
Allergenic Activity of Individual Cat Allergen Molecules

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
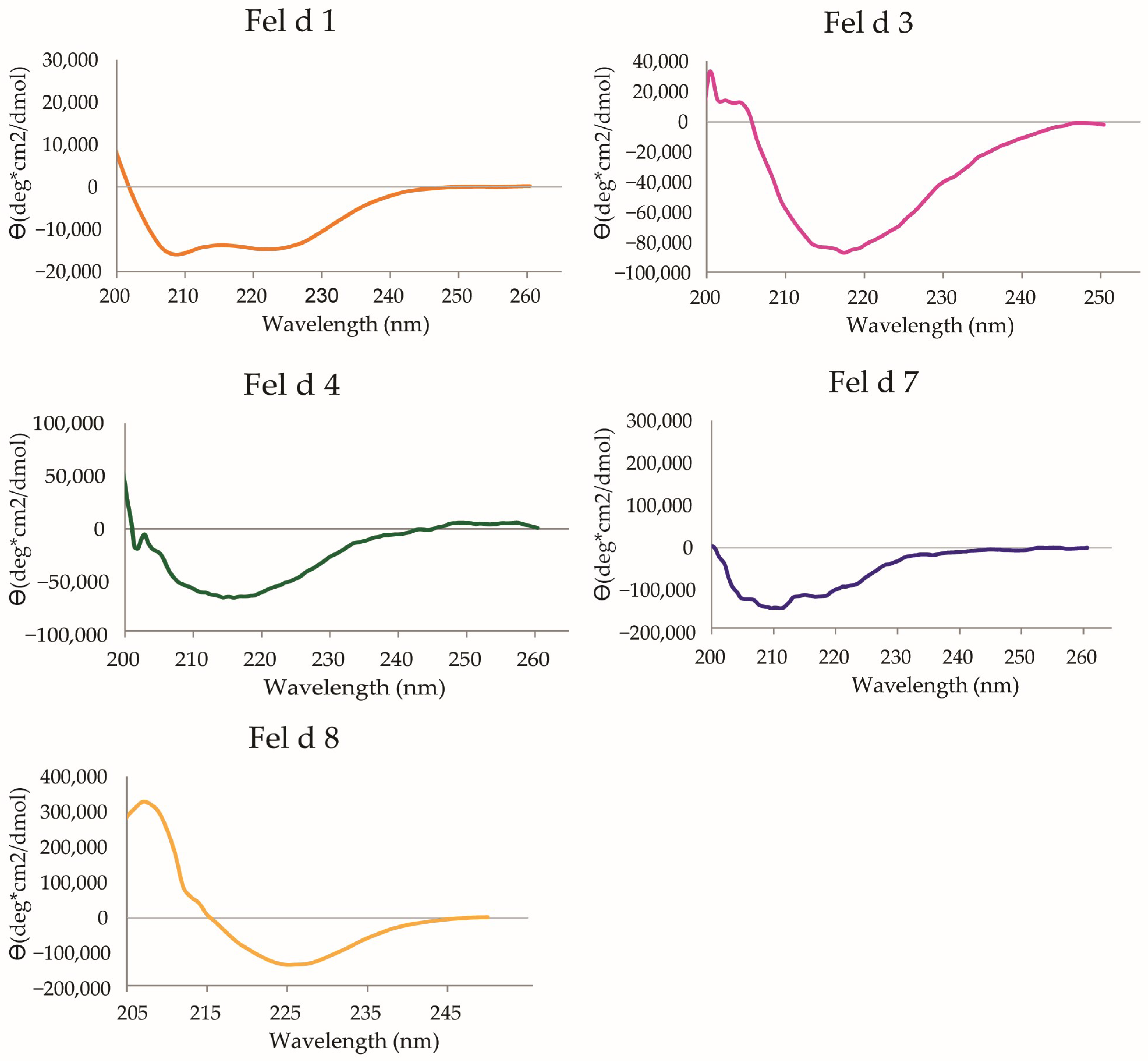
2.1. Characterization of Cat Allergen Molecules
2.2. Characterization of Subjects with IgE Sensitization to Cat Exposure
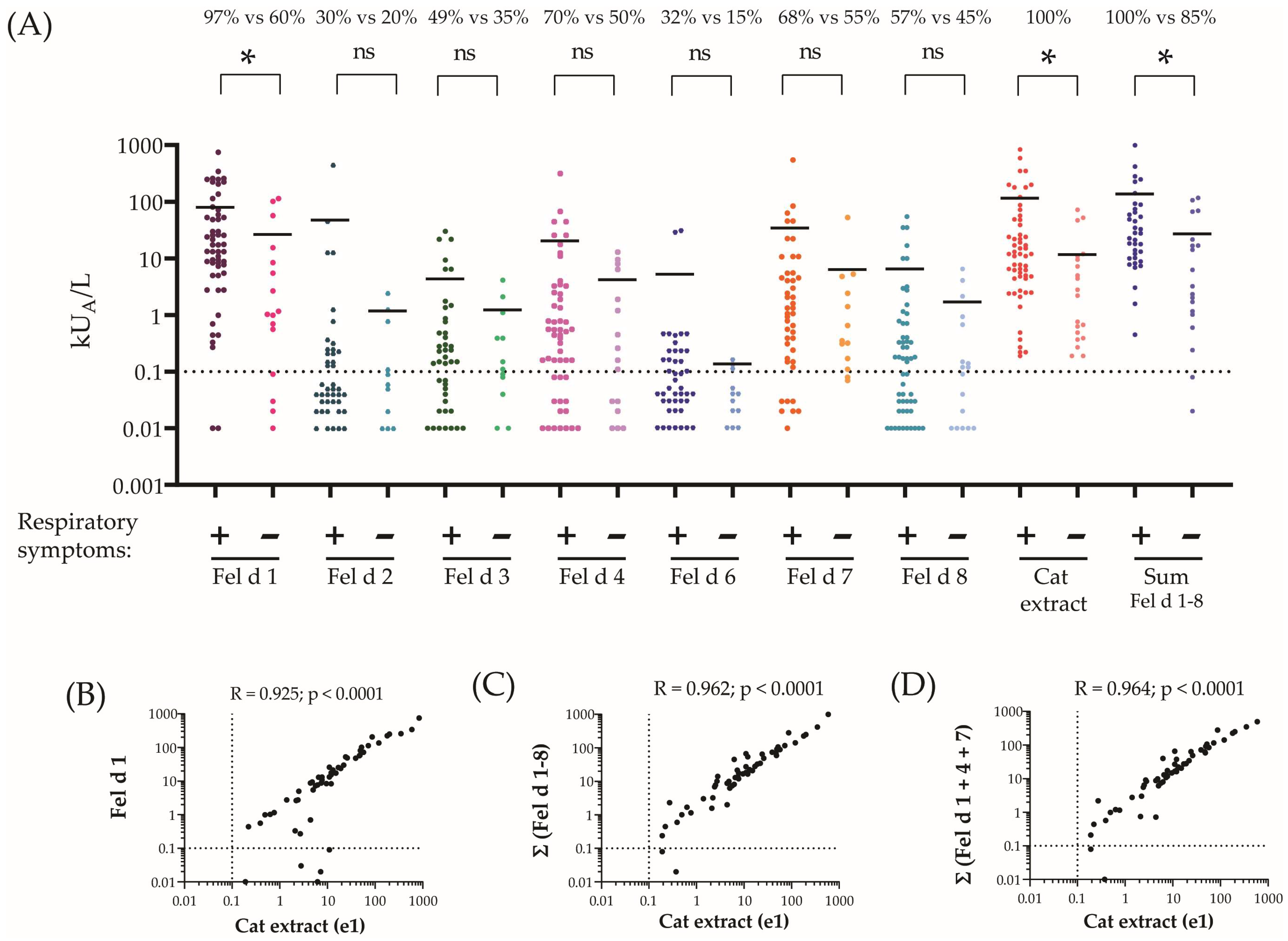
2.3. Fel d 1, Fel d 4 and Fel d 7 Are the Most Frequently Recognized Cat Allergens and Account for the Majority of Cat-Allergen-Specific IgE
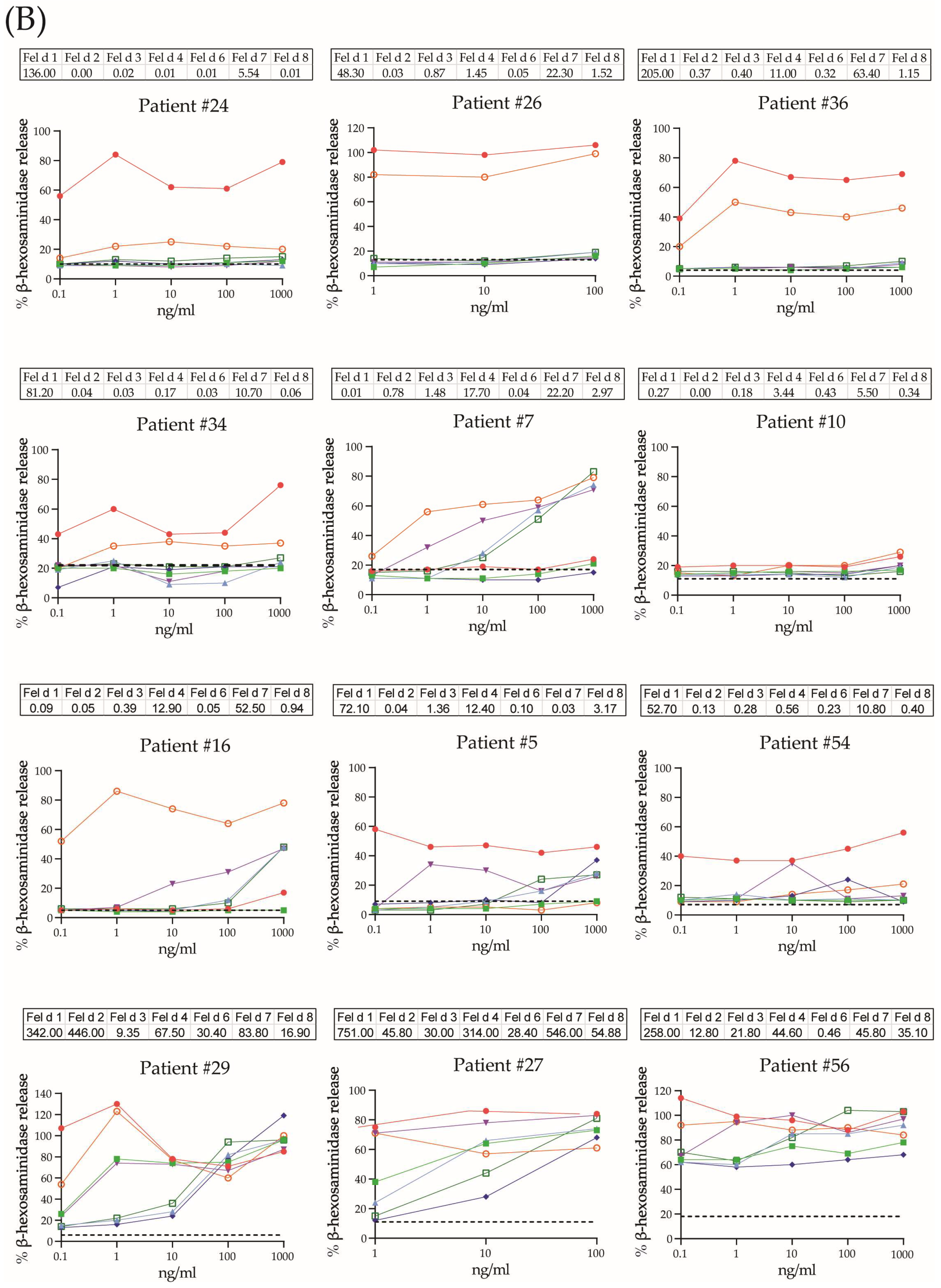
2.4. Allergenic Activity of Cat Allergens
3. Discussion
4. Materials and Methods
4.1. Cat-Allergic Patients’ Sera
4.2. Allergen Molecules
4.3. Allergen-Specific IgE Levels Quantified by ImmunoCAP
4.4. Rat Basophil Leukemia (RBL) Cells Assay for Testing Allergenic Activity
4.5. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AIT | Allergen-specific immunotherapy |
| Fel d 1–8 | nomenclature of cat (Felis domesticus) allergens |
| IgE, IgM, IgG and IgG4 | Immunoglobulin E, immunoglobulin M, immunoglobulin G and immunoglobulin G4 subclass, respectively |
| IUIS | International Union of Immunological Societies’ database for allergen nomenclature |
| CD | Circular dichroism |
| PDB | Protein data bank |
| BPI | Bactericidal/permeability-increasing protein |
| kUA/L | IgE levels in ImmunoCAP standardized units |
| RBL | Rat basophil leukemia cells |
References
- Zahradnik, E.; Raulf, M. Respiratory Allergens from Furred Mammals: Environmental and Occupational Exposure. Vet. Sci. 2017, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Heinzerling, L.M.; Burbach, G.J.; Edenharter, G.; Bachert, C.; Bindslev-Jensen, C.; Bonini, S.; Bousquet, J.; Bousquet-Rouanet, L.; Bousquet, P.J.; Bresciani, M.; et