
WISP1 Is Involved in the Pathogenesis of Kashin-Beck Disease via the Autophagy Pathway

 , , and
, , and 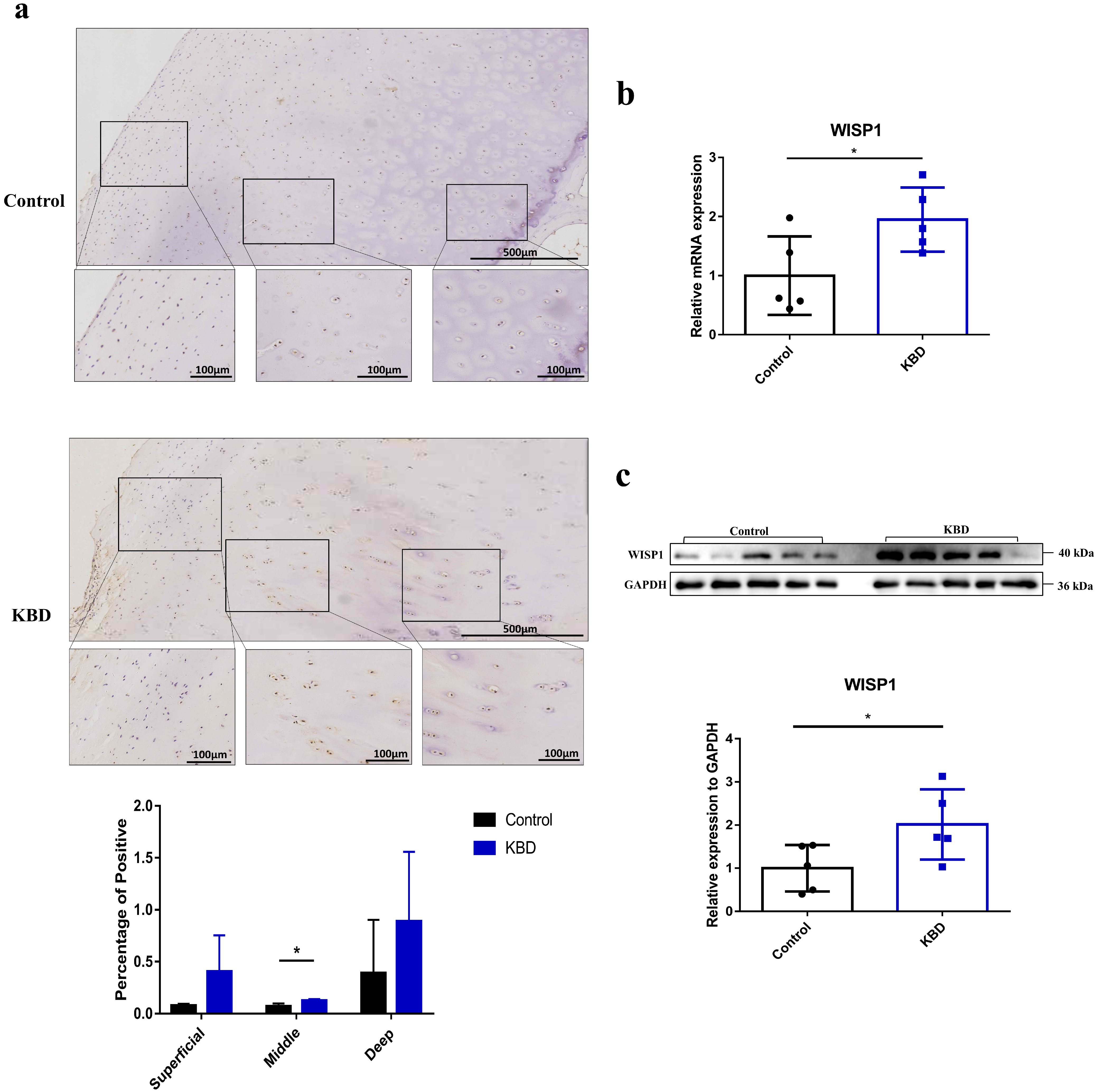{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Expression of WISP1 in KBD and Normal Control Articular Cartilage
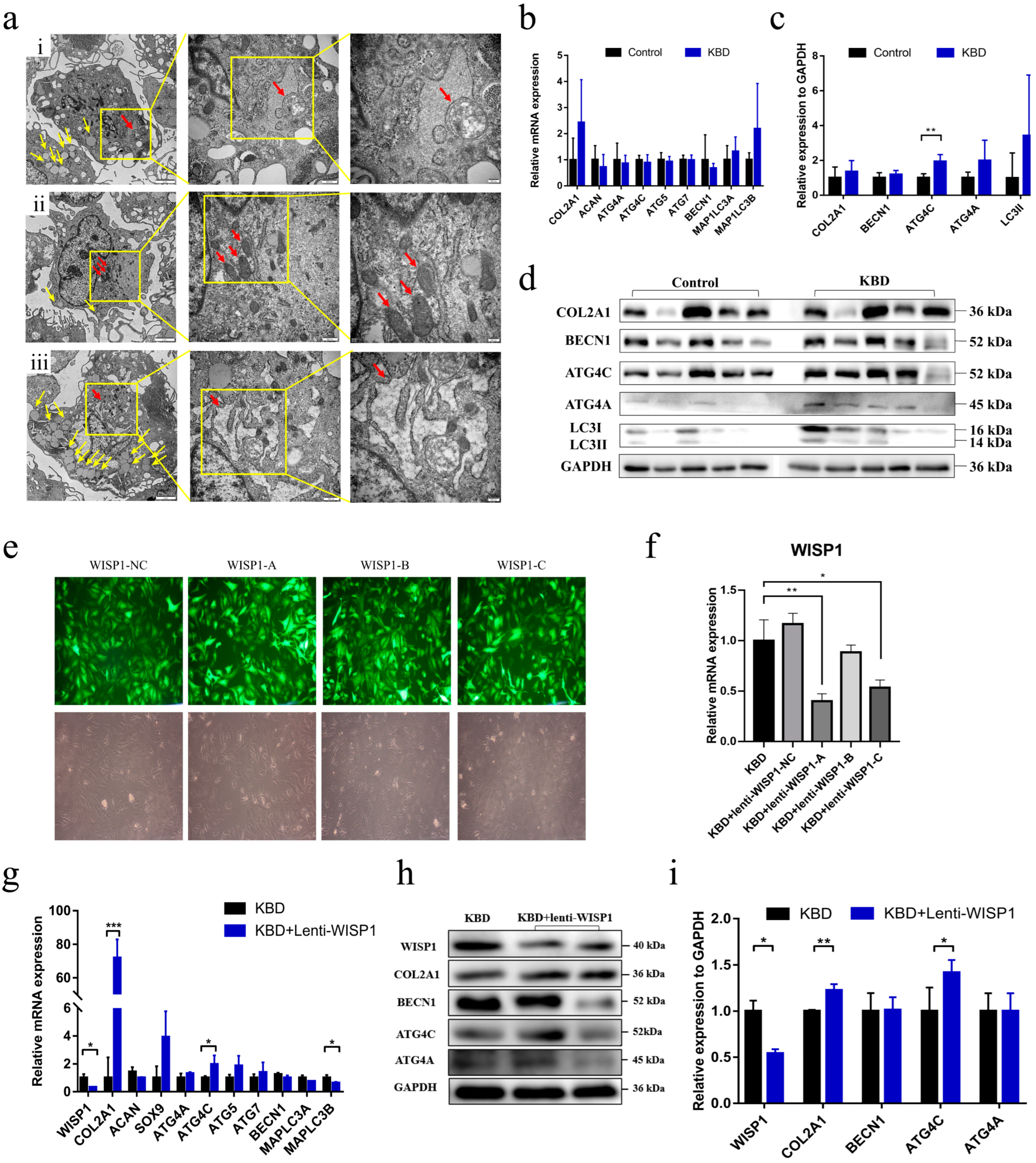
2.2. The Exploration of the Functional Role of WISP1 in KBD Chondrocyte Damage
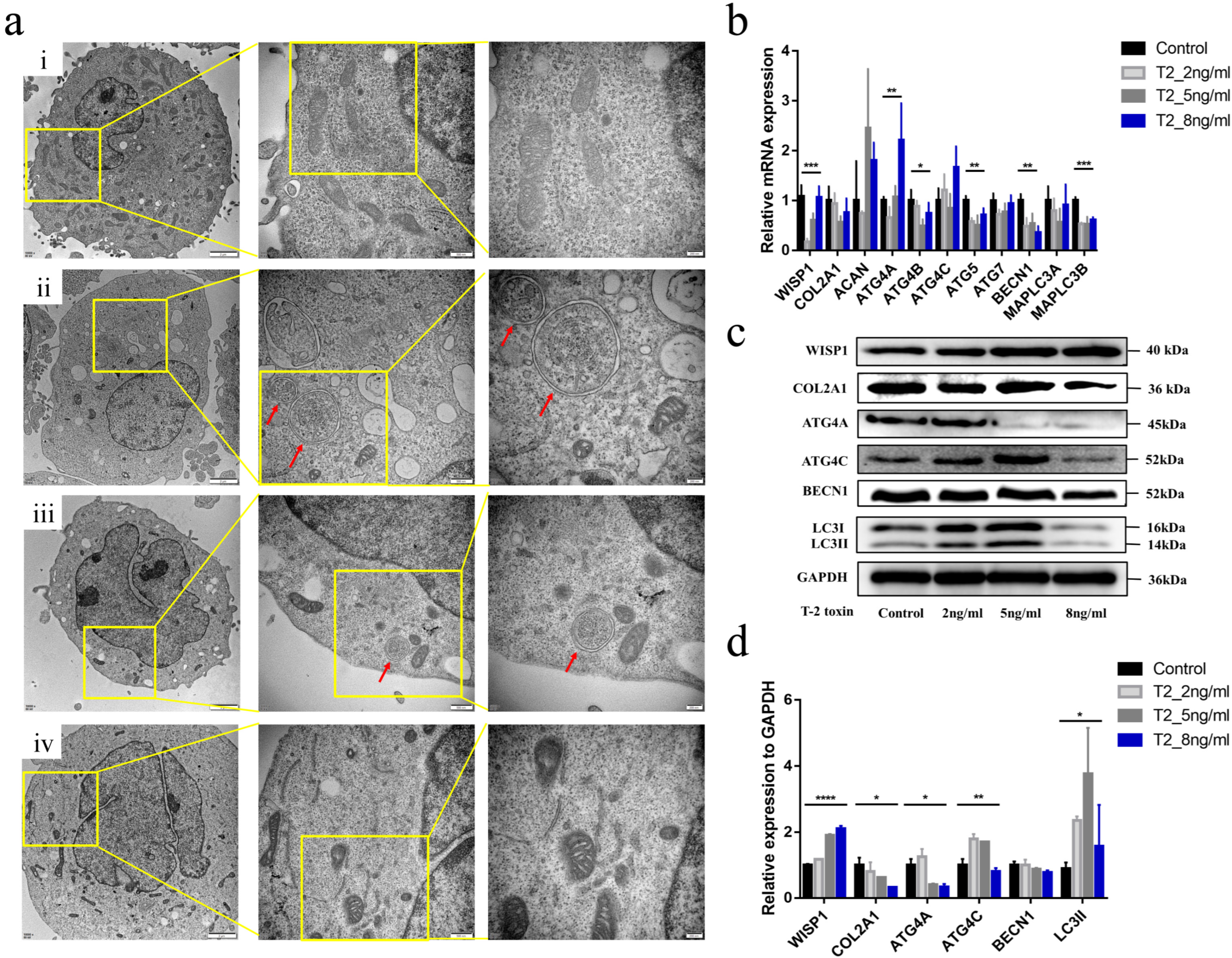
2.3. Effects of T-2 Toxin Intervention on WISP1 Expression in C28/I2 Human Chondrocytes
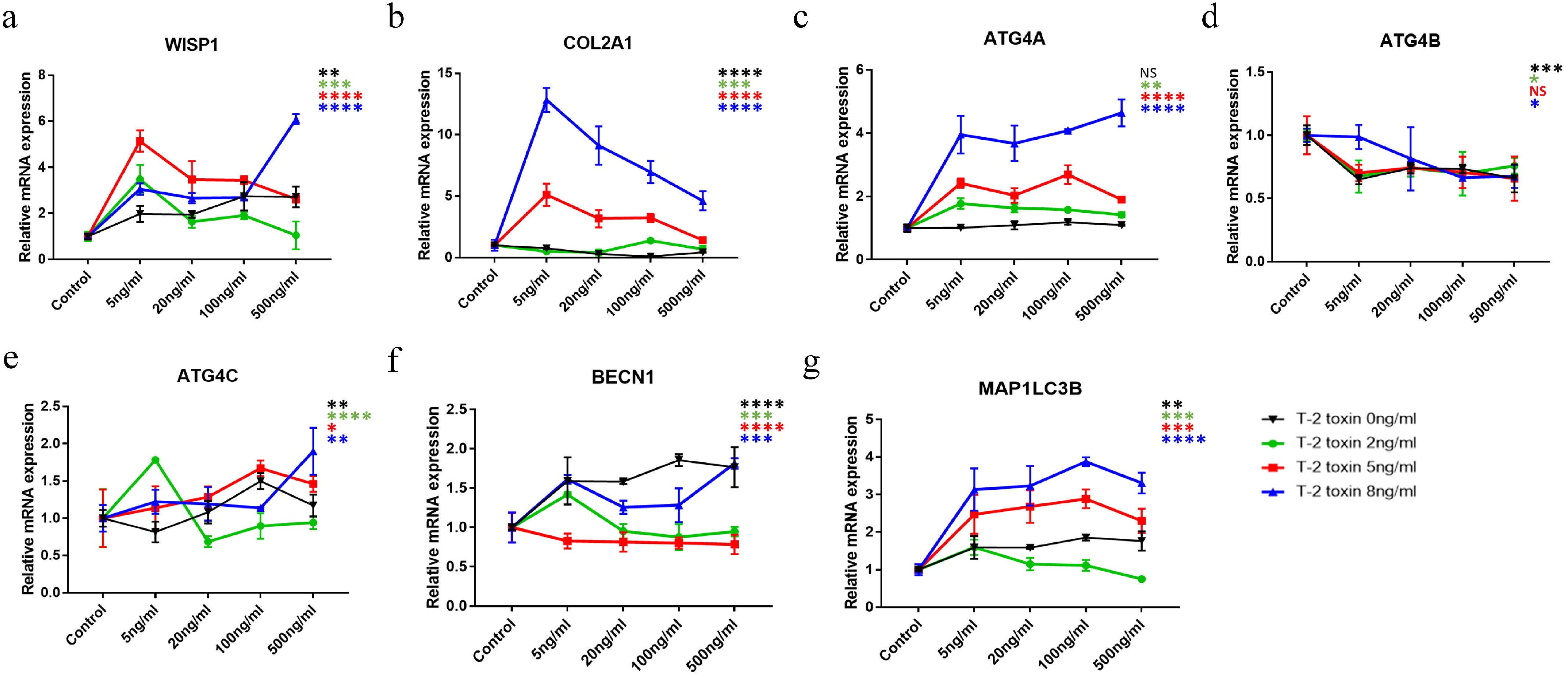
2.4. The Combined Effect of WISP1 and T-2 Toxin on Chondrocytes
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Specimen Collection
4.3. Chondrocyte Culture
4.4. Tissue Preparation and Hematoxylin-Eosin (H&E) Staining
4.5. IHC
4.6. Morphology Observation by Transmission Electron Microscope (TEM)
4.7. Cell Viability Detection by the Methyl-Thiazol-Tetrazolium (MTT) Assay
4.8. Intervention of rWISP1 Protein in C28/I2 Human Chondrocytes
4.9. WISP1 Knock-Down Cell Model by Lentiviral Transduction
4.10. mRNA Detection by qPCR
4.11. Protein Detection by Western Blot
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, X.; Ma, W.J.; Zhang, F.; Ren, F.L.; Qu, C.J.; Lammi, M.J. Recent advances in the research of an endemic osteochondropathy in China: Kashin-Beck disease. Osteoarthr. Cartil. 2014, 22, 1774–1783. [Google Scholar] [CrossRef] [PubMed]
