
Enhanced Intervertebral Disc Repair via Genetically Engineered Mesenchymal Stem Cells with Tetracycline Regulatory System

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
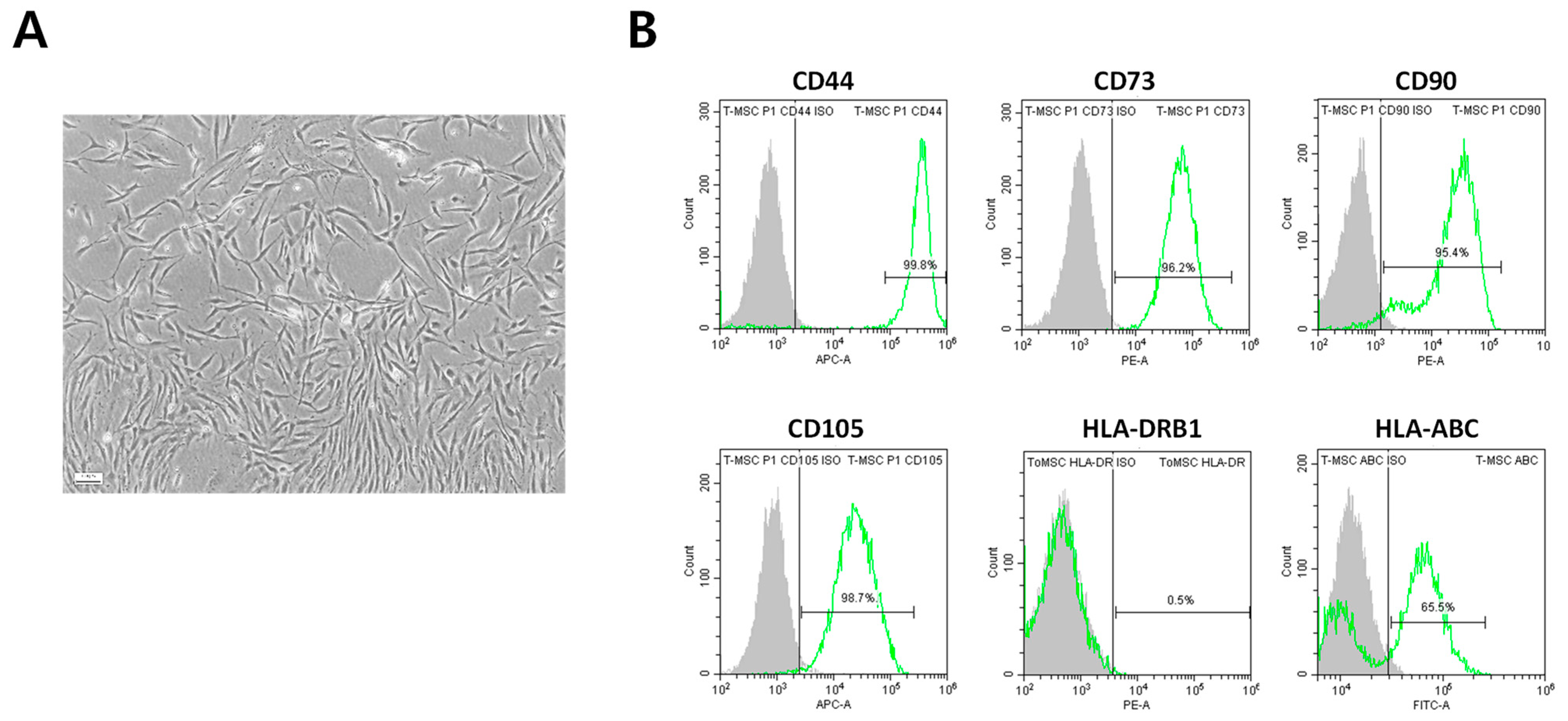
2.1. Immunophenotypic Characterization of ToMSCs
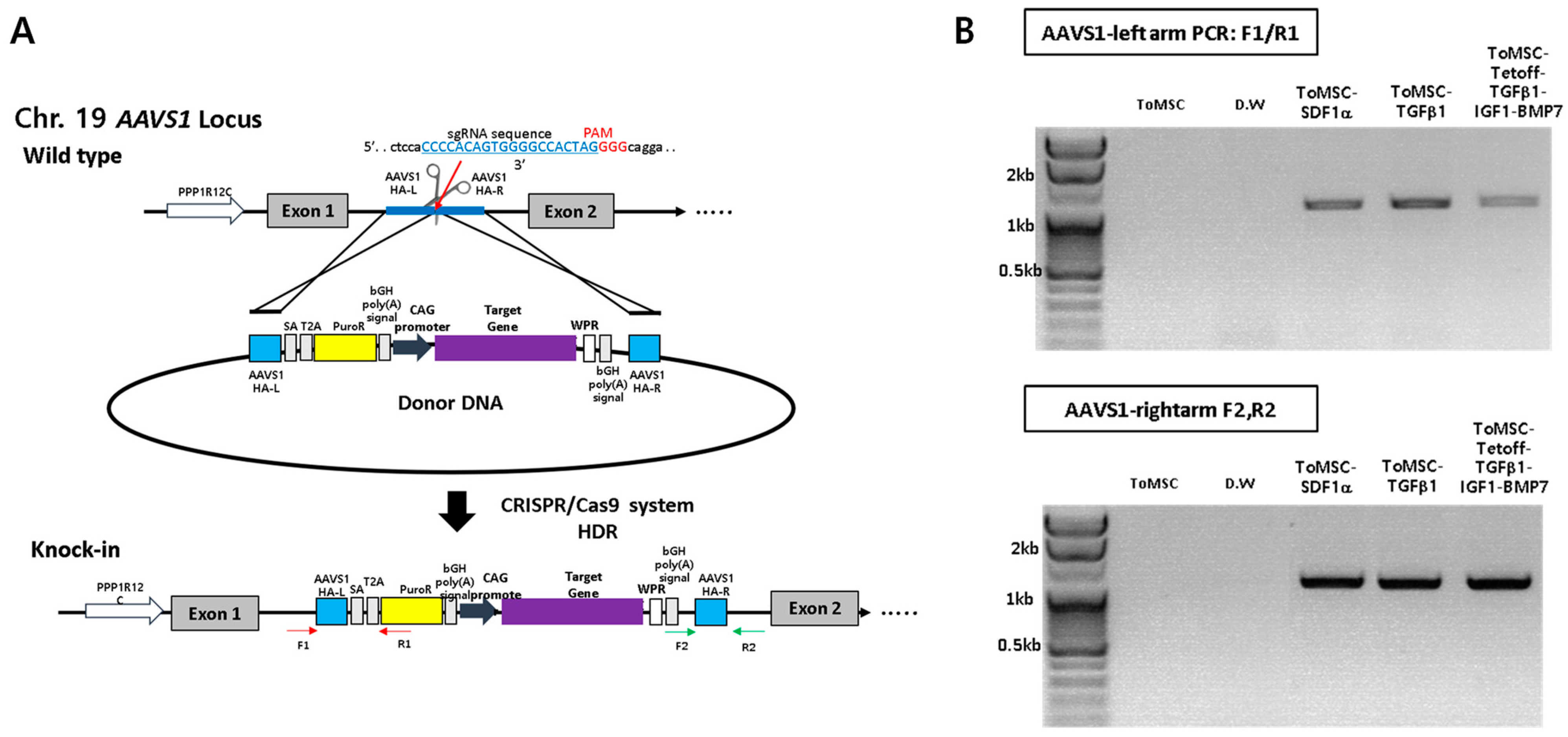
2.2. CRISPR/Cas9-Mediated Knock-In of Transgenes into a Safe-Harbor Site on the ToMSC Chromosome
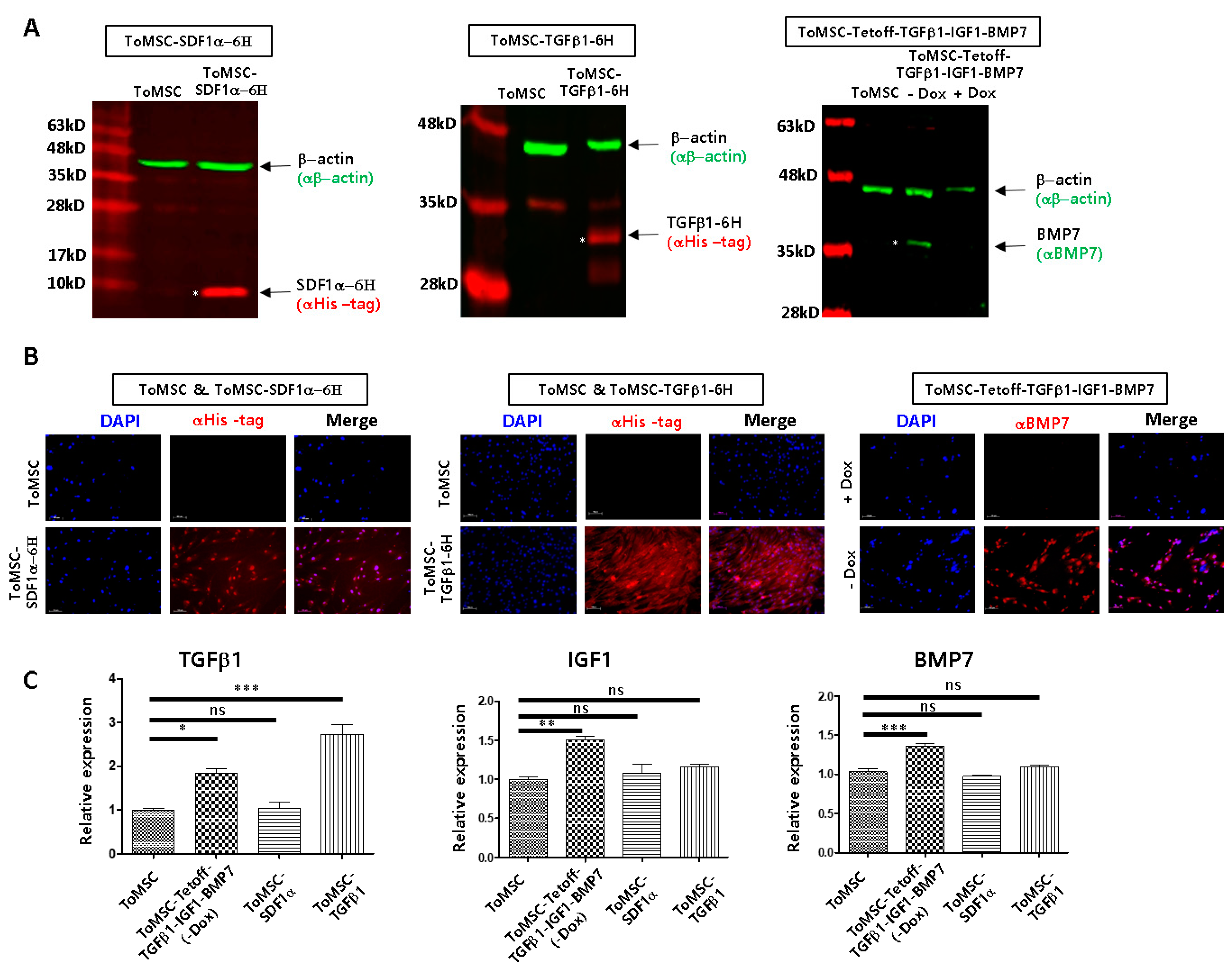
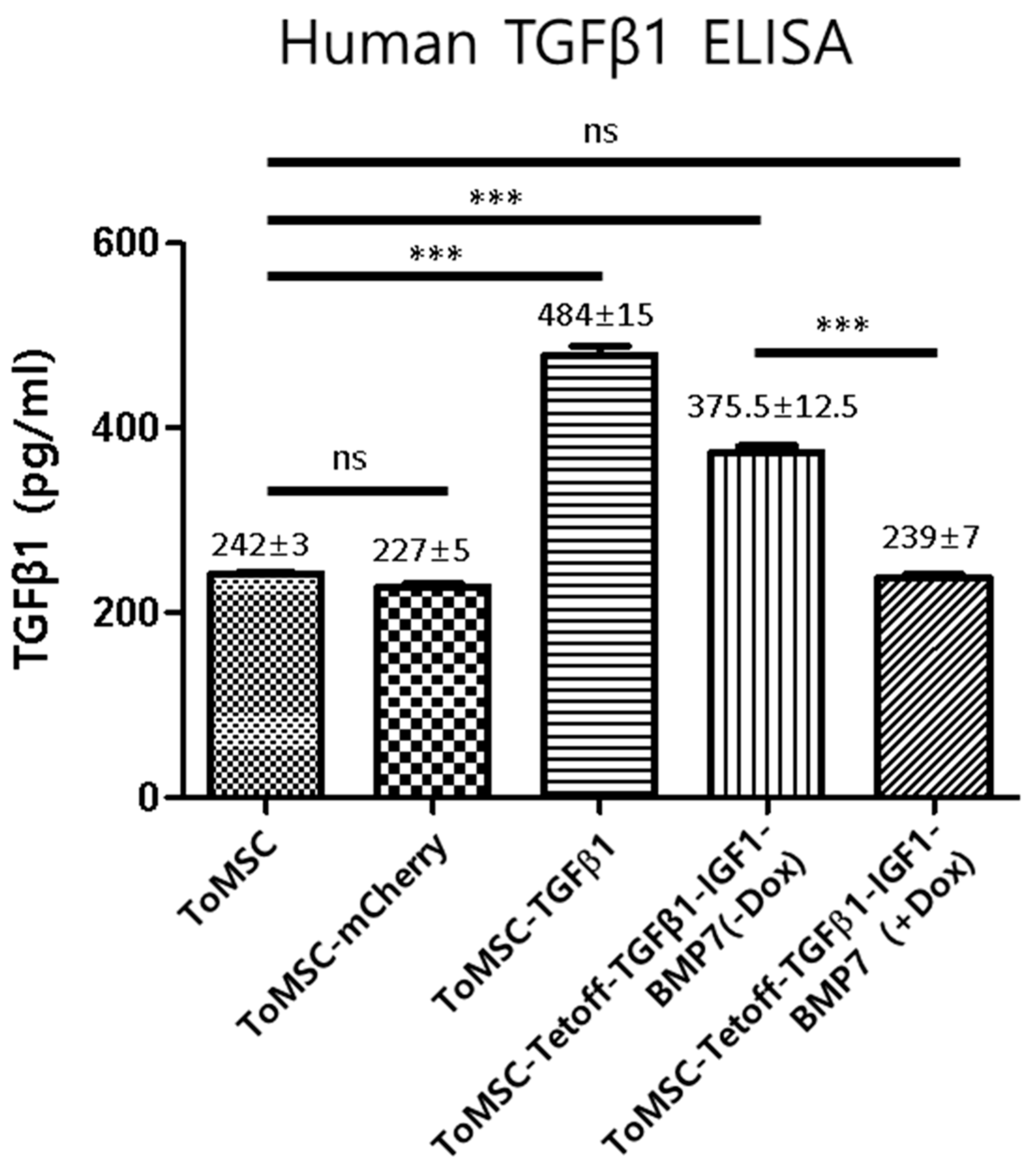
2.3. Assessment of Transgenic Expression through qRT-PCR, Western Blots, Immunocytochemistry, and ELISA
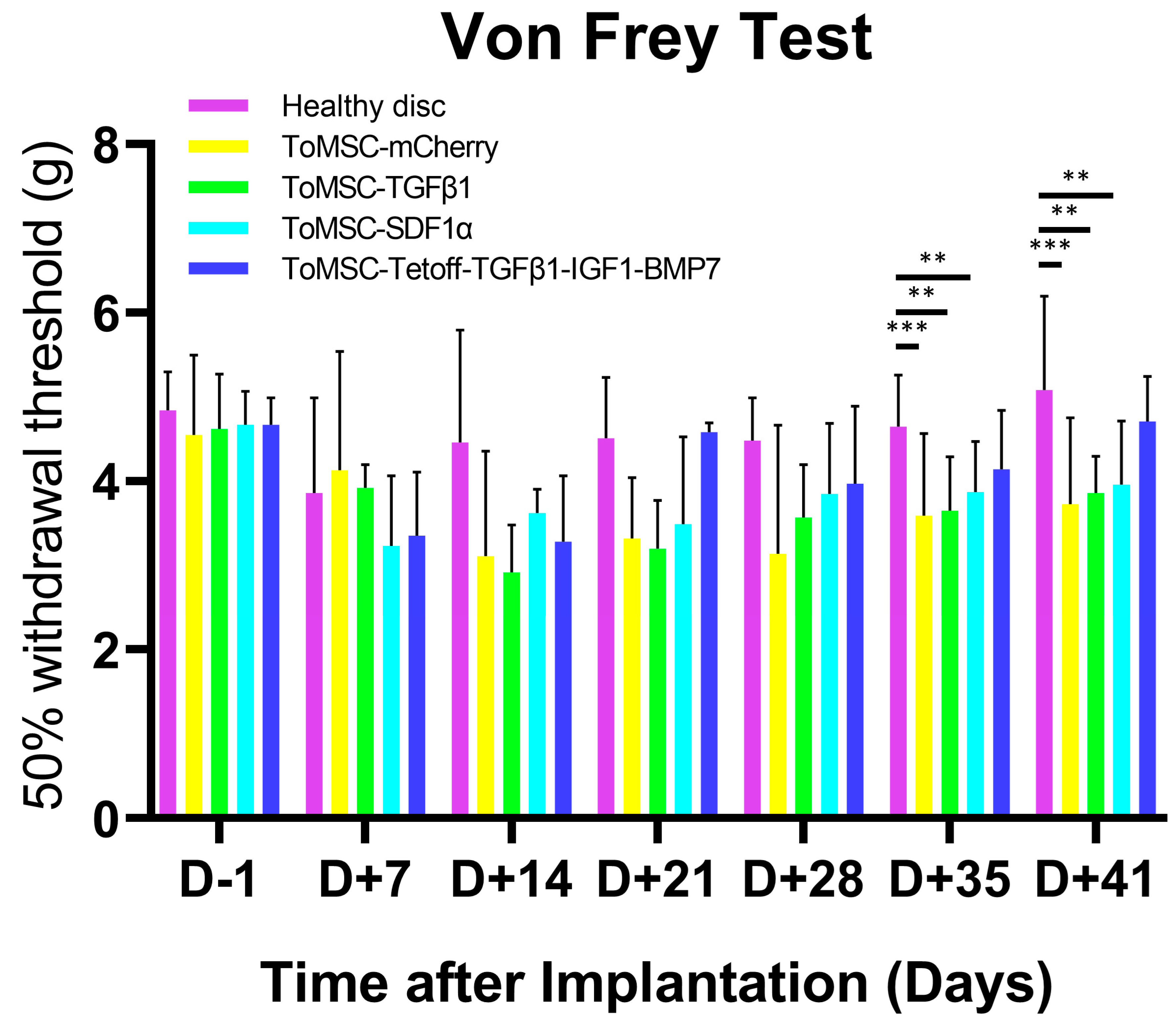
2.4. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Exhibited the Best Anti-Allodynic Effect
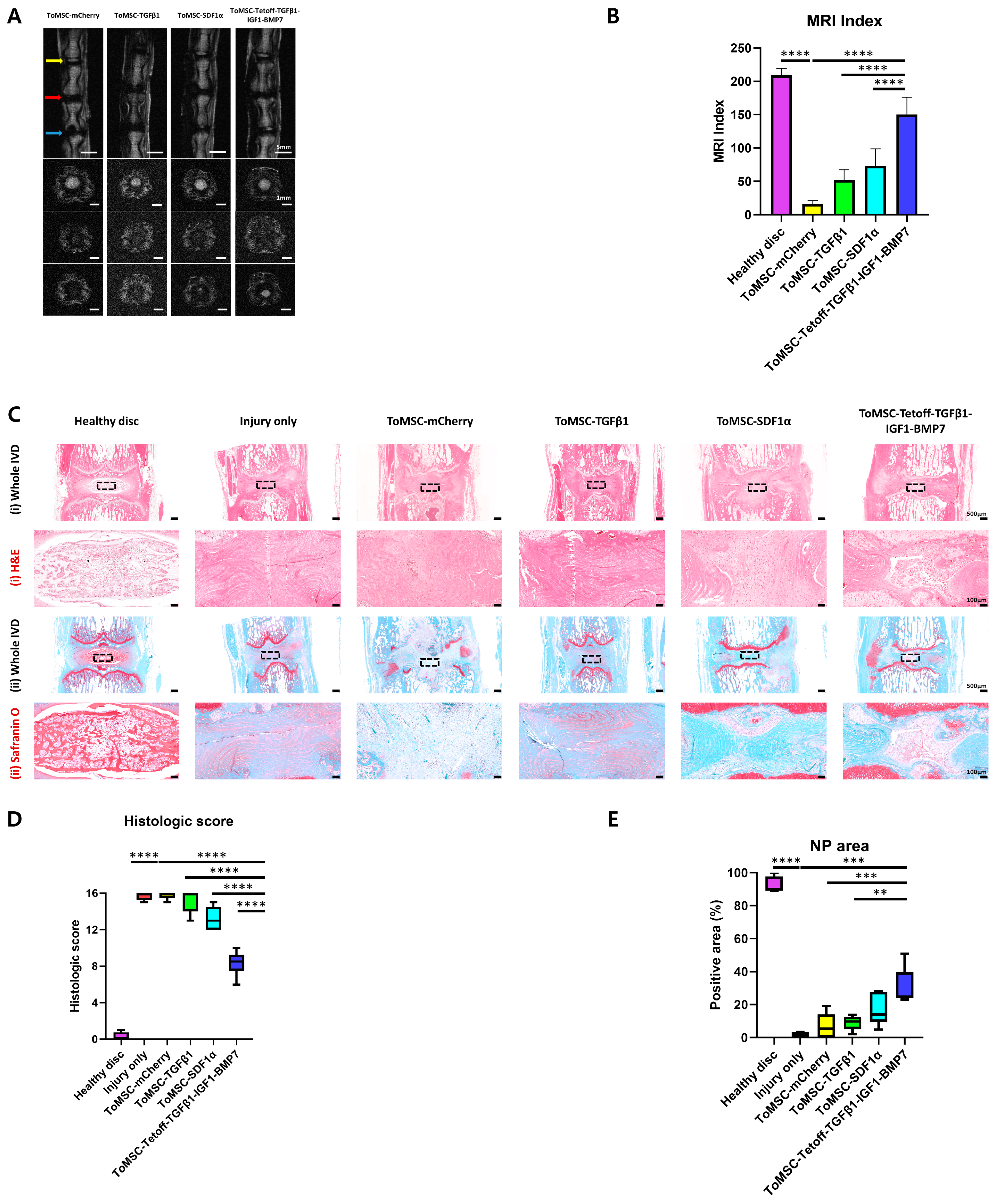
2.5. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Demonstrated the Best Restoration of Disc Anatomy and Hydration in the Rat Tail Needle Puncture Injury Model
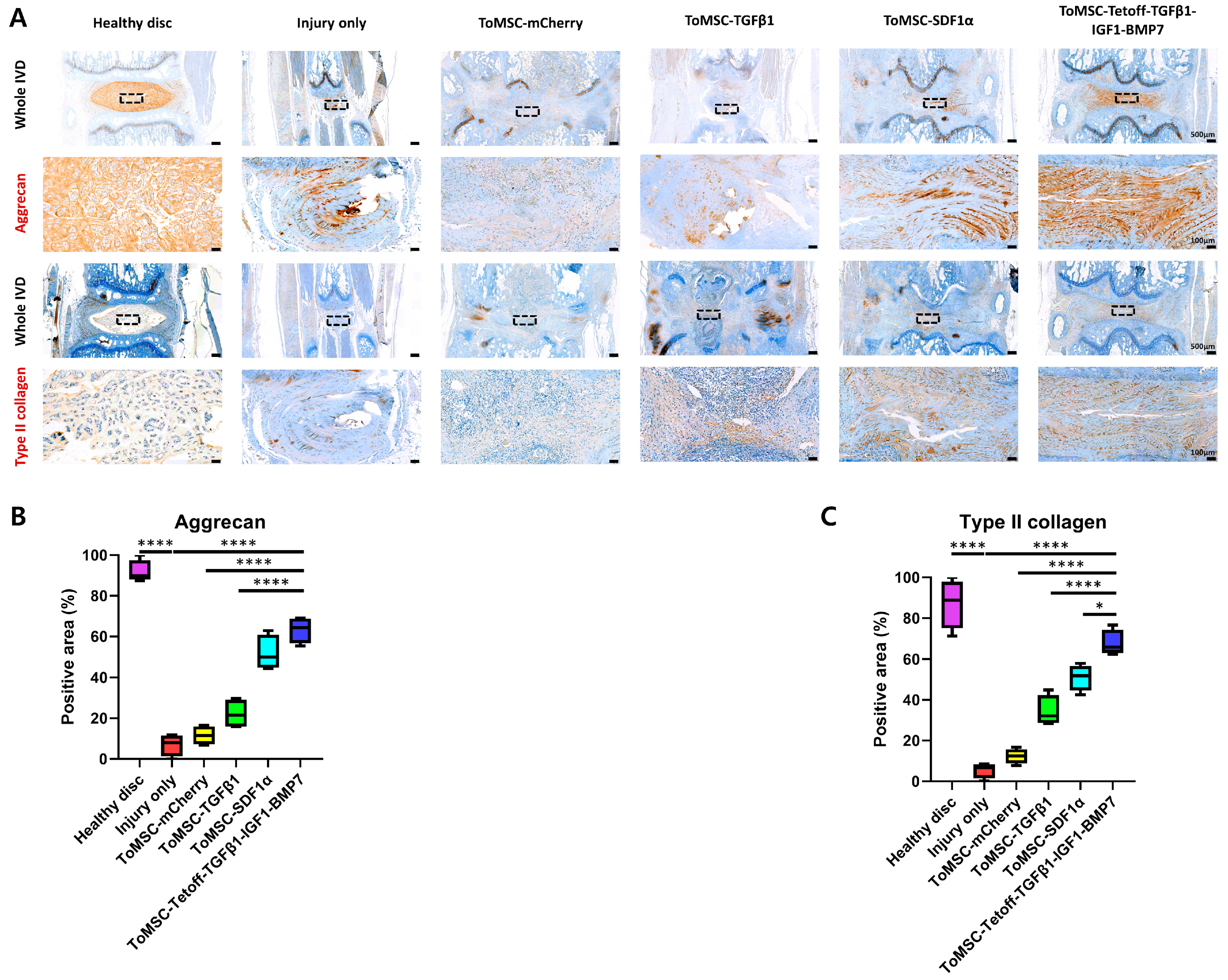
2.6. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Demonstrated the Best Preservation of Matrix Proteins in the Disc NP of the Rat Tail Needle Puncture Injury Model
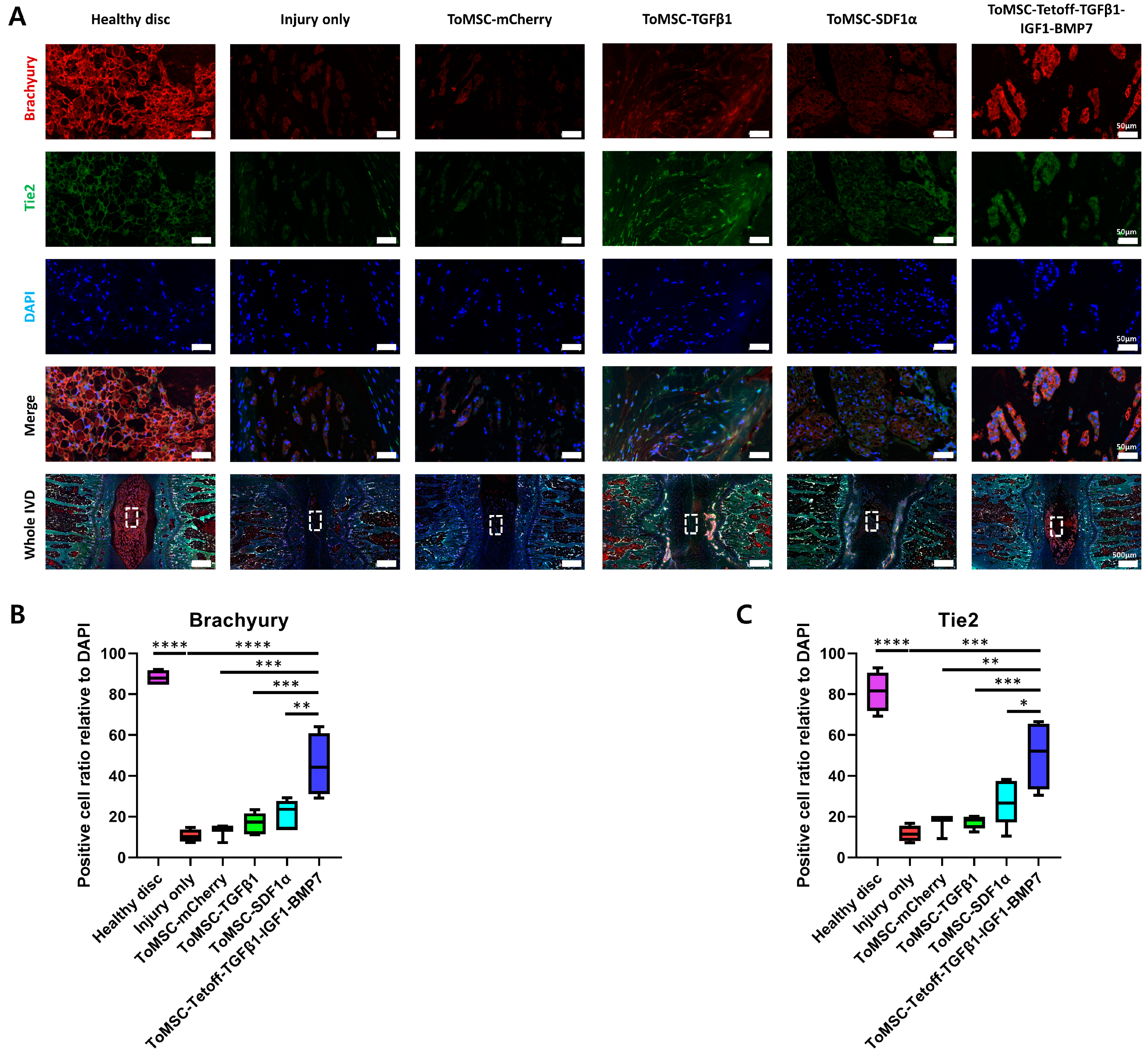
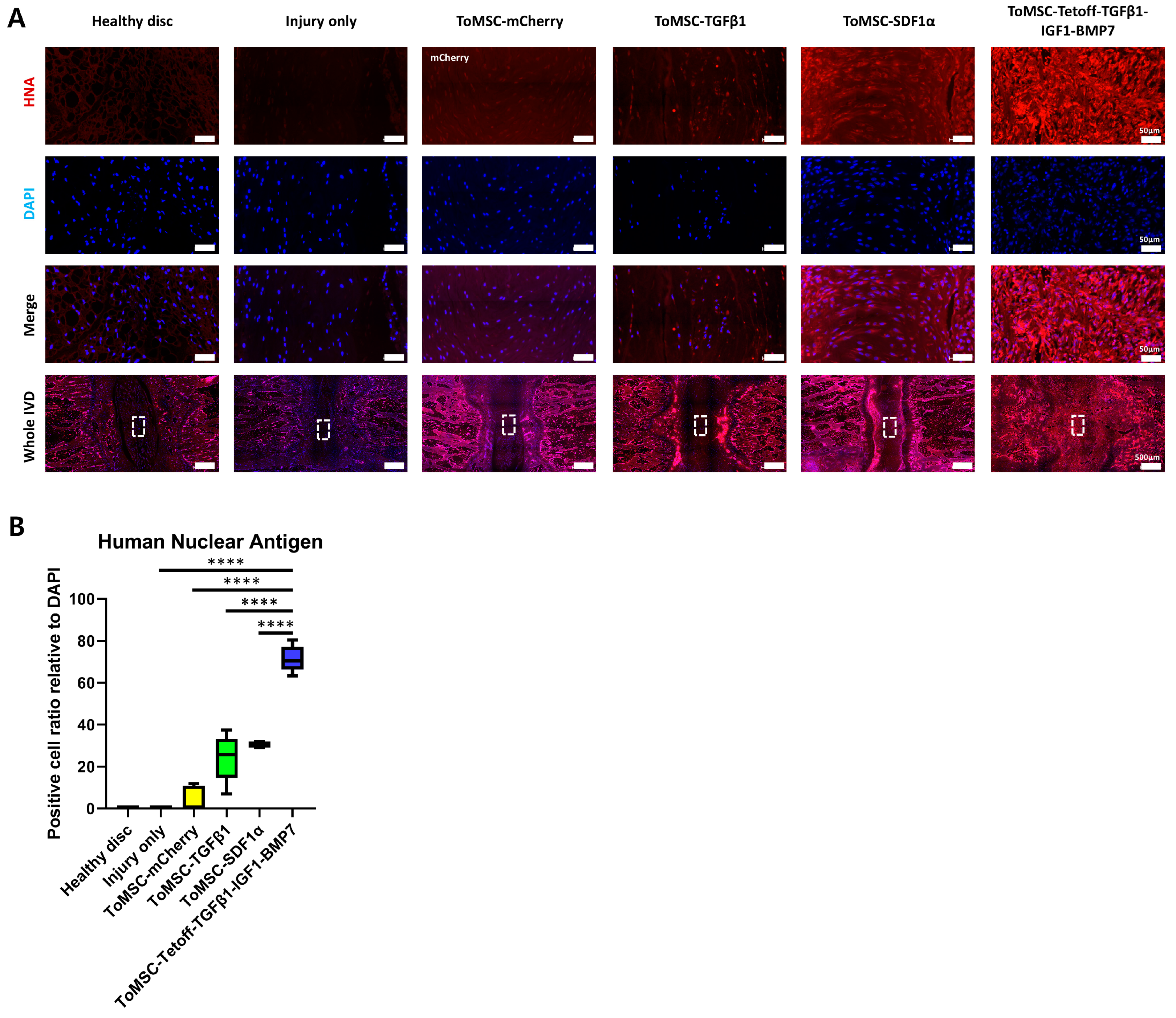
2.7. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Exhibited the Best Preservation of Endogenous Disc NP Progenitor Cells
2.8. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Displays the Greatest Cell Proliferation
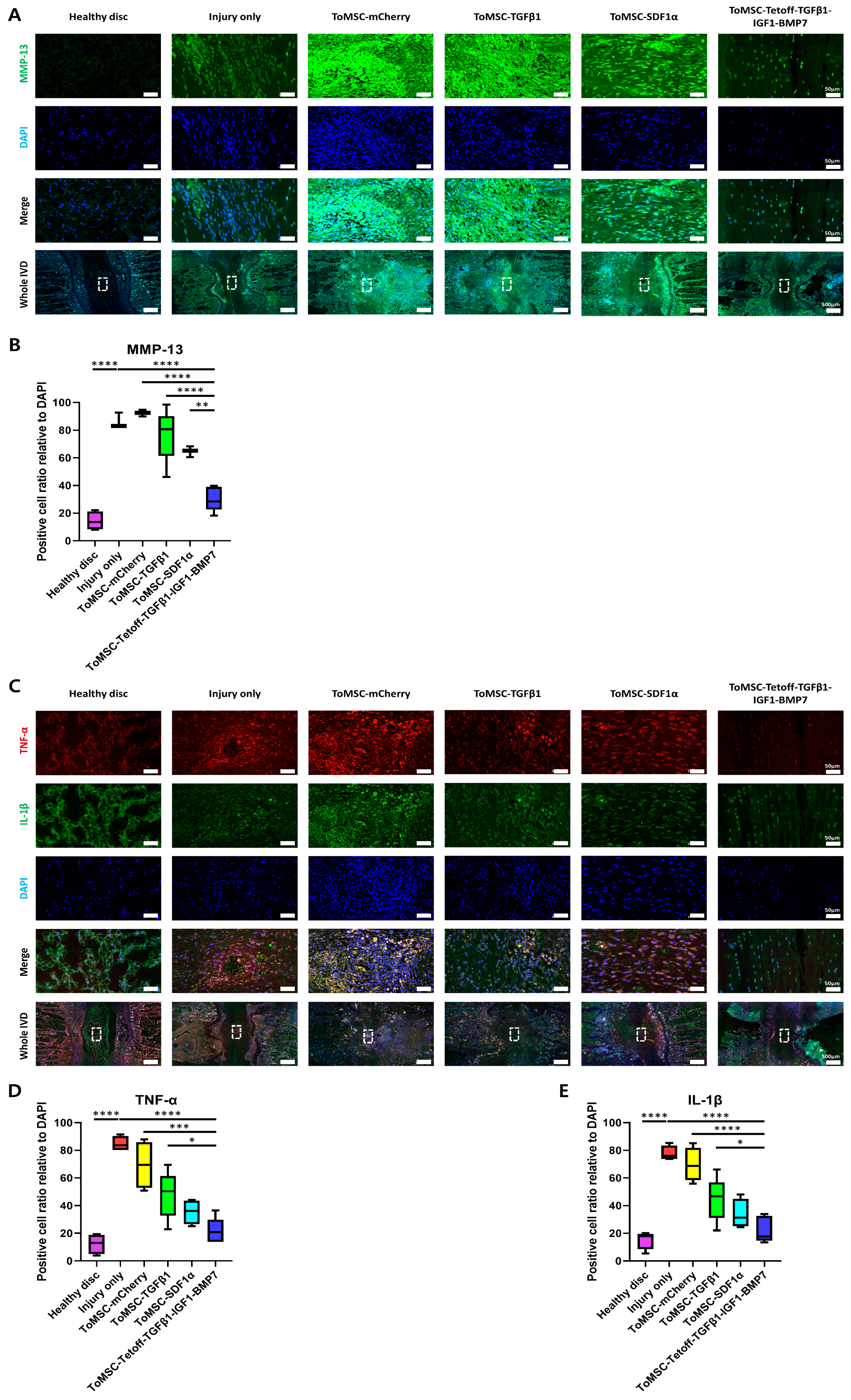
2.9. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Demonstrated the Greatest Decrease in Catabolic Enzymes and Pro-Inflammatory Cytokines in the Disc NP
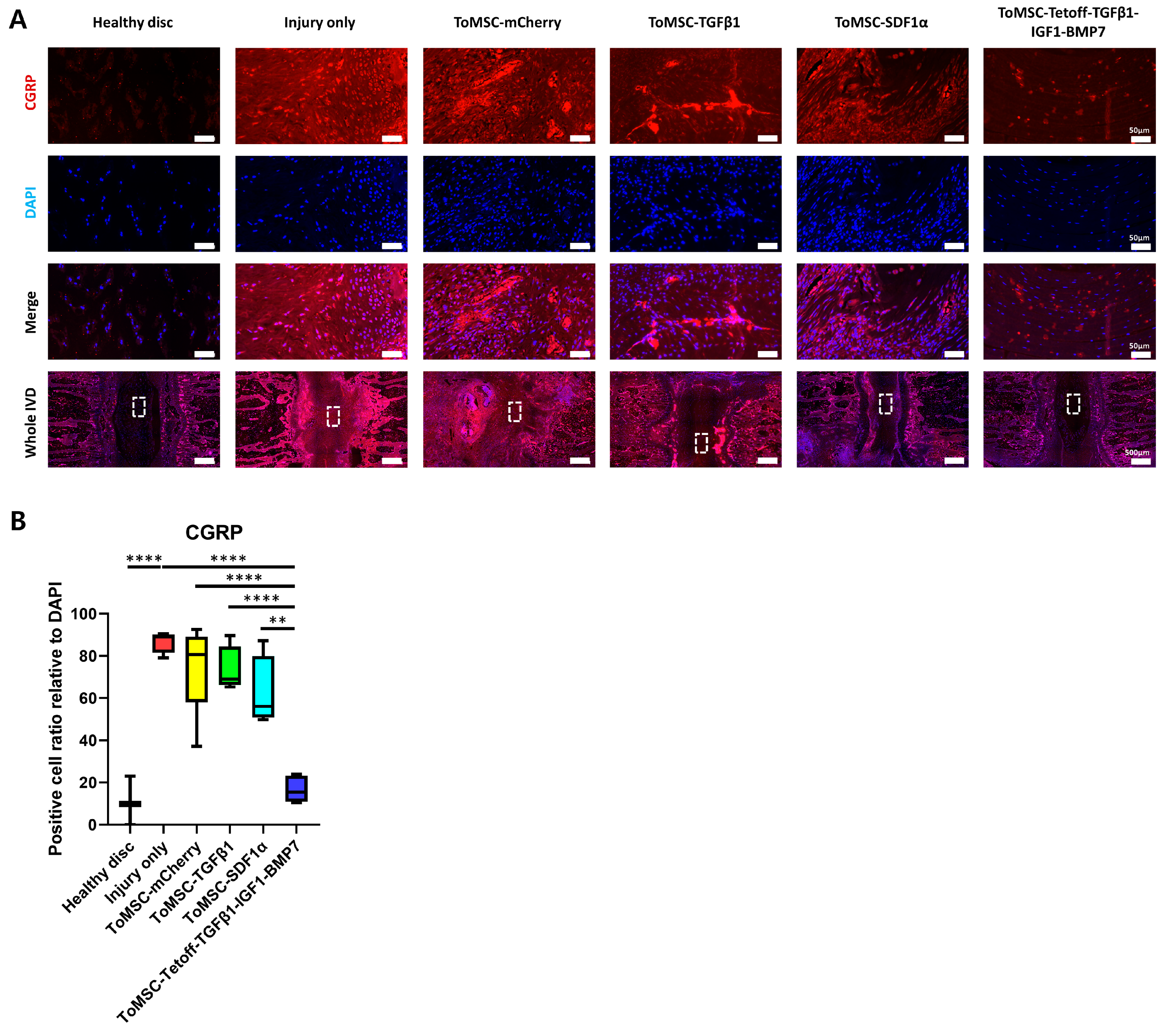
2.10. ToMSC-Tetoff-TGFβ1-IGF1-BMP7 Shows the Greatest Reduction of Injury-Induced Pain
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of Tonsil-Derived Mesenchymal Stem Cells
4.2. Flow Cytometry Analysis
4.3. Plasmid Construction
4.3.1. pAAVS1-Puro-CAG-SDF1α-6H
4.3.2. pAAVS1-Puro-CAG-TGFβ1-6H
4.3.3. pAAVS1-Puro-CAG-mCherry
4.3.4. pAAVS1-Puro-Tetoff-TGFβ1-IGF1-BMP7-CAG-tTA-Advanced
4.4. CRISPR/Cas9-Mediated Knock-In Gene Editing
4.5. Immunofluorescent Detection of Transgene Expression
4.6. Western Blots
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Enzyme-Linked Immunosorbent Assay
4.9. Animal Surgical Procedures
4.10. Experimental Design in a Rat Disc Degeneration Model
4.11. Quantitative Behavioral Nociception Assay
4.12. Magnetic Resonance Imaging
4.13. Histological Analysis
4.14. Immunofluorescence and Immunohistochemistry
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goel, A. Cervical facet joint degeneration. Neurospine 2022, 19, 847–850. [Google Scholar] [CrossRef]
- Yingsakmongkol, W.; Jitpakdee, K.; Kerr, S.; Limthongkul, W.; Kotheeranurak, V.; Singhatanadgige, W. Successful criteria for indirect decompression with lateral lumbar interbody fusion. Neurospine 2022, 19, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Joaquim, A.F.; Lee, N.J.; Riew, K.D. Revision surgeries at the index level after cervical disc arthroplasty—A systematic review. Neurospine 2021, 18, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, J.; Pernaa, K.; Rantakokko, J.; Laaksonen, I.; Saltychev, M. Effect of lumbar laminectomy on spinal sagittal alignment: A systematic review. Eur. Spine J. 2021, 30, 2413–2426. