, J.; Cohen, C.; Pichat, P.; et al. AVE1625, a cannabinoid CB1 receptor antagonist, as a co-treatment with antipsychotics for schizophrenia: Improvement in cognitive function and re-duction of an-tipsychotic-side effects in rodents. Psychopharmacology 2011, 215, 149–163. [Google Scholar] [CrossRef]
- Patel, S.; Hillard, C.J. Pharmacological evaluation of cannabinoid receptor ligands in a mouse model of anxiety: Further evidence for an anxiolytic role for endogenous cannabinoid signaling. J. Pharmacol. Exp. Ther. 2006, 318, 304–311. [Google Scholar] [CrossRef]
- Hill, M.N.; Gorzalka, B.B. Enhancement of anxiety-like responsiveness to the cannabinoid CB(1) receptor agonist HU-210 following chronic stress. Eur. J. Pharmacol. 2004, 499, 291–295. [Google Scholar] [CrossRef]
- Marco, E.M.; Pérez-Alvarez, L.; Borcel, E.; Rubio, M.; Guaza, C.; Ambrosio, E.; File, S.E.; Viveros, M.P. In-volvement of 5-HT1A receptors in behavioural effects of the cannabinoid receptor agonist CP 55,940 in male rats. Behav. Pharmacol. 2004, 15, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Célérier, E.; Ahdepil, T.; Wikander, H.; Berrendero, F.; Nyberg, F.; Maldonado, R. Influence of the anabol-ic-androgenic steroid nandrolone on cannabinoid dependence. Neuropharmacology 2006, 50, 788–806. [Google Scholar] [CrossRef] [PubMed]
- Malheiro, R.F.; Carmo, H.; Carvalho, F.; Silva, J.P. Cannabinoid-mediated targeting of mitochondria on the mod-ulation of mitochondrial function and dynamics. Pharmacol. Res. 2023, 187, 106603. [Google Scholar] [CrossRef] [PubMed]
- Oubraim, S.; Wang, R.; Hausknecht, K.A.; Shen, R.Y.; Haj-Dahmane, S. Tonic Endocannabinoid Signaling Gates Synaptic Plasticity in Dorsal Raphe Nucleus Serotonin Neurons Through Peroxisome Proliferator-Activated Receptors. Front. Pharmacol. 2021, 12, 691219. [Google Scholar] [CrossRef] [PubMed]
- Piomelli, D. The molecular logic of endocannabinoid signalling. Nat. Rev. Neurosci. 2003, 4, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Goodenough, S.; Monory, K.; Hermann, H.; Eder, M.; Cannich, A.; Azad, S.C.; Cascio, M.G.; Gutiérrez, S.O.; van der Stelt, M.; et al. CB1 cannabinoid receptors and on-demand defense against excitotoxicity. Science 2003, 302, 84–88. [Google Scholar] [CrossRef]
- Rancz, E.A.; Häusser, M. Dendritic calcium spikes are tunable triggers of cannabinoid release and short-term synaptic plasticity in cerebellar Purkinje neurons. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 5428–5437. [Google Scholar] [CrossRef]
- Tsou, K.; Brown, S.; Sañudo-Peña, M.C.; Mackie, K.; Walker, J.M. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience 1998, 83, 393–411. [Google Scholar] [CrossRef]
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The emerging role of the endocannabinoid system in en-docrine regulation and energy balance. Endocr. Rev. 2006, 27, 73–100. [Google Scholar] [CrossRef]
- Medeiros, D.C.; Cota, V.R.; Oliveira, A.C.P.; Moreira, F.A.; Moraes, M.F.D. The Endocannabinoid System Ac-tivation as a Neural Network Desynchronizing Mediator for Seizure Suppression. Front. Behav. Neurosci. 2020, 14, 603245. [Google Scholar] [CrossRef]
- Deutsch, D.G.; Ueda, N.; Yamamoto, S. The fatty acid amide hydrolase (FAAH). Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Egertová, M.; Giang, D.K.; Cravatt, B.F.; Elphick, M.R. A new perspective on cannabinoid signalling: Comple-mentary localization of fatty acid amide hydrolase and the CB1 receptor in rat brain. Proc. Biol. Sci. 1998, 265, 2081–2085. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ordóñez, A.; Martín-Fontecha, M.; Ortega-Gutiérrez, S.; López-Rodríguez, M.L. Monoacylglycerol lipase (MAGL) as a promising therapeutic target. Biochem. Pharmacol. 2018, 157, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Savinainen, J.R.; Saario, S.M.; Laitinen, J.T. The serine hydrolases MAGL, ABHD6 and ABHD12 as guardians of 2-arachidonoylglycerol signalling through cannabinoid receptors. Acta Physiol. 2012, 204, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; Arrabal, S.; Cifuentes, M.; Grondona, J.M.; Pérez-Martín, M.; Rubio, L.; Vargas, A.; Serrano, A.; Pavón, F.J.; Suárez, J.; et al. Localization of the cannabinoid CB1 receptor and the 2-AG synthesizing (DAGLα) and degrading (MAGL, FAAH) enzymes in cells expressing the Ca2+-binding proteins calbindin, calretinin, and parvalbumin in the adult rat hippocampus. Front. Neuroanat. 2014, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Kathuria, S.; Gaetani, S.; Fegley, D.; Valiño, F.; Duranti, A.; Tontini, A.; Mor, M.; Tarzia, G.; La Rana, G.; Calignano, A.; et al. Modulation of anxiety through blockade of anandamide hydrolysis. Nat. Med. 2003, 9, 76–81. [Google Scholar] [CrossRef]
