


Folate-Appended Hydroxypropyl-β-Cyclodextrin Induces Autophagic Cell Death in Acute Myeloid Leukemia Cells

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
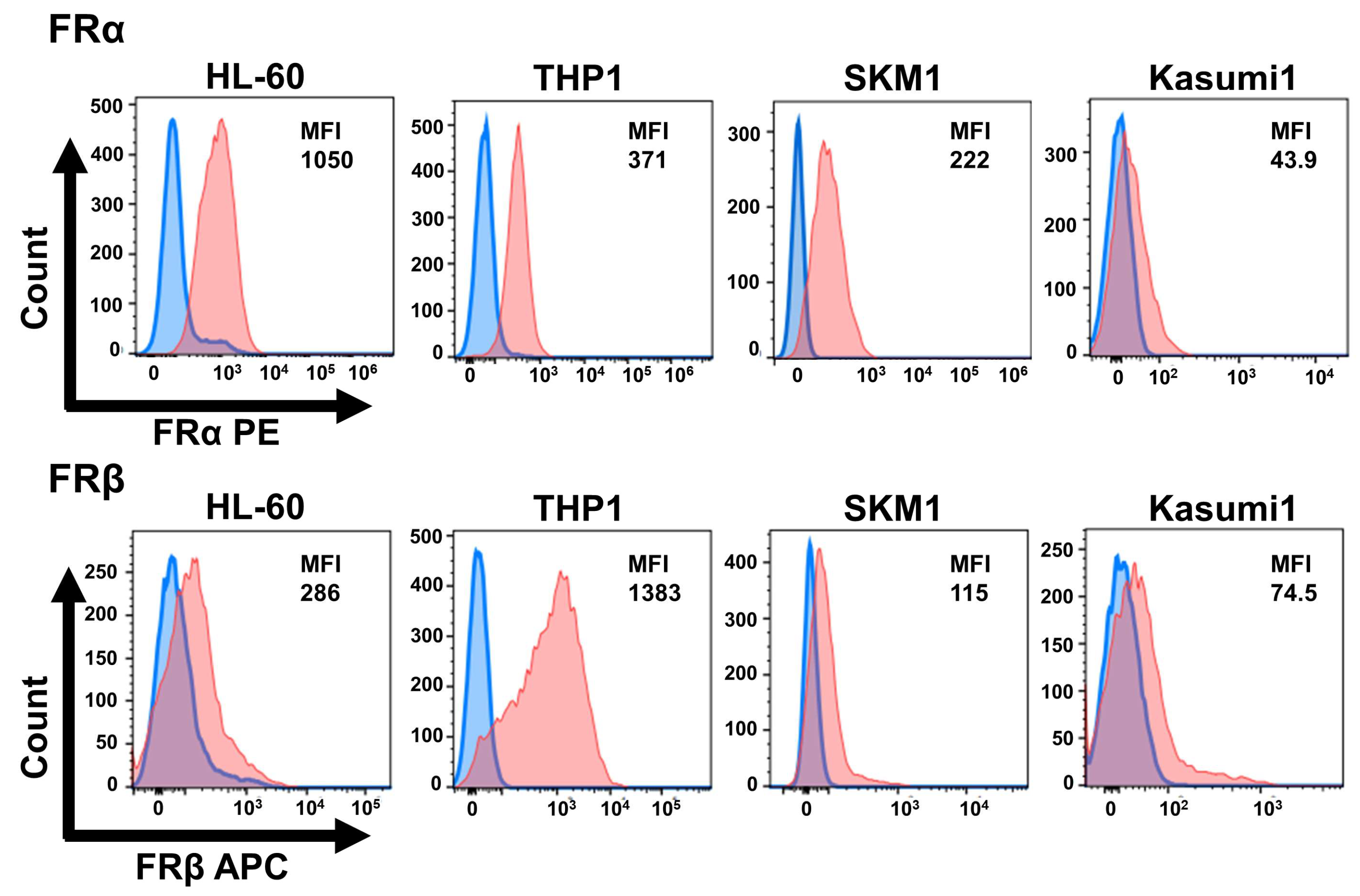
2.1. AML Cell Lines Express High Levels of the FR
2.2. FA-HP-β-CyD Inhibits Growth of AML Cell Lines More Potently Than HP-β-CyD
2.3. FA-HP-β-CyD Inhibits Cell Growth by Inducing Apoptosis
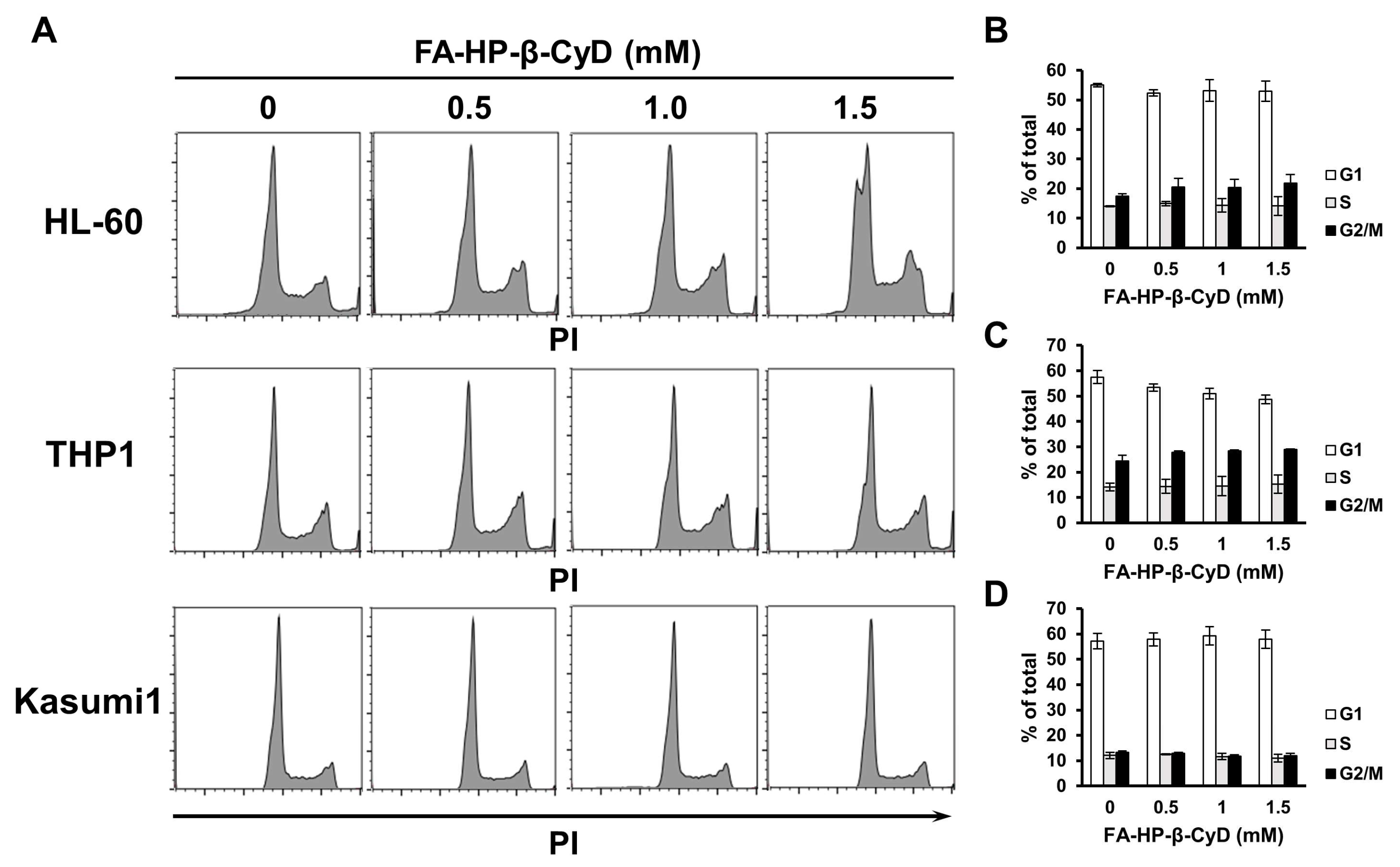
2.4. FA-HP-β-CyD Does Not Affect the Cell Cycle Status of AML Cells
2.5. FA-HP-β-CyD Induces Autophagosome Formation
2.6. Inhibiting Autophagy Dampens the Antitumor Activity of FA-HP-β-CyD
2.7. FA-HP-β-CyD Acts Synergistically to Enhance the Effects of Ara-C and Venetoclax
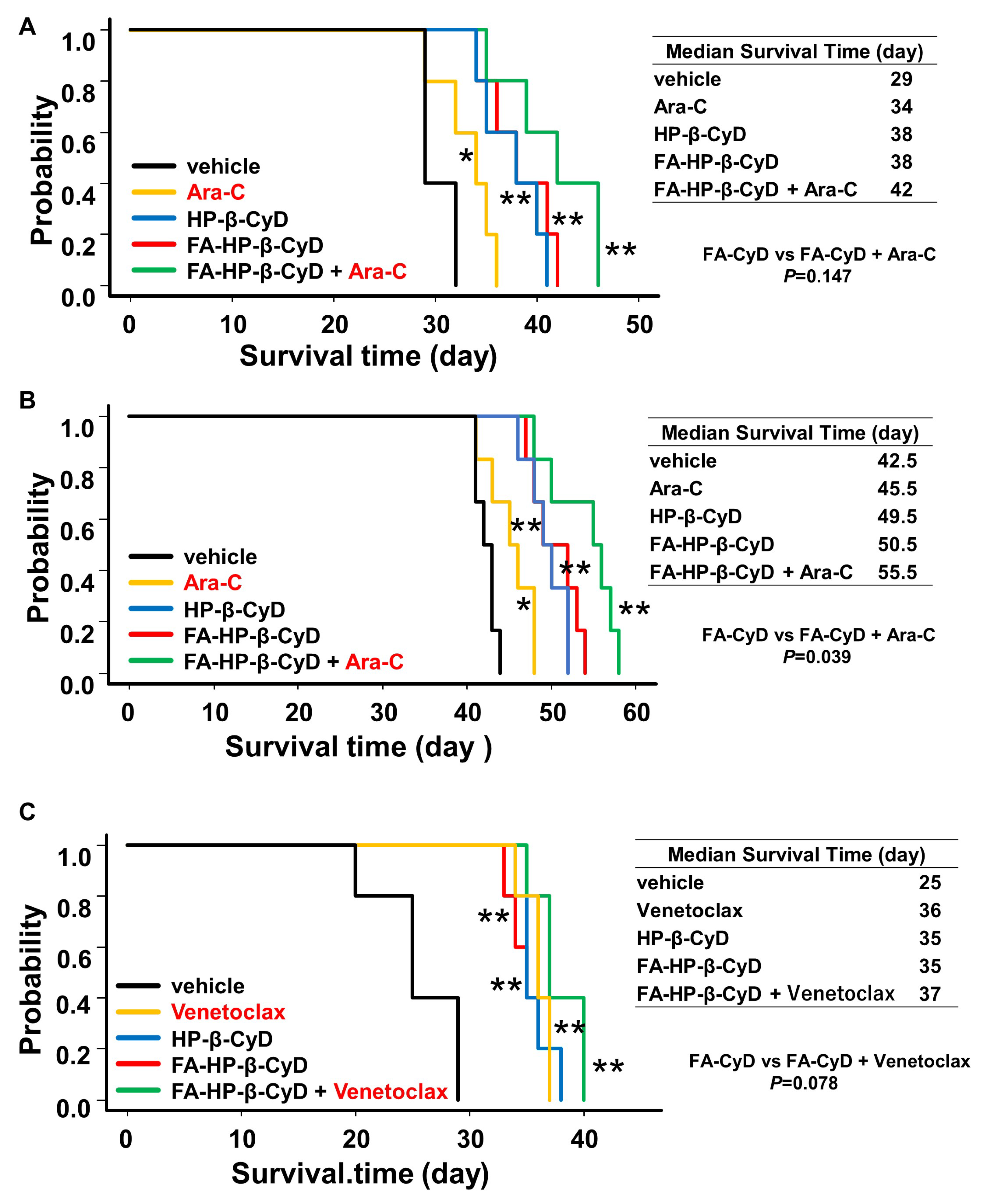
2.8. FA-HP-β-CyD Prolongs Survival of Acute Myeloid Leukemia Xenografts
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Cultures
4.3. Mice
4.4. Analysis of Cell Growth and Viability
4.5. Flow Cytometry Analysis
4.6. Cell Cycle Analysis
4.7. Autophagy Detection
4.8. Culture of Cells with Autophagy Inhibitors
4.9. Murine Leukemia Model
4.10. Blood Counts
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DiNardo, C.D.; Erba, H.P.; Freeman, S.D.; Wei, A.H. Acute myeloid leukaemia. Lancet 2023, 401, 2073–2086. [Google Scholar] [CrossRef]
- Ohtake, S.; Miyawaki, S.; Fujita, H.; Kiyoi, H.; Shinagawa, K.; Usui, N.; Okumura, H.; Miyamura, K.; Nakaseko, C.; Miyazaki, Y.; et al. Randomized study of induction therapy comparing standard-dose idarubicin with high-dose daunorubicin in adult patients with previously untreated acute myeloid leukemia: The JALSG AML201 Study. Blood 2011, 117, 2358–2365. [Google Scholar] [CrossRef]
- Choi, J.; Shukla, M.; Abdul-Hay, M. Acute Myeloid Leukemia Treatment in the Elderly: A Comprehensive Review of the Present and Future. Acta Haematol. 2023. [Google Scholar] [CrossRef]
- DiNardo, C.D.; Pratz, K.; Pullarkat, V.; Jonas, B.A.; Arellano, M.; Becker, P.S.; Frankfurt, O.; Konopleva, M.; Wei, A.H.; Kantarjian, H.M.; et al. Venetoclax combined with decitabine or azacitidine in treatment-naive, elderly patients with acute myeloid leukemia. Blood 2019, 133, 7–17. [Google Scholar] [CrossRef]
