
Therapeutic Effect and Safety Evaluation of Naringin on Klebsiella pneumoniae in Mice


Abstract
:1. Introduction
2. Results
2.1. Therapeutic Effect of NAR on Mice with Kpn
2.2. Effect of NAR on the Pathological Injury of the Lung in Mice
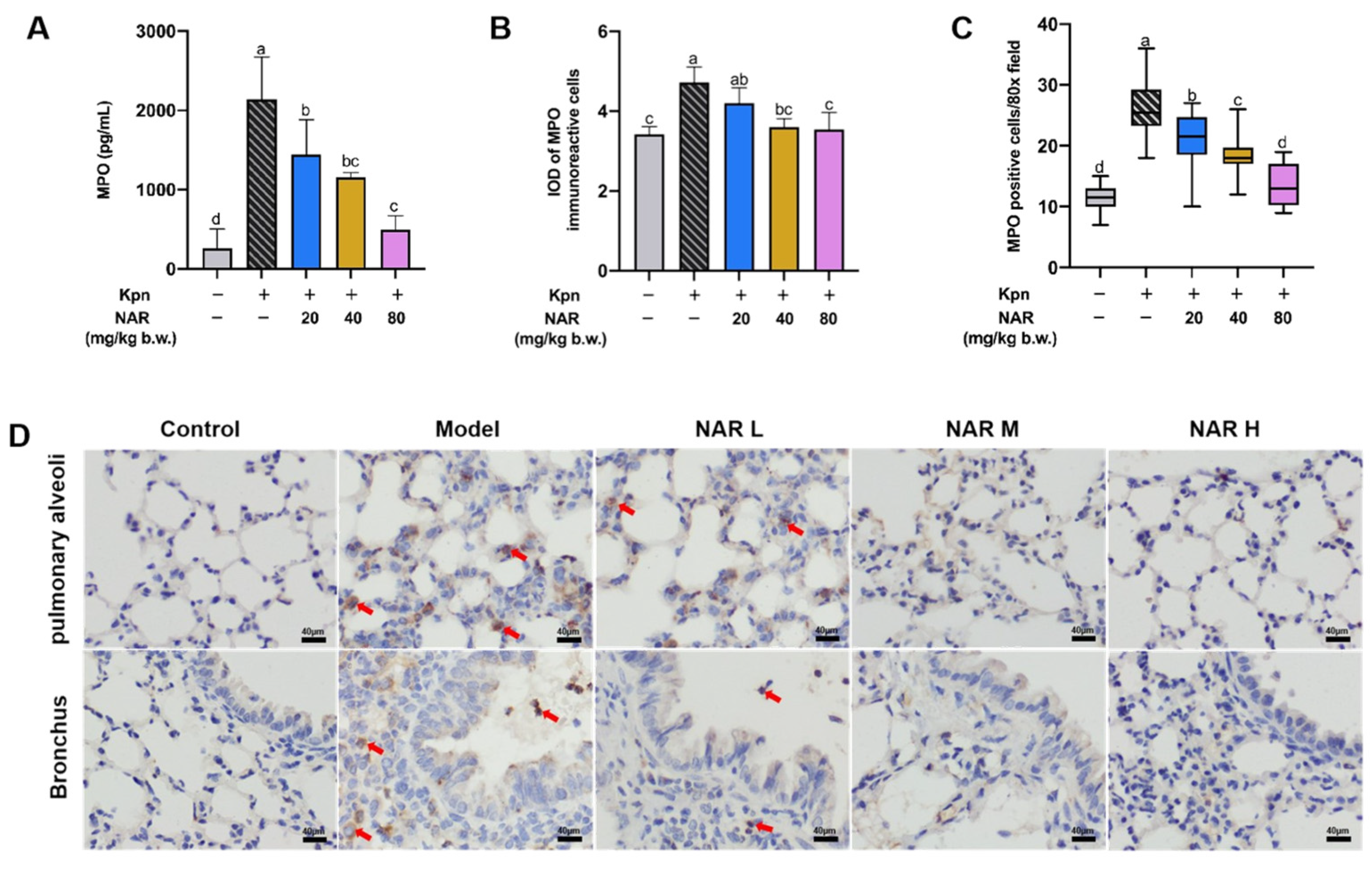
2.3. Regulation Effect of NAR on Pulmonary Inflammatory Infiltration in Mice
2.4. Regulation of NAR on the Classical Inflammatory Signaling Pathway
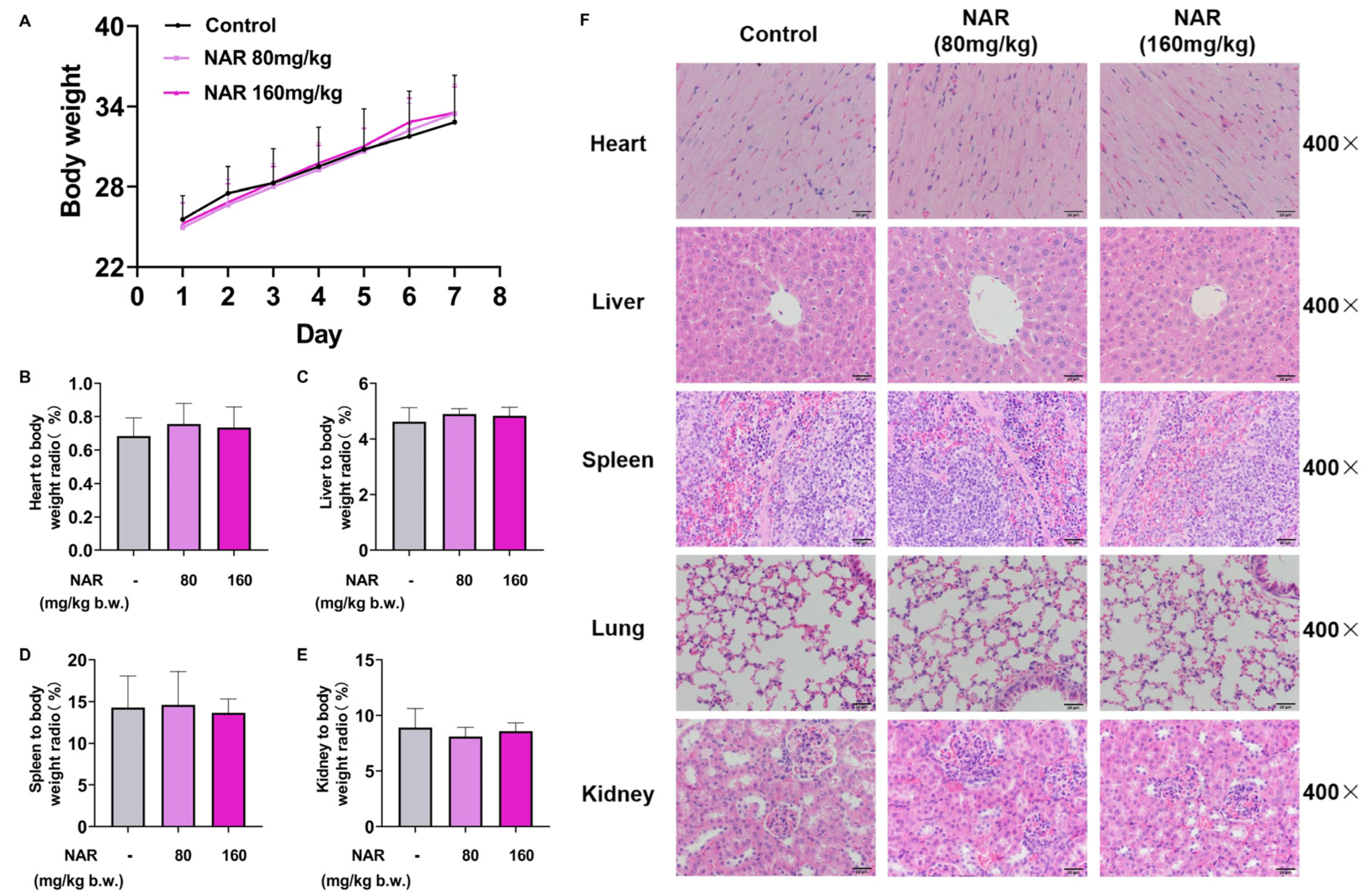
2.5. Medication Safety Evaluation of NAR
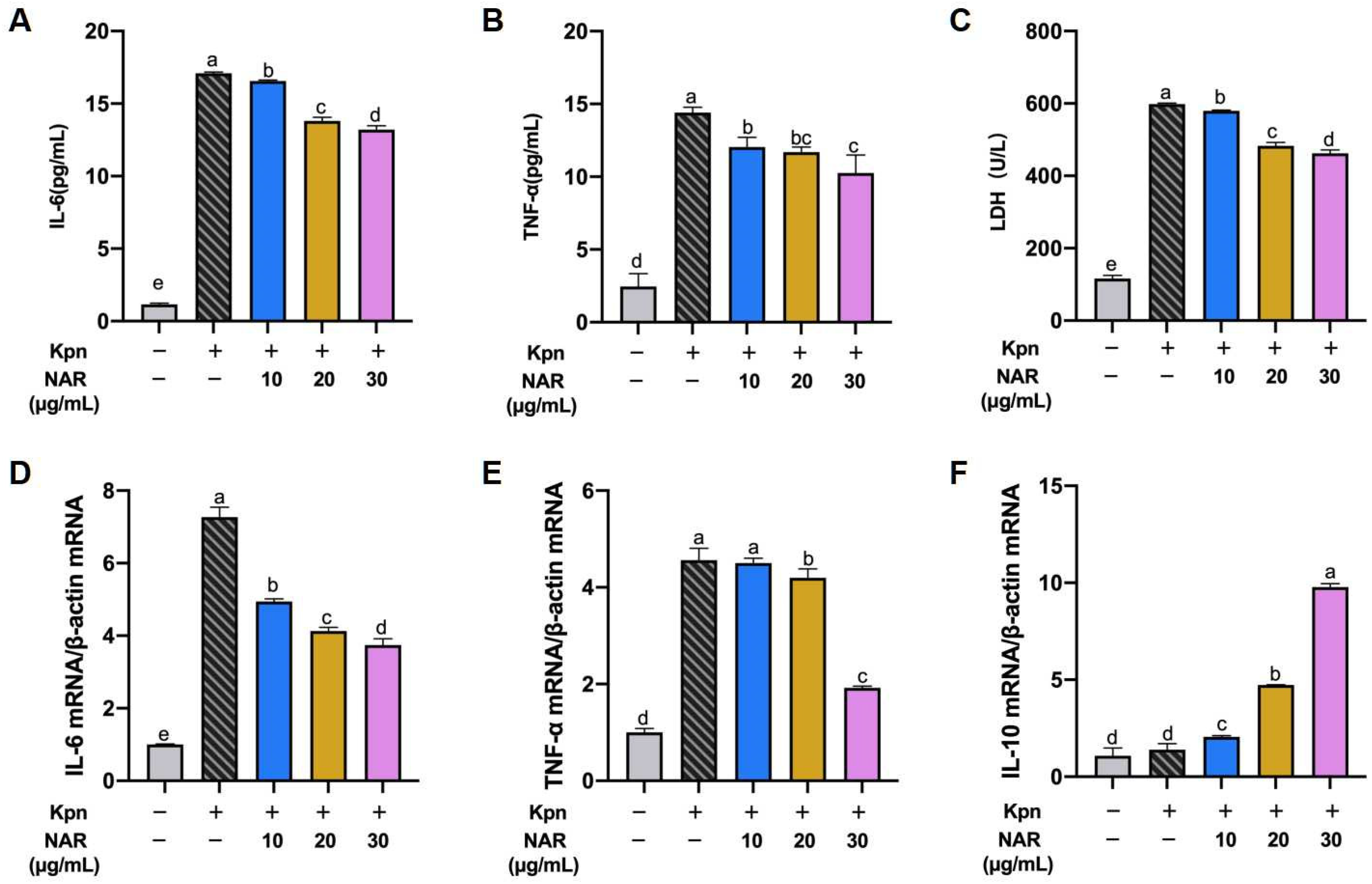
2.6. Effect of NAR on the Viability and Production of Proinflammatory Cytokines in the MH-S Cell Line
2.7. Effect of NAR on the Expression of Proinflammatory Cytokines in Inflammatory Cell Models
2.8. Regulation of the NF-κB Signaling Pathway by NAR
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Establishment of Animal Model and Treatment of NAR
4.3. In Vivo Safety Evaluation of NAR
4.4. Bacteria, Cellular Inflammation Models, and NAR Treatment
4.5. Blood Analysis
4.6. Histopathologic Examination
4.7. Immunohistochemistry (IHC) Assay
4.8. Quantitative Real-Time PCR Analysis of the Expression of Inflammatory Factors
4.9. ELISA Analysis of the Expression of Inflammatory Factors
4.10. Western Blotting Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- So, M.; Kabata, H.; Fukunaga, K.; Takagi, H.; Kuno, T. Radiological and functional lung sequelae of COVID-19: A systematic review and meta-analysis. BMC Pulm. Med. 2021, 21, 97. [Google Scholar] [CrossRef] [PubMed]