al. GA2LEN skin test study I: GA2LEN harmonization of skin prick testing: Novel sensitization patterns for inhalant allergens in Europe. Allergy 2009, 64, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Lupinek, C.; Vrtala, S.; Wieser, S.; Baar, A.; Kiss, R.; Kull, I.; Melén, E.; Wickman, M.; Porta, D.; et al. A molecular sensitization map of European children reveals exposome- and climate-dependent sensitization profiles. Allergy 2023, 78, 2007–2018. [Google Scholar] [CrossRef]
- D’souza, N.; Weber, M.; Sarzsinszky, E.; Vrtala, S.; Curin, M.; Schaar, M.; Garib, V.; Focke-Tejkl, M.; Li, Y.; Jones, R.; et al. The Molecular Allergen Recognition Profile in China as Basis for Allergen-Specific Immunotherapy. Front. Immunol. 2021, 12, 719573. [Google Scholar] [CrossRef] [PubMed]
- González-Pérez, R.; Poza-Guedes, P.; Pineda, F.; Galán, T.; Mederos-Luis, E.; Abel-Fernández, E.; Martínez, M.J.; Sánchez-Machín, I. Molecular Mapping of Allergen Exposome among Different Atopic Phenotypes. Int. J. Mol. Sci. 2023, 24, 10467. [Google Scholar] [CrossRef]
- van Hage, M.; Käck, U.; Asarnoj, A.; Konradsen, J.R. An update on the prevalence and diagnosis of cat and dog allergy—Emphasizing the role of molecular allergy diagnostics. Mol. Immunol. 2023, 157, 1–7. [Google Scholar] [CrossRef]
- Landzaat, L.J.; Emons, J.A.M.; Sonneveld, L.J.H.; Schreurs, M.W.J.; Arends, N.J.T. Early inhalant allergen sensitization at component level: An analysis in atopic Dutch children. Front. Allergy 2023, 4, 1173540. [Google Scholar] [CrossRef]
