
Sphingomyelin Metabolism Modifies Luminal A Breast Cancer Cell Line under a High Dose of Vitamin C

 , , ,
, , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
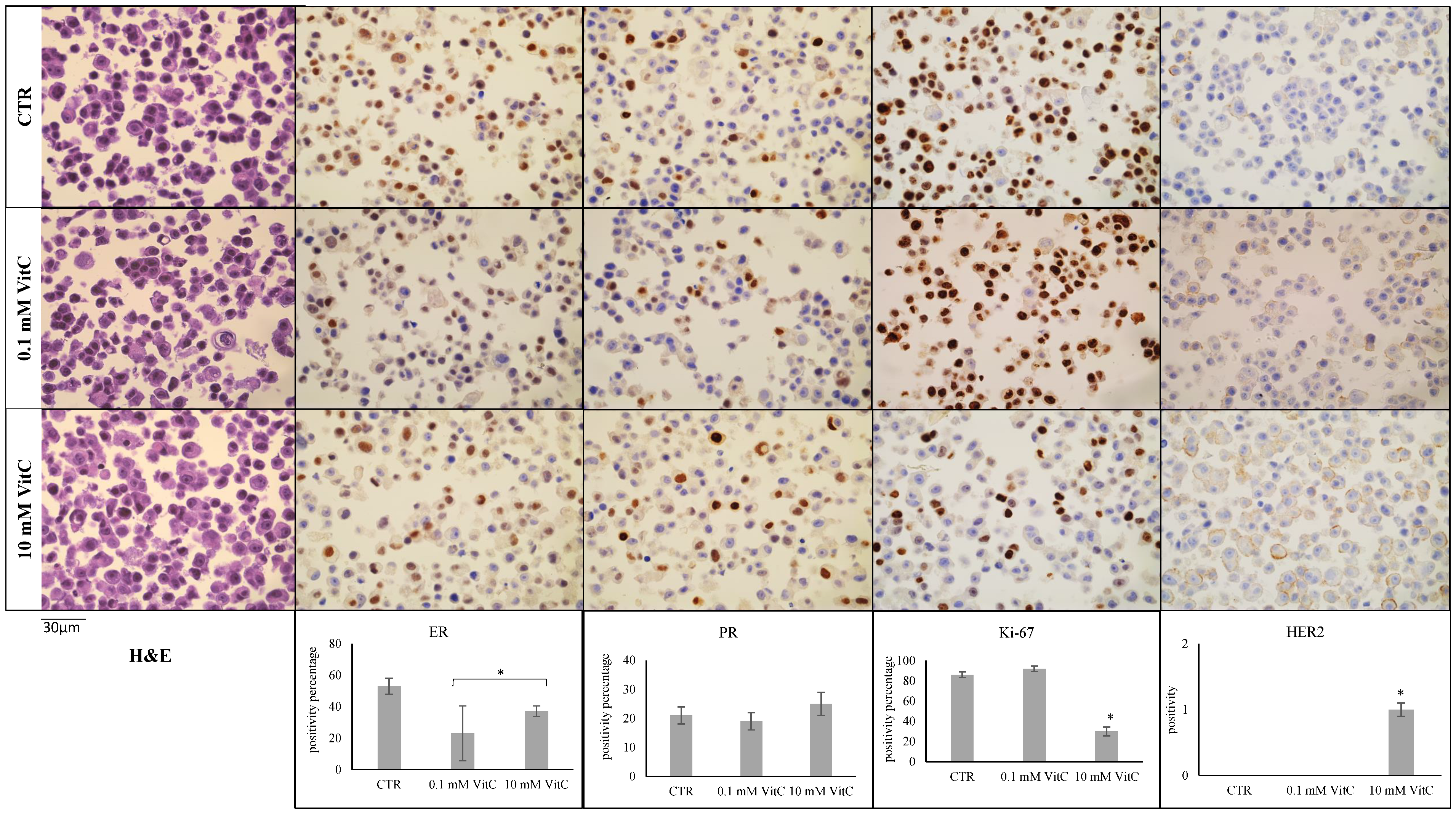
2.1. Effect of Vitamin C on MCF7 and MB-231 Cell Phenotypes
2.2. Vitamin C Influences Sphingomyelin Metabolism in Human Breast Cancer
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture and Treatments
4.3. Cell Viability
4.4. Immunocytochemistry
4.5. Reverse Transcription Quantitative PCR (RTqPCR)
4.6. Protein Concentration and Western Blotting
4.7. Ultrafast Liquid Chromatography–Tandem Mass Spectrometry
4.8. Immunofluorescence
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smirnoff, N.; Conklin, P.L.; Loewus, F.A. Biosynthesis of Ascorbic Acid in Plants: A Renaissance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 437–467. [Google Scholar] [CrossRef]
- Linster, C.L.; Van Schaftingen, E. Vitamin C. Biosynthesis, recycling and degradation in mammals. FEBS J. 2007, 274, 1–22. [Google Scholar] [CrossRef]
- Tsukaguchi, H.; Tokui, T.; Mackenzie, B.; Berger, U.V.; Chen, X.Z.; Wang, Y.; Brubaker, R.F.; Hediger, M.A. A family of mammalian Na+-dependent L-ascorbic acid transporters. Nature 1999, 399, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Hasselholt, S.; Tveden-Nyborg, P.; Lykkesfeldt, J. Distribution of vitamin C is tissue specific with early saturation of the brain and adrenal glands following differential oral dose regimens in guinea pigs. Br. J. Nutr. 2015, 113, 1539–1549. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Poulsen, H.E. Is vitamin C supplementation beneficial? Lessons learned from randomised controlled trials. Br. J. Nutr. 2010, 103, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.; Frei, B. Does vitamin C act as a pro-oxidant under physiological conditions? FASEB J. 1999, 13, 1007–1024. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Loft, S.; Nielsen, J.B.; Poulsen, H.E. Ascorbic acid and dehydroascorbic acid as biomarkers of oxidative stress caused by smoking. Am. J. Clin. Nutr. 1997, 65, 959–963. [Google Scholar] [CrossRef]
- Ting, H.H.; Timimi, F.K.; Boles, K.S.; Creager, S.J.; Ganz, P.; Creager, M.A. Vitamin C improves endothelium-dependent vasodilation in patients with non-insulin-dependent diabetes mellitus. J. Clin. Investig. 1996, 97, 22–28. [Google Scholar] [CrossRef]
- Ramdas, W.D.; Schouten, J.; Webers, C.A.B. The Effect of Vitamins on Glaucoma: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 359. [Google Scholar] [CrossRef]
- Seddon, J.M.; Ajani, U.A.; Sperduto, R.D.; Hiller, R.; Blair, N.; Burton, T.C.; Farber, M.D.; Gragoudas, E.S.; Haller, J.; Miller, D.T.; et al. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study Group. JAMA 1994, 272, 1413–1420. [Google Scholar] [CrossRef]
- Chen, G.C.; Lu, D.B.; Pang, Z.; Liu, Q.F. Vitamin C intake, circulating vitamin C and risk of stroke: A meta-analysis of prospective studies. J. Am. Heart Assoc. 2013, 2, e000329. [Google Scholar] [CrossRef] [PubMed]