- Lü, A.L.; Guo, X.; Aisha, M.M.; Shi, X.W.; Zhang, Y.; Zhang, Y.Y. Kashin-Beck disease and Sayiwak disease in China: Prevalence and a comparison of the clinical manifestations, familial aggregation, and heritability. Bone 2011, 48, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, C.; Guo, X.; Wen, Y.; Liu, L.; Liang, X.; Du, Y.; Zhang, L.; Ma, M.; Cheng, S.; et al. Integrative Analysis of Genome-Wide Association Studies and DNA Methylation Profile Identified Genetic Control Genes of DNA Methylation for Kashin-Beck Disease. Cartilage 2021, 13 (Suppl. S1), 780s–788s. [Google Scholar] [CrossRef] [PubMed]
- Gurbuz, I.; Chiquet-Ehrismann, R. CCN4/WISP1 (WNT1 inducible signaling pathway protein 1): A focus on its role in cancer. Int. J. Biochem. Cell Biol. 2015, 62, 142–146. [Google Scholar] [CrossRef]
- van den Bosch, M.H.; Gleissl, T.A.; Blom, A.B.; van den Berg, W.B.; van Lent, P.L.; van der Kraan, P.M. Wnts talking with the TGF-β superfamily: WISPers about modulation of osteoarthritis. Rheumatology 2016, 55, 1536–1547. [Google Scholar] [CrossRef]
- Blom, A.B.; Brockbank, S.M.; van Lent, P.L.; van Beuningen, H.M.; Geurts, J.; Takahashi, N.; van der Kraan, P.M.; van de Loo, F.A.; Schreurs, B.W.; Clements, K.; et al. Involvement of the Wnt signaling pathway in experimental and human osteoarthritis: Prominent role of Wnt-induced signaling protein 1. Arthritis Rheum. 