- Tanni, S.E.; Fabro, A.T.; de Albuquerque, A.; Ferreira, E.V.M.; Verrastro, C.G.Y.; Sawamura, M.V.Y.; Ribeiro, S.M.; Baldi, B.G. Pulmonary fibrosis secondary to COVID-19: A narrative review. Expert Rev. Respir. Med. 2021, 15, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Rusu, L.C.; Ardelean, L.C.; Tigmeanu, C.V.; Matichescu, A.; Sauciur, I.; Bratu, E.A. COVID-19 and Its Repercussions on Oral Health: A Review. Medicina 2021, 57, 1189. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, S. Bioactivity of naringin and related mechanisms. Die Pharm. 2021, 76, 359–363. [Google Scholar]
- Parasher, A. COVID-19: Current understanding of its Pathophysiology, Clinical presentation and Treatment. Postgrad. Med. J. 2021, 97, 312–320. [Google Scholar] [CrossRef]
- Wu, H.Y.; Chang, P.H.; Chen, K.Y.; Lin, I.F.; Hsih, W.H.; Tsai, W.L.; Chen, J.A.; Lee, S.S. Coronavirus disease 2019 (COVID-19) associated bacterial coinfection: Incidence, diagnosis and treatment. J. Microbiol. Immunol. Infect. Wei Mian Yu Gan Ran Za Zhi 2022, 55 Pt 1, 985–992. [Google Scholar] [CrossRef]
- Yang, C.P.; Liu, M.H.; Zou, W.; Guan, X.L.; Lai, L.; Su, W.W. Toxicokinetics of naringin and its metabolite naringenin after 180-day repeated oral administration in beagle dogs assayed by a rapid resolution liquid chromatography/tandem mass spectrometric method. J. Asian Nat. Prod. Res. 2012, 14, 68–75. [Google Scholar] [CrossRef]
- Liu, G.; Li, S.; Zhang, N.; Wei, N.; Wang, M.; Liu, J.; Xu, Y.; Li, Y.; Sun, Q.; Li, Y.; et al. Sequential grade evaluation method exploration of Exocarpium Citri Grandis (Huajuhong) decoction pieces based on “network prediction → grading quantization → efficacy validation”. J. Ethnopharmacol. 2022, 291, 115149. [Google Scholar] [CrossRef]