- Cameron, E.; Pauling, L. Supplemental ascorbate in the supportive treatment of cancer: Prolongation of survival times in terminal human cancer. Proc. Natl. Acad. Sci. USA 1976, 73, 3685–3689. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Miyake, T.; Tani, M.; Uemoto, S. Diverse antitumor effects of ascorbic acid on cancer cells and the tumor microenvironment. Front. Oncol. 2022, 12, 981547. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Huang, Y.; Cao, D.; Qiu, S.; Chen, B.; Li, J.; Bao, Y.; Wei, Q.; Han, P.; Liu, L. Vitamin C Intake and Cancers: An Umbrella Review. Front. Nutr. 2022, 8, 812394. [Google Scholar] [CrossRef] [PubMed]
- Villagran, M.; Ferreira, J.; Martorell, M.; Mardones, L. The Role of Vitamin C in Cancer Prevention and Therapy: A Literature Review. Antioxidants 2021, 10, 1894. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.; Van Riper, J.M.; Cantley, L.C.; Yun, J. Targeting cancer vulnerabilities with high-dose vitamin C. Nat. Rev. Cancer 2019, 19, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Unlu, A.; Kirca, O.; Ozdogan, M.; Nayır, E. High-dose vitamin C and cancer. J. Oncol. Sci. 2016, 1, 10–12. [Google Scholar] [CrossRef]
- Padayatty, S.J.; Sun, H.; Wang, Y.; Riordan, H.D.; Hewitt, S.M.; Katz, A.; Wesley, R.A.; Levine, M. Vitamin C pharmacokinetics: Implications for oral and intravenous use. Ann. Intern. Med. 2004, 140, 533–537. [Google Scholar] [CrossRef]
- Hoffer, L.J.; Levine, M.; Assouline, S.; Melnychuk, D.; Padayatty, S.J.; Rosadiuk, K.; Rousseau, C.; Robitaille, L.; Miller, W.H., Jr. Phase I clinical trial of i.v. ascorbic acid in advanced malignancy. Ann. Oncol. 2008, 19, 1969–1974. [Google Scholar] [CrossRef]
- Zasowska-Nowak, A.; Nowak, P.J.; Ciałkowska-Rysz, A. High-Dose Vitamin C in Advanced-Stage Cancer Patients. Nutrients 2021, 13, 735. [Google Scholar] [CrossRef]
- Mayland, C.R.; Bennett, M.I.; Allan, K. Vitamin C deficiency in cancer patients. Palliat. Med. 2005, 19, 17–20. [Google Scholar] [CrossRef]
- Park, S. The effects of high concentrations of vitamin C on cancer cells. Nutrients 2013, 5, 3496–3505. [Google Scholar] [CrossRef] [PubMed]
- Poulter, J.M.; White, W.F.; Dickerson, J. W Ascorbic acid supplementation and five year survival rates in women with early breast cancer. Acta Vitaminol. Enzymol. 1984, 6, 175–182. [Google Scholar] [PubMed]
- Harris, H.R.; Orsini, N.; Wolk, A. Vitamin C and survival among women with breast cancer: A meta-analysis. Eur. J. Cancer 2014, 50, 1223–1231. [Google Scholar] [CrossRef]
- Codini, M. Why Vitamin C Could Be an Excellent Complementary Remedy to Conventional Therapies for Breast Cancer. Int. J. Mo.l Sci. 2020, 21, 8397. [Google Scholar] [CrossRef]
