
Effect of a Total Extract and Saponins from Astragalus glycyphyllos L. on Human Coronavirus Replication In Vitro

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
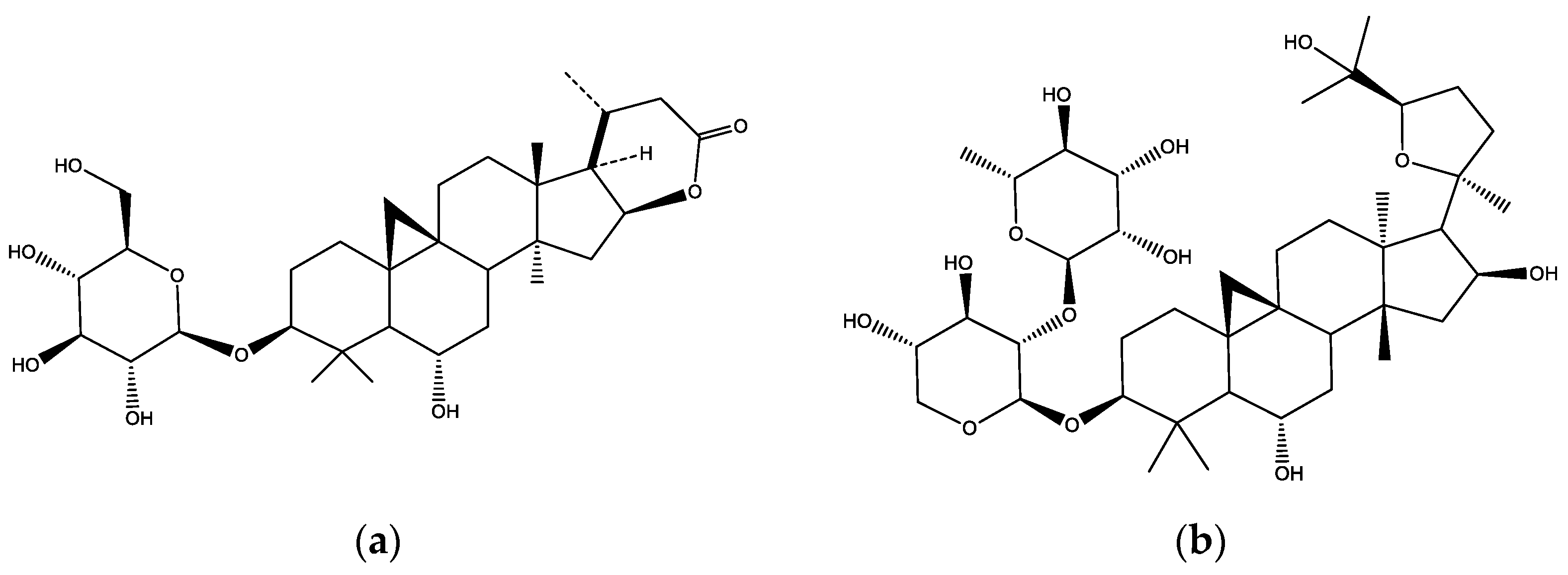
2.1. Phytochemistry of DEAG, PSM, and Saponins S1, S3, S9
2.2. Cultivation of Human Coronavirus 229E in Cell Culture
2.3. Cytotoxicity Assay
2.4. Antiviral Assay
3. Discussion
4. Materials and Methods
4.1. Preparation of the Dried Extract (DEAG)
4.2. Preparation of the Purified Saponin Mixture (PSM)
4.3. Isolation of Saponins S1, S3 and S9
4.4. Analysis of DEAG and PSM
4.5. Cells
4.6. Cultivation of Human Coronavirus 229E in Cell Culture
4.7. Cytotoxicity Assay
4.8. MTT-Based Colorimetric Assay for Detection of HcoV-229E Replication Inhibition
- Control cells (not infected with virus and not treated with extract)—0.2 mL of supporting nutrient medium are added to the wells designated for cell control (at least 3);
- Virus control (virus-infected and extract-untreated cells)—to the wells designated for virus control (at least three), 0.1 mL of supporting nutrient medium is instilled;
- Cells exposed to the extract—(infected with a virus and treated with different dilutions of the studied extract)—0.1 mL of the previously prepared dilutions of the extract.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Xu, H.; Yan, C.; Fu, Q.; Xiao, K.; Yu, Y.; Han, D.; Wang, W.; Cheng, J. Possible environmental effects on the spread of COVID-19 in China. Sci. Total Environ. 2020, 731, 139211. [Google Scholar] [CrossRef]
- Salata, C.; Calistri, A.; Parolin, C.; Palu, G. Coronaviruses: A paradigm of new emerging zoonotic diseases. Pathog. Dis. 2019, 77, ftaa006. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Hua, F.; Bian, Z. Coronavirus disease 2019 (COVID-19): Emerging and future challenges for dental and oral medicine. J. Dent. Res. 2020, 99, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Sun, X.; Ye, J.; Ding, L.; Liu, M.; Yang, Z.; Lu, X.; Zhang, Y.; Ma, L.; Gu, W.; et al. Key residues of the receptor binding motif in the spike protein of SARS-CoV-2 that interact with ACE2 and neutralizing antibodies. Cell Mol. Immunol. 2020, 17, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Roldan, E.Q.; Biasiotto, G.; Magro, P.; Zanella, I. The possible mechanisms of action of 4-aminoquinolines (chloroquine/hydroxychloroquine) against SARS-CoV-2 infection (COVID-19): A role for iron homeostasis? Pharmacol. Res. 2020, 158, 104904. [Google Scholar] [CrossRef]
- Arunkumar, G.; Mudgal, P.P.; Maity, H.; Dowarha, D.; Devadiga, S.; Nag, S.; Arunkumar, G. Herbal plants and plant preparations as remedial approach for viral diseases. Virusdisease 2015, 26, 225–236. [Google Scholar] [CrossRef]
- Ben-Shabat, S.; Yarmolinsky, L.; Porat, D.; Dahan, A. Antiviral effect of phytochemicals from medicinal plants: Applications and drug delivery strategies. Drug Deliv. Transl. Res. 2020, 10, 354–367. [Google Scholar] [CrossRef]