- Long, J.Z.; Li, W.; Booker, L.; Burston, J.J.; Kinsey, S.G.; Schlosburg, J.E.; Pavón, F.J.; Serrano, A.M.; Selley, D.E.; Parsons, L.H.; et al. Selective blockade of 2-arachidonoylglycerol hydrolysis produces canna-binoid behavioral effects. Nat. Chem. Biol. 2009, 5, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Cravatt, B.F.; Demarest, K.; Patricelli, M.P.; Bracey, M.H.; Giang, D.K.; Martin, B.R.; Lichtman, A.H. Su-persensitivity to anandamide and enhanced endogenous cannabinoid signaling in mice lacking fatty acid amide hydrolase. Proc. Natl. Acad. Sci. USA 2001, 98, 9371–9376. [Google Scholar] [CrossRef]
- Rubino, T.; Realini, N.; Castiglioni, C.; Guidali, C.; Viganó, D.; Marras, E.; Petrosino, S.; Perletti, G.; Maccarrone, M.; Di Marzo, V.; et al. Role in anxiety behavior of the endocannabinoid system in the prefrontal cortex. Cereb. Cortex 2008, 18, 1292–1301. [Google Scholar] [CrossRef]
- Navia-Paldanius, D.; Aaltonen, N.; Lehtonen, M.; Savinainen, J.R.; Taschler, U.; Radner, F.P.; Zimmermann, R.; Laitinen, J.T. Increased tonic cannabinoid CB1R activity and brain region-specific desensitization of CB1R Gi/o signaling axis in mice with global genetic knockout of monoacylglycerol lipase. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2015, 77, 180–188. [Google Scholar] [CrossRef]
- Hill, M.N.; Karacabeyli, E.S.; Gorzalka, B.B. Estrogen recruits the endocannabinoid system to modulate emo-tionality. Psychoneuroendocrinology 2007, 32, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Cippitelli, A.; Cannella, N.; Braconi, S.; Duranti, A.; Tontini, A.; Bilbao, A.; Defonseca, F.R.; Piomelli, D.; Ciccocioppo, R. Increase of brain endocannabinoid anandamide levels by FAAH inhibition and alcohol abuse behaviours in the rat. Psychopharmacology 2008, 198, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Moreira, F.A.; Kaiser, N.; Monory, K.; Lutz, B. Reduced anxiety-like behaviour induced by genetic and phar-macological inhibition of the endocannabinoid-degrading enzyme fatty acid amide hydrolase (FAAH) is mediated by CB1 receptors. Neuropharmacology 2008, 54, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Scherma, M.; Medalie, J.; Fratta, W.; Vadivel, S.K.; Makriyannis, A.; Piomelli, D.; Mikics, E.; Haller, J.; Yasar, S.; Tanda, G.; et al. The endogenous cannabinoid anandamide has effects on motivation and anxiety that are revealed by fatty acid amide hydrolase (FAAH) inhibition. Neuropharmacology 2008, 54, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Naidu, P.S.; Varvel, S.A.; Ahn, K.; Cravatt, B.F.; Martin, B.R.; Lichtman, A.H. Evaluation of fatty acid amide hydrolase inhibition in murine models of emotionality. Psychopharmacology 2007, 192, 61–70. [Google Scholar] [CrossRef]
- Naderi, N.; Haghparast, A.; Saber-Tehrani, A.; Rezaii, N.; Alizadeh, A.M.; Khani, A.; Motamedi, F. Interaction between cannabinoid compounds and diazepam on anxiety-like behaviour of mice. Pharmacol. Biochem. Behav. 2008, 89, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Barna, I.; Barsvari, B.; Gyimesi Pelczer, K.; Yasar, S.; Panlilio, L.V.; Goldberg, S. Interactions between environmental aversiveness and the anxiolytic effects of enhanced cannabinoid signaling by FAAH inhibition in rats. Psychopharmacology 2009, 204, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Cippitelli, A.; Astarita, G.; Duranti, A.; Caprioli, G.; Ubaldi, M.; Stopponi, S.; Kallupi, M.; Sagratini, G.; Rodrìguez de Fonseca, F.; Piomelli, D.; et al. Endocannabinoid regulation of acute and protracted nicotine with-drawal: Effect of FAAH inhibition. PLoS ONE 2011, 6, e28142. [Google Scholar] [CrossRef]
- Hill, M.N.; Kumar, S.A.; Filipski, S.B.; Iverson, M.; Stuhr, K.L.; Keith, J.M.; Cravatt, B.F.; Hillard, C.J.; Chattarji, S.; McEwen, B.S. Disruption of fatty acid amide hydrolase activity prevents the effects of chronic stress on anxiety and amygdalar microstructure. Mol. Psychiatry 2013, 18, 1125–1135. [Google Scholar] [CrossRef]
- Bluett, R.J.; Gamble-George, J.C.; Hermanson, D.J.; Hartley, N.D.; Marnett, L.J.; Patel, S. Central anandamide deficiency predicts stress-induced anxiety: Behavioral reversal through endocannabinoid augmentation. Transl. Psychiatry 2014, 4, e408. [Google Scholar] [CrossRef]
- Burman, M.A.; Szolusha, K.; Bind, R.; Kerney, K.; Boger, D.L.; Bilsky, E.J. FAAH inhibitor OL-135 disrupts con-textual, but not auditory, fear conditioning in rats. Behav. Brain Res. 2016, 308, 1–5. [Google Scholar] [CrossRef]
- Bedse, G.; Bluett, R.J.; Patrick, T.A.; Romness, N.K.; Gaulden, A.D.; Kingsley, P.J.; Plath, N.; Marnett, L.J.; Patel, S. Therapeutic endocannabinoid augmentation for mood and anxiety disorders: Comparative profiling of FAAH, MAGL and dual inhibitors. Transl. Psychiatry 2018, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Berardi, A.; Colucci, P.; Palmery, M.; Trezza, V.; Hill, M.N.; Campolongo, P. Enhancing En-do-cannabinoid Neurotransmission Augments the Efficacy of Extinction Training and Ameliorates Traumatic Stress-Induced Behavioral Alterations in Rats. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2018, 43, 1284–1296. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.; Pavon, F.J.; Buczynski, M.W.; Schlosburg, J.; Natividad, L.A.; Polis, I.Y.; Stouffer, D.G.; Zorrilla, E.P.; Roberto, M.; Cravatt, B.F.; et al. Deficient endocanna-binoid signaling in the central amygdala contributes to alcohol dependence-related anxiety-like behavior and excessive alcohol intake. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2018, 43, 1840–1850. [Google Scholar] [CrossRef] [PubMed]