- Albi, E.; Mandarano, M.; Cataldi, S.; Ceccarini, M.R.; Fiorani, F.; Beccari, T.; Sidoni, A.; Codini, M. The Effect of Cholesterol in MCF7 Human Breast Cancer Cells. Int. J. Mol. Sci. 2023, 24, 5935. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wu, S.; Liu, H.; Luo, Z.; Sun, J.; Jin, X. Relationship between food-derived antioxidant vitamin intake and breast cancer risk: A mendelian randomized study. Eur. J. Nutr. 2023, 62, 2365–2373. [Google Scholar] [CrossRef]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef]
- Subramani, T.; Yeap, S.K.; Ho, W.Y.; Ho, C.L.; Omar, A.R.; Aziz, S.A.; Rahman, N.M.; Alitheen, N.B. Vitamin C suppresses cell death in MCF-7 human breast cancer cells induced by tamoxifen. J. Cell. Mol. Med. 2014, 18, 305–313. [Google Scholar] [CrossRef]
- Bober, P.; Alexovic, M.; Talian, I.; Tomkova, Z.; Viscorova, Z.; Benckova, M.; Andrasina, I.; Ciccocioppo, R.; Petrovic, D.; Adamek, M.; et al. Proteomic analysis of the vitamin C effect on the doxorubicin cytotoxicity in the MCF-7 breast cancer cell line. J. Cancer Res. Clin. Oncol. 2017, 143, 35–42. [Google Scholar] [CrossRef]
- Ibrahim, S.S.A.; El-Aal, S.A.A.; Reda, A.M.; Achy, S.E.; Shahine, Y. Anti-neoplastic action of Cimetidine/Vitamin C on histamine and the PI3K/AKT/mTOR pathway in Ehrlich breast cancer. Sci. Rep. 2022, 12, 11514. [Google Scholar] [CrossRef] [PubMed]
- Khazaei, S.; Nilsson, L.; Adrian, G.; Tryggvadottir, H.; Konradsson, E.; Borgquist, S.; Isaksson, K.; Ceberg, C.; Jernström, H. Impact of combining vitamin C with radiation therapy in human breast cancer: Does it matter? Oncotarget 2022, 13, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Corsetto, P.A.; Zava, S.; Rizzo, A.M.; Colombo, I. The Critical Impact of Sphingolipid Metabolism in Breast Cancer Progression and Drug Response. Int. J. Mol. Sci. 2023, 24, 2107. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, P.; Conte, C.; Albi, E. The Multiple Roles of Sphingomyelin in Parkinson’s Disease. Biomolecules 2021, 11, 1311. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Atilla-Gokcumen, G.E.; Frasor, J. Emerging Roles of Ceramides in Breast Cancer Biology and Therapy. Int. J. Mol. Sci. 2022, 23, 11178. [Google Scholar] [CrossRef]
- Kar, A.; Medatwal, N.; Rajput, K.; Mandal, S.; Pani, T.; Khan, A.; Sharma, P.; Oberoi, A.S.; Vishwakarma, G.; Deo, S.; et al. Unique sphingolipid signature identifies luminal and triple-negative breast cancer subtypes. Int. J. Cancer 2023, 152, 2410–2423. [Google Scholar] [CrossRef]
- Rajput, K.; Ansari, M.N.; Jha, S.K.; Pani, T.; Medatwal, N.; Chattopadhyay, S.; Bajaj, A.; Dasgupta, U. Ceramide Kinase (CERK) Emerges as a Common Therapeutic Target for Triple Positive and Triple Negative Breast Cancer Cells. Cancers 2022, 14, 4496. [Google Scholar] [CrossRef]
- Huang, C.; Su, L.; Chen, Y.; Wu, S.; Sun, R.; Xu, Q.; Qiu, X.; Yang, C.; Kong, X.; Qin, H.; et al. Ceramide kinase confers tamoxifen resistance in estrogen receptor-positive breast cancer by altering sphingolipid metabolism. Pharmacol. Res. 2023, 187, 106558. [Google Scholar] [CrossRef]
- Bhadwal, P.; Randhawa, V.; Vaiphei, K.; Dahiya, D.; Agnihotri, N. Clinical relevance of CERK and SPHK1 in breast cancer and their association with metastasis and drug resistance. Sci. Rep. 2022, 12, 18239. [Google Scholar] [CrossRef]