- DiNardo, C.D.; Jonas, B.A.; Pullarkat, V.; Thirman, M.J.; Garcia, J.S.; Wei, A.H.; Konopleva, M.; Döhner, H.; Letai, A.; Fenaux, P.; et al. Azacitidine and Venetoclax in Previously Untreated Acute Myeloid Leukemia. N. Engl. J. Med. 2020, 383, 617–629. [Google Scholar] [CrossRef]
- Wei, A.H.; Montesinos, P.; Ivanov, V.; DiNardo, C.D.; Novak, J.; Laribi, K.; Kim, I.; Stevens, D.A.; Fiedler, W.; Pagoni, M.; et al. Venetoclax plus LDAC for newly diagnosed AML ineligible for intensive chemotherapy: A phase 3 randomized placebo-controlled trial. Blood 2020, 135, 2137–2145. [Google Scholar] [CrossRef]
- Cherry, E.M.; Abbott, D.; Amaya, M.; McMahon, C.; Schwartz, M.; Rosser, J.; Sato, A.; Schowinsky, J.; Inguva, A.; Minhajuddin, M.; et al. Venetoclax and azacitidine compared with induction chemotherapy for newly diagnosed patients with acute myeloid leukemia. Blood Adv. 2021, 5, 5565–5573. [Google Scholar] [CrossRef]
- Freeman, T.; Williams, K.; Puto, M.; Waller, A.; McLaughlin, E.M.; Blachly, J.S.; Roddy, J. Real-World Experience of Adults with Acute Myeloid Leukemia on Hypomethylating Agents with or without Venetoclax at a Comprehensive Cancer Center. World J. Oncol. 2023, 14, 40–50. [Google Scholar] [CrossRef]
- Arber, D.A.; Orazi, A.; Hasserjian, R.P.; Borowitz, M.J.; Calvo, K.R.; Kvasnicka, H.M.; Wang, S.A.; Bagg, A.; Barbui, T.; Branford, S.; et al. International Consensus Classification of Myeloid Neoplasms and Acute Leukemias: Integrating morphologic, clinical, and genomic data. Blood 2022, 140, 1200–1228. [Google Scholar] [CrossRef]
- Khoury, J.D.; Solary, E.; Abla, O.; Akkari, Y.; Alaggio, R.; Apperley, J.F.; Bejar, R.; Berti, E.; Busque, L.; Chan, J.K.C.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Myeloid and Histiocytic/Dendritic Neoplasms. Leukemia 2022, 36, 1703–1719. [Google Scholar] [CrossRef]
- Döhner, H.; Wei, A.H.; Appelbaum, F.R.; Craddock, C.; DiNardo, C.D.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Godley, L.A.; Hasserjian, R.P.; et al. Diagnosis and management of AML in adults: 2022 recommendations from an international expert panel on behalf of the ELN. Blood 2022, 140, 1345–1377. [Google Scholar] [CrossRef]
