
Involvement of TRPM7 in Alcohol-Induced Damage of the Blood–Brain Barrier in the Presence of HIV Viral Proteins

Abstract
:1. Introduction
2. Results
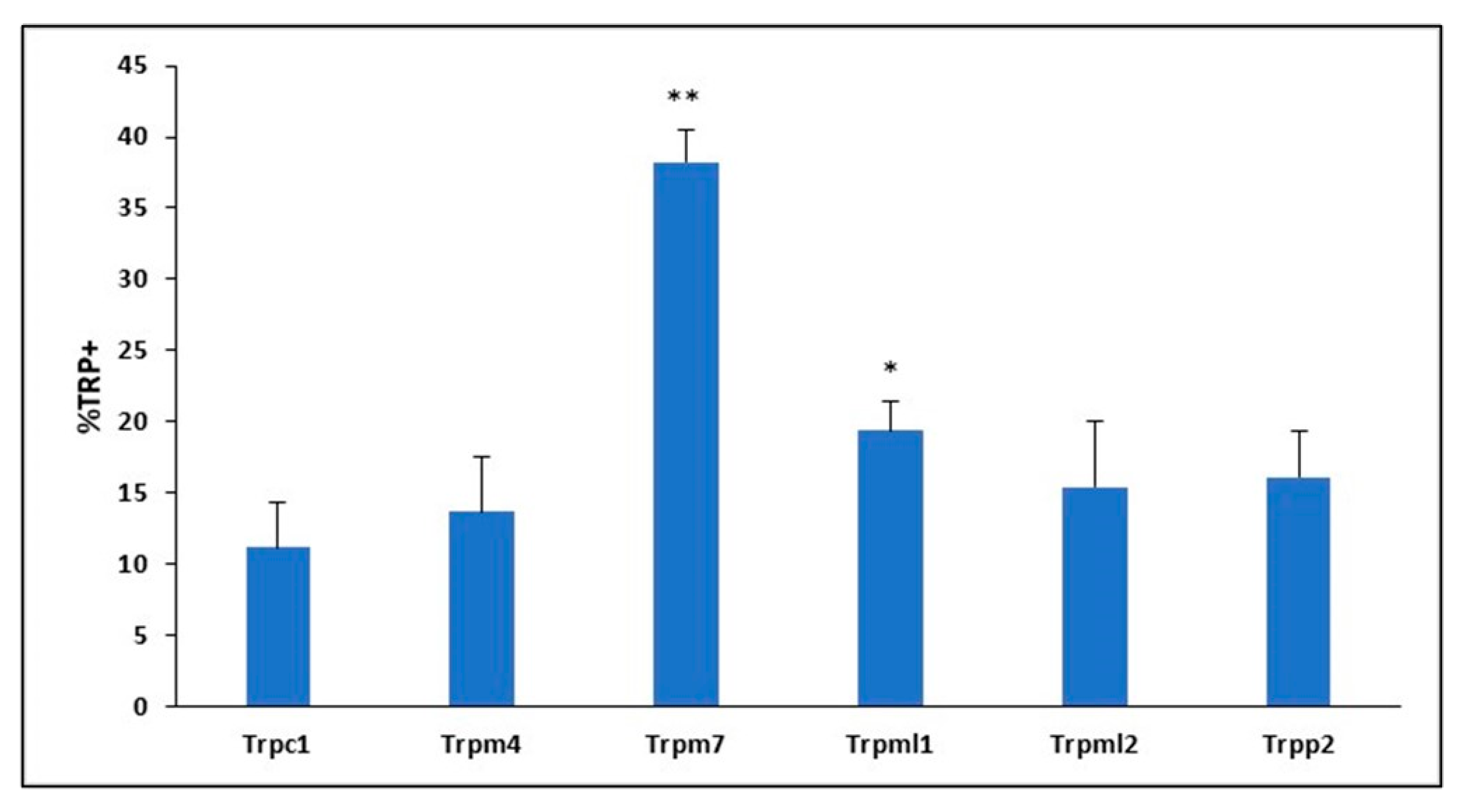
2.1. Members of 6 TRP Subfamilies Are Expressed in Mouse BMVECs
2.2. TRP Channel Protein Expression in Mouse BMVECs
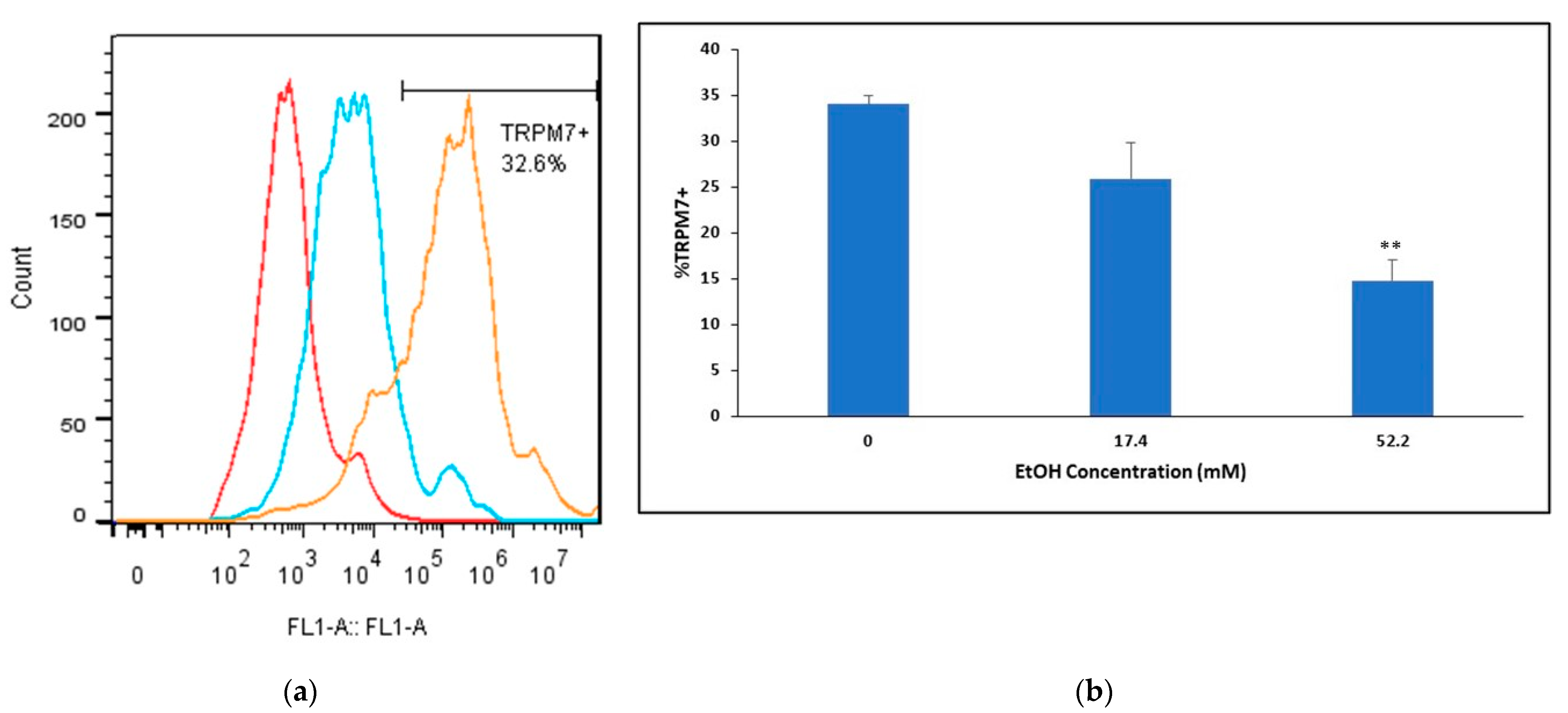
2.3. Ethanol’s Effects on TRPM7 Protein Expression in Mouse BMVECs
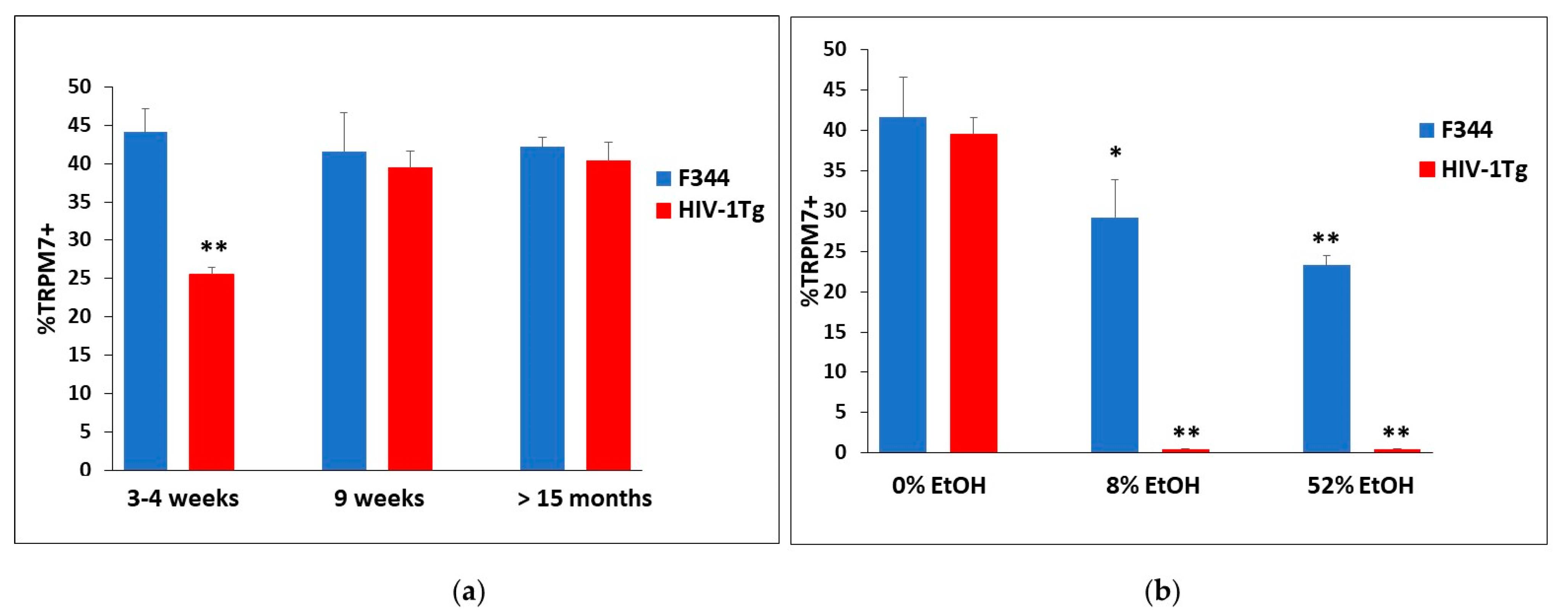
2.4. Ethanol’s Modulation on the Differential Expression of TRPM7 in BMVECs from F344 and HIV-1Tg Rats
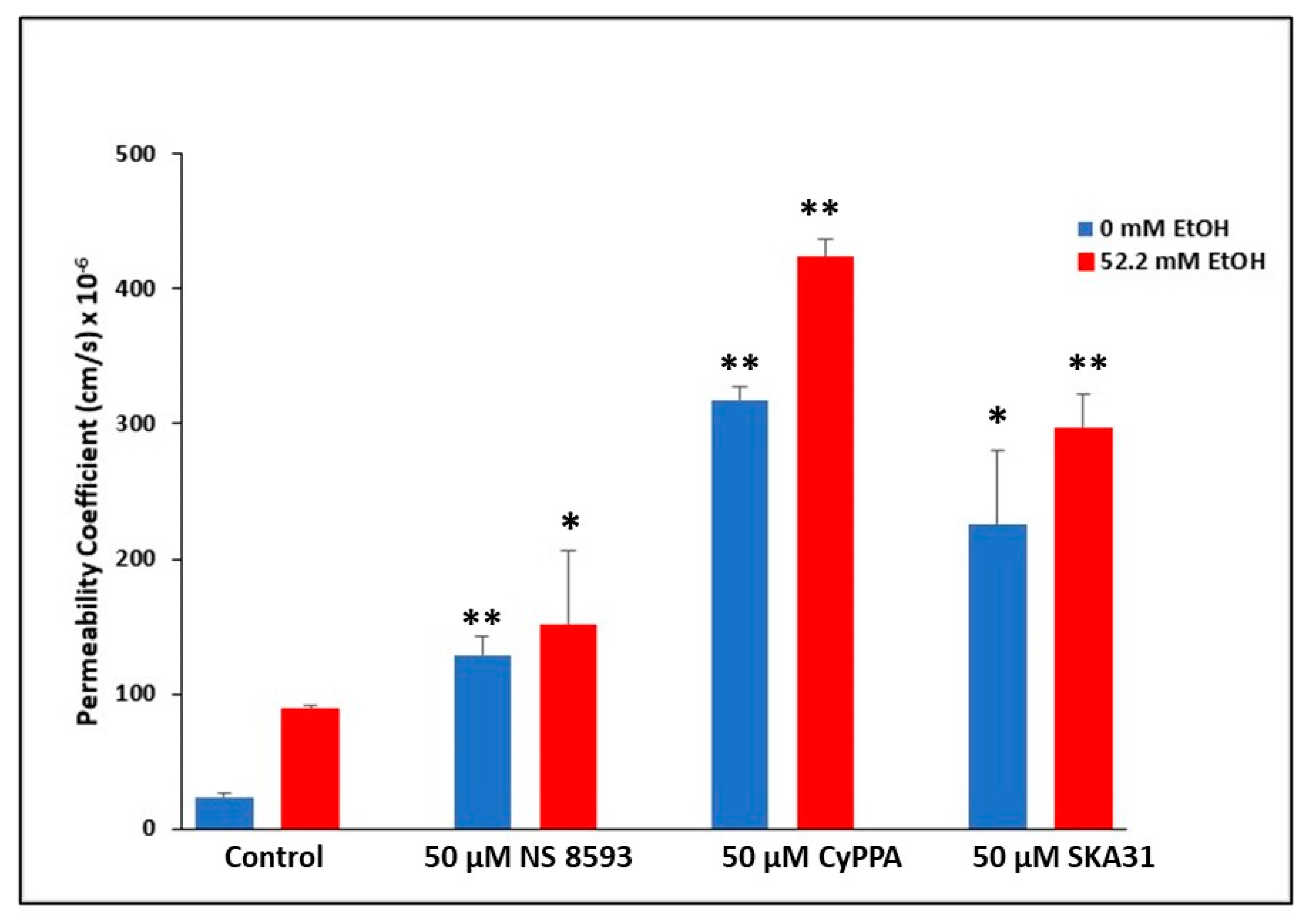
2.5. Involvement of TRPM7 in Ethanol-Mediated BBB Permeability Changes
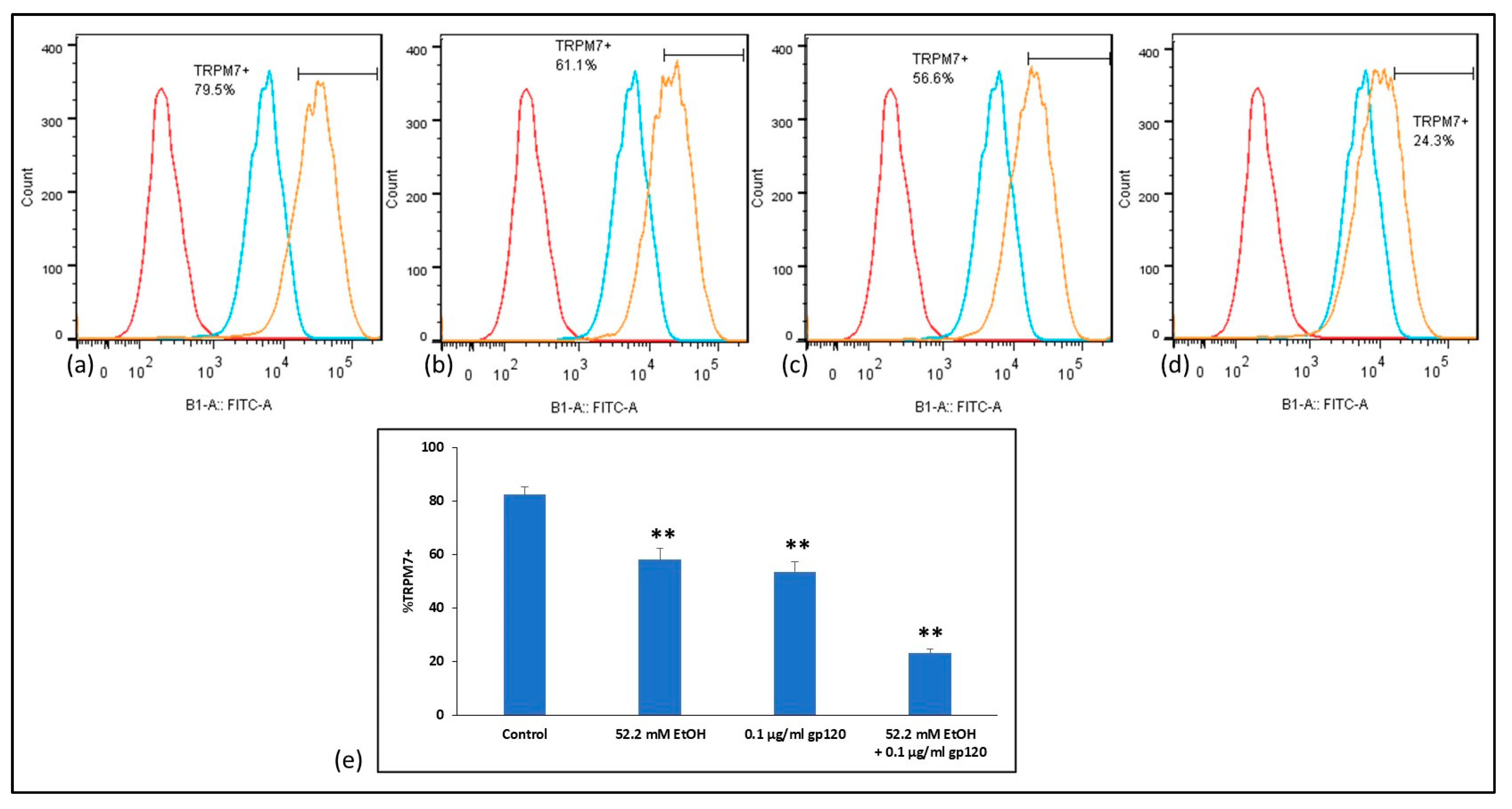
2.6. Analysis of TRPM7 Expression in Human BMVECs after Ethanol and HIV-1 gp120 Combination Treatment
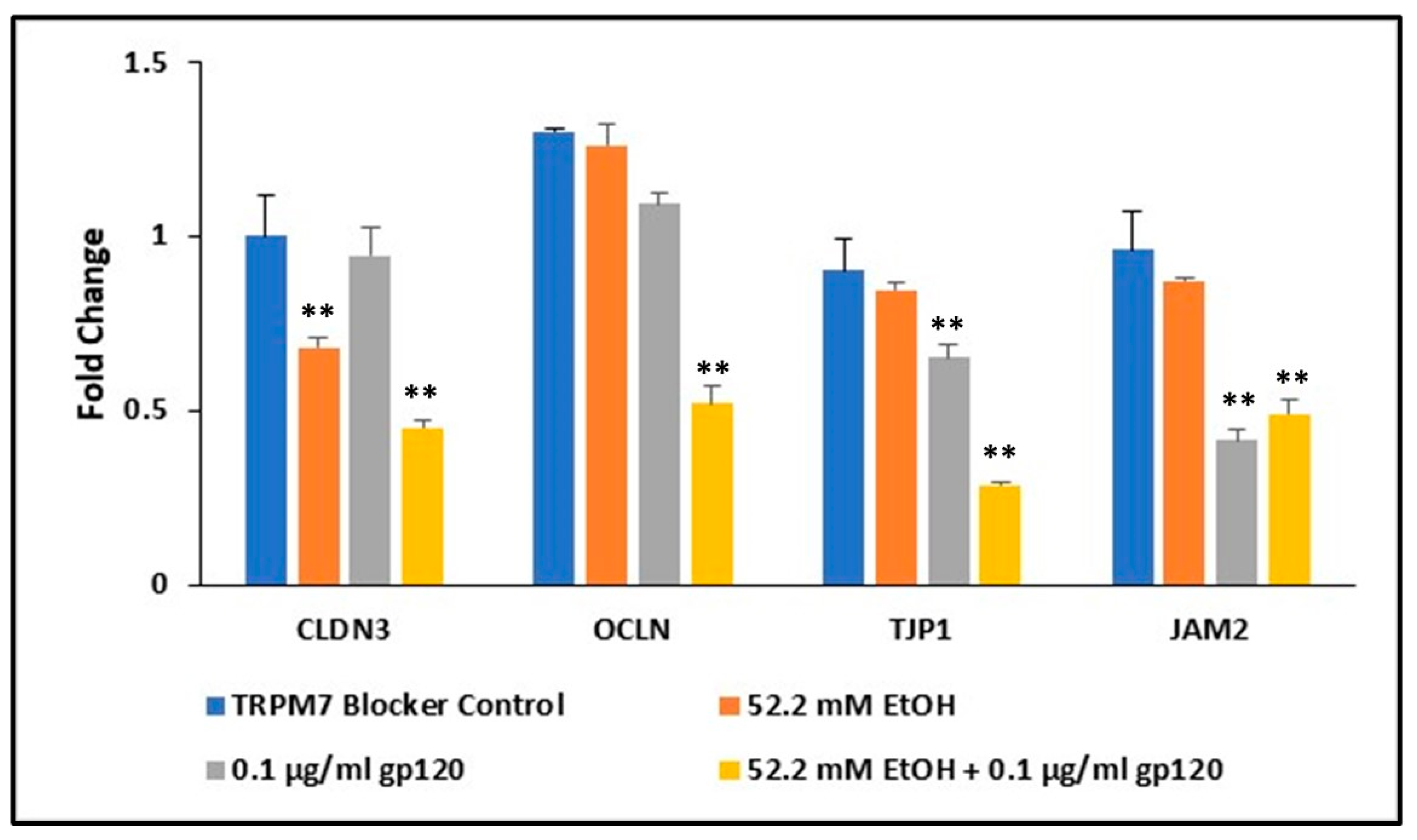
2.7. Ethanol and HIV-1 Viral Protein gp120 Effects on Tight Junction Complexes
2.8. Involvement of TRPM7 in Alcohol Induced Damage of the BBB in the Presence of HIV Viral Protein gp120
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. EtOH Administration
4.3. Tissue Collection and BMVEC Isolation
4.4. Cell Culture
4.5. Flow Cytometry
4.6. In Vitro BBB Permeability Assay
4.7. RNA Isolation, Purification, Reverse Transcription, and Quantitative RT-PCR Array
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reid, W.; Sadowska, M.; Denaro, F.; Rao, S.; Foulke, J.; Hayes, N.; Jones, O.; Doodnauth, D.; Davis, H.; Sill, A.; et al. An HIV-1 transgenic rat that develops HIV-related pathology and immunologic dysfunction. Proc. Natl. Acad. Sci. USA 2001, 98, 9271–9276. [Google Scholar] [CrossRef] [Green Version]
- Mazzucchelli, R.; Amadio, M.; Curreli, S.; Denaro, F.; Bemis, K.; Reid, W.; Bryant, J.; Riva, A.; Galli, M.; Zella, D. Establishment of an ex vivo model of monocytes-derived macrophages differentiated from peripheral blood mononuclear cells (PBMCs) from HIV-1 transgenic rats. Mol. Immunol. 2004, 41, 979–984. [Google Scholar] [CrossRef]
- Reid, W.; Abdelwahab, S.; Sadowska, M.; Huso, D.; Neal, A.; Ahearn, A.; Bryant, J.; Gallo, R.C.; Lewis, G.K.; Reitz, M. HIV-1 transgenic rats develop T cell abnormalities. Virology 2004, 321, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Vigorito, M.; Connaghan, K.P.; Chang, S.L. The HIV-1 transgenic rat model of neuroHIV. Brain, Behav. Immun. 2015, 48, 336–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.L.; Beltran, J.A.; Swarup, S. Expression of the Mu Opioid Receptor in the Human Immunodeficiency Virus Type 1 Transgenic Rat Model. J. Virol. 2007, 81, 8406–8411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigorito, M.; Lashomb, A.L.; Chang, S.L. Spatial Learning and Memory in HIV-1 Transgenic Rats. J. Neuroimmune Pharmacol. 2007, 2, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Chang, S.L. Ethanol Concentration-Dependent Alterations in Gene Expression During Acute Binge Drinking in the HIV-1 Transgenic Rat. Alcohol. Clin. Exp. Res. 2013, 37, 1082–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.; Mao, X.; Liu, C.; Chang, S.L. Age- and Ethanol Concentration-Dependent Effects of Acute Binge Drinking in the HIV-1 Transgenic Rat. Alcohol. Clin. Exp. Res. 2013, 37 (Suppl. 1), E70–E78. [Google Scholar] [CrossRef]
- Benedikt, J.; Teisinger, J.; Vyklicky, L.; Vlachova, V. Ethanol inhibits cold-menthol receptor TRPM8 by modulating its interaction with membrane phosphatidylinositol 4,5-bisphosphate. J. Neurochem. 2007, 100, 211–224. [Google Scholar] [CrossRef]
- Blednov, Y.; Harris, R. Deletion of vanilloid receptor (TRPV1) in mice alters behavioral effects of ethanol. Neuropharmacology 2009, 56, 814–820. [Google Scholar] [CrossRef]