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.P.; Cha, E.D.K.; Jadczak, C.N.; Mohan, S.; Geoghegan, C.E.; Singh, K. What can legacy patient-reported outcome measures tell us about participation bias in patient-reported outcomes measurement information system scores among lumbar spine patients? Neurospine 2022, 19, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, N.; Pathak, Z.; Kumar, H. Extra cellular matrix remodeling: An adjunctive target for spinal cord injury and intervertebral disc degeneration. Neurospine 2022, 19, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, A.; Takeshima, Y.; Yokoyama, S.; Nishimura, F.; Nakagawa, I.; Park, Y.S.; Nakase, H. Prevalence and clinical impact of cervical facet joint degeneration on degenerative cervical myelopathy: A novel computed tomography classification study. Neurospine 2022, 19, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; He, L.; Huang, L.; Liu, Z.; Dong, J.; Liu, B.; Chen, R.; Zhang, L.; Xie, P.; Rong, L. Risk factors for poor outcomes following minimally invasive discectomy: A post hoc subgroup analysis of 2-year follow-up prospective data. Neurospine 2022, 19, 224–235. [Google Scholar] [CrossRef]
- Yoo, B.R.; Son, S.; Lee, S.G.; Kim, W.K.; Jung, J.M. Factors predicting the clinical outcome after trans-sacral epiduroscopic laser decompression for lumbar disc herniation. Neurospine 2021, 18, 336–343. [Google Scholar] [CrossRef]
- Perez-Roman, R.J.; McCarthy, D.; Luther, E.M.; Lugo-Pico, J.G.; Leon-Correa, R.; Gaztanaga, W.; Madhavan, K.; Vanni, S. Effects of body mass index on perioperative outcomes in patients undergoing anterior cervical discectomy and fusion surgery. Neurospine 2021, 18, 79–86. [Google Scholar] [CrossRef]
- Sebaaly, A.; Lahoud, M.J.; Rizkallah, M.; Kreichati, G.; Kharrat, K. Etiology, evaluation, and treatment of failed back surgery syndrome. Asian Spine J. 2018, 12, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.W.; Peng, P. Failed back surgery syndrome. Pain Med. (Malden Mass.) 2011, 12, 577–606. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, C.M. The failed back surgery syndrome: Pitfalls surrounding evaluation and treatment. Phys. Med. Rehabil. Clin. N. Am. 2014, 25, 319–340. [Google Scholar] [CrossRef]
- Ju, D.G.; Kanim, L.E.; Bae, H.W. Intervertebral disc repair: Current concepts. Glob. Spine J. 2020, 10, 130s–136s. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, F.J.; Eiro, N.; Costa, L.; Esparza, P.; Landin, M.; Diaz-Rodriguez, P.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cells in homeostasis and systemic diseases: Hypothesis, evidences, and therapeutic opportunities. Int. J. Mol. Sci. 2019, 20, 3738. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Francos, S.; Eiro, N.; Costa, L.A.; Escudero-Cernuda, S.; Fernández-Sánchez, M.L.; Vizoso, F.J. Mesenchymal stem cells as a cornerstone in a galaxy of intercellular signals: Basis for a new era of medicine. Int. J. Mol. Sci. 2021, 22, 3576. [Google Scholar] [CrossRef]
- Sun, S.J.; Wei, R.; Li, F.; Liao, S.Y.; Tse, H.F. Mesenchymal stromal cell-derived exosomes in cardiac regeneration and repair. Stem Cell Rep. 2021, 16, 1662–1673. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.C.; Niu, Z.F.; Fan, H.J.; Hou, S.K.; Guo, X.Q.; Sang, L.; Lv, Q. Mesenchymal stem cell-derived small extracellular vesicles in the treatment of human diseases: Progress and prospect. World J. Stem Cells 2021, 13, 49–63. [Google Scholar] [CrossRef]