- Segev, A.; Korem, N.; Mizrachi Zer-Aviv, T.; Abush, H.; Lange, R.; Sauber, G.; Hillard, C.J.; Akirav, I. Role of endocannabinoids in the hippocampus and amygdala in emotional memory and plasticity. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2018, 43, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Mizrachi Zer-Aviv, T.; Islami, L.; Hamilton, P.J.; Parise, E.M.; Nestler, E.J.; Sbarski, B.; Akirav, I. Enhancing En-docannabinoid Signaling via β-Catenin in the Nucleus Accumbens Attenuates PTSD- and Depression-like Behavior of Male Rats. Biomedicines 2022, 10, 1789. [Google Scholar] [CrossRef] [PubMed]
- Sciolino, N.R.; Zhou, W.; Hohmann, A.G. Enhancement of endocannabinoid signaling with JZL184, an inhibitor of the 2-arachidonoylglycerol hydrolyzing enzyme monoacylglycerol lipase, produces anxiolytic effects under conditions of high environmental aversiveness in rats. Pharmacol. Res. 2011, 64, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Aliczki, M.; Zelena, D.; Mikics, E.; Varga, Z.K.; Pinter, O.; Bakos, N.V.; Varga, J.; Haller, J. Monoacylglycerol lipase inhibition-induced changes in plasma corticosterone levels, anxiety and locomotor activity in male CD1 mice. Horm. Behav. 2013, 63, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Bluett, R.J.; Báldi, R.; Haymer, A.; Gaulden, A.D.; Hartley, N.D.; Parrish, W.P.; Baechle, J.; Marcus, D.J.; Mardam-Bey, R.; Shonesy, B.C.; et al. Endocannabinoid signalling modulates susceptibility to traumatic stress exposure. Nat. Commun. 2017, 8, 14782. [Google Scholar] [CrossRef]
- Gamage, T.F.; Ignatowska-Jankowska, B.M.; Muldoon, P.P.; Cravatt, B.F.; Damaj, M.I.; Lichtman, A.H. Differ-ential effects of endocannabinoid catabolic inhibitors on morphine withdrawal in mice. Drug Alcohol Depend. 2015, 146, 7–16. [Google Scholar] [CrossRef]
- Gregus, A.M.; Buczynski, M.W. Druggable Targets in Endocannabinoid Signaling. Adv. Exp. Med. Biol. 2020, 1274, 177–201. [Google Scholar] [CrossRef] [PubMed]
- Beltramo, M.; Stella, N.; Calignano, A.; Lin, S.Y.; Makriyannis, A.; Piomelli, D. Functional role of high-affinity anandamide transport, as revealed by selective inhibition. Science 1997, 277, 1094–1097. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Fontana, A.; Cadas, H.; Schinelli, S.; Cimino, G.; Schwartz, J.C.; Piomelli, D. Formation and inacti-vation of endogenous cannabinoid anandamide in central neurons. Nature 1994, 372, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Kaczocha, M.; Haj-Dahmane, S. Mechanisms of endocannabinoid transport in the brain. Br. J. Pharmacol. 2022, 179, 4300–4310. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, A.; Beltramo, M.; Piomelli, D. Mechanisms of endocannabinoid inactivation: Biochemistry and pharmacology. J. Pharmacol. Exp. Ther. 2001, 298, 7–14. [Google Scholar] [PubMed]
- Metna-Laurent, M.; Soria-Gómez, E.; Verrier, D.; Conforzi, M.; Jégo, P.; Lafenêtre, P.; Marsicano, G. Bimodal control of fear-coping strategies by CB1 cannabinoid receptors. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 7109–7118. [Google Scholar] [CrossRef] [PubMed]
- Vickstrom, C.R.; Liu, X.; Liu, S.; Hu, M.M.; Mu, L.; Hu, Y.; Yu, H.; Love, S.L.; Hillard, C.J.; Liu, Q.S. Role of endocannabinoid signaling in a septohabenular pathway in the regulation of anxiety- and depressive-like behavior. Mol. Psychiatry 2021, 26, 3178–3191. [Google Scholar] [CrossRef]
- Han, X.; Liang, Y.; Hempel, B.; Jordan, C.J.; Shen, H.; Bi, G.H.; Li, J.; Xi, Z.X. Cannabinoid CB1 Receptors Are Expressed in a Subset of Dopamine Neurons and Underlie Cannabinoid-Induced Aversion, Hypoactivity, and Anxiolytic Ef-fects in Mice. J. Neurosci. Off. J. Soc. Neurosci. 2023, 43, 373–385. [Google Scholar] [CrossRef]
- Canseco-Alba, A.; Sanabria, B.; Hammouda, M.; Bernadin, R.; Mina, M.; Liu, Q.R.; Onaivi, E.S. Cell-Type Specific Deletion of CB2 Cannabinoid Receptors in Dopamine Neurons Induced Hyperactivity Phenotype: Possible Rele-vance to At-tention-Deficit Hyperactivity Disorder. Front. Psychiatry 2022, 12, 803394. [Google Scholar] [CrossRef]
- Liu, Q.R.; Canseco-Alba, A.; Zhang, H.Y.; Tagliaferro, P.; Chung, M.; Dennis, E.; Sanabria, B.; Schanz, N.; Esco-steguy-Neto, J.C.; Ishiguro, H.; et al. Cannabinoid type 2 receptors in dopamine neurons inhibits psychomotor behaviors, alters anxiety, depression and alcohol preference. Sci. Rep. 2017, 7, 17410. [Google Scholar] [CrossRef]