- Zhou, T.; Liu, L.; Wang, Q.; Gao, Y. Naringenin alleviates cognition deficits in high-fat diet-fed SAMP8 mice. J. Food Biochem. 2020, 44, e13375. [Google Scholar] [CrossRef]
- Sun, M.; Li, L.; Wang, C.; Wang, L.; Lu, D.; Shen, D.; Wang, J.; Jiang, C.; Cheng, L.; Pan, X.; et al. Naringenin confers defence against Phytophthora nicotianae through antimicrobial activity and induction of pathogen resistance in tobacco. Mol. Plant Pathol. 2022, 23, 1737–1750. [Google Scholar] [CrossRef]
- Nie, Y.C.; Wu, H.; Li, P.B.; Luo, Y.L.; Long, K.; Xie, L.M.; Shen, J.G.; Su, W.W. Anti-inflammatory effects of naringin in chronic pulmonary neutrophilic inflammation in cigarette smoke-exposed rats. J. Med. Food 2012, 15, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Attia, S.M.; Bakheet, S.A.; Zoheir, K.M.; Ansari, M.A.; Korashy, H.M.; Abdel-Hamied, H.E.; Ashour, A.E.; Abd-Allah, A.R. Naringin attenuates the development of carrageenan-induced acute lung inflammation through inhibition of NF-κb, STAT3 and proinflammatory mediators and enhancement of IκBα and anti-inflammatory cytokines. Inflammation 2015, 38, 846–857. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, R.F. The interface between COVID-19 and bacterial healthcare-associated infections. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2021, 27, 1772–1776. [Google Scholar] [CrossRef] [PubMed]
- Bardelčíková, A.; Miroššay, A.; Šoltýs, J.; Mojžiš, J. Therapeutic and prophylactic effect of flavonoids in post-COVID-19 therapy. Phytother. Res. 2022, 36, 2042–2060. [Google Scholar] [CrossRef]
- Santana, F.P.R.; Thevenard, F.; Gomes, K.S.; Taguchi, L.; Câmara, N.O.S.; Stilhano, R.S.; Ureshino, R.P.; Prado, C.M.; Lago, J.H.G. New perspectives on natural flavonoids on COVID-19-induced lung injuries. Phytother. Res. 2021, 35, 4988–5006. [Google Scholar] [CrossRef]