- Hu, S.; Xing, H.; Zhang, J.; Zhu, Z.; Yin, Y.; Zhang, N.; Qi, Y. Mesenchymal stem cell-derived extracellular vesicles: Immunomodulatory effects and potential applications in intervertebral disc degeneration. Stem Cells Int. 2022, 2022, 7538025. [Google Scholar] [CrossRef]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Suzuki, S.; Fujita, N.; Hosogane, N.; Watanabe, K.; Ishii, K.; Toyama, Y.; Takubo, K.; Horiuchi, K.; Miyamoto, T.; Nakamura, M.; et al. Excessive reactive oxygen species are therapeutic targets for intervertebral disc degeneration. Arthritis Res. Ther. 2015, 17, 316. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Yang, M.; Lan, M.; Liu, C.; Zhang, Y.; Huang, B.; Liu, H.; Zhou, Y. Ros: Crucial intermediators in the pathogenesis of intervertebral disc degeneration. Oxid. Med. Cell. Longev. 2017, 2017, 5601593. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.J.; Cui, H.; Pan, H.; Mc Cheung, K.; Cao, X.; Iatridis, J.C.; Zheng, Z. Painful intervertebral disc degeneration and inflammation: From laboratory evidence to clinical interventions. Bone Res. 2021, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Fabre, H.; Ducret, M.; Degoul, O.; Rodriguez, J.; Perrier-Groult, E.; Aubert-Foucher, E.; Pasdeloup, M.; Auxenfans, C.; McGuckin, C.; Forraz, N.; et al. Characterization of different sources of human mscs expanded in serum-free conditions with quantification of chondrogenic induction in 3d. Stem Cells Int. 2019, 2019, 2186728. [Google Scholar] [CrossRef]
- Smith, L.J.; Silverman, L.; Sakai, D.; Le Maitre, C.L.; Mauck, R.L.; Malhotra, N.R.; Lotz, J.C.; Buckley, C.T. Advancing cell therapies for intervertebral disc regeneration from the lab to the clinic: Recommendations of the ors spine section. JOR Spine 2018, 1, e1036. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, J.; Dai, G.; Li, X.; Zhang, Z.; Li, L.; Shen, B. Therapeutic effect of human bone marrow- and adipose-derived stem cells on intervertebral disc degeneration: A comparative study on rats. Discov. Med. 2023, 35, 831–844. [Google Scholar] [CrossRef]
- Khalid, S.; Ekram, S.; Ramzan, F.; Salim, A.; Khan, I. Co-regulation of sox9 and tgfβ1 transcription factors in mesenchymal stem cells regenerated the intervertebral disc degeneration. Front. Med. 2023, 10, 1127303. [Google Scholar] [CrossRef]
- Vadalà, G.; Russo, F.; Ambrosio, L.; Papalia, R.; Denaro, V. Mesenchymal stem cells for intervertebral disc regeneration. J. Biol. Regul. Homeost. Agents 2016, 30, 173–179. [Google Scholar]
- Morita, K.; Schol, J.; Volleman, T.N.E.; Sakai, D.; Sato, M.; Watanabe, M. Screening for growth-factor combinations enabling synergistic differentiation of human msc to nucleus pulposus cell-like cells. Appl. Sci. 2021, 11, 3673. [Google Scholar] [CrossRef]
- Russo, F.; Ambrosio, L.; Peroglio, M.; Guo, W.; Wangler, S.; Gewiess, J.; Grad, S.; Alini, M.; Papalia, R.; Vadalà, G.; et al. A hyaluronan and platelet-rich plasma hydrogel for mesenchymal stem cell delivery in the intervertebral disc: An organ culture study. Int. J. Mol. Sci. 2021, 22, 2963. [Google Scholar] [CrossRef]
- Barakat, A.H.; Elwell, V.A.; Lam, K.S. Stem cell therapy in discogenic back pain. J. Spine Surg. 2019, 5, 561–583. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.S.; Jeong, E.J.; Kim, J.Y.; Park, S.J.; Ju, W.S.; Kim, C.H.; Kim, J.S.; Choo, Y.K. Application of mesenchymal stem cells in inflammatory and fibrotic diseases. Int. J. Mol. Sci. 2020, 21, 8366. [Google Scholar] [CrossRef] [PubMed]
- Mackiewicz, Z.; Salo, J.; Konttinen, Y.T.; Kaigle Holm, A.; Indahl, A.; Pajarinen, J.; Holm, S. Receptor activator of nuclear factor kappa b ligand in an experimental intervertebral disc degeneration. Clin. Exp. Rheumatol. 2009, 27, 299–306. [Google Scholar]