- Lazary, J.; Lazary, A.; Gonda, X.; Benko, A.; Molnar, E.; Hunyady, L.; Juhasz, G.; Bagdy, G. Promoter variants of the cannabinoid receptor 1 gene (CNR1) in interaction with 5-HTTLPR affect the anxious phenotype. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. Off. Publ. Int. Soc. Psychiatr. Genet. 2009, 150, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Heitland, I.; Klumpers, F.; Oosting, R.S.; Evers, D.J.; Leon Kenemans, J.; Baas, J.M. Failure to extinguish fear and genetic variability in the human cannabinoid receptor 1. Transl. Psychiatry 2012, 2, e162. [Google Scholar] [CrossRef] [PubMed]
- Peiró, A.M.; García-Gutiérrez, M.S.; Planelles, B.; Femenía, T.; Mingote, C.; Jiménez-Treviño, L.; Martínez-Barrondo, S.; García-Portilla, M.P.; Saiz, P.A.; Bobes, J.; et al. Association of cannabinoid receptor genes (CNR1 and CNR2) polymorphisms and panic disorder. Anxiety Stress Coping 2020, 33, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Abood, M.; Alexander, S.P.; Barth, F.; Bonner, T.I.; Bradshaw, H.; Cabral, G.; Casellas, P.; Cravatt, B.F.; Devane, W.A.; Di Marzo, V.; et al. Cannabinoid receptors in GtoPdb v.2023.1. IUPHAR/BPS Guide Pharmacol. CITE 2023, 2023. [Google Scholar] [CrossRef]
- Haller, J.; Rompos, É.; Szabó, Í.; Humli, V.; Christián, L. Drug regulations and trafficking: Synthetic cannabinoids and cathinones in Hungary. Forensic Sci. Int. 2023, 349, 111778. [Google Scholar] [CrossRef] [PubMed]
- Gambi, F.; De Berardis, D.; Sepede, G.; Quartesan, R.; Calcagni, E.; Salerno, R.M.; Conti, C.M.; Ferro, F.M. Can-nabinoid receptors and their relationships with neuropsychiatric disorders. Int. J. Immunopathol. Pharmacol. 2005, 18, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, B.; Forget, B.; Aubin, H.J.; Goldberg, S.R. Blocking cannabinoid CB1 receptors for the treatment of nicotine dependence: Insights from pre-clinical and clinical studies. Addict. Biol. 2008, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Seely, K.A.; Brents, L.K.; Franks, L.N.; Rajasekaran, M.; Zimmerman, S.M.; Fantegrossi, W.E.; Prather, P.L. AM-251 and rimonabant act as direct antagonists at mu-opioid receptors: Implications for opioid/cannabinoid interaction studies. Neuropharmacology 2012, 63, 905–915. [Google Scholar] [CrossRef]
- Sharir, H.; Abood, M.E. Pharmacological characterization of GPR55, a putative cannabinoid receptor. Pharmacol. Ther. 2010, 126, 301–313. [Google Scholar] [CrossRef]
- Pulgar, V.M.; Howlett, A.C.; Eldeeb, K. WIN55212-2 Modulates Intracellular Calcium via CB1 Recep-tor-Dependent and Independent Mechanisms in Neuroblastoma Cells. Cells 2022, 11, 2947. [Google Scholar] [CrossRef]
- Barna, I.; Till, I.; Haller, J. Blood, adipose tissue and brain levels of the cannabinoid ligands WIN-55,212 and SR-141716A after their intraperitoneal injection in mice: Compound-specific and area-specific distribution within the brain. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2009, 19, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Hillard, C.J. Endocannabinoids and the Endocrine System in Health and Disease. Handb. Exp. Pharmacol. 2015, 231, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Strobel, C.; Hunt, S.; Sullivan, R.; Sun, J.; Sah, P. Emotional regulation of pain: The role of noradrenaline in the amygdala. Sci. China Life Sci. 2014, 57, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Hogri, R.; Teuchmann, H.L.; Heinke, B.; Holzinger, R.; Trofimova, L.; Sandkühler, J. GABAergic CaMKIIα+ Amygdala Output Attenuates Pain and Modulates Emotional-Motivational Behavior via Parabrachial Inhibition. J. Neurosci. Off. J. Soc. Neurosci. 2022, 42, 5373–5388. [Google Scholar] [CrossRef] [PubMed]
- Jurik, A.; Ressle, A.; Schmid, R.M.; Wotjak, C.T.; Thoeringer, C.K. Supraspinal TRPV1 modulates the emotional expression of abdominal pain. Pain 2014, 155, 2153–2160. [Google Scholar] [CrossRef] [PubMed]
- Sokolaj, E.; Assareh, N.; Anderson, K.; Aubrey, K.R.; Vaughan, C.W. Cannabis constituents for chronic neuropathic pain; reconciling the clinical and animal evidence. J. Neurochem. 2023; early view. [Google Scholar] [CrossRef]
- Zvolensky, M.J.; Bernstein, A.; Sachs-Ericsson, N.; Schmidt, N.B.; Buckner, J.D.; Bonn-Miller, M.O. Lifetime associations be-tween cannabis, use, abuse, and dependence and panic attacks in a representative sample. J. Psychiatr. Res. 2006, 40, 477–486. [Google Scholar] [CrossRef]
- Rifkin-Zybutz, R.; Erridge, S.; Holvey, C.; Coomber, R.; Gaffney, J.; Lawn, W.; Barros, D.; Bhoskar, U.; Mwimba, G.; Praveen, K.; et al. Clinical outcome data of anxiety patients treated with cannabis-based medicinal products in the United Kingdom: A cohort study from the UK Medical Cannabis Registry. Psychopharmacology 2023, 240, 1735–1745. [Google Scholar] [CrossRef]