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar] [CrossRef]
- Uekama, K.; Hirayama, F.; Irie, T. Cyclodextrin Drug Carrier Systems. Chem. Rev. 1998, 98, 2045–2076. [Google Scholar] [CrossRef]
- Szente, L.; Szejtli, J. Highly soluble cyclodextrin derivatives: Chemistry, properties, and trends in development. Adv. Drug Deliv. Rev. 1999, 36, 17–28. [Google Scholar] [CrossRef]
- George, K.S.; Wu, S. Lipid raft: A floating island of death or survival. Toxicol. Appl. Pharmacol. 2012, 259, 311–319. [Google Scholar] [CrossRef]
- Simons, K.; Ehehalt, R. Cholesterol, lipid rafts, and disease. J. Clin. Investig. 2002, 110, 597–603. [Google Scholar] [CrossRef]
- Galbiati, F.; Razani, B.; Lisanti, M.P. Emerging themes in lipid rafts and caveolae. Cell 2001, 106, 403–411. [Google Scholar] [CrossRef]
- Anderson, R.G.; Jacobson, K. A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science 2002, 296, 1821–1825. [Google Scholar] [CrossRef]
- Matsuo, M.; Togawa, M.; Hirabaru, K.; Mochinaga, S.; Narita, A.; Adachi, M.; Egashira, M.; Irie, T.; Ohno, K. Effects of cyclodextrin in two patients with Niemann-Pick Type C disease. Mol. Genet. Metab. 2013, 108, 76–81. [Google Scholar] [CrossRef]
- Yamada, Y.; Ishitsuka, Y.; Kondo, Y.; Nakahara, S.; Nishiyama, A.; Takeo, T.; Nakagata, N.; Motoyama, K.; Higashi, T.; Arima, H.; et al. Differential mode of cholesterol inclusion with 2-hydroxypropyl-cyclodextrins increases safety margin in treatment of Niemann-Pick disease type C. Br. J. Pharmacol. 2021, 178, 2727–2746. [Google Scholar] [CrossRef]
- Yokoo, M.; Kubota, Y.; Motoyama, K.; Higashi, T.; Taniyoshi, M.; Tokumaru, H.; Nishiyama, R.; Tabe, Y.; Mochinaga, S.; Sato, A.; et al. 2-Hydroxypropyl-beta-Cyclodextrin Acts as a Novel Anticancer Agent. PLoS ONE 2015, 10, e0141946. [Google Scholar] [CrossRef]
- Hoshiko, T.; Kubota, Y.; Onodera, R.; Higashi, T.; Yokoo, M.; Motoyama, K.; Kimura, S. Folic Acid-Appended Hydroxypropyl-beta-Cyclodextrin Exhibits Potent Antitumor Activity in Chronic Myeloid Leukemia Cells via Autophagic Cell Death. Cancers 2021, 13, 5413. [Google Scholar] [CrossRef]