- Hanikoglu, A.; Kucuksayan, E.; Hanikoglu, F.; Ozben, T.; Menounou, G.; Sansone, A.; Chatgilialoglu, C.; Di Bella, G.; Ferreri, C. Effects of Somatostatin and Vitamin C on the Fatty Acid Profile of Breast Cancer Cell Membranes. Anticancer Agents Med. Chem. 2019, 19, 1899–1909. [Google Scholar] [CrossRef]
- Kim, J.; Yun, H.; Cho, Y. Analysis of ceramide metabolites in differentiating epidermal keratinocytes treated with calcium or vitamin C. Nutr. Res. Pract. 2011, 5, 396–403. [Google Scholar] [CrossRef]
- Raheem, F.; Karikalan, S.A.; Batalini, F.; El Masry, A.; Mina, L. Metastatic ER+ Breast Cancer: Mechanisms of Resistance and Future Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 16198. [Google Scholar] [CrossRef]
- Santoro, A.L.; Drummond, R.D.; Silva, I.T.; Ferreira, S.S.; Juliano, L.; Vendramini, P.H.; Lemos, M.B.D.C.; Eberlin, M.N.; Andrade, V.P. In Situ DESI-MSI Lipidomic Profiles of Breast Cancer Molecular Subtypes and Precursor Lesions. Cancer Res. 2020, 80, 1246–1257. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Jeong, J.H.; Lee, I.H.; Lee, J.; Jung, J.H.; Park, H.Y.; Lee, D.H.; Chae, Y.S. Effect of High-dose Vitamin C Combined With Anti-cancer Treatment on Breast Cancer Cells. Anticancer Res. 2019, 39, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi-Pour, Z.; Ramezani, F.; Keshavarzi, F.; Samadi, N. The role of quercetin and vitamin C in Nrf2-dependent oxidative stress production in breast cancer cells. Oncol Lett. 2017, 13, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Coombes, R.C. Endocrine-responsive breast cancer and strategies for combating resistance. Nat. Rev. Cancer 2002, 2, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, Z.; Liu, Y.; Ding, X.; Wang, Y.; Fan, S. The ceramide synthase (CERS/LASS) family: Functions involved in cancer progression. Cell. Oncol. 2023, 46, 825–845. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; He, B.; Ke, F. Ceramide Synthase 6 Mediates Triple-Negative Breast Cancer Response to Chemotherapy through RhoA- and EGFR-Mediated Signaling Pathways. J. Breast Cancer 2022, 25, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Knox, A.J.; Scaling, A.L.; Pinto, M.P.; Bliesner, B.S.; Haughian, J.M.; Abdel-Hafiz, H.A.; Horwitz, K.B. Modeling luminal breast cancer heterogeneity: Combination therapy to suppress a hormone receptor-negative, cytokeratin 5-positive subpopulation in luminal disease. Breast Cancer Res. 2014, 16, 418. [Google Scholar] [CrossRef]
- Cataldi, S.; Ceccarini, M.R.; Patria, F.; Beccari, T.; Mandarano, M.; Ferri, I.; Lazzarini, A.; Curcio, F.; Albi, E. The Effect of Vitamin D3 and Silver Nanoparticles on HaCaT Cell Viability. Int. J. Mol. Sci. 2022, 23, 1410. [Google Scholar] [CrossRef]