- Asarnoj, A.; Hamsten, C.; Wadén, K.; Lupinek, C.; Andersson, N.; Kull, I.; Curin, M.; Anto, J.; Bousquet, J.; Valenta, R.; et al. Sensitization to cat and dog allergen molecules in childhood and prediction of symptoms of cat and dog allergy in adolescence: A BAMSE/MeDALL study. J. Allergy Clin. Immunol. 2016, 137, 813–821.e7. [Google Scholar] [CrossRef]
- Elisyutina, O.; Lupinek, C.; Fedenko, E.; Litovkina, A.; Smolnikov, E.; Ilina, N.; Kudlay, D.; Shilovskiy, I.; Valenta, R.; Khaitov, M. IgE-reactivity profiles to allergen molecules in Russian children with and without symptoms of allergy revealed by micro-array analysis. Pediatr. Allergy Immunol. 2021, 32, 251–263. [Google Scholar] [CrossRef]
- Hemmer, W.; Sestak-Greinecker, G.; Braunsteiner, T.; Wantke, F.; Wöhrl, S. Molecular sensitization patterns in animal allergy: Relationship with clinical relevance and pet ownership. Allergy 2021, 76, 3687–3696. [Google Scholar] [CrossRef]
- Nwaru, B.I.; Suzuki, S.; Ekerljung, L.; Sjölander, S.; Mincheva, R.; Rönmark, E.P.; Rådinger, M.; Rönmark, E.; Borres, M.P.; Lundbäck, B.; et al. Furry Animal Allergen Component Sensitization and Clinical Outcomes in Adult Asthma and Rhinitis. J. Allergy Clin. Immunol. Pract. 2019, 7, 1230–1238.e4. [Google Scholar] [CrossRef]
- Wickman, M.; Lupinek, C.; Andersson, N.; Belgrave, D.; Asarnoj, A.; Benet, M.; Pinart, M.; Wieser, S.; Garcia-Aymerich, J.; Baar, A.; et al. Detection of IgE Reactivity to a Handful of Allergen Molecules in Early Childhood Predicts Respiratory Allergy in Adolescence. EBioMedicine 2017, 26, 91–99. [Google Scholar] [CrossRef]