- Hourfane, S.; Mechqoq, H.; Bekkali, A.Y.; Rocha, J.M.; El Aouad, N. A Comprehensive Review on Cannabis sativa Ethnobotany, Phytochemistry, Molecular Docking and Biological Activities. Plants 2023, 12, 1245. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Morrison, P.D.; Fusar-Poli, P.; Martin-Santos, R.; Borgwardt, S.; Winton-Brown, T.; Nosarti, C.; O’Carroll, C.M.; Seal, M.; Allen, P.; et al. Opposite effects of delta-9-tetrahydrocannabinol and cannabidiol on human brain function and psychopathology. Neuropsychopharmacology 2010, 35, 764–774. [Google Scholar] [CrossRef]
- Madras, B.K. Tinkering with THC-to-CBD ratios in Marijuana. Neuropsychopharmacology 2019, 44, 215–216. [Google Scholar] [CrossRef]
- Sharpe, L.; Sinclair, J.; Kramer, A.; de Manincor, M.; Sarris, J. Cannabis, a cause for anxiety? A critical appraisal of the anxiogenic and anxiolytic properties. J. Transl. Med. 2020, 18, 374. [Google Scholar] [CrossRef]
- Englund, A.; Freeman, T.P.; Murray, R.M.; McGuire, P. Can we make cannabis safer? Lancet Psychiatry 2017, 4, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Karschner, E.L.; Darwin, W.D.; McMahon, R.P.; Liu, F.; Wright, S.; Goodwin, R.S.; Huestis, M.A. Subjective and physiological effects after controlled Sativex and oral THC administration. Clin. Pharmacol. Ther. 2011, 89, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Zuardi, A.W.; Shirakawa, I.; Finkelfarb, E.; Karniol, I.G. Action of cannabidiol on the anxiety and other effects produced by delta 9-THC in normal subjects. Psychopharmacology 1982, 76, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Naveed, S.; Mian, N.; Fida, A.; Raafey, M.A.; Aedma, K.K. The therapeutic role of Cannabidiol in mental health: A systematic review. J. Cannabis Res. 2020, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Hutten, N.R.P.W.; Arkell, T.R.; Vinckenbosch, F.; Schepers, J.; Kevin, R.C.; Theunissen, E.L.; Kuypers, K.P.C.; McGregor, I.S.; Ramaekers, J.G. Cannabis containing equivalent concentrations of delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD) induces less state anxiety than THC-dominant cannabis. Psychopharmacology 2022, 239, 3731–3741. [Google Scholar] [CrossRef]
- Patton, G.C.; Coffey, C.; Carlin, J.B.; Degenhardt, L.; Lynskey, M.; Hall, W. Cannabis use and mental health in young people: Cohort study. BMJ 2002, 325, 1195–1198. [Google Scholar] [CrossRef]
- Zvolensky, M.J.; Lewinsohn, P.; Bernstein, A.; Schmidt, N.B.; Buckner, J.D.; Seeley, J.; Bonn-Miller, M.O. Prospective associations between cannabis use, abuse, and dependence and panic attacks and disorder. J. Psychiatr. Res. 2008, 42, 1017–1023. [Google Scholar] [CrossRef]
- Duperrouzel, J.; Hawes, S.W.; Lopez-Quintero, C.; Pacheco-Colón, I.; Comer, J.; Gonzalez, R. The association be-tween adolescent cannabis use and anxiety: A parallel process analysis. Addict. Behav. 2018, 78, 107–113. [Google Scholar] [CrossRef]
- Allsop, D.J.; Copeland, J.; Lintzeris, N.; Dunlop, A.J.; Montebello, M.; Sadler, C.; Rivas, G.R.; Holland, R.M.; Muhleisen, P.; Norberg, M.M.; et al. Nabiximols as an agonist replacement therapy during cannabis withdrawal: A randomized clinical trial. JAMA Psychiatry 2014, 71, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Crippa, J.A.; Hallak, J.E.; Machado-de-Sousa, J.P.; Queiroz, R.H.; Bergamaschi, M.; Chagas, M.H.; Zuardi, A.W. Cannabidiol for the treatment of cannabis withdrawal syndrome: A case report. J. Clin. Pharm. Ther. 2013, 38, 162–164. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Prasad, S.; Jain, E.; Dogrul, B.N.; Al-Oleimat, A.; Pokhrel, B.; Chowdhury, S.; Co, E.L.; Mitra, S.; Quinonez, J.; et al. Medical Cannabis for Chronic Nonmalignant Pain Management. Curr. Pain Headache Rep. 2023, 27, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.D.; Vitetta, L.; Hall, S. Tetrahydrocannabinol and cannabidiol medicines for chronic pain and mental health conditions. Inflammopharmacology 2022, 30, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Zieglgänsberger, W.; Brenneisen, R.; Berthele, A.; Wotjak, C.T.; Bandelow, B.; Tölle, T.R.; Lutz, B. Chronic Pain and the Endocannabinoid System: Smart Lipids—A Novel Therapeutic Option? Med. Cannabis Cannabinoids 2022, 5, 61–75. [Google Scholar] [CrossRef]
- Martinez Naya, N.; Kelly, J.; Corna, G.; Golino, M.; Abbate, A.; Toldo, S. Molecular and Cellular Mechanisms of Action of Cannabidiol. Molecules 2023, 28, 5980. [Google Scholar] [CrossRef]
- Patel, S.; Cravatt, B.F.; Hillard, C.J. Synergistic interactions between cannabinoids and environmental stress in the activation of the central amygdala. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2005, 30, 497–507. [Google Scholar] [CrossRef]
- Patel, S.; Roelke, C.T.; Rademacher, D.J.; Cullinan, W.E.; Hillard, C.J. Endocannabinoid signaling negatively modulates stress-induced activation of the hypothalamic-pituitary-adrenal axis. Endocrinology 2004, 145, 5431–5438. [Google Scholar] [CrossRef]