- Parker, N.; Turk, M.J.; Westrick, E.; Lewis, J.D.; Low, P.S.; Leamon, C.P. Folate receptor expression in carcinomas and normal tissues determined by a quantitative radioligand binding assay. Anal. Biochem. 2005, 338, 284–293. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Leamon, C.P.; Reddy, J.A. The folate receptor as a rational therapeutic target for personalized cancer treatment. Drug Resist. Updat. 2014, 17, 89–95. [Google Scholar] [CrossRef]
- Roy, A.G.; Robinson, J.M.; Sharma, P.; Rodriguez-Garcia, A.; Poussin, M.A.; Nickerson-Nutter, C.; Powell, D.J., Jr. Folate Receptor Beta as a Direct and Indirect Target for Antibody-Based Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 5572. [Google Scholar] [CrossRef]
- Coney, L.R.; Tomassetti, A.; Carayannopoulos, L.; Frasca, V.; Kamen, B.A.; Colnaghi, M.I.; Zurawski, V.R., Jr. Cloning of a tumor-associated antigen: Mov18 and Mov19 antibodies recognize a folate-binding protein. Cancer Res. 1991, 51, 6125–6132. [Google Scholar] [PubMed]
- Ross, J.F.; Wang, H.; Behm, F.G.; Mathew, P.; Wu, M.; Booth, R.; Ratnum, M. Folate receptor type beta is a neutrophilic lineage marker and is differentially expressed in myeloid leukemia. Cancer 1999, 85, 348–357. [Google Scholar] [CrossRef]
- Ross, J.F.; Chaudhuri, P.K.; Ratnum, M. Differential regulation of folate receptor isoforms in normal and malignant tis sues in vivo and in established cell lines. Physiologic and clinical implications. Cancer 1994, 73, 2432–2443. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Ross, J.F.; Wang, X.; Raatnum, M. Identification of a novel folate receptor, a truncated receptor, and receptor type beta in hematopoietic cells: cDNA cloning, expression, immunoreactivity, and tissue specificity. Biochemistry 1994, 33, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Onodera, R.; Motoyama, K.; Tanaka, N.; Ohyama, A.; Okamatsu, A.; Higashi, T.; Kariya, R.; Okada, S.; Arima, H. Involvement of autophagy in antitumor activity of folate-appended methyl-beta-cyclodextrin. Sci. Rep. 2014, 4, 4417. [Google Scholar] [CrossRef]