- Konradsen, J.R.; Nordlund, B.; Onell, A.; Borres, M.P.; Grönlund, H.; Hedlin, G. Severe childhood asthma and allergy to furry animals: Refined assessment using molecular-based allergy diagnostics. Pediatr. Allergy Immunol. 2014, 25, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.A.; Agrawal, R.; Minnicozzi, S.; Xin, W.; Patrie, J.; Heymann, P.W.; Workman, L.; Platts-Mills, T.A.; Song, T.W.; Moloney, M.; et al. Sensitization to food and inhalant allergens in relation to age and wheeze among children with atopic dermatitis. Clin. Exp. Allergy 2013, 43, 1160–1170. [Google Scholar] [CrossRef]
- Grönlund, H.; Adédoyin, J.; Reininger, R.; Varga, E.-M.; Zach, M.; Fredriksson, M.; Kronqvist, M.; Szepfalusi, Z.; Spitzauer, S.; Grönneberg, R.; et al. Higher immunoglobulin E antibody levels to recombinant Fel d 1 in cat-allergic children with asthma compared with rhinoconjunctivitis. Clin. Exp. Allergy 2008, 38, 1275–1281. [Google Scholar] [CrossRef]
- Nordlund, B.; Konradsen, J.R.; Kull, I.; Borres, M.P.; Önell, A.; Hedlin, G.; Grönlund, H. IgE antibodies to animal-derived lipocalin, kallikrein and secretoglobin are markers of bronchial inflammation in severe childhood asthma. Allergy 2012, 67, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.R. The advent of recombinant allergens and allergen cloning. J. Allergy Clin. Immunol. 2011, 127, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Breda, D.; Spisni, A.; Burastero, S.E. Component-Resolved Diagnosis Based on a Recombinant Variant of Mus m 1 Lipocalin Allergen. Int. J. Mol. Sci. 2023, 24, 1193. [Google Scholar] [CrossRef]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatr. Allergy Immunol. 2016, 27, 1–250. [Google Scholar] [CrossRef]
- WHO/IUIS Allergen Nomenclature. Available online: http://www.allergen.org/search.php?Species=Felis%20domesticus%20(F.%20catus) (accessed on 2 November 2023).