- Chavanne, A.V.; Robinson, O.J. The Overlapping Neurobiology of Induced and Pathological Anxiety: A Me-ta-Analysis of Functional Neural Activation. Am. J. Psychiatry 2021, 178, 156–164. [Google Scholar] [CrossRef]
- Muigg, P.; Scheiber, S.; Salchner, P.; Bunck, M.; Landgraf, R.; Singewald, N. Differential stress-induced neuronal ac-tivation patterns in mouse lines selectively bred for high, normal or low anxiety. PLoS ONE 2009, 4, e5346. [Google Scholar] [CrossRef]
- Imperatore, R.; Morello, G.; Luongo, L.; Taschler, U.; Romano, R.; De Gregorio, D.; Belardo, C.; Maione, S.; Di Marzo, V.; Cristino, L. Genetic deletion of monoacylglycerol lipase leads to impaired cannabinoid receptor CB1R signaling and anxiety-like behavior. J. Neurochem. 2015, 135, 799–813. [Google Scholar] [CrossRef] [PubMed]
- Schlosburg, J.E.; Blankman, J.L.; Long, J.Z.; Nomura, D.K.; Pan, B.; Kinsey, S.G.; Nguyen, P.T.; Ramesh, D.; Booker, L.; Burston, J.J.; et al. Chronic mon-oacylglycerol lipase blockade causes functional antagonism of the endocannabinoid system. Nat. Neurosci. 2010, 13, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Leishman, E.; Cornett, B.; Spork, K.; Straiker, A.; Mackie, K.; Bradshaw, H.B. Broad impact of deleting endogenous cannabinoid hydrolyzing enzymes and the CB1 cannabinoid receptor on the endogenous cannabinoid-related lipidome in eight regions of the mouse brain. Pharmacol. Res. 2016, 110, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Schlosburg, J.E.; Kinsey, S.G.; Ignatowska-Jankowska, B.; Ramesh, D.; Abdullah, R.A.; Tao, Q.; Booker, L.; Long, J.Z.; Selley, D.E.; Cravatt, B.F.; et al. Prolonged monoacylglycerol lipase blockade causes equivalent cannabinoid receptor type 1 receptor-mediated adaptations in fatty acid amide hydrolase wild-type and knockout mice. J. Pharmacol. Exp. Ther. 2014, 350, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Micale, V.; Stepan, J.; Jurik, A.; Pamplona, F.A.; Marsch, R.; Drago, F.; Eder, M.; Wotjak, C.T. Extinction of avoidance behavior by safety learning depends on endocannabinoid signaling in the hippocampus. J. Psychiatr. Res. 2017, 90, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Bortolato, M.; Campolongo, P.; Mangieri, R.A.; Scattoni, M.L.; Frau, R.; Trezza, V.; La Rana, G.; Russo, R.; Calignano, A.; Gessa, G.L.; et al. Anxiolytic-like properties of the anandamide transport inhibitor AM404. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2006, 31, 2652–2659. [Google Scholar] [CrossRef] [PubMed]
- Braida, D.; Limonta, V.; Malabarba, L.; Zani, A.; Sala, M. 5-HT1A receptors are involved in the anxiolytic effect of Delta9-tetrahydrocannabinol and AM 404, the anandamide transport inhibitor, in Sprague-Dawley rats. Eur. J. Pharmacol. 2007, 555, 156–163. [Google Scholar] [CrossRef]
- Lisboa, S.F.; Resstel, L.B.; Aguiar, D.C.; Guimarães, F.S. Activation of cannabinoid CB1 receptors in the dorsolateral periaqueductal gray induces anxiolytic effects in rats submitted to the Vogel conflict test. Eur. J. Pharmacol. 2008, 593, 73–78. [Google Scholar] [CrossRef]
- Resstel, L.B.; Lisboa, S.F.; Aguiar, D.C.; Corrêa, F.M.; Guimarães, F.S. Activation of CB1 cannabinoid receptors in the dorsolateral periaqueductal gray reduces the expression of contextual fear conditioning in rats. Psychopharmacology 2008, 198, 405–411. [Google Scholar] [CrossRef]
- Campos, A.C.; Ferreira, F.R.; Guimarães, F.S.; Lemos, J.I. Facilitation of endocannabinoid effects in the ventral hippocampus modulates anxiety-like behaviors depending on previous stress experience. Neuroscience 2010, 167, 238–246. [Google Scholar] [CrossRef]
- Hill, M.N.; Campolongo, P.; Yehuda, R.; Patel, S. Integrating Endocannabinoid Signaling and Cannabinoids into the Biology and Treatment of Posttraumatic Stress Disorder. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2018, 43, 80–102. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.; Rauf, K.; Zada, W.; McCarthy, M.; Abbas, G.; Anwar, F.; Shah, A.J. Kaempferol Facilitated Ex-tinction Learning in Contextual Fear Conditioned Rats via Inhibition of Fatty-Acid Amide Hydrolase. Molecules 2020, 25, 4683. [Google Scholar] [CrossRef] [PubMed]
- Danandeh, A.; Vozella, V.; Lim, J.; Oveisi, F.; Ramirez, G.L.; Mears, D.; Wynn, G.; Piomelli, D. Effects of fatty acid amide hydrolase inhibitor URB597 in a rat model of trauma-induced long-term anxiety. Psychopharmacology 2018, 235, 3211–3221. [Google Scholar] [CrossRef] [PubMed]
- Fotio, Y.; Mabou Tagne, A.; Jung, K.M.; Piomelli, D. Fatty acid amide hydrolase inhibition alleviates anxiety-like symptoms in a rat model used to study post-traumatic stress disorder. Psychopharmacology, 2023; early view. [Google Scholar] [CrossRef] [PubMed]