- Joshi, B.; Panda, S.K.; Jouneghani, R.S.; Liu, M.; Parajuli, N.; Leyssen, P.; Neyts, J.; Luyten, W. Antibacterial, antifungal, antiviral, and anthelmintic activities of medicinal plants of Nepal selected based on ethnobotanical evidence. Evid. Based Complement. Altern. Med. 2020, 2020, 1043471. [Google Scholar] [CrossRef] [PubMed]
- Zambounis, A.; Sytar, O.; Valasiadis, D.; Hilioti, Z. Effect of photosensitisers on growth and morphology of Phytophthora citrophthora coupled with leaf bioassays in pear seedlings. Plant Protect. Sci. 2020, 56, 74–82. [Google Scholar] [CrossRef]
- Ríos, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef]
- Ghildiyal, R.; Prakash, V.; Chaudhary, V.K.; Gupta, V.; Gabrani, R. Phytochemicals as Antiviral Agents: Recent Updates. In Plant-Derived Bioactives; Swamy, M., Ed.; Springer: Singapore, 2020; pp. 279–295. [Google Scholar] [CrossRef]
- Adhikari, B.; Marasini, B.P.; Rayamajhee, B.; Bhattarai, B.R.; Lamichhane, G.; Khadayat, K.; Adhikari, A.; Khanal, S.; Parajuli, N. Potential roles of medicinal plants for the treatment of viral diseases focusing on COVID-19: A review. Phytother. Res. 2021, 35, 1298–1312. [Google Scholar] [CrossRef]
- Wen, C.C.; Shyur, L.F.; Jan, J.T.; Liang, P.H.; Kuo, C.J.; Arulselvan, P.; Wu, J.B.; Kuo, S.C.; Yang, N.S. Traditional Chinese medicine herbal extracts of Cibotium barometz, Gentiana scabra, Dioscorea batatas, Cassia tora, and Taxillus chinensis inhibit SARS-CoV replication. J. Tradit. Complement. Med. 2011, 1, 41–50. [Google Scholar] [CrossRef]
- Kim, H.Y.; Eo, E.Y.; Park, H.; Kim, Y.C.; Park, S.; Shin, H.J.; Kim, K. Medicinal herbal extracts of sophorae radix ACANTHOPANACIS cortex sanguisorbae radix and Torilis fructus inhibit coronavirus replication in vitro. Antivir. Ther. 2010, 15, 697–709. [Google Scholar] [CrossRef]
- Kim, H.Y.; Shin, H.S.; Park, H.; Kim, Y.C.; Yun, Y.G.; Park, S.; Shin, H.J.; Kim, K. In vitro inhibition of coronavirus replications by the traditionally used medicinal herbal extracts, Cimicifuga rhizoma, Meliae cortex, Coptidis rhizoma, and Phellodendron cortex. J. Clin. Virol. 2008, 41, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Carneiro, J.N.P.; Rocha, J.E.; Coutinho, H.D.M.; Braga, M.F.B.M.; Sharifi-Rad, J.; Semwal, P.; Painuli, S.; Moujir, L.M.; Machado, V.Z.; et al. Astragalus species: Insights on its chemical composition toward pharmacological applications. Phytother. Res. 2021, 35, 2445–2476. [Google Scholar] [CrossRef]
- Pistelli, L.F. Secondary metabolites of genus Astragalus: Structure and biological activity. In Studies in Natural Products Chemistry; Rahman, A.U., Ed.; Elsivier Science BV: Karachi, Pakistan, 2002; pp. 443–545. [Google Scholar]
- Zhang, P.; Liu, X.; Liu, H.; Wang, W.; Liu, X.; Li, X.; Wu, X. Astragalus polysaccharides inhibit avian infectious bronchitis virus infection by regulating viral replication. Microb. Pathog. 2018, 114, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Shkondrov, A.; Hinkov, A.; Cvetkov, V.; Shishkova, K.; Todorov, D.; Shishkov, S.; Stambolov, I.; Yoncheva, K.; Krasteva, I. Astragalus glycyphyllos L.: Antiviral Activity and Tablet Dosage Formulation of a Standardized Dry Extract. Biotechnol. Biotechnol. Equip. 2023, 37, 2221752. [Google Scholar] [CrossRef]