- Cornett, P.M.; Matta, J.A.; Ahern, G.P. General Anesthetics Sensitize the Capsaicin Receptor Transient Receptor Potential V1. Mol. Pharmacol. 2008, 74, 1261–1268. [Google Scholar] [CrossRef] [Green Version]
- Ellingson, J.M.; Silbaugh, B.C.; Brasser, S.M. Reduced Oral Ethanol Avoidance in Mice Lacking Transient Receptor Potential Channel Vanilloid Receptor 1. Behav. Genet. 2009, 39, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, R.J.; Slesinger, P.A.; Davies, D.L.; Das, J.; Trudell, J.R.; Harris, R.A. Alcohol-Binding Sites in Distinct Brain Proteins: The Quest for Atomic Level Resolution. Alcohol. Clin. Exp. Res. 2011, 35, 1561–1573. [Google Scholar] [CrossRef]
- Matta, J.A.; Cornett, P.M.; Miyares, R.L.; Abe, K.; Sahibzada, N.; Ahern, G.P. General anesthetics activate a nociceptive ion channel to enhance pain and inflammation. Proc. Natl. Acad. Sci. USA 2008, 105, 8784–8789. [Google Scholar] [CrossRef] [Green Version]
- Trevisani, M.; Smart, D.J.; Gunthorpe, M.J.; Tognetto, M.; Barbieri, M.; Campi, B.; Amadesi, S.; Gray, J.C.; Jerman, J.C.; Brough, S.J.; et al. Ethanol elicits and potentiates nociceptor responses via the vanilloid receptor-1. Nat. Neurosci. 2002, 5, 546–551. [Google Scholar] [CrossRef]
- Visser, D.; Middelbeek, J.; van Leeuwen, F.N.; Jalink, K. Function and regulation of the channel-kinase TRPM7 in health and disease. Eur. J. Cell Biol. 2014, 93, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Zaelzer, C.; Brauchi, S. Structure–functional intimacies of transient receptor potential channels. Q. Rev. Biophys. 2009, 42, 201–246. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Voets, T.; Talavera, K.; Owsianik, G.; Nilius, B. Sensing with TRP channels. Nat. Chem. Biol. 2005, 1, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Bandell, M.; Macpherson, L.J.; Patapoutian, A. From chills to chilis: Mechanisms for thermosensation and chemesthesis via thermoTRPs. Curr. Opin. Neurobiol. 2007, 17, 490–497. [Google Scholar] [CrossRef]
- Patapoutian, A.; Peier, A.M.; Story, G.M.; Viswanath, V. ThermoTRP channels and beyond: Mechanisms of temperature sensation. Nat. Rev. Neurosci. 2003, 4, 529–539. [Google Scholar] [CrossRef]
- Cortright, D.N.; Szallasi, A. TRP channels and pain. Curr. Pharm. Des. 2009, 15, 1736–1749. [Google Scholar] [CrossRef]
- Montell, C. The TRP Superfamily of Cation Channels. Sci. STKE 2005, 2005, re3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An Introduction to TRP Channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwanyanya, A.; Sipido, K.R.; Vereecke, J.; Mubagwa, K. ATP and PIP2 dependence of the magnesium-inhibited, TRPM7-like cation channel in cardiac myocytes. Am. J. Physiol. Cell Physiol. 2006, 291, C627–C635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, J.A.; Matsushita, M.; Nairn, A.C.; Cahalan, M.D. Charge Screening by Internal pH and Polyvalent Cations as a Mechanism for Activation, Inhibition, and Rundown of TRPM7/MIC Channels. J. Gen. Physiol. 2005, 126, 499–514. [Google Scholar] [CrossRef] [Green Version]
- Langeslag, M.; Clark, K.; Moolenaar, W.H.; van Leeuwen, F.N.; Jalink, K. Activation of TRPM7 Channels by Phospholipase C-coupled Receptor Agonists. J. Biol. Chem. 2007, 282, 232–239. [Google Scholar] [CrossRef] [Green Version]
- Macianskiene, R.; Gwanyanya, A.; Vereecke, J.; Mubagwa, K. Inhibition of the Magnesium-Sensitive TRPM7-like Channel in Cardiac Myocytes by Nonhydrolysable GTP Analogs: Involvement of Phosphoinositide Metabolism. Cell. Physiol. Biochem. 2008, 22, 109–118. [Google Scholar] [CrossRef]