- Motoyama, K.; Onodera, R.; Tanaka, N.; Kameyama, K.; Higashi, T.; Kariya, R.; Okada, S.; Arima, H. Evaluation of antitumor effects of folate-conjugated methyl-beta-cyclodextrin in melanoma. Biol. Pharm. Bull. 2015, 38, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.; Motoyama, K.; Tanaka, N.; Yamashita, Y.; Higashi, T.; Arima, H. Induction of mitophagy-mediated antitumor activity with folate-appended methyl-beta-cyclodextrin. Int. J. Nanomed. 2017, 12, 3433–3446. [Google Scholar] [CrossRef] [PubMed]
- Hoshiko, T.; Kubota, Y.; Akisawa, T.; Watanabe, T.; Tanigawara, K.; Yano, J.; Kimura, S. Naked antisense double-stranded DNA oligonucleotide efficiently suppresses BCR-ABL positive leukemic cells. Investig. New Drugs 2020, 38, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Sun, J.; Feng, L.; Wang, K.; Li, D.; Pan, Q.; Chen, Y.; Jin, W.; Wang, X.; Pan, H.; et al. Autophagy inhibition enhances daunorubicin-induced apoptosis in K562 cells. PLoS ONE 2011, 6, e28491. [Google Scholar] [CrossRef]
- Bosnjak, M.; Ristic, B.; Arsikin, K.; Mircic, A.; Suzin-Zivkovic, V.; Perovic, V.; Bogdanovic, A.; Paunovic, V.; Markovic, I.; Bumbasirevic, V.; et al. Inhibition of mTOR-dependent autophagy sensitizes leukemic cells to cytarabine-induced apoptotic death. PLoS ONE 2014, 9, e94374. [Google Scholar] [CrossRef]
- Piya, S.; Kornblau, S.M.; Ruvolo, V.R.; Mu, H.; Ruvolo, P.P.; McQueen, T.; Davis, R.E.; Hail, N., Jr.; Kantarjian, H.; Andreeff, M.; et al. Atg7 suppression enhances chemotherapeutic agent sensitivity and overcomes stroma-mediated chemoresistance in acute myeloid leukemia. Blood 2016, 128, 1260–1269. [Google Scholar] [CrossRef]
- Isakson, P.; Bjoras, M.; Boe, S.O.; Simonsen, A. Autophagy contributes to therapy-induced degradation of the PML/RARA oncoprotein. Blood 2010, 116, 2324–2331. [Google Scholar] [CrossRef]
- Orfali, N.; O’Donovan, T.R.; Nyhan, M.J.; Britschgi, A.; Tschan, M.P.; Cahill, M.R.; Mongan, N.P.; Gudas, L.J.; McKenna, S.L. Induction of autophagy is a key component of all-trans-retinoic acid-induced differentiation in leukemia cells and a potential target for pharmacologic modulation. Exp. Hematol. 2015, 43, 781–793e782. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, L.; Kang, R.; Yang, M.; Liu, L.; Zhao, Y.; Yu, Y.; Xie, M.; Yin, X.; Livesey, K.M.; et al. Autophagy regulates myeloid cell differentiation by p62/SQSTM1-mediated degradation of PML-RARalpha oncoprotein. Autophagy 2011, 7, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Goussetis, D.J.; Gounaris, E.; Wu, E.J.; Vakana, E.; Sharma, B.; Bogyo, M.; Altman, J.K.; Platanias, L.C. Autophagic degradation of the BCR-ABL oncoprotein and generation of antileukemic responses by arsenic trioxide. Blood 2012, 120, 3555–3562. [Google Scholar] [CrossRef] [PubMed]
- Stankov, M.V.; El Khatib, M.; Kumar Thakur, B.; Heitmann, K.; Panayotova-Dimitrova, D.; Schoening, J.; Bourquin, J.P.; Schweitzer, N.; Leverkus, M.; Welte, K.; et al. Histone deacetylase inhibitors induce apoptosis in myeloid leukemia by suppressing autophagy. Leukemia 2014, 28, 577–588. [Google Scholar] [CrossRef]
- Torgersen, M.L.; Engedal, N.; Boe, S.O.; Hokland, P.; Simonsen, A. Targeting autophagy potentiates the apoptotic effect of histone deacetylase inhibitors in t(8;21) AML cells. Blood 2013, 122, 2467–2476. [Google Scholar] [CrossRef]
- Li, L.; Liu, W.; Sun, Q.; Zhu, H.; Hong, M.; Qian, S. Decitabine Downregulates TIGAR to Induce Apoptosis and Autophagy in Myeloid Leukemia Cells. Oxid. Med. Cell Longev. 2021, 2021, 8877460. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Deng, J.; Fan, D.; Duan, Z.; Zhu, C.; Fu, R.; Wang, S. Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem. Pharmacol. 2018, 148, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Sachan, R.; Kundu, A.; Jeon, Y.; Choi, W.S.; Yoon, K.; Kim, I.S.; Kwak, J.H.; Kim, H.S. Afrocyclamin A, a triterpene saponin, induces apoptosis and autophagic cell death via the PI3K/Akt/mTOR pathway in human prostate cancer cells. Phytomedicine 2018, 51, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Chollat-Namy, M.; Ben Safta-Saadoun, T.; Haferssas, D.; Meurice, G.; Chouaib, S.; Thiery, J. The pharmalogical reactivation of p53 function improves breast tumor cell lysis by granzyme B and NK cells through induction of autophagy. Cell Death Dis. 2019, 10, 695. [Google Scholar] [CrossRef] [PubMed]