- Morgenstern, J.P.; Griffith, I.J.; Brauer, A.W.; Rogers, B.L.; Bond, J.F.; Chapman, M.D.; Kuo, M.C. Amino acid sequence of Fel dI, the major allergen of the domestic cat: Protein sequence analysis and cDNA cloning. Proc. Natl. Acad. Sci. USA 1991, 88, 9690–9694. [Google Scholar] [CrossRef]
- Kaiser, L.; Grönlund, H.; Sandalova, T.; Ljunggren, H.-G.; van Hage-Hamsten, M.; Achour, A.; Schneider, G. The crystal structure of the major cat allergen Fel d 1, a member of the secretoglobin family. J. Biol. Chem. 2003, 278, 37730–37735. [Google Scholar] [CrossRef] [PubMed]
- Norman, P.S.; Ohman, J.L., Jr.; Long, A.A.; Creticos, P.S.; Gefter, M.A.; Shaked, Z.E.; Wood, R.A.; Eggleston, P.A.; Hafner, K.B.; Rao, P. Treatment of cat allergy with T-cell reactive peptides. Am. J. Respir. Crit. Care Med. 1996, 154 Pt 1, 1623–1628. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, W.; Larché, M.; Kay, A. Effect of T-cell peptides derived from Fel d 1 on allergic reactions and cytokine production in patients sensitive to cats: A randomised controlled trial. Lancet 2002, 360, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, N.; Dietmeier, K.; Bauer, M.; Maudrich, M.; Utzinger, S.; Muntwiler, S.; Saudan, P.; Bachmann, M.F. Displaying Fel d1 on virus-like particles prevents reactogenicity despite greatly enhanced immunogenicity: A novel therapy for cat allergy. J. Exp. Med. 2009, 206, 1941–1955. [Google Scholar] [CrossRef] [PubMed]
- Orengo, J.M.; Radin, A.R.; Kamat, V.; Badithe, A.; Ben, L.H.; Bennett, B.L.; Zhong, S.; Birchard, D.; Limnander, A.; Rafique, A.; et al. Treating cat allergy with monoclonal IgG antibodies that bind allergen and prevent IgE engagement. Nat. Commun. 2018, 9, 1421. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, B.; Messaoudi, K.; Jacomet, F.; Michaud, E.; Fauquert, J.L.; Caillaud, D.; Evrard, B. An update on molecular cat allergens: Fel d 1 and what else? Chapter 1: Fel d 1, the major cat allergen. Allergy Asthma Clin. Immunol. 2018, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Dávila, I.; Domínguez-Ortega, J.; Navarro-Pulido, A.; Alonso, A.; Antolín-Amerigo, D.; González-Mancebo, E.; Martín-García, C.; Núñez-Acevedo, B.; Prior, N.; Reche, M.; et al. Consensus document on dog and cat allergy. Allergy 2018, 73, 1206–1222. [Google Scholar] [CrossRef] [PubMed]
- Hales, B.J.; Chai, L.Y.; Hazell, L.; Elliot, C.E.; Stone, S.; O’Neil, S.E.; Smith, W.-A.; Thomas, W.R. IgE and IgG Binding Patterns and T-cell Recognition of Fel d 1 and Non–Fel d 1 Cat Allergens. J. Allergy Clin. Immunol. Pract. 2013, 1, 656–665.e5. [Google Scholar] [CrossRef]
- Riabova, K.; Karsonova, A.V.; van Hage, M.; Käck, U.; Konradsen, J.R.; Grönlund, H.; Fomina, D.; Beltyukov, E.; Glazkova, P.A.; Semenov, D.Y.; et al. Molecular Allergen-Specific IgE Recognition Profiles and Cumulative Specific IgE Levels Associated with Phenotypes of Cat Allergy. Int. J. Mol. Sci. 2022, 23, 6984. [Google Scholar] [CrossRef]
- Popescu, F.-D.; Ganea, C.S.; Panaitescu, C.; Vieru, M. Molecular diagnosis in cat allergy. World J. Methodol. 2021, 11, 46–60. [Google Scholar] [CrossRef]
- Caraballo, L.; Valenta, R.; Puerta, L.; Pomés, A.; Zakzuk, J.; Fernandez-Caldas, E.; Acevedo, N.; Sanchez-Borges, M.; Ansotegui, I.; Zhang, L.; et al. The allergenic activity and clinical impact of individual IgE-antibody binding molecules from indoor allergen sources. World Allergy Organ. J. 2020, 13, 100118. [Google Scholar] [CrossRef]
- Westritschnig, K.; Horak, F.; Swoboda, I.; Balic, N.; Spitzauer, S.; Kundi, M.; Fiebig, H.; Suck, R.; Cromwell, O.; Valenta, R. Different allergenic activity of grass pollen allergens revealed by skin testing. Eur. J. Clin. Investig. 2008, 38, 260–267. [Google Scholar] [CrossRef]
- Jutel, M.; Jaeger, L.; Suck, R.; Meyer, H.; Fiebig, H.; Cromwell, O. Allergen-specific immunotherapy with recombinant grass pollen allergens. J. Allergy Clin. Immunol. 2005, 116, 608–613. [Google Scholar] [CrossRef]
- Niederberger, V.; Neubauer, A.; Gevaert, P.; Zidarn, M.; Worm, M.; Aberer, W.; Malling, H.J.; Pfaar, O.; Klimek, L.; Pfützner, W.; et al. Safety and efficacy of immunotherapy with the recombinant B-cell epitope–based grass pollen vaccine BM32. J. Allergy Clin. Immunol. 2018, 142, 497–509.e9. [Google Scholar] [CrossRef] [PubMed]