- Chhatwal, J.P.; Davis, M.; Maguschak, K.A.; Ressler, K.J. Enhancing cannabinoid neurotransmission augments the extinction of conditioned fear. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2005, 30, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Spohrs, J.; Ulrich, M.; Grön, G.; Plener, P.L.; Abler, B. FAAH polymor-phism (rs324420) modulates extinction recall in healthy humans: An fMRI study. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Paulus, M.P.; Stein, M.B.; Simmons, A.N.; Risbrough, V.B.; Halter, R.; Chaplan, S.R. The effects of FAAH inhibition on the neural basis of anxiety-related pro-cessing in healthy male subjects: A randomized clinical trial. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2021, 46, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Mayo, L.M.; Asratian, A.; Lindé, J.; Morena, M.; Haataja, R.; Hammar, V.; Augier, G.; Hill, M.N.; Heilig, M. Elevated Anandamide, Enhanced Recall of Fear Ex-tinction, and Attenuated Stress Responses Following Inhibition of Fatty Acid Amide Hydrolase: A Randomized, Controlled Experimental Medicine Trial. Biol. Psychiatry 2020, 87, 538–547. [Google Scholar] [CrossRef]
- Bourke, S.L.; Schlag, A.K.; O’Sullivan, S.E.; Nutt, D.J.; Finn, D.P. Cannabinoids and the endocannabinoid system in fibromyalgia: A review of preclinical and clinical research. Pharmacol. Ther. 2022, 240, 108216. [Google Scholar] [CrossRef]
- Petrosino, S.; Di Marzo, V. FAAH and MAGL inhibitors: Therapeutic opportunities from regulating endocannabinoid levels. Curr. Opin. Investig. Drugs 2010, 11, 51–62. [Google Scholar]
- McLaughlin, R.J.; Hill, M.N.; Bambico, F.R.; Stuhr, K.L.; Gobbi, G.; Hillard, C.J.; Gorzalka, B.B. Prefrontal cortical anandamide signaling coordinates coping responses to stress through a serotonergic pathway. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2012, 22, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M. Coping style and immunity in animals: Making sense of individual variation. Brain Behav. Immun. 2008, 22, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Price, R.H.; Wortman, C.B. Social factors in psychopathology: Stress, social support, and coping processes. Annu. Rev. Psychol. 1985, 36, 531–572. [Google Scholar] [CrossRef] [PubMed]
- Temoshok, L.R. Complex coping patterns and their role in adaptation and neuroimmunomodulation. Theory, meth-odology, and research. Ann. N. Y. Acad. Sci. 2000, 917, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Goldberg, S.R.; Pelczer, K.G.; Aliczki, M.; Panlilio, L.V. The effects of anandamide signaling enhanced by the FAAH inhibitor URB597 on coping styles in rats. Psychopharmacology 2013, 230, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, O.; Lecca, D.; Piras, G.; Driscoll, P.; Corda, M.G. Dissociation between mesocortical dopamine release and fear-related behaviours in two psychogenetically selected lines of rats that differ in coping strategies to aversive conditions. Eur. J. Neurosci. 2003, 17, 2716–2726. [Google Scholar] [CrossRef]
- Haller, J.; Aliczki, M.; Pelczer, K.G.; Spitzer, K.; Balogh, Z.; Kantor, S. Effects of the fatty acid amide hydrolase inhibitor URB597 on coping behavior under challenging conditions in mice. Psychopharmacology 2014, 231, 593–601. [Google Scholar] [CrossRef]
- Hawley, D.F.; Bardi, M.; Everette, A.M.; Higgins, T.J.; Tu, K.M.; Kinsley, C.H.; Lambert, K.G. Neurobiological constituents of active, passive, and variable coping strategies in rats: Integration of regional brain neuropeptide Y levels and cardiovascular responses. Stress 2010, 13, 172–183. [Google Scholar] [CrossRef]
- Gunduz-Cinar, O.; MacPherson, K.P.; Cinar, R.; Gamble-George, J.; Sugden, K.; Williams, B.; Godlewski, G.; Ramikie, T.S.; Gorka, A.X.; Alapafuja, S.O.; et al. Convergent translational evidence of a role for anandamide in amygdala-mediated fear extinction, threat processing and stress-reactivity. Mol. Psychiatry 2013, 18, 813–823. [Google Scholar] [CrossRef]
- Hill, M.N.; McEwen, B.S. Involvement of the endocannabinoid system in the neurobehavioural effects of stress and glucocorticoids. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 791–797. [Google Scholar] [CrossRef]
- Lutz, B.; Marsicano, G.; Maldonado, R.; Hillard, C.J. The endocannabinoid system in guarding against fear, anxiety and stress. Nat. Rev. Neurosci. 2015, 16, 705–718. [Google Scholar] [CrossRef]
- Aliczki, M.; Haller, J. Interactions Between Cannabinoid Signaling and Anxiety: A Comparative Analysis of In-tervention Tools and Behavioral Effects. In Cannabinoid Modulation of Emotion, Memory, and Motivation; Campolongo, P., Fattore, L., Eds.; Springer: New York, NY, USA, 2015. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CB1-KO | |||
| Background (sex) | Effects on anxiety | Anxiety test | Reference |
| CD1 (♂) | ↑ | LD | [47] |
| CD1 (♂) | ↑ | EPM | [48] |
| CD1 (♂) | ↑ | EPM | [49] |
| CD1 (♂) | ↑ (increased stress response) | EPM, LD, SI | [50] |
| CD1 (♂) | ↑ (high light) | EPM | [51] |
| C57BL/6NCrl (♂) | ↑ | EPM, LD | [52] |
| CD1 (♂) | ↑ (0.7, 1.5 mA) | CF | [53] |