- Herberg, S.; Fulzele, S.; Yang, N.; Shi, X.; Hess, M.; Periyasamy-Thandavan, S.; Hamrick, M.W.; Isales, C.M.; Hill, W.D. Stromal cell-derived factor-1β potentiates bone morphogenetic protein-2-stimulated osteoinduction of genetically engineered bone marrow-derived mesenchymal stem cells in vitro. Tissue Eng. Part A 2013, 19, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Gossen, M.; Freundlieb, S.; Bender, G.; Müller, G.; Hillen, W.; Bujard, H. Transcriptional activation by tetracyclines in mammalian cells. Science 1995, 268, 1766–1769. [Google Scholar] [CrossRef]
- Golchin, A.; Shams, F.; Karami, F. Advancing mesenchymal stem cell therapy with crispr/cas9 for clinical trial studies. Adv. Exp. Med. Biol. 2020, 1247, 89–100. [Google Scholar]
- Khatri, R.; Arad, M.; Ortlip, T.; Portney, B.A.; Meltzer, W.A.; Diaconu, S.; Silipino, L.E.; Wang, Y.; Kaetzel, D.M.; Taylor, R.J.; et al. Harvesting multipotent progenitor cells from a small sample of tonsillar biopsy for clinical applications. Stem Cell Res. Ther. 2017, 8, 174. [Google Scholar] [CrossRef]
- Cho, K.A.; Park, M.; Kim, Y.H.; Woo, S.Y.; Ryu, K.H. Rna sequencing reveals a transcriptomic portrait of human mesenchymal stem cells from bone marrow, adipose tissue, and palatine tonsils. Sci. Rep. 2017, 7, 17114. [Google Scholar] [CrossRef]
- Ryu, K.H.; Cho, K.A.; Park, H.S.; Kim, J.Y.; Woo, S.Y.; Jo, I.; Choi, Y.H.; Park, Y.M.; Jung, S.C.; Chung, S.M.; et al. Tonsil-derived mesenchymal stromal cells: Evaluation of biologic, immunologic and genetic factors for successful banking. Cytotherapy 2012, 14, 1193–1202. [Google Scholar] [CrossRef]
- Park, M.; Kim, Y.H.; Woo, S.Y.; Lee, H.J.; Yu, Y.; Kim, H.S.; Park, Y.S.; Jo, I.; Park, J.W.; Jung, S.C.; et al. Tonsil-derived mesenchymal stem cells ameliorate ccl4-induced liver fibrosis in mice via autophagy activation. Sci. Rep. 2015, 5, 8616. [Google Scholar] [CrossRef]
- Park, S.; Choi, Y.; Jung, N.; Yu, Y.; Ryu, K.H.; Kim, H.S.; Jo, I.; Choi, B.O.; Jung, S.C. Myogenic differentiation potential of human tonsil-derived mesenchymal stem cells and their potential for use to promote skeletal muscle regeneration. Int. J. Mol. Med. 2016, 37, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Yu, Y.L.; Tang, C.L.; Lv, F.Z. Effects of tgf-β1 and il-1β on expression of adamts enzymes and timp-3 in human intervertebral disc degeneration. Exp. Ther. Med. 2013, 6, 1522–1526. [Google Scholar] [CrossRef]
- Travascio, F.; Elmasry, S.; Asfour, S. Modeling the role of igf-1 on extracellular matrix biosynthesis and cellularity in intervertebral disc. J. Biomech. 2014, 47, 2269–2276. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lang, G.; Karfeld-Sulzer, L.S.; Mader, K.T.; Richards, R.G.; Weber, F.E.; Sammon, C.; Sacks, H.; Yayon, A.; Alini, M.; et al. Heterodimeric bmp-2/7 for nucleus pulposus regeneration-in vitro and ex vivo studies. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2017, 35, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Davidson, D.; Blanc, A.; Filion, D.; Wang, H.; Plut, P.; Pfeffer, G.; Buschmann, M.D.; Henderson, J.E. Fibroblast growth factor (fgf) 18 signals through fgf receptor 3 to promote chondrogenesis. J. Biol. Chem. 2005, 280, 20509–20515. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.G.; Kim, T.K.; Taboas, A.; Malik, A.; Manson, P.; Elisseeff, J. In vitro chondrogenesis of bone marrow-derived mesenchymal stem cells in a photopolymerizing hydrogel. Tissue Eng. 2003, 9, 679–688. [Google Scholar] [CrossRef]
- Indrawattana, N.; Chen, G.; Tadokoro, M.; Shann, L.H.; Ohgushi, H.; Tateishi, T.; Tanaka, J.; Bunyaratvej, A. Growth factor combination for chondrogenic induction from human mesenchymal stem cell. Biochem. Biophys. Res. Commun. 2004, 320, 914–919. [Google Scholar] [CrossRef]