- Cutrignelli, A.; Sanarica, F.; Lopalco, A.; Lopedota, A.; Laquintana, V.; Franco, M.; Boccanegra, B.; Mantuano, P.; De Luca, A.; Denora, N. Dasatinib/HP-β-CD Inclusion Complex Based Aqueous Formulation as a Promising Tool for the Treatment of Paediatric Neuromuscular Disorders. Int. J. Mol. Sci. 2019, 20, 591. [Google Scholar] [CrossRef] [PubMed]
- Eid, E.E.M.; Alshehade, S.A.; Almaiman, A.A.; Kamran, S.; Lee, V.S.; Alshawsh, M.A. Enhancing the Anti-Leukemic Potential of Thymoquinone/Sulfobutylether-β-cyclodextrin (SBE-β-CD) Inclusion Complexes. Biomedicines 2023, 11, 1891. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Chean, C.S.; Ng, Y.Y.; Chan, S.Y.; Ho, P.C. 2-hydroxypropyl-beta-cyclodextrin increases aqueous solubility and photostability of all-trans-retinoic acid. J. Clin. Pharm. Ther. 2000, 25, 265–269. [Google Scholar] [CrossRef]
- Jin, S.; Cojocari, D.; Purkal, J.J.; Popovic, R.; Talaty, N.N.; Xiao, Y.; Solomon, L.R.; Boghaert, E.R.; Leverson, J.D.; Phillips, D.C. 5-Azacitidine Induces NOXA to Prime AML Cells for Venetoclax-Mediated Apoptosis. Clin. Cancer Res. 2020, 26, 3371–3383. [Google Scholar] [CrossRef]
- Ono, A.; Hattori, S.; Kariya, R.; Iwanaga, S.; Taura, M.; Harada, H.; Suzu, S.; Okada, S. Comparative study of human hematopoietic cell engraftment into BALB/c and C57BL/6 strain of rag-2/jak3 double-deficient mice. J. Biomed. Biotechnol. 2011, 2011, 539748. [Google Scholar] [CrossRef]
- Chow, K.U.; Ries, J.; Weidmann, E.; Pourebrahim, F.; Napieralski, S.; Stieler, M.; Boehrer, S.; Rummel, M.J.; Stein, J.; Hoelzer, D.; et al. Induction of apoptosis using 2′,2′ difluorodeoxycytidine (gemcitabine) in combination with antimetabolites or anthracyclines on malignant lymphatic and myeloid cells. Antagonism or synergism depends on incubation schedule and origin of neoplastic cells. Ann Hematol. 2000, 79, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, R.; Hattori, Y.; Hosogi, S.; Toda, Y.; Takata, K.; Ashihara, E. A novel dipeptide type inhibitor of the Wnt/β-catenin pathway suppresses proliferation of acute myelogenous leukemia cells. Biochem. Biophys. Res. Commun. 2021, 535, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (mM) | ||