2009, 60, 501–512. [Google Scholar] [CrossRef]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. Tuberous sclerosis protein 2 (TSC2) modulates CCN4 cytoprotection during apoptotic amyloid toxicity in microglia. Curr. Neurovascular Res. 2013, 10, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. Wnt1 inducible signaling pathway protein 1 (WISP1) targets PRAS40 to govern beta-amyloid apoptotic injury of microglia. Curr. Neurovascular Res. 2012, 9, 239–249. [Google Scholar] [CrossRef]
- Wu, C.; Wen, Y.; Guo, X.; Yang, T.; Shen, H.; Chen, X.; Tian, Q.; Tan, L.; Deng, H.W.; Zhang, F. Genetic association, mRNA and protein expression analysis identify ATG4C as a susceptibility gene for Kashin-Beck disease. Osteoarthr. Cartil. 2017, 25, 281–286. [Google Scholar] [CrossRef]
- Wu, C.; Zheng, J.; Yao, X.; Shan, H.; Li, Y.; Xu, P.; Guo, X. Defective autophagy in chondrocytes with Kashin-Beck disease but higher than osteoarthritis. Osteoarthr. Cartil. 2014, 22, 1936–1946. [Google Scholar] [CrossRef]
- Srinivas, V.; Shapiro, I.M. Chondrocytes embedded in the epiphyseal growth plates of long bones undergo autophagy prior to the induction of osteogenesis. Autophagy 2006, 2, 215–216. [Google Scholar] [CrossRef] [PubMed]