- Tutunchi, H.; Naeini, F.; Ostadrahimi, A.; Hosseinzadeh-Attar, M.J. Naringenin, a flavanone with antiviral and anti-inflammatory effects: A promising treatment strategy against COVID-19. Phytother. Res. 2020, 34, 3137–3147. [Google Scholar] [CrossRef]
- Zhu, Z.; Wu, H.; Su, W.; Shi, R.; Li, P.; Liao, Y.; Wang, Y.; Li, P. Effects of Total Flavonoids from Exocarpium Citri Grandis on Air Pollution Particle-Induced Pulmonary Inflammation and Oxidative Stress in Mice. J. Food Sci. 2019, 84, 3843–3849. [Google Scholar] [CrossRef]
- Haggag, Y.A.; El-Ashmawy, N.E.; Okasha, K.M. Is hesperidin essential for prophylaxis and treatment of COVID-19 Infection? Med. Hypotheses 2020, 144, 109957. [Google Scholar] [CrossRef]
- Wang, Z.; Ding, Z.; Li, Z.; Ding, Y.; Jiang, F.; Liu, J. Antioxidant and antibacterial study of 10 flavonoids revealed rutin as a potential antibiofilm agent in Klebsiella pneumoniae strains isolated from hospitalized patients. Microb. Pathog. 2021, 159, 105121. [Google Scholar] [CrossRef]
- Ozçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Biol. 2011, 49, 396–402. [Google Scholar] [CrossRef]
- Shilpa, V.S.; Shams, R.; Dash, K.K.; Pandey, V.K.; Dar, A.H.; Ayaz Mukarram, S.; Harsányi, E.; Kovács, B. Phytochemical Properties, Extraction, and Pharmacological Benefits of Naringin: A Review. Molecules 2023, 28, 5623. [Google Scholar] [CrossRef]
- Zeng, X.; Su, W.; Liu, B.; Chai, L.; Shi, R.; Yao, H. A Review on the Pharmacokinetic Properties of Naringin and Its Therapeutic Efficacies in Respiratory Diseases. Mini Rev. Med. Chem. 2020, 20, 286–293. [Google Scholar] [CrossRef]