- Park, Y.; Sugimoto, M.; Watrin, A.; Chiquet, M.; Hunziker, E.B. Bmp-2 induces the expression of chondrocyte-specific genes in bovine synovium-derived progenitor cells cultured in three-dimensional alginate hydrogel. Osteoarthr. Cartil. 2005, 13, 527–536. [Google Scholar] [CrossRef]
- Worster, A.A.; Brower-Toland, B.D.; Fortier, L.A.; Bent, S.J.; Williams, J.; Nixon, A.J. Chondrocytic differentiation of mesenchymal stem cells sequentially exposed to transforming growth factor-beta1 in monolayer and insulin-like growth factor-i in a three-dimensional matrix. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2001, 19, 738–749. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, B.; Zhao, J.; Pan, W.; Xu, J.; Chen, S. Igf-i induces adipose derived mesenchymal cell chondrogenic differentiation in vitro and enhances chondrogenesis in vivo. Vitr. Cell. Dev. Biol. Anim. 2016, 52, 356–364. [Google Scholar] [CrossRef]
- Lee, P.T.; Li, W.J. Chondrogenesis of embryonic stem cell-derived mesenchymal stem cells induced by tgfβ1 and bmp7 through increased tgfβ receptor expression and endogenous tgfβ1 production. J. Cell. Biochem. 2017, 118, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Maguire, S.; Davis, L.A.; Alexander, M.; Yang, F.; Chandran, S.; ffrench-Constant, C.; Pedersen, R.A. Robust, persistent transgene expression in human embryonic stem cells is achieved with aavs1-targeted integration. Stem Cells (Dayt. Ohio) 2008, 26, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Janjanin, S.; Djouad, F.; Shanti, R.M.; Baksh, D.; Gollapudi, K.; Prgomet, D.; Rackwitz, L.; Joshi, A.S.; Tuan, R.S. Human palatine tonsil: A new potential tissue source of multipotent mesenchymal progenitor cells. Arthritis Res. Ther. 2008, 10, R83. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hong, J.Y.; Lee, J.; Jeon, W.J.; Ha, I.H. Il-1β promotes disc degeneration and inflammation through direct injection of intervertebral disc in a rat lumbar disc herniation model. Spine J. Off. J. N. Am. Spine Soc. 2021, 21, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kroin, J.S.; Li, X.; An, H.S.; Buvanendran, A.; Yan, D.; Tuman, K.J.; van Wijnen, A.J.; Chen, D.; Im, H.J. The rat intervertebral disk degeneration pain model: Relationships between biological and structural alterations and pain. Arthritis Res. Ther. 2011, 13, R165. [Google Scholar] [CrossRef]
- Bhujel, B.; Yang, S.S.; Kim, H.R.; Kim, S.B.; Min, B.H.; Choi, B.H.; Han, I. An injectable engineered cartilage gel improves intervertebral disc repair in a rat nucleotomy model. Int. J. Mol. Sci. 2023, 24, 3146. [Google Scholar] [CrossRef]
- Lai, A.; Moon, A.; Purmessur, D.; Skovrlj, B.; Winkelstein, B.A.; Cho, S.K.; Hecht, A.C.; Iatridis, J.C. Assessment of functional and behavioral changes sensitive to painful disc degeneration. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2015, 33, 755–764. [Google Scholar] [CrossRef]
- Deuis, J.R.; Dvorakova, L.S.; Vetter, I. Methods used to evaluate pain behaviors in rodents. Front. Mol. Neurosci. 2017, 10, 284. [Google Scholar] [CrossRef]
- Ohnishi, T.; Sudo, H.; Iwasaki, K.; Tsujimoto, T.; Ito, Y.M.; Iwasaki, N. In vivo mouse intervertebral disc degeneration model based on a new histological classification. PLoS ONE 2016, 11, e0160486. [Google Scholar] [CrossRef]
- Tam, V.; Chan, W.C.W.; Leung, V.Y.L.; Cheah, K.S.E.; Cheung, K.M.C.; Sakai, D.; McCann, M.R.; Bedore, J.; Seguin, C.A.; Chan, D. Histological and reference system for the analysis of mouse intervertebral disc. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2018, 36, 233–243. [Google Scholar] [CrossRef]