- Sokoloff, L. Kashin-Beck disease: Current status. Nutr. Rev. 1988, 46, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ning, Y.; Zhang, P.; Yang, L.; Wang, Y.; Guo, X. Chondrocytes damage induced by T-2 toxin via Wnt/β-catenin signaling pathway is involved in the pathogenesis of an endemic osteochondropathy, Kashin-Beck disease. Exp. Cell Res. 2017, 361, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Han, J.; Guo, X.; Qu, C.; Yu, F.; Wu, X. The effects of T-2 toxin on the prevalence and development of Kashin-Beck disease in China: A meta-analysis and systematic review. Toxicol. Res. 2016, 5, 731–751. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, H.; Tu, Y.; Li, Y.; Zou, Y.; Li, G.; Wang, L.; Zhong, X. WNT1-inducible signaling protein-1 mediates TGF-β1-induced renal fibrosis in tubular epithelial cells and unilateral ureteral obstruction mouse models via autophagy. J. Cell. Physiol. 2020, 235, 2009–2022. [Google Scholar] [CrossRef] [PubMed]
- Agrotis, A.; Pengo, N.; Burden, J.J.; Ketteler, R. Redundancy of human ATG4 protease isoforms in autophagy and LC3/GABARAP processing revealed in cells. Autophagy 2019, 15, 976–997. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Noda, N.N. Autophagy-regulating protease Atg4: Structure, function, regulation and inhibition. J. Antibiot. 2017, 71, 72–78. [Google Scholar] [CrossRef]
- Li, M.; Hou, Y.; Wang, J.; Chen, X.; Shao, Z.M.; Yin, X.M. Kinetics comparisons of mammalian Atg4 homologues indicate selective preferences toward diverse Atg8 substrates. J. Biol. Chem. 2011, 286, 7327–7338. [Google Scholar] [CrossRef]