- Oh, H.G.; Chun, Y.S.; Kim, Y.; Youn, S.H.; Shin, S.; Park, M.K.; Kim, T.-W.; Chung, S. Modulation of transient receptor potential melastatin related 7 channel by presenilins. Dev. Neurobiol. 2012, 72, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Runnels, L.; Yue, L.; Clapham, D. The TRPM7 channel is inactivated by PIP2 hydrolysis. Nature 2002, 4, 329–336. [Google Scholar] [CrossRef]
- Xie, J.; Sun, B.; Du, J.; Yang, W.; Chen, H.C.; Overton, J.D.; Runnels, L.W.; Yue, L. Phosphatidylinositol 4,5-bisphosphate (PIP(2)) controls magnesium gatekeeper TRPM6 activity. Sci. Rep. 2011, 1, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abed, E.; Moreau, R. Importance of melastatin-like transient receptor potential 7 and cations (magnesium, calcium) in human osteoblast-like cell proliferation. Cell Prolif. 2007, 40, 849–865. [Google Scholar] [CrossRef] [Green Version]
- Chubanov, V.; Mederos y Schnitzler, M.; Meißner, M.; Schäfer, S.; Abstiens, K.; Hofmann, T.; Gudermann, T. Natural and synthetic modulators of SK (K(ca)2) potassium channels inhibit magnesium-dependent activity of the kinase-coupled cation channel TRPM7. Br. J. Pharmacol. 2012, 166, 1357–1376. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Xie, J.; Zhang, Z.; Tsujikawa, H.; Fusco, D.; Silverman, D.; Liang, B.; Yue, L. TRPM7-Mediated Ca 2+ Signals Confer Fibrogenesis in Human Atrial Fibrillation. Circ. Res. 2010, 106, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Elizondo, M.R.; Arduini, B.L.; Paulsen, J.; MacDonald, E.L.; Sabel, J.L.; Henion, P.D.; Cornell, R.A.; Parichy, D.M. Defective skeletogenesis with kidney stone formation in dwarf zebrafish mutant for trpm7. Curr. Biol. 2005, 15, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Li, M.-H.; Inoue, K.; Chu, X.-P.; Seeds, J.; Xiong, Z.-G. Transient Receptor Potential Melastatin 7–like Current in Human Head and Neck Carcinoma Cells: Role in Cell Proliferation. Cancer Res 2007, 67, 10929–10938. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Desai, B.N.; Navarro, B.; Donovan, A.; Andrews, N.C.; Clapham, D.E. Deletion of Trpm7 disrupts embryonic development and thymopoiesis without altering Mg2+ homeostasis. Science 2008, 322, 756–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.J.; Park, E.-J.; Lee, J.H.; Jeon, J.-H.; Kim, S.J.; So, I. Suppression of transient receptor potential melastatin 7 channel induces cell death in gastric cancer. Cancer Sci. 2008, 99, 2502–2509. [Google Scholar] [CrossRef] [PubMed]
- Ryazanova, L.V.; Rondon, L.J.; Zierler, S.; Hu, Z.; Galli, J.; Yamaguchi, T.P.; Mazur, A.; Fleig, A.; Ryazanov, A.G. TRPM7 is essential for Mg2+ homeostasis in mammals. Nat. Commun. 2010, 1, 109. [Google Scholar] [CrossRef] [Green Version]
- Sah, R.; Mesirca, P.; Mason, X.; Gibson, W.; Bates-Withers, C.; Van den Boogert, M.; Chaudhuri, D.; Pu, W.T.; Mangoni, M.E.; Clapham, D.E. Timing of myocardial trpm7 deletion during cardiogenesis variably disrupts adult ventricular function, conduction, and repolarization. Circulation 2013, 128, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Sahni, J.; Tamura, R.; Sweet, I.R.; Scharenberg, A.M. TRPM7 regulates quiescent/proliferative metabolic transitions in lymphocytes. Cell Cycle 2010, 9, 3565–3574. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Selvaraj, S.; Varma, A.; Derry, S.; Sahmoun, A.E.; Singh, B.B. Increase in serum Ca2+/Mg2+ ratio promotes proliferation of prostate cancer cells by activating TRPM7 channels. J. Biol. Chem. 2013, 288, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Yee, N.S.; Zhou, W.; Liang, I.-C. Transient receptor potential ion channel Trpm7 regulates exocrine pancreatic epithelial proliferation by Mg2+-sensitive Socs3a signaling in development and cancer. Dis. Model. Mech. 2011, 4, 240–254. [Google Scholar] [CrossRef] [Green Version]