- De Geer, C.M. Intervertebral disk nutrients and transport mechanisms in relation to disk degeneration: A narrative literature review. J. Chiropr. Med. 2018, 17, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Roberts, S.; Smith, S.; Menage, J.; Ghosh, P. Increased nerve and blood vessel ingrowth associated with proteoglycan depletion in an ovine anular lesion model of experimental disc degeneration. Spine 2002, 27, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.P.G.; Fairbank, J.C.T. Current perspectives on the role of biomechanical loading and genetics in development of disc degeneration and low back pain; a narrative review. J. Biomech. 2020, 102, 109573. [Google Scholar] [CrossRef]
- Zhang, Y.; He, F.; Chen, Z.; Su, Q.; Yan, M.; Zhang, Q.; Tan, J.; Qian, L.; Han, Y. Melatonin modulates il-1β-induced extracellular matrix remodeling in human nucleus pulposus cells and attenuates rat intervertebral disc degeneration and inflammation. Aging 2019, 11, 10499–10512. [Google Scholar] [CrossRef] [PubMed]
- Nakamichi, R.; Asahara, H. The transcription factors regulating intervertebral disc development. JOR Spine 2020, 3, e1081. [Google Scholar] [CrossRef]
- Tekari, A.; Chan, S.C.W.; Sakai, D.; Grad, S.; Gantenbein, B. Angiopoietin-1 receptor tie2 distinguishes multipotent differentiation capability in bovine coccygeal nucleus pulposus cells. Stem Cell Res. Ther. 2016, 7, 75. [Google Scholar] [CrossRef]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.; et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef]
- Iimori, Y.; Morioka, M.; Koyamatsu, S.; Tsumaki, N. Implantation of human-induced pluripotent stem cell-derived cartilage in bone defects of mice. Tissue Eng. Part A 2021, 27, 1355–1367. [Google Scholar] [CrossRef]
- Gilbert, H.T.J.; Hodson, N.; Baird, P.; Richardson, S.M.; Hoyland, J.A. Acidic ph promotes intervertebral disc degeneration: Acid-sensing ion channel -3 as a potential therapeutic target. Sci. Rep. 2016, 6, 37360. [Google Scholar] [CrossRef]
- Rossignol, J.; Boyer, C.; Thinard, R.; Remy, S.; Dugast, A.S.; Dubayle, D.; Dey, N.D.; Boeffard, F.; Delecrin, J.; Heymann, D.; et al. Mesenchymal stem cells induce a weak immune response in the rat striatum after allo or xenotransplantation. J. Cell. Mol. Med. 2009, 13, 2547–2558. [Google Scholar] [CrossRef]
- Cízková, D.; Rosocha, J.; Vanický, I.; Jergová, S.; Cízek, M. Transplants of human mesenchymal stem cells improve functional recovery after spinal cord injury in the rat. Cell. Mol. Neurobiol. 2006, 26, 1167–1180. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.K.; Lin, K.J.; Lin, C.Y.; Chang, Y.H.; Yen, T.C.; Hwang, S.M.; Sung, L.Y.; Chen, H.C.; Hu, Y.C. Xenotransplantation of human mesenchymal stem cells into immunocompetent rats for calvarial bone repair. Tissue Eng. Part A 2010, 16, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Zhang, Z.; Mao, Q.; Wang, C.; Zhou, Y.; Zhou, X.; Ying, L.; Xu, H.; Hu, S.; Zhang, N. Injectable exosome-functionalized extracellular matrix hydrogel for metabolism balance and pyroptosis regulation in intervertebral disc degeneration. J. Nanobiotechnol. 2021, 19, 264. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. Accelerated cellular senescence in degenerate intervertebral discs: A possible role in the pathogenesis of intervertebral disc degeneration. Arthritis Res. Ther. 2007, 9, R45. [Google Scholar] [CrossRef]
- Cai, F.; Zhu, L.; Wang, F.; Shi, R.; Xie, X.H.; Hong, X.; Wang, X.H.; Wu, X.T. The paracrine effect of degenerated disc cells on healthy human nucleus pulposus cells is mediated by mapk and nf-κb pathways and can be reduced by tgf-β1. DNA Cell Biol. 2017, 36, 143–158. [Google Scholar] [CrossRef]