- Song, S.J.; Tao, J.J.; Li, S.F.; Qian, X.W.; Niu, R.W.; Wang, C.; Zhang, Y.H.; Chen, Y.; Wang, K.; Zhu, F.; et al. 17beta-estradiol attenuates rat articular chondrocyte injury by targeting ASIC1a-mediated apoptosis. Mol. Cell. Endocrinol. 2020, 505, 110742. [Google Scholar] [CrossRef]
- Zheng, K.; Li, Y.; Wang, S.; Wang, X.; Liao, C.; Hu, X.; Fan, L.; Kang, Q.; Zeng, Y.; Wu, X.; et al. Inhibition of autophagosome-lysosome fusion by ginsenoside Ro via the ESR2-NCF1-ROS pathway sensitizes esophageal cancer cells to 5-fluorouracil-induced cell death via the CHEK1-mediated DNA damage checkpoint. Autophagy 2016, 12, 1593–1613. [Google Scholar] [CrossRef]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar] [PubMed]
- van den Bosch, M.H.J.; Ramos, Y.F.M.; den Hollander, W.; Bomer, N.; Nelissen, R.; Bovée, J.; van den Berg, W.B.; van Lent, P.; Blom, A.B.; van der Kraan, P.M.; et al. Increased WISP1 expression in human osteoarthritic articular cartilage is epigenetically regulated and decreases cartilage matrix production. Rheumatology 2019, 58, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zou, N.; Wang, J.; Wang, K.W.; Li, F.Y.; Chen, F.X.; Sun, B.Y.; Sun, D.J. TGF-beta1/Smad3 Signaling Pathway Mediates T-2 Toxin-Induced Decrease of Type II Collagen in Cultured Rat Chondrocytes. Toxins 