- Shkondrov, A.; Krasteva, I. High Resolution LC-MS/MS Screening for Secondary Metabolites in Bulgarian Species of Genus Astragalus L. Quim. Nova 2021, 44, 683–688. [Google Scholar] [CrossRef]
- Shkondrov, A.; Krasteva, I.; Ionkova, I.; Popova, P.; Zarev, Y.; Mihaylova, R.; Konstantinov, S. Production of saponins from in vitro cultures of Astragalus glycyphyllos and their antineoplastic activity. Biotechnol. Biotechnol. Equip. 2019, 33, 1413–1418. [Google Scholar] [CrossRef]
- Shkondrov, A.; Krasteva, I.; Bucar, F.; Kunert, O.; Kondeva-Burdina, M.; Ionkova, I. A New Tetracyclic Saponin from Astragalus glycyphyllos L. and its Neuroprotective and HMAO-B Inhibiting Activity. Nat. Prod. Res. 2020, 34, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Stambolov, I.; Shkondrov, A.; Kunert, O.; Bucar, F.; Kondeva-Burdina, M.; Krasteva, I. Cycloartane Saponins from Astragalus glycyphyllos and Their In Vitro Neuroprotective, Antioxidant, and hMAO-B-Inhibiting Effects. Metabolites 2023, 13, 857. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Yao, J.; Liu, J.; He, Y.; Liu, L.; Xu, Z.; Lin, X.; Liu, N.; Kai, G. Systems Pharmacology Reveals the Mechanism of Astragaloside IV in Improving Immune Activity on Cyclophosphamide-Induced Immunosuppressed Mice. J. Ethnopharmacol. 2023, 313, 116533. [Google Scholar] [CrossRef]
- Cho, W.C.S.; Leung, K.N. In Vitro and in Vivo Immunomodulating and Immunorestorative Effects of Astragalus membranaceus. J. Ethnopharmacol. 2007, 113, 132–141. [Google Scholar] [CrossRef]
- Hui, X.; Wang, S.; Lu, S.; Liu, Y.; Peng Zhang, Z.; Jing Lian, W.; Zhang, H.; Yuan Zhou, S.; Zhang, J.; Wang, J. Clinical Effect of Astragalus membranaceus Injection on Viral Myocarditis in Children: An Overview of Systematic Reviews and Evidence Mapping. Pharmacol. Res.—Mod. Chin. Med. 2023, 8, 100270. [Google Scholar] [CrossRef]
- Handa, R.K.; Harding, J.W.; Simasko, S.M. Characterization and function of the bovine kidney epithelial angiotensin receptor subtype 4 using angiotensin IV and divalinal angiotensin IV as receptor ligands. J. Pharmacol. Exp. Ther. 1999, 291, 1242–1249. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Sudo, K.; Konno, K.; Yokota, T.; Shigeta, S. A sensitive assay system screening antiviral compounds against herpes simplex virus type 1 and type 2. J. Virol. Methods 1994, 49, 169–178. [Google Scholar] [CrossRef]



 ) and one and a half hours after the inoculation of cell monolayer (
) and one and a half hours after the inoculation of cell monolayer ( ) with HCoV 229E; (b) PSM added simultaneously with the inoculation of cell monolayer (
) with HCoV 229E; (b) PSM added simultaneously with the inoculation of cell monolayer ( ) and one and a half hours after the inoculation of cell monolayer (
) and one and a half hours after the inoculation of cell monolayer ( ) with HCoV 229E.