- Niederberger, V.; Eckl-Dorna, J.; Pauli, G. Recombinant allergen-based provocation testing. Methods 2014, 66, 96–105. [Google Scholar] [CrossRef]
- Blackley, C.H. Experimental Researches on the Causes and Nature of Catarrhus aestivus (Hay Fever or Hay-Asthma); Balliere Tindall & Cox: London, UK, 1873. [Google Scholar]
- Pfaar, O.; Bergmann, K.; Bonini, S.; Compalati, E.; Domis, N.; de Blay, F.; de Kam, P.; Devillier, P.; Durham, S.R.; Ellis, A.K.; et al. Technical standards in allergen exposure chambers worldwide—An EAACI Task Force Report. Allergy 2021, 76, 3589–3612. [Google Scholar] [CrossRef]
- Santos, A.F.; Du Toit, G.; O’rourke, C.; Becares, N.; Couto-Francisco, N.; Radulovic, S.; Khaleva, E.; Basting, M.; Harris, K.M.; Larson, D.; et al. Biomarkers of severity and threshold of allergic reactions during oral peanut challenges. J. Allergy Clin. Immunol. 2020, 146, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Cañas, J.A.; Pérez-Sánchez, N.; Lopera-Doblas, L.; Palomares, F.; Molina, A.; Bartra, J.; Torres, M.J.; Gómez, F.; Mayorga, C. Basophil Activation Test Utility as a Diagnostic Tool in LTP Allergy. Int. J. Mol. Sci. 2022, 23, 4979. [Google Scholar] [CrossRef] [PubMed]
- Mondol, E.; Donado, K.; Regino, R.; Hernandez, K.; Mercado, D.; Mercado, A.C.; Benedetti, I.; Puerta, L.; Zakzuk, J.; Caraballo, L. The Allergenic Activity of Blo t 2, a Blomia tropicalis IgE-Binding Molecule. Int. J. Mol. Sci. 2023, 24, 5543. [Google Scholar] [CrossRef]
- Hoffmann, A.; Jamin, A.; Foetisch, K.; May, S.; Aulepp, H.; Haustein, D.; Vieths, S. Determination of the allergenic activity of birch pollen and apple prick test solutions by measurement of β-hexosaminidase release from RBL-2H3 cells. Comparison with classical methods in allergen standardization. Allergy 1999, 54, 446–454. [Google Scholar] [CrossRef]
- Adédoyin, J.; Johansson, S.; Grönlund, H.; van Hage, M. Interference in immunoassays by human IgM with specificity for the carbohydrate moiety of animal proteins. J. Immunol. Methods 2006, 310, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Schooltink, L.; Sagmeister, T.; Gottstein, N.; Pavkov-Keller, T.; Keller, W. Crystal Structure of cat allergen Fel d 4. In Full wwPDB X-ray Structure Validation Report; PDB8AMC; wwPDB: Piscataway, NJ, USA, 2023. [Google Scholar] [CrossRef]
- Min, J.; Pedersen, L.C.; Geoffrey, M.A. 2.2 A crystal structure of the lipocalin cat allergen Fel d 7. In Full wwPDB X-ray Structure Validation Report; PDB8AMC; wwPDB: Piscataway, NJ, USA, 2023. [Google Scholar] [CrossRef]
- Smith, W.; O’neil, S.; Hales, B.; Chai, T.; Hazell, L.; Tanyaratsrisakul, S.; Piboonpocanum, S.; Thomas, W. Two newly identified cat allergens: The von Ebner gland protein Fel d 7 and the latherin-like protein Fel d 8. Int. Arch. Allergy Immunol. 2011, 156, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Botros, H.G.; Poncet, P.; Rabillon, J.; Fontaine, T.; Laval, J.; David, B. Biochemical characterization and surfactant properties of horse allergens. JBIC J. Biol. Inorg. Chem. 2001, 268, 3126–3136. [Google Scholar] [CrossRef]
- Li, J.; Xu, P.; Wang, L.; Feng, M.; Chen, D.; Yu, X.; Lu, Y. Molecular biology of BPIFB1 and its advances in disease. Ann. Transl. Med. 2020, 8, 651. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ishibashi, O.; Sugiura, K.; Ubatani, M.; Sakaguchi, M.; Nakatsuji, M.; Shimamoto, S.; Noda, M.; Uchiyama, S.; Fukutomi, Y.; et al. Crystal structure of the dog allergen Can f 6 and structure-based implications of its cross-reactivity with the cat allergen Fel d 4. Sci. Rep. 2019, 9, 1503. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.B.; Binnmyr, J.; Zoltowska, A.; Saarne, T.; van Hage, M.; Grönlund, H. Characterization of the dog lipocalin allergen Can f 6: The role in cross-reactivity with cat and horse. Allergy 2012, 67, 751–757. [Google Scholar] [CrossRef]
- Lichtenstein, L.M.; Osler, A.G. Studies on the mechanisms of hypersensitivity phenomena. IX. Histamine release from human leukocytes by ragweed pollen antigen. J. Exp. Med. 1964, 120, 507–530. [Google Scholar] [CrossRef]
- Lichtenstein, L.M.; King, T.P.; Osler, A.G. In vitro assay of allergenic properties of ragweed pollen antigens. J. Allergy 1966, 38, 174–182. [Google Scholar] [CrossRef]
- Blanc, F.; Adel-Patient, K.; Drumare, M.; Paty, E.; Wal, J.; Bernard, H. Capacity of purified peanut allergens to induce degranulation in a functional in vitro assay: Ara h 2 and Ara h 6 are the most efficient elicitors. Clin. Exp. Allergy 2009, 39, 1277–1285. [Google Scholar] [CrossRef]