2017, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Poole, A.R.; Kobayashi, M.; Yasuda, T.; Laverty, S.; Mwale, F.; Kojima, T.; Sakai, T.; Wahl, C.; El-Maadawy, S.; Webb, G.; et al. Type II collagen degradation and its regulation in articular cartilage in osteoarthritis. Ann. Rheum. Dis. 2002, 61 (Suppl. S2), ii78–ii81. [Google Scholar] [CrossRef] [PubMed]
- Pascarelli, N.A.; Collodel, G.; Moretti, E.; Cheleschi, S.; Fioravanti, A. Changes in Ultrastructure and Cytoskeletal Aspects of Human Normal and Osteoarthritic Chondrocytes Exposed to Interleukin-1β and Cyclical Hydrostatic Pressure. Int. J. Mol. Sci. 2015, 16, 26019–26034. [Google Scholar] [CrossRef]
- Claassen, H.; Schicht, M.; Brandt, J.; Reuse, K.; Schadlich, R.; Goldring, M.B.; Guddat, S.S.; Thate, A.; Paulsen, F. C-28/I2 and T/C-28a2 chondrocytes as well as human primary articular chondrocytes express sex hormone and insulin receptors--Useful cells in study of cartilage metabolism. Ann. Anat. Anat. Anz. Off. Organ Anat. Ges. 2011, 193, 23–29. [Google Scholar] [CrossRef]
- Li, P.; Ning, Y.; Wang, W.; Guo, X.; Poulet, B.; Wang, X.; Wen, Y.; Han, J.; Hao, J.; Liang, X.; et al. The integrative analysis of DNA methylation and mRNA expression profiles confirmed the role of selenocompound metabolism pathway in Kashin-Beck disease. Cell Cycle 2020, 19, 2351–2366. [Google Scholar] [CrossRef]