- Zierler, S.; Yao, G.; Zhang, Z.; Kuo, W.C.; Pörzgen, P.; Penner, R.; Horgen, F.D.; Fleig, A. Waixenicin A Inhibits Cell Proliferation through Magnesium-dependent Block of Transient Receptor Potential Melastatin 7 (TRPM7) Channels. J. Biol. Chem. 2011, 286, 39328–39335. [Google Scholar] [CrossRef] [Green Version]
- Davis, F.M.; Azimi, I.; Faville, R.A.; Peters, A.A.; Jalink, K.; Putney, J.W.; Goodhill, G.J.; Thompson, E.W.; Roberts-Thomson, S.J.; Monteith, G.R. Induction of epithelial–mesenchymal transition (EMT) in breast cancer cells is calcium signal dependent. Oncogene 2014, 33, 2307–2316. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Wu, L.-J.; Jun, J.; Cheng, X.; Xu, H.; Andrews, N.C.; Clapham, D.E. The channel kinase, TRPM7, is required for early embryonic development. Proc. Natl. Acad. Sci. USA 2012, 109, E225–E233. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Su, L.-T.; Khadka, D.K.; Mezzacappa, C.; Komiya, Y.; Sato, A.; Habas, R.; Runnels, L.W. TRPM7 regulates gastrulation during vertebrate embryogenesis. Dev. Biol. 2011, 350, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Low, S.E.; Amburgey, K.; Horstick, E.; Linsley, J.; Sprague, S.M.; Cui, W.W.; Zhou, W.; Hirata, H.; Saint-Amant, L.; Hume, R.I.; et al. TRPM7 Is Required within Zebrafish Sensory Neurons for the Activation of Touch-Evoked Escape Behaviors. J. Neurosci. 2011, 31, 11633–11644. [Google Scholar] [CrossRef] [Green Version]
- Dhennin-Duthille, I.; Gautier, M.; Faouzi, M.; Guilbert, A.; Brevet, M.; Vaudry, D.; Ahidouch, A.; Sevestre, H.; Ouadid-Ahidouch, H. High Expression of Transient Receptor Potential Channels in Human Breast Cancer Epithelial Cells and Tissues: Correlation with Pathological Parameters. Cell. Physiol. Biochem. 2011, 28, 813–822. [Google Scholar] [CrossRef]
- Guilbert, A.; Gautier, M.; Dhennin-Duthille, I.; Haren, N.; Sevestre, H.; Ouadid-Ahidouch, H. Evidence that TRPM7 is required for breast cancer cell proliferation. Am. J. Physiol. Cell Physiol. 2009, 297, C493–C502. [Google Scholar] [CrossRef]
- Guilbert, A.; Gautier, M.; Dhennin-Duthille, I.; Rybarczyk, P.; Sahni, J.; Sevestre, H.; Scharenberg, A.M.; Ouadid-Ahidouch, H. Transient receptor potential melastatin 7 is involved in oestrogen receptor-negative metastatic breast cancer cells migration through its kinase domain. Eur. J. Cancer 2013, 49, 3694–3707. [Google Scholar] [CrossRef] [PubMed]
- Bae, C.Y.-J.; Sun, H.-S. TRPM7 in cerebral ischemia and potential target for drug development in stroke. Acta Pharmacol. Sin. 2011, 32, 725–733. [Google Scholar] [CrossRef]
- Paravicini, T.M.; Yogi, A.; Mazur, A.; Touyz, R.M. Dysregulation of Vascular TRPM7 and Annexin-1 Is Associated With Endothelial Dysfunction in Inherited Hypomagnesemia. Hypertension 2009, 53, 423–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touyz, R.M. Transient receptor potential melastatin 6 and 7 channels, magnesium transport, and vascular biology: Implications in hypertension. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1103–H1118. [Google Scholar] [CrossRef] [Green Version]
- Yogi, A.; Callera, G.E.; Antunes, T.T.; Tostes, R.C.; Touyz, R.M. Transient Receptor Potential Melastatin 7 (TRPM7) Cation Channels, Magnesium and the Vascular System in Hypertension. Circ. J. 2011, 75, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogi, A.; Callera, G.E.; Tostes, R.; Touyz, R.M. Bradykinin regulates calpain and proinflammatory signaling through TRPM7-sensitive pathways in vascular smooth muscle cells. Am. J. Physiol. Integr. Comp. Physiol. 2009, 296, R201–R207. [Google Scholar] [CrossRef] [Green Version]