|---|---|---|
| Cell Line | HP-β-CyD | FA-HP-β-CyD |
| HL-60 | 7.66 | 0.62 |
| THP-1 | 9.05 | 0.32 |
| SKM-1 | 9.44 | 0.2 |
| Kasumi-1 | 3.14 | 0.3 |
| HP-β-CyD + Ara-C | FA-HP-β-CyD + Ara-C | |
|---|---|---|
| HL-60 | 1.412 | 0.507 |
| THP1 | 1.085 | 0.469 |
| SKM1 | 1.299 | 0.706 |
| HP-β-CyD + Venetoclax | FA-HP-β-CyD + Venetoclax | |
|---|---|---|
| HL-60 | 2.013 | 1.056 |
| THP1 | 1.415 | 0.82 |
| SKM1 | 1.26 | 0.905 |
| HP-β-CyD (mg/kg) | FA-HP-β-CyD (mg/kg) | ||||||
|---|---|---|---|---|---|---|---|
| vehicle | 2000 | 6000 | 10,000 | 200 | 600 | 1000 | |
| WBC (×102/μL) | 57.0 ± 10.67 | 44.7 ± 12.65 | 50.0 ± 2.34 | 30.5 ± 1.65 * | 50.7 ± 13.36 | 52.5 ± 3.84 | 55.7 ± 14.28 |
| Hb (g/dL) | 15.1 ± 0.60 | 12.4 ± 0.67 | 14.3 ± 0.25 | 14.5 ± 0.78 | 13.8 ± 1.12 | 14.5 ± 0.27 | 14.7 ± 0.28 |
| Plt (×104/μL) | 59.7 ± 11.0 | 64.5 ± 2.29 | 60.2 ± 10.69 | 75.5 ± 17.83 | 55.8 ± 15.67 | 61.1 ± 12.50 | 55.7 ± 18.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubota, Y.; Hoshiko, T.; Higashi, T.; Motoyama, K.; Okada, S.; Kimura, S. Folate-Appended Hydroxypropyl-β-Cyclodextrin Induces Autophagic Cell Death in Acute Myeloid Leukemia Cells. Int. J. Mol. Sci. 2023, 24, 16720. https://doi.org/10.3390/ijms242316720
Kubota Y, Hoshiko T, Higashi T, Motoyama K, Okada S, Kimura S. Folate-Appended Hydroxypropyl-β-Cyclodextrin Induces Autophagic Cell Death in Acute Myeloid Leukemia Cells. International Journal of Molecular Sciences. 2023; 24(23):16720. https://doi.org/10.3390/ijms242316720
Chicago/Turabian StyleKubota, Yasushi, Toshimi Hoshiko, Taishi Higashi, Keiichi Motoyama, Seiji Okada, and Shinya Kimura. 2023. "Folate-Appended Hydroxypropyl-β-Cyclodextrin Induces Autophagic Cell Death in Acute Myeloid Leukemia Cells" International Journal of Molecular Sciences 24, no. 23: 16720. https://doi.org/10.3390/ijms242316720
APA StyleKubota, Y., Hoshiko, T., Higashi, T., Motoyama, K., Okada, S., & Kimura, S. (2023). Folate-Appended Hydroxypropyl-β-Cyclodextrin Induces Autophagic Cell Death in Acute Myeloid Leukemia Cells. International Journal of Molecular Sciences, 24(23), 16720. https://doi.org/10.3390/ijms242316720