- Li, F.J.; Wang, X.J.; Zhou, X.L. WISP-1 overexpression upregulates cell proliferation in human salivary gland carcinomas via regulating MMP-2 expression. OncoTargets Ther. 2016, 9, 6539–6548. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Cheng, B.; Yao, Y.; Yu, W.; Liu, L.; Cheng, S.; Zhang, L.; Ma, M.; Qi, X.; Liang, C.; et al. WISP1 Is Involved in the Pathogenesis of Kashin-Beck Disease via the Autophagy Pathway. Int. J. Mol. Sci. 2023, 24, 16037. https://doi.org/10.3390/ijms242216037
Li P, Cheng B, Yao Y, Yu W, Liu L, Cheng S, Zhang L, Ma M, Qi X, Liang C, et al. WISP1 Is Involved in the Pathogenesis of Kashin-Beck Disease via the Autophagy Pathway. International Journal of Molecular Sciences. 2023; 24(22):16037. https://doi.org/10.3390/ijms242216037
Chicago/Turabian StyleLi, Ping, Bolun Cheng, Yao Yao, Wenxing Yu, Li Liu, Shiqiang Cheng, Lu Zhang, Mei Ma, Xin Qi, Chujun Liang, and et al. 2023. "WISP1 Is Involved in the Pathogenesis of Kashin-Beck Disease via the Autophagy Pathway" International Journal of Molecular Sciences 24, no. 22: 16037. https://doi.org/10.3390/ijms242216037
APA StyleLi, P., Cheng, B., Yao, Y., Yu, W., Liu, L., Cheng, S., Zhang, L., Ma, M., Qi, X., Liang, C., Chu, X., Ye, J., Sun, S., Jia, Y., Guo, X., Wen, Y., & Zhang, F. (2023). WISP1 Is Involved in the Pathogenesis of Kashin-Beck Disease via the Autophagy Pathway. International Journal of Molecular Sciences, 24(22), 16037. https://doi.org/10.3390/ijms242216037