- Berrout, J.; Jin, M.; O’Neil, R.G. Critical role of TRPP2 and TRPC1 channels in stretch-induced injury of blood–brain barrier endothelial cells. Brain Res. 2012, 1436, 1–12. [Google Scholar] [CrossRef]
- Brown, R.C.; Wu, L.; Hicks, K.; O’Neil, R.G. Regulation of Blood-Brain Barrier Permeability by Transient Receptor Potential Type C and Type V Calcium-Permeable Channels. Microcirculation 2008, 15, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, S.; Al-Ahmad, A.J.; Gassmann, M.; Ogunshola, O.O. Hypoxia selectively disrupts brain microvascular endothelial tight junction complexes through a hypoxia-inducible factor-1 (HIF-1) dependent mechanism. J. Cell. Physiol. 2014, 229, 1096–1105. [Google Scholar] [CrossRef]
- Persidsky, Y.; Ramirez, S.H.; Haorah, J.; Kanmogne, G.D. Blood–brain Barrier: Structural Components and Function under Physiologic and Pathologic Conditions. J. Neuroimmune Pharmacol. 2006, 1, 223–236. [Google Scholar] [CrossRef]
- Burger, D.; Touyz, R.M. Cellular biomarkers of endothelial health: Microparticles, endothelial progenitor cells, and circulating endothelial cells. J. Am. Soc. Hypertens. 2012, 6, 85–99. [Google Scholar] [CrossRef]
- Wen, J.; Doerner, J.; Weidenheim, K.; Xia, Y.; Stock, A.; Michaelson, J.S.; Baruch, K.; Deczkowska, A.; Gulinello, M.; Schwartz, M.; et al. TNF-like weak inducer of apoptosis promotes blood brain barrier disruption and increases neuronal cell death in MRL/lpr mice. J. Autoimmun. 2015, 60, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Calcagno, A.; Alberione, M.C.; Romito, A.; Imperiale, D.; Ghisetti, V.; Audagnotto, S.; Lipani, F.; Raviolo, S.; Di Perri, G.; Bonora, S. Prevalence and predictors of blood-brain barrier damage in the HAART era. J. NeuroVirol. 2014, 20, 521–525. [Google Scholar] [CrossRef]
- Chodobski, A.; Zink, B.J.; Szmydynger-Chodobska, J. Blood–Brain Barrier Pathophysiology in Traumatic Brain Injury. Transl. Stroke Res. 2011, 2, 492–516. [Google Scholar] [CrossRef] [Green Version]
- Haorah, J.; Knipe, B.; Gorantla, S.; Zheng, J.; Persidsky, Y. Alcohol-induced blood-brain barrier dysfunction is mediated via inositol 1,4,5-triphosphate receptor (IP3R)-gated intracellular calcium release. J. Neurochem. 2007, 100, 324–336. [Google Scholar] [CrossRef]
- Persidsky, Y.; Ho, W.; Ramirez, S.H.; Potula, R.; Abood, M.E.; Unterwald, E.; Tuma, R. HIV-1 infection and alcohol abuse: Neurocognitive impairment, mechanisms of neurodegeneration and therapeutic interventions. Brain, Behav. Immun. 2011, 25 (Suppl. 1), S61–S70. [Google Scholar] [CrossRef] [Green Version]
- Haorah, J.; Heilman, D.; Knipe, B.; Chrastil, J.; Leibhart, J.; Ghorpade, A.; Miller, D.W.; Persidsky, Y. Ethanol-Induced Activation of Myosin Light Chain Kinase Leads to Dysfunction of Tight Junctions and Blood-Brain Barrier Compromise. Alcohol. Clin. Exp. Res. 2005, 29, 999–1009. [Google Scholar] [CrossRef]
- McRae, M. HIV and viral protein effects on the blood brain barrier. Tissue Barriers 2016, 4, e1143543. [Google Scholar] [CrossRef] [Green Version]
- Petito, C.K.; Cash, K.S. Blood-brain barrier abnormalities in acquired immunodeficiency syndrome: Immunohistochemical localization of serum proteins in postmortem brain. Ann. Neurol. 1992, 32, 658–666. [Google Scholar] [CrossRef]