| ICR (♂) | ↑ (chronic stress-like phenotype) | EPM | [54] |
| CD1 (♂) | ↑ (increased stress response) | EPM | [55] |
| CD1 (♂) | ↑ (chronic stress) | CF | [56] |
| C57/BL6J (♂) | ↑ | SI | [57] |
| C57BL/6J (♂) | ↑ | EPM, LD | [58] |
| C57BL/6J (♂) | ↑ | EPM, LD | [59] |
| C57BL/6N (♂, ♀) | ↑ | SI | [60] |
| CD1 (♂) | ↑ (young) | OF, LD | [61] |
| C57BL/6J (♂, ♀) | ↑ (chronic pain) | EZM, LD | [62] |
| C57BL/6J (♂) | ↑ (male) | EPM | [63] |
| CD1 (♂) | → (old) | OF, LD | [61] |
| CD1 (♂) | → (low light) | EPM | [51] |
| CD1 (♂) | → | EPM, OF | [64] |
| CD1 (♂) | → (unstressed) | CF | [56] |
| CD1 (♂) | → (0.5 mA) | CF | [53] |
| CD1 (♂) | → | SI | [65] |
| C57BL/6J (♂, ♀) | → (control) | EZM, LD | [62] |
| C57BL/6J (♀) | → (female) | EPM | [63] |
| C57BL⁄6 (♂) | ↓ | SPBT | [66] |
| CB2-KO | |||
| Background | Effects on anxiety | Anxiety test | Reference |
| C57BL/6J (♂) | ↑ | EPM, LD | [67] |
| C57BL/J6 (♀) | ↑ | OF | [68] |
| C57BL/6J (♂, ♀) | → | EZM, OF | [69] |
| Gene Overexpression | |||
| Receptor | Effects on anxiety | Anxiety test | Reference |
| CB1 * (♂) | ↑ | SI | [45] |
| CB2 (♂) | ↓ | EPM, LD | [70] |
| The Effects of CB1 Antagonists Is Conflicting | |||
| Antagonist | Effect on Anxiety | Effective Dose (Range) | Reference |
| Rimonabant | anxiogenesis (EPM, ETM, OF) | 3 (1–3) mg/kg | [64] |
| anxiolysis (EPM) | 10 (1–10) mg/kg | [75] | |
| anxiogenesis (EPM, DW) | 3 (0.1–3) mg/kg | [76] | |
| anxiolysis (EPM) | 3 (1–3) mg/kg | [48] | |
| AM-281 | no effect (LD) | none (1–4) mg/kg | [77] |
| AM4113 | no effect (EPM) | none (3.0–12.0) mg/kg | [78] |
| AVE1625 | no effect (LD) | none (10–100) mg/kg | [79] |
| Agonists and Antagonists May Have Similar Effects | |||
| Ligand | Effect on Anxiety | Effective Dose (Range) | Reference |
| Δ9-THC | anxiogenesis (EPM) | 1, 2.5, 10 (0.25–10) mg/kg | [80] |
| Rimonabant | anxiogenesis (EPM) | 3, 10 (1–10) mg/kg | |
| AM-251 | anxiogenesis (EPM) | 3, 10 (1–10) mg/kg | |
| Agonists Have Biphasic Effects | |||
| Ligand | Effect on Anxiety | Effective Dose (Range) | Reference |
| HU-210 | anxiolysis (EPM) | 10 μg/kg | [81] |
| anxiogenesis (EPM) | 50 μg/kg | ||
| CP 55,940 | anxiolysis (EPM) | 1 μg/kg | [82] |
| anxiogenesis (EPM) | 50 μg/kg | ||
| Δ9-THC | anxiolysis (LD, OF) | 0.2 mg/kg | [83] |
| anxiogenesis (LD, OF) | 7.5 mg/kg | ||
| A. Early Studies with URB-597 | |||
| Effective dose (range) | Species (test; sex) | Effects on anxiety | Reference |
| 0.1 (0.05–0.1) mg/kg | rat (EZM; ♂) | ↓ | [97] |
| 0.1, 0.3 (0.03–0.3) mg/kg | mouse (EPM; ♂) | ↓ | [80] |
| 0.1, 0.3 (0.1–0.3) mg/kg | mouse (EPM; ♀) | ↓ | [102] |
| 0.3, 1 (0.3–1) mg/kg | rat (EPM; ♂) | ↓ | [103] |
| 1 (1) mg/kg | mouse (EPM; ♂) | ↓ | [104] |
| 0.1, 0.3 (0.1–0.3) mg/kg | rat (LD; ♂) | ↓ | [105] |
| none (0.1–10) mg/kg | mouse (EPM; ♂, ♀) | → | [106] |
| none (0.03–0.3) mg/kg | mouse (EPM; ♂) | → | [107] |
| 1 (1) mg/kg | mouse (LD; ♂) | → | [104] |
| B. FAAH Inhibition Ameliorates Stress-Induced Anxiety | |||
| FAAH inhibitor | Species | Stress factor | Reference |
| URB-597 | mouse (EPM; ♂, ♀) | light contrast * | [106] |
| URB-597 | rat (EPM; ♂) | test aversiveness | [108] |
| URB-597 | rat (EPM, SPBT; ♂) | nicotine withdrawal | [109] |
| JNJ-5003 | mouse (EPM; ♂) | chronic stress | [110] |
| PF-3845 | mouse (LD, NH; ♂) | contextual fear | [111] |
| OL-135 | rat (FRE; ♂) | contextual fear | [112] |
| PF-3845 | rat (LD; ♂) | chronic stress | [113] |
| URB-597 | rat (FRE, SI; ♂) | contextual fear | [114] |
| PF-3845 | rat (EPM; ♂) | alcohol withdrawal | [115] |
| URB-597 | rat (IA; ♂) | contextual fear | [116] |
| URB-597 | rat (FRE, ASR; ♂) | contextual fear | [117] |
| C. MAGL Inhibition Ameliorates Stress-Induced Anxiety | |||
| MAGL inhibitor | Species | Stress factor | Reference |
| JZL-184 | rat (EPM; ♂) | test aversiveness | [118] |
| JZL-184 | mouse (EPM; ♂) | hormone manipulation | [119] |
| JZL-184 | mouse (LD, NH; ♂, ♀) | Susceptibility † | [120] |
| JZL-184 | rat (LD; ♂) | chronic stress | [113] |
| JZL-184 | rat (FRE, SI; ♂) | contextual fear | [114] |
| JZL-184 | rat (EPM; ♂) | alcohol withdrawal | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haller, J. Anxiety Modulation by Cannabinoids—The Role of Stress Responses and Coping. Int. J. Mol. Sci. 2023, 24, 15777. https://doi.org/10.3390/ijms242115777
Haller J. Anxiety Modulation by Cannabinoids—The Role of Stress Responses and Coping. International Journal of Molecular Sciences. 2023; 24(21):15777. https://doi.org/10.3390/ijms242115777
Chicago/Turabian StyleHaller, József. 2023. "Anxiety Modulation by Cannabinoids—The Role of Stress Responses and Coping" International Journal of Molecular Sciences 24, no. 21: 15777. https://doi.org/10.3390/ijms242115777
APA StyleHaller, J. (2023). Anxiety Modulation by Cannabinoids—The Role of Stress Responses and Coping. International Journal of Molecular Sciences, 24(21), 15777. https://doi.org/10.3390/ijms242115777