- Oz, E.S.; Aydemir, E.; Fışkın, K. DMSO exhibits similar cytotoxicity effects to thalidomide in mouse breast cancer cells. Oncol. Lett. 2012, 3, 927–929. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.C.; Hammond, M.E.H.; Allison, K.H.; Harvey, B.E.; Mangu, P.B.; Bartlett, J.M.S.; Bilous, M.; Ellis, I.O.; Fitzgibbons, P.; Hanna, W.; et al. Human Epidermal Growth Factor Receptor 2 Testing in Breast Cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice Guideline Focused Update. J Clin. Oncol. 2018, 36, 2105–2122. [Google Scholar] [CrossRef] [PubMed]
- Codini, M.; Conte, C.; Cataldi, S.; Arcuri, C.; Lazzarini, A.; Ceccarini, M.R.; Patria, F.; Floridi, A.; Mecca, C.; Ambesi-Impiombato, F.S.; et al. Nuclear Lipid Microdomains Regulate Daunorubicin Resistance in Hepatoma Cells. Int. J. Mol. Sci. 2018, 19, 3424. [Google Scholar] [CrossRef] [PubMed]
- Conte, C.; Cataldi, S.; Arcuri, C.; Mirarchi, A.; Lazzarini, A.; Garcia-Gil, M.; Beccari, T.; Curcio, F.; Albi, E. Vitamin D3 Enriches Ceramide Content in Exosomes Released by Embryonic Hippocampal Cells. Int. J. Mol. Sci. 2021, 22, 9287. [Google Scholar] [CrossRef]
- Albi, E.; Arcuri, C.; Kobayashi, T.; Tomishige, N.; Cas, M.D.; Paroni, R.; Signorelli, P.; Cerquiglini, L.; Troiani, S.; Gizzi, C.; et al. Sphingomyelin in Human Breast Milk might be Essential for the Hippocampus Maturation. Front. Biosci. (Landmark Ed.) 2022, 27, 247. [Google Scholar] [CrossRef]
- Tomishige, N.; Murate, M.; Didier, P.; Richert, L.; Mely, Y.; Kobayashi, T. The use of pore-forming toxins to image lipids and lipid domains. Methods Enzymol. 2021, 649, 503–542. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Codini, M.; Fiorani, F.; Mandarano, M.; Cataldi, S.; Arcuri, C.; Mirarchi, A.; Ceccarini, M.R.; Beccari, T.; Kobayashi, T.; Tomishige, N.; et al. Sphingomyelin Metabolism Modifies Luminal A Breast Cancer Cell Line under a High Dose of Vitamin C. Int. J. Mol. Sci. 2023, 24, 17263. https://doi.org/10.3390/ijms242417263
Codini M, Fiorani F, Mandarano M, Cataldi S, Arcuri C, Mirarchi A, Ceccarini MR, Beccari T, Kobayashi T, Tomishige N, et al. Sphingomyelin Metabolism Modifies Luminal A Breast Cancer Cell Line under a High Dose of Vitamin C. International Journal of Molecular Sciences. 2023; 24(24):17263. https://doi.org/10.3390/ijms242417263
Chicago/Turabian StyleCodini, Michela, Federico Fiorani, Martina Mandarano, Samuela Cataldi, Cataldo Arcuri, Alessandra Mirarchi, Maria Rachele Ceccarini, Tommaso Beccari, Toshihide Kobayashi, Nario Tomishige, and et al. 2023. "Sphingomyelin Metabolism Modifies Luminal A Breast Cancer Cell Line under a High Dose of Vitamin C" International Journal of Molecular Sciences 24, no. 24: 17263. https://doi.org/10.3390/ijms242417263
APA StyleCodini, M., Fiorani, F., Mandarano, M., Cataldi, S., Arcuri, C., Mirarchi, A., Ceccarini, M. R., Beccari, T., Kobayashi, T., Tomishige, N., Sidoni, A., & Albi, E. (2023). Sphingomyelin Metabolism Modifies Luminal A Breast Cancer Cell Line under a High Dose of Vitamin C. International Journal of Molecular Sciences, 24(24), 17263. https://doi.org/10.3390/ijms242417263