- Power, C.; Kong, P.-A.; Crawford, T.O.; Wesselingh, S.; Glass, J.D.; McArthur, J.C.; Trapp, B.D. Cerebral white matter changes in acquired immunodeficiency syndrome dementia: Alterations of the blood-brain barrier. Ann. Neurol. 1993, 34, 339–350. [Google Scholar] [CrossRef]
- Smith, T.W.; DeGirolami, U.; Hénin, D.; Bolgert, F.; Hauw, J.-J. Human Immunodeficiency Virus (HIV) Leukoencephalopathy and the Microcirculation. J. Neuropathol. Exp. Neurol. 1990, 49, 357–370. [Google Scholar] [CrossRef]
- Eugenin, E.A.; Gamss, R.; Buckner, C.; Buono, D.; Klein, R.S.; Schoenbaum, E.E.; Calderon, T.M.; Berman, J.W. Shedding of PECAM-1 during HIV infection: A potential role for soluble PECAM-1 in the pathogenesis of NeuroAIDS. J. Leukoc. Biol. 2006, 79, 444–452. [Google Scholar] [CrossRef] [Green Version]
- Miguez, M.J.; Shor-Posner, G.; Morales, G.; Rodriguez, A.; Burbano, X. HIV treatment in drug abusers: Impact of alcohol use. Addict. Biol. 2003, 8, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Bombardier, C.H.; Temkin, N.R.; Machamer, J.; Dikmen, S.S. The natural history of drinking and alcohol-related problems after traumatic brain injury. Arch. Phys. Med. Rehabil. 2003, 84, 185–191. [Google Scholar] [CrossRef]
- Chang, S.L.; Connaghan, K.P.; Wei, Y.; Li, M.D. NeuroHIV and Use of Addictive Substances. Int. Rev. Neurobiol. 2014, 118, 403–440. [Google Scholar] [CrossRef]
- Galvan, F.H.; Bing, E.G.; Fleishman, J.A.; London, A.S.; Caetano, R.; Burnam, M.A.; Longshore, D.; Morton, S.C.; Orlando, M.; Shapiro, M. The prevalence of alcohol consumption and heavy drinking among people with HIV in the United States: Results from the HIV Cost and Services Utilization Study. J. Stud. Alcohol 2002, 63, 179–186. [Google Scholar] [CrossRef]
- Hasin, D.S.; Stinson, F.S.; Ogburn, E.; Grant, B.F. Prevalence, correlates, disability, and comorbidity of DSM-IV alcohol abuse and dependence in the United States: Results from the National Epidemiologic Survey on Alcohol and Related Conditions. Arch. Gen. Psychiatry 2007, 64, 830–842. [Google Scholar] [CrossRef] [Green Version]
- Heaton, R.K.; Franklin, D.R.; Ellis, R.J.; McCutchan, J.A.; Letendre, S.L.; Leblanc, S.; Corkran, S.H.; Duarte, N.A.; Clifford, D.B.; Woods, S.P.; et al. HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: Differences in rates, nature, and predictors. J. NeuroVirol. 2011, 17, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.L.; Huang, W.; Mao, X.; Mack, M.L. Ethanol’s Effects on Transient Receptor Potential Channel Expression in Brain Microvascular Endothelial Cells. J. Neuroimmune Pharmacol. 2018, 13, 498–508. [Google Scholar] [CrossRef] [PubMed]
- De, S.K.; Wohlenberg, C.R.; Marinos, N.J.; Doodnauth, D.; Bryant, J.L.; Notkins, A.L. Human chorionic gonadotropin hormone prevents wasting syndrome and death in HIV-1 transgenic mice. J. Clin. Investig. 1997, 99, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- Dickie, P.; Felser, J.; Eckhaus, M.; Bryant, J.; Silver, J.; Marinos, N.; Notkins, A.L. HIV-associated nephropathy in transgenic mice expressing HIV-1 genes. Virology 1991, 185, 109–119. [Google Scholar] [CrossRef]
- Santoro, T.J.; Bryant, J.L.; Pellicoro, J.; Klotman, M.E.; Kopp, J.; Bruggeman, L.A.; Franks, R.R.; Notkins, A.L.; Klotman, P.E. Growth Failure and AIDS-like Cachexia Syndrome in HIV-1 Transgenic Mice. Virology 1994, 201, 147–151. [Google Scholar] [CrossRef]