) and one and a half hours after the inoculation of cell monolayer () with HCoV 229E; (b) PSM added simultaneously with the inoculation of cell monolayer () and one and a half hours after the inoculation of cell monolayer () with HCoV 229E.
) with HCoV 229E.
) and one and a half hours after the inoculation of cell monolayer () with HCoV 229E; (b) PSM added simultaneously with the inoculation of cell monolayer () and one and a half hours after the inoculation of cell monolayer () with HCoV 229E.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Test Sample | Cell Viability (Cytotoxicity) | Cell Protection (Antiviral Activity) | ||||||
|---|---|---|---|---|---|---|---|---|
| Test Sample Added Simultaneously with Inoculation of Cell Monolayer | Test Sample Added 1.5 h after Inoculation of Cell Monolayer | |||||||
| MTC (mg/mL) | CC50 a,b (mg/mL) | Cell Protection (%) When the Extracts are Added in MTC a | EC50 a,b (mg/mL) | SI c | Cell Protection (%) When the Extracts Are Added in MTC a | EC50 a,b (mg/mL) | SI c | |
| DEAG | 1 | 1.272 (±0.11) | 100 | 0.091 (±0.06) | 13.97 | 100 | 0.114 (±0.10) | 11.15 |
| PSM | 0.025 | 0.0748 (±0.18) | 100 | 0.00432 (±0.14) | 17.31 | 100 | 0.00418 (±0.31) | 17.89 |
| S1 | 0.00625 | 0.117 (±0.03) | n.d. d | n.d. d | n.d. d | n.d. d | n.d. d | n.d. d |
| S3 | 0.025 | 0.163 (±0.21) | n.d. d | n.d. d | n.d. d | n.d. d | n.d. d | n.d. d |
| S9 | 0.00625 | 0.104 (±0.53) | n.d. d | n.d. d | n.d. d | n.d. d | n.d. d | n.d. d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinkov, A.; Tsvetkov, V.; Shkondrov, A.; Krasteva, I.; Shishkov, S.; Shishkova, K. Effect of a Total Extract and Saponins from Astragalus glycyphyllos L. on Human Coronavirus Replication In Vitro. Int. J. Mol. Sci. 2023, 24, 16525. https://doi.org/10.3390/ijms242216525
Hinkov A, Tsvetkov V, Shkondrov A, Krasteva I, Shishkov S, Shishkova K. Effect of a Total Extract and Saponins from Astragalus glycyphyllos L. on Human Coronavirus Replication In Vitro. International Journal of Molecular Sciences. 2023; 24(22):16525. https://doi.org/10.3390/ijms242216525
Chicago/Turabian StyleHinkov, Anton, Venelin Tsvetkov, Aleksandar Shkondrov, Ilina Krasteva, Stoyan Shishkov, and Kalina Shishkova. 2023. "Effect of a Total Extract and Saponins from Astragalus glycyphyllos L. on Human Coronavirus Replication In Vitro" International Journal of Molecular Sciences 24, no. 22: 16525. https://doi.org/10.3390/ijms242216525
APA StyleHinkov, A., Tsvetkov, V., Shkondrov, A., Krasteva, I., Shishkov, S., & Shishkova, K. (2023). Effect of a Total Extract and Saponins from Astragalus glycyphyllos L. on Human Coronavirus Replication In Vitro. International Journal of Molecular Sciences, 24(22), 16525. https://doi.org/10.3390/ijms242216525