- Kaul, S.; Lüttkopf, D.; Kastner, B.; Vogel, L.; Höltz, G.; Vieths, S.; Hoffmann, A. Mediator release assays based on human or murine immunoglobulin E in allergen standardization. Clin. Exp. Allergy 2006, 37, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Vogel, L.; Lüttkopf, D.; Hatahet, L.; Haustein, D.; Vieths, S. Development of a functional in vitro assay as a novel tool for the standardization of allergen extracts in the human system. Allergy 2005, 60, 1021–1028. [Google Scholar] [CrossRef]
- Miller, L.; Blank, U.; Metzger, H.; Kinet, J.-P. Expression of high-affinity binding of human immunoglobulin E by transfected cells. Science 1989, 244, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Falcone, F.H.; Alcocer, M.J.C.; Okamoto-Uchida, Y.; Nakamura, R. Use of humanized rat basophilic leukemia reporter cell lines as a diagnostic tool for detection of allergen-specific IgE in allergic patients: Time for a reappraisal? Curr. Allergy Asthma Rep. 2015, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Falcone, F.H.; Wan, D.; Barwary, N.; Sagi-Eisenberg, R. RBL cells as models for in vitro studies of mast cells and basophils. Immunol. Rev. 2018, 282, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Dibbern, D.A., Jr.; Palmer, G.; Williams, P.; Bock, S.; Dreskin, S.C. RBL cells expressing human FcεRI are a sensitive tool for exploring functional IgE–allergen interactions: Studies with sera from peanut-sensitive patients. J. Immunol. Methods 2003, 274, 37–45. [Google Scholar] [CrossRef]
- Kleine-Tebbe, J.; Erdmann, S.; Knol, E.F.; MacGlashan, D.W., Jr.; Poulsen, L.K.; Gibbs, B.F. Diagnostic tests based on human basophils: Potentials, pitfalls and perspectives. Int. Arch. Allergy Immunol. 2006, 141, 79–90. [Google Scholar] [CrossRef]
- Grönlund, H.; Bergman, T.; Sandström, K.; Alvelius, G.; Reininger, R.; Verdino, P.; Hauswirth, A.; Liderot, K.; Valent, P.; Spitzauer, S.; et al. Formation of disulfide bonds and homodimers of the major cat allergen Fel d 1 equivalent to the natural Allergen by expression in Escherichia coli. J. Biol. Chem. 2003, 278, 40144–40151. [Google Scholar] [CrossRef]
- Gabet, S.; Just, J.; Couderc, R.; Seta, N.; Momas, I. Allergic sensitisation in early childhood: Patterns and related factors in PARIS birth cohort. Int. J. Hyg. Environ. Health 2016, 219, 792–800. [Google Scholar] [CrossRef]
- Bertelsen, R.J.; Instanes, C.; Granum, B.; Carlsen, K.C.L.; Hetland, G.; Mowinckel, P.; Løvik, M. Gender differences in indoor allergen exposure and association with current rhinitis. Clin. Exp. Allergy 2010, 40, 1388–1397. [Google Scholar] [CrossRef]
- Konradsen, J.R.; Nordlund, B.; Nilsson, O.B.; van Hage, M.; Nopp, A.; Hedlin, G.; Grönlund, H. High basophil allergen sensitivity (CD-sens) is associated with severe allergic asthma in children. Pediatr. Allergy Immunol. 2012, 23, 376–384. [Google Scholar] [CrossRef]
- Hoffmann, H.J.; Santos, A.F.; Mayorga, C.; Nopp, A.; Eberlein, B.; Ferrer, M.; Rouzaire, P.; Ebo, D.G.; Sabato, V.; Sanz, M.L.; et al. The clinical utility of basophil activation testing in diagnosis and monitoring of allergic disease. Allergy 2015, 70, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Gieras, A.; Linhart, B.; Roux, K.H.; Dutta, M.; Khodoun, M.; Zafred, D.; Cabauatan, C.R.; Lupinek, C.; Weber, M.; Focke-Tejkl, M.; et al. IgE epitope proximity determines immune complex shape and effector cell activation capacity. J. Allergy Clin. Immunol. 2016, 137, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Hjort, C.; Schiøtz, P.O.; Ohlin, M.; Würtzen, P.A.; Christensen, L.H.; Hoffmann, H.J. The number and affinity of productive IgE pairs determine allergen activation of mast cells. J. Allergy Clin. Immunol. 2017, 140, 1167–1170.e2. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-J.; Resch-Marat, Y.; Casset, A.; Weghofer, M.; Zieglmayer, P.; Zieglmayer, R.; Lemell, P.; Horak, F.; Chen, K.-W.; Potapova, E.; et al. IgE recognition of the house dust mite allergen Der p 37 is associated with asthma. J. Allergy Clin. Immunol. 2022, 149, 1031–1043. [Google Scholar] [CrossRef]