- Altavilla, G.; Caputo, A.; Trabanelli, C.; Cofano, E.B.; Sabbioni, S.; Menegatti, M.; Barbanti-Brodano, G.; Corallini, A. Prevalence of liver tumours in HIV-1 tat-transgenic mice treated with urethane. Eur. J. Cancer 2004, 40, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Altavilla, G.; Trabanelli, C.; Merlin, M.; Caputo, A.; Lanfredi, M.; Barbanti-Brodano, G.; Corallini, A. Morphological, Histochemical, Immunohistochemical, and Ultrastructural Characterization of Tumors and Dysplastic and Non-Neoplastic Lesions Arising in BK Virus/tat Transgenic Mice. Am. J. Pathol. 1999, 154, 1231–1244. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Liu, R.-M.; Kundu, R.K.; Sangiorgi, F.; Wu, W.; Maxson, R.; Forman, H.J. Molecular Mechanism of Decreased Glutathione Content in Human Immunodeficiency Virus Type 1 Tat-transgenic Mice. J. Biol. Chem. 2000, 275, 3693–3698. [Google Scholar] [CrossRef] [Green Version]
- Corallini, A.; Altavilla, G.; Pozzi, L.; Bignozzi, F.; Negrini, M.; Rimessi, P.; Gualandi, F.; Barbanti-Brodano, G. Systemic expression of HIV-1 tat gene in transgenic mice induces endothelial proliferation and tumors of different histotypes. Cancer Res. 1993, 53, 5569–5575. [Google Scholar] [PubMed]
- De Benedictis, L.; Mariotti, M.; Dragoni, I.; Maier, J.A. Cloning and characterization of murine EDF-1. Gene 2001, 275, 299–304. [Google Scholar] [CrossRef]
- Garza, H.H.; Prakash, O.; Carr, D.J. Aberrant regulation of cytokines in HIV-1 TAT72-transgenic mice. J. Immunol. 1996, 156, 3631–3637. [Google Scholar] [CrossRef]
- Vellutini, C.; Horschowski, N.; Philippon, V.; Gambarelli, D.; Nave, K.A.; Filippi, P. Development of lymphoid hyperplasia in transgenic mice expressing the HIV tat gene. AIDS Res. Hum. Retrovir. 1995, 11, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Cepeda, M.; Enk, A.; Ngo, L.; Jay, G. The Hiv Tat Gene Is a Promoter of Epidermal Skin Tumors. Int. J. Oncol. 1995, 7, 727–733. [Google Scholar] [CrossRef]
- Toggas, S.M.; Masliah, E.; Rockenstein, E.M.; Rail, G.F.; Abraham, C.R.; Mucke, L. Central nervous system damage produced by expression of the HIV-1 coat protein gp120 in transgenic mice. Nature 1994, 367, 188–193. [Google Scholar] [CrossRef]
- Thomas, F.P.; Chalk, C.; Lalonde, R.; Robitaille, Y.; Jolicoeur, P. Expression of human immunodeficiency virus type 1 in the nervous system of transgenic mice leads to neurological disease. J. Virol. 1994, 68, 7099–7107. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.A.; Trudell, J.R.; Mihic, S.J. Ethanol’s Molecular Targets. Sci. Signal. 2008, 1, re7. [Google Scholar] [CrossRef] [Green Version]
- Haorah, J.; Schall, K.; Ramirez, S.; Persidsky, Y. Activation of protein tyrosine kinases and matrix metalloproteinases causes blood-brain barrier injury: Novel mechanism for neurodegeneration associated with alcohol abuse. Glia 2008, 56, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Haorah, J.; Knipe, B.; Leibhart, J.; Ghorpade, A.; Persidsky, Y. Alcohol-induced oxidative stress in brain endothelial cells causes blood-brain barrier dysfunction. J. Leukoc. Biol. 2005, 78, 1223–1232. [Google Scholar] [CrossRef] [Green Version]
- Howard, R.J.; Murail, S.; Ondricek, K.E.; Corringer, P.-J.; Lindahl, E.; Trudell, J.R.; Harris, R.A. Structural basis for alcohol modulation of a pentameric ligand-gated ion channel. Proc. Natl. Acad. Sci. USA 2011, 108, 12149–12154. [Google Scholar] [CrossRef] [Green Version]
- Paravicini, T.M.; Chubanov, V.; Gudermann, T. TRPM7: A unique channel involved in magnesium homeostasis. Int. J. Biochem. Cell Biol. 2012, 44, 1381–1384. [Google Scholar] [CrossRef]
- Brandao, K.; Deason-Towne, F.; Perraud, A.-L.; Schmitz, C. The role of Mg2+ in immune cells. Immunol. Res. 2013, 55, 261–269. [Google Scholar] [CrossRef]
- Guinamard, R.; Sallé, L.; Simard, C. The non-selective monovalent cationic channels TRPM4 and TRPM5. Adv. Exp. Med. Biol. 2011, 704, 147–171. [Google Scholar]