- Wanford, J.J.; Hames, R.G.; Carreno, D.; Jasiunaite, Z.; Chung, W.Y.; Arena, F.; Di Pilato, V.; Straatman, K.; West, K.; Farzand, R.; et al. Interaction of Klebsiella pneumoniae with tissue macrophages in a mouse infection model and ex-vivo pig organ perfusions: An exploratory investigation. Lancet Microbe 2021, 2, e695–e703. [Google Scholar] [CrossRef]
- Gonzalez-Ferrer, S.; Peñaloza, H.F.; Budnick, J.A.; Bain, W.G.; Nordstrom, H.R.; Lee, J.S.; Van Tyne, D. Finding Order in the Chaos: Outstanding Questions in Klebsiella pneumoniae Pathogenesis. Infect. Immun. 2021, 89, e00693-20. [Google Scholar] [CrossRef]
- Liu, X.; Lu, J.; Liao, Y.; Liu, S.; Chen, Y.; He, R.; Men, L.; Lu, C.; Chen, Z.; Li, S.; et al. Dihydroartemisinin attenuates lipopolysaccharide-induced acute kidney injury by inhibiting inflammation and oxidative stress. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 117, 109070. [Google Scholar] [CrossRef]
- Gong, J.; Li, J.; Dong, H.; Chen, G.; Qin, X.; Hu, M.; Yuan, F.; Fang, K.; Wang, D.; Jiang, S.; et al. Inhibitory effects of berberine on proinflammatory M1 macrophage polarization through interfering with the interaction between TLR4 and MyD88. BMC Complement. Altern. Med. 2019, 19, 314. [Google Scholar] [CrossRef]
- Lu, H.; Wu, L.; Liu, L.; Ruan, Q.; Zhang, X.; Hong, W.; Wu, S.; Jin, G.; Bai, Y. Quercetin ameliorates kidney injury and fibrosis by modulating M1/M2 macrophage polarization. Biochem. Pharmacol. 2018, 154, 203–212. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | NAR (80 mg/kg) | NAR (160 mg/kg) | |
|---|---|---|---|
| ALB (g/L) | 36.28 ± 1.34 | 37.42 ± 1.69 | 36.26 ± 1.20 |
| ALP (U/L) | 265.64 ± 19.42 | 256.18 ± 23.21 | 271.50 ± 30.64 |