- Evashwick-Rogler, T.W.; Lai, A.; Watanabe, H.; Salandra, J.M.; Winkelstein, B.A.; Cho, S.K.; Hecht, A.C.; Iatridis, J.C. Inhibiting tumor necrosis factor-alpha at time of induced intervertebral disc injury limits long-term pain and degeneration in a rat model. JOR Spine 2018, 1, e1014. [Google Scholar] [CrossRef]
- Wang, F.; Cai, F.; Shi, R.; Wang, X.H.; Wu, X.T. Aging and age related stresses: A senescence mechanism of intervertebral disc degeneration. Osteoarthr. Cartil. 2016, 24, 398–408. [Google Scholar] [CrossRef]
- Wang, Y.; Che, M.; Xin, J.; Zheng, Z.; Li, J.; Zhang, S. The role of il-1β and tnf-α in intervertebral disc degeneration. Biomed. Pharmacother. = Biomed. Pharmacother. 2020, 131, 110660. [Google Scholar] [CrossRef]
- Sun, K.; Zhu, J.; Yan, C.; Li, F.; Kong, F.; Sun, J.; Sun, X.; Shi, J.; Wang, Y. Cgrp regulates nucleus pulposus cell apoptosis and inflammation via the mapk/nf-κb signaling pathways during intervertebral disc degeneration. Oxid. Med. Cell. Longev. 2021, 2021, 2958584. [Google Scholar] [CrossRef]
- Osada, R.; Ohshima, H.; Ishihara, H.; Yudoh, K.; Sakai, K.; Matsui, H.; Tsuji, H. Autocrine/paracrine mechanism of insulin-like growth factor-1 secretion, and the effect of insulin-like growth factor-1 on proteoglycan synthesis in bovine intervertebral discs. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 1996, 14, 690–699. [Google Scholar] [CrossRef]
- Thompson, J.P.; Oegema, T.R., Jr.; Bradford, D.S. Stimulation of mature canine intervertebral disc by growth factors. Spine 1991, 16, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Tang, X.; Li, Q.; Wang, H.; Sun, H.; Tian, J. Mesenchymal stem cell extracellular vesicles-derived microrna-194-5p delays the development of intervertebral disc degeneration by targeting traf6. Regen. Ther. 2022, 19, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Luo, B.; Liu, Z.H.; Samartzis, D.; Liu, Z.; Gao, B.; Huang, L.; Luo, Z.J. Adipose-derived stromal cells protect intervertebral disc cells in compression: Implications for stem cell regenerative disc therapy. Int. J. Biol. Sci. 2015, 11, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Inoue, G.; Ohtori, S.; Aoki, Y.; Ozawa, T.; Doya, H.; Saito, T.; Ito, T.; Akazawa, T.; Moriya, H.; Takahashi, K. Exposure of the nucleus pulposus to the outside of the anulus fibrosus induces nerve injury and regeneration of the afferent fibers innervating the lumbar intervertebral discs in rats. Spine 2006, 31, 1433–1438. [Google Scholar] [CrossRef]
- Takahashi, K.; Aoki, Y.; Ohtori, S. Resolving discogenic pain. Eur. Spine J. 2008, 17, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, M. Novel treatment strategies for intervertebral disc degeneration. Saudi J. Health Sci. 2015, 4, 5–15. [Google Scholar] [CrossRef]
- Krock, E.; Rosenzweig, D.H.; Chabot-Doré, A.J.; Jarzem, P.; Weber, M.H.; Ouellet, J.A.; Stone, L.S.; Haglund, L. Painful, degenerating intervertebral discs up-regulate neurite sprouting and cgrp through nociceptive factors. J. Cell. Mol. Med. 2014, 18, 1213–1225. [Google Scholar] [CrossRef]
- Jiao, Y.; Yuan, Y.; Lin, Y.; Zhou, Z.; Zheng, Y.; Wu, W.; Tang, G.; Chen, Y.; Xiao, J.; Li, C.; et al. Propionibacterium acnes induces discogenic low back pain via stimulating nucleus pulposus cells to secrete pro-algesic factor of il-8/cinc-1 through tlr2-nf-κb p65 pathway. J. Mol. Med. 2019, 97, 25–35. [Google Scholar] [CrossRef]