- Erwin, E.; Custis, N.; Satinover, S.; Perzanowski, M.; Woodfolk, J.; Crane, J.; Wickens, K.; Plattsmills, T. Quantitative measurement of IgE antibodies to purified allergens using streptavidin linked to a high-capacity solid phase. J. Allergy Clin. Immunol. 2005, 115, 1029–1035. [Google Scholar] [CrossRef]
- Van Hage, M.; Hamsten, C.; Valenta, R. ImmunoCAP assays: Pros and cons in allergology. J. Allergy Clin. Immunol. 2017, 140, 974–977. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographics | All Patients | Respiratory+ | Respiratory− | Difference + vs. − |
|---|---|---|---|---|
| Number of patients | 57 | 37 | 20 | |
| Age, mean (min–max) | 13.1 (10–17) | 13.2 (10–17) | 12.9 (10–17) | n.s. |
| Female sex, N (%) | 21 (37%) | 13 (35%) | 8 (40%) | n.a. |
| Cat-related clinical symptoms | ||||
| Reported asthma, N (%) | 17 (29.8%) | 17 (45.9%) | n.a. | n.a. |
| Reported rhinitis, N (%) | 35 (61.4%) | 35 (94.6%) | n.a. | n.a. |
| Reported respiratory symptoms | 37 (64.9%) | 37 (100%) | n.a. | n.a. |
| From them, only asthma, N (%) | 3 (8%) | 3 (8.1%) | n.a. | n.a. |
| From them, only rhinitis, N (%) | 21 (36.8%) | 21 (56.7%) | n.a. | n.a. |
| From them, asthma with rhinitis, N (%) | 14 (24.56%) | 14 (37.8%) | n.a. | n.a. |
| Cat allergy | ||||
| IgE to cat dander (e1), mean kUA/L (min–max) | 53.2 (0.19–840) | 75.9 (0.2–840) | 11.5 (0.19–72) | <0.05 |
| N (%) | 57 (100%) | 37 (100%) | 20 (100%) | |
| IgE to Fel d 1, mean kUA/L (min–max) | 64.8 (0–751) | 77.8 (0–751) | 25.7 (0–113.5) | <0.05 |
| N (%) | 48 (84%) | 36 (97%) | 12 (60%) | |
| IgE to Fel d 2, mean kUA/L (min–max) | 31.9 (0–446) | 46.1 (0–446) | 1.15 (0–2.45) | n.s. |
| N (%) | 15 (26%) | 11 (30%) | 4 (20%) | |
| IgE to Fel d 3, mean kUA/L (min–max) | 3.4 (0–30) | 4.2 (0–30) | 1.2 (0–4.2) | n.s. |
| N (%) | 25 (44%) | 18 (49%) | 7 (33%) | |
| IgE to Fel d 4, mean kUA/L (min–max) | 15.4 (0–314) | 19.8 (0–314) | 4.1 (0–12.9) | n.s. |
| N (%) | 36 (63%) | 26 (70%) | 10 (50%) | |
| IgE to Fel d 6, mean kUA/L (min–max) | 4.1 (0–30.4) | 5.12 (0–30.4) | 0.13 (0–0.16) | n.s. |
| N (%) | 15 (26%) | 12 (32%) | 3 (15%) | |
| IgE to Fel d 7, mean kUA/L (min–max) | 25.15 (0–546) | 33.5 (0–546) | 6.2 (0–52.5) | n.s. |
| N (%) | 36 (63%) | 25 (68%) | 11 (55%) | |
| IgE to Fel d 8, mean kUA/L (min–max) | 4.9 (0–54.9) | 6.3 (0–54.9) | 1.65 (0–6.5) | n.s. |
| N (%) | 30 (53%) | 21 (57%) | 9 (45%) | |
| IgE to sum Fel d 1–8, mean kUA/L (min–max) | 99.6 (0–1770) | 133.4 (0.5–1770) | 26.3 (0–116.7) | <0.05 |
| N (%) | 54 (95%) | 37 (100%) | 17 (85%) | |
| Other allergy | ||||
| Other pet, N (%) | 51 (89.5%) | 34 (91.9%) | 17 (85%) | |
| Pollen, N (%) | 44 (77.3%) | 31 (83.8%) | 13 (65%) | |
| Food (class 1 and class 2 allergens), N (%) | 33 (57.9%) | 21 (56.8%) | 12 (60%) |
| Allergens | Patients (N = 17) | Plateau of Mediator Release Reached | |
|---|---|---|---|
| Positive, N/% | Range, ng/mL | Median Conc., ng/mL | |
| Fel d 1 | 15/88% | ≤0.1–1 | 0.1 |
| Fel d 2 | 4/23.5% | ≤0.1–10 | 1 |
| Fel d 3 | 6/35.3% | 10–≥1000 | 100 |
| Fel d 4 | 9/59.2% | 0.1–100 | 1 |
| Fel d 6 | 4/23.5% | ≥100 | ≥100 |
| Fel d 7 | 11/64.7% | ≤1 | 1 |
| Fel d 8 | 7/41.2% | ≥100 | ≥100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trifonova, D.; Curin, M.; Riabova, K.; Karsonova, A.; Keller, W.; Grönlund, H.; Käck, U.; Konradsen, J.R.; van Hage, M.; Karaulov, A.; et al. Allergenic Activity of Individual Cat Allergen Molecules. Int. J. Mol. Sci. 2023, 24, 16729. https://doi.org/10.3390/ijms242316729
Trifonova D, Curin M, Riabova K, Karsonova A, Keller W, Grönlund H, Käck U, Konradsen JR, van Hage M, Karaulov A, et al. Allergenic Activity of Individual Cat Allergen Molecules. International Journal of Molecular Sciences. 2023; 24(23):16729. https://doi.org/10.3390/ijms242316729
Chicago/Turabian StyleTrifonova, Daria, Mirela Curin, Ksenja Riabova, Antonina Karsonova, Walter Keller, Hans Grönlund, Ulrika Käck, Jon R. Konradsen, Marianne van Hage, Alexander Karaulov, and et al. 2023. "Allergenic Activity of Individual Cat Allergen Molecules" International Journal of Molecular Sciences 24, no. 23: 16729. https://doi.org/10.3390/ijms242316729
APA StyleTrifonova, D., Curin, M., Riabova, K., Karsonova, A., Keller, W., Grönlund, H., Käck, U., Konradsen, J. R., van Hage, M., Karaulov, A., & Valenta, R. (2023). Allergenic Activity of Individual Cat Allergen Molecules. International Journal of Molecular Sciences, 24(23), 16729. https://doi.org/10.3390/ijms242316729