| ALT (U/L) | 49.94 ± 7.36 | 46.19 ± 11.12 | 55.83 ± 14.06 |
| AST (U/L) | 158.97 ± 17.72 | 156.77 ± 26.52 | 165.93 ± 10.89 |
| GREA (μmol/L) | 34.72 ± 6.06 | 35.59 ± 8.14 | 38.32 ± 5.20 |
| TBiL (μmol/L) | 0.97 ± 0.13 | 0.83 ± 0.24 | 0.91 ± 0.14 |
| TC (mmol/L) | 2.87 ± 0.26 | 2.86 ± 0.33 | 2.97 ± 0.36 |
| TG (mmol/L) | 1.11 ± 0.19 | 1.26 ± 0.43 | 1.13 ± 0.27 |
| TP (g/L) | 77.34 ± 2.17 | 76.28 ± 4.27 | 76.10 ± 2.43 |
| UREA (mmol/L) | 9.13 ± 0.81 | 8.90 ± 0.82 | 9.41 ± 0.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, G.; Huang, Q.; Jing, X.; Huang, L.; Liu, C.; Pan, X.; Li, Z.; Li, S.; Qiu, Z.; Xin, R. Therapeutic Effect and Safety Evaluation of Naringin on Klebsiella pneumoniae in Mice. Int. J. Mol. Sci. 2023, 24, 15940. https://doi.org/10.3390/ijms242115940
Zhao G, Huang Q, Jing X, Huang L, Liu C, Pan X, Li Z, Li S, Qiu Z, Xin R. Therapeutic Effect and Safety Evaluation of Naringin on Klebsiella pneumoniae in Mice. International Journal of Molecular Sciences. 2023; 24(21):15940. https://doi.org/10.3390/ijms242115940
Chicago/Turabian StyleZhao, Guanyu, Qilin Huang, Xiaohan Jing, Lina Huang, Chen Liu, Xiangyi Pan, Zhaorong Li, Sifan Li, Zhengying Qiu, and Ruihua Xin. 2023. "Therapeutic Effect and Safety Evaluation of Naringin on Klebsiella pneumoniae in Mice" International Journal of Molecular Sciences 24, no. 21: 15940. https://doi.org/10.3390/ijms242115940
APA StyleZhao, G., Huang, Q., Jing, X., Huang, L., Liu, C., Pan, X., Li, Z., Li, S., Qiu, Z., & Xin, R. (2023). Therapeutic Effect and Safety Evaluation of Naringin on Klebsiella pneumoniae in Mice. International Journal of Molecular Sciences, 24(21), 15940. https://doi.org/10.3390/ijms242115940