- Melrose, J. Strategies in regenerative medicine for intervertebral disc repair using mesenchymal stem cells and bioscaffolds. Regen. Med. 2016, 11, 705–724. [Google Scholar] [CrossRef]
- Cunha, C.; Leite Pereira, C.; Ferreira, J.R.; Ribeiro-Machado, C.; Grad, S.; Santos, S.G.; Gonçalves, R.M. Therapeutic strategies for ivd regeneration through hyaluronan/sdf-1-based hydrogel and intravenous administration of mscs. Int. J. Mol. Sci. 2021, 22, 9609. [Google Scholar] [CrossRef]
- Ha, D.H.; Kim, H.K.; Lee, J.; Kwon, H.H.; Park, G.H.; Yang, S.H.; Jung, J.Y.; Choi, H.; Lee, J.H.; Sung, S.; et al. Mesenchymal stem/stromal cell-derived exosomes for immunomodulatory therapeutics and skin regeneration. Cells 2020, 9, 1157. [Google Scholar] [CrossRef] [PubMed]
- Hockemeyer, D.; Soldner, F.; Beard, C.; Gao, Q.; Mitalipova, M.; DeKelver, R.C.; Katibah, G.E.; Amora, R.; Boydston, E.A.; Zeitler, B.; et al. Efficient targeting of expressed and silent genes in human escs and ipscs using zinc-finger nucleases. Nat. Biotechnol. 2009, 27, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Heo, C.H.; Roh, E.J.; Kim, J.; Choi, H.; Jang, H.Y.; Lee, G.; Lim, C.S.; Han, I. Development of a cox-2-selective fluorescent probe for the observation of early intervertebral disc degeneration. J. Funct. Biomater. 2023, 14, 192. [Google Scholar] [CrossRef]
- Choi, U.Y.; Joshi, H.P.; Payne, S.; Kim, K.T.; Kyung, J.W.; Choi, H.; Cooke, M.J.; Kwon, S.Y.; Roh, E.J.; Sohn, S.; et al. An injectable hyaluronan-methylcellulose (hamc) hydrogel combined with wharton’s jelly-derived mesenchymal stromal cells (wj-mscs) promotes degenerative disc repair. Int. J. Mol. Sci. 2020, 21, 7391. [Google Scholar] [CrossRef]
- Lim, S.; An, S.B.; Jung, M.; Joshi, H.P.; Kumar, H.; Kim, C.; Song, S.Y.; Lee, J.R.; Kang, M.; Han, I.J.A.H.M. Local delivery of senolytic drug inhibits intervertebral disc degeneration and restores intervertebral disc structure. Adv. Healthc. Mater. 2022, 11, 2101483. [Google Scholar] [CrossRef] [PubMed]
- Lai, A.; Gansau, J.; Gullbrand, S.E.; Crowley, J.; Cunha, C.; Dudli, S.; Engiles, J.B.; Fusellier, M.; Goncalves, R.M.; Nakashima, D.; et al. Development of a standardized histopathology scoring system for intervertebral disc degeneration in rat models: An initiative of the ors spine section. JOR Spine 2021, 4, e1150. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; An, S.B.; Lee, S.-H.; Lee, J.J.; Kim, S.B.; Ahn, J.-C.; Hwang, D.-Y.; Han, I. Enhanced Intervertebral Disc Repair via Genetically Engineered Mesenchymal Stem Cells with Tetracycline Regulatory System. Int. J. Mol. Sci. 2023, 24, 16024. https://doi.org/10.3390/ijms242216024
Kim Y, An SB, Lee S-H, Lee JJ, Kim SB, Ahn J-C, Hwang D-Y, Han I. Enhanced Intervertebral Disc Repair via Genetically Engineered Mesenchymal Stem Cells with Tetracycline Regulatory System. International Journal of Molecular Sciences. 2023; 24(22):16024. https://doi.org/10.3390/ijms242216024
Chicago/Turabian StyleKim, Yeji, Seong Bae An, Sang-Hyuk Lee, Jong Joo Lee, Sung Bum Kim, Jae-Cheul Ahn, Dong-Youn Hwang, and Inbo Han. 2023. "Enhanced Intervertebral Disc Repair via Genetically Engineered Mesenchymal Stem Cells with Tetracycline Regulatory System" International Journal of Molecular Sciences 24, no. 22: 16024. https://doi.org/10.3390/ijms242216024
APA StyleKim, Y., An, S. B., Lee, S. -H., Lee, J. J., Kim, S. B., Ahn, J. -C., Hwang, D. -Y., & Han, I. (2023). Enhanced Intervertebral Disc Repair via Genetically Engineered Mesenchymal Stem Cells with Tetracycline Regulatory System. International Journal of Molecular Sciences, 24(22), 16024. https://doi.org/10.3390/ijms242216024