- Sarmiento, D.; Montorfano, I.; Cerda, O.; Cáceres, M.; Becerra, A.; Cabello-Verrugio, C.; Elorza, A.A.; Riedel, C.; Tapia, P.; Velásquez, L.A.; et al. Increases in reactive oxygen species enhance vascular endothelial cell migration through a mechanism dependent on the transient receptor potential melastatin 4 ion channel. Microvasc. Res. 2015, 98, 187–196. [Google Scholar] [CrossRef]
- Grimm, C.; Hassan, S.; Wahl-Schott, C.; Biel, M. Role of TRPML and Two-Pore Channels in Endolysosomal Cation Homeostasis. J. Pharmacol. Exp. Ther. 2012, 342, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Maggi, F.; Amantini, C.; Marinelli, O.; Nabissi, M.; Morelli, M.B. Pathophysiological Role of Transient Receptor Potential Mucolipin Channel 1 in Calcium-Mediated Stress-Induced Neurodegenerative Diseases. Front. Physiol. 2020, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Mackie, A.R.; Krishnamurthy, P.; Verma, S.K.; Thorne, T.; Ramirez, V.; Qin, G.; Abramova, T.; Hamada, H.; Losordo, D.; Kishore, R. Alcohol Consumption Negates Estrogen-mediated Myocardial Repair in Ovariectomized Mice by Inhibiting Endothelial Progenitor Cell Mobilization and Function. J. Biol. Chem. 2013, 288, 18022–18034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocker, M. Ca2+-activated K+ channels: Molecular determinants and function of the SK family. Nat. Rev. Neurosci. 2004, 5, 758–770. [Google Scholar] [CrossRef]
- Domen, J. The role of apoptosis in regulating hematopoietic stem cell numbers. Apoptosis 2001, 6, 239–252. [Google Scholar] [CrossRef]
- Hougaard, C.; Eriksen, B.L.; Jørgensen, S.; Johansen, T.H.; Dyhring, T.; Madsen, L.S.; Strøbaek, D.; Christophersen, P. Selective positive modulation of the SK3 and SK2 subtypes of small conductance Ca2+-activated K+ channels. Br. J. Pharmacol. 2007, 151, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Sankaranarayanan, A.; Raman, G.; Busch, C.; Schultz, T.; Zimin, P.I.; Hoyer, J.; Köhler, R.; Wulff, H. Naphtho[1,2-d]thiazol-2-ylamine (SKA-31), a New Activator of KCa2 and KCa3.1 Potassium Channels, Potentiates the Endothelium-Derived Hyperpolarizing Factor Response and Lowers Blood Pressure. Mol. Pharmacol. 2009, 75, 281–295. [Google Scholar] [CrossRef]
- Strøbæk, D.; Hougaard, C.; Johansen, T.H.; Sørensen, U.S.; Nielsen, E.Ø.; Nielsen, K.S.; Taylor, R.D.; Pedarzani, P.; Christophersen, P. Inhibitory gating modulation of small conductance Ca2+-activated K+ channels by the synthetic compound (R)-N-(benzimidazol-2-yl)-1,2,3,4-tetrahydro-1-naphtylamine (NS8593) reduces afterhyperpolarizing current in hippocampal CA1 neurons. Mol. Pharmacol. 2006, 70, 1771–1782. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Mao, X.; Chang, S.L. Altered gene expression in the spleen of adolescent rats following high ethanol concentration binge drinking. Int. J. Clin. Exp. Med. 2011, 4, 252–257. [Google Scholar]
- Takata, F.; Dohgu, S.; Yamauchi, A.; Matsumoto, J.; Machida, T.; Fujishita, K.; Shibata, K.; Shinozaki, Y.; Sato, K.; Kataoka, Y.; et al. In Vitro Blood-Brain Barrier Models Using Brain Capillary Endothelial Cells Isolated from Neonatal and Adult Rats Retain Age-Related Barrier Properties. PLoS ONE 2013, 8, e55166. [Google Scholar] [CrossRef] [Green Version]
- Molino, Y.; Jabès, F.; Lacassagne, E.; Gaudin, N.; Khrestchatisky, M. Setting-up an In Vitro Model of Rat Blood-brain Barrier (BBB): A Focus on BBB Impermeability and Receptor-mediated Transport. J. Vis. Exp. 2014, e51278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freese, C.; Reinhardt, S.; Hefner, G.; Unger, R.E.; Kirkpatrick, C.J.; Endres, K. A Novel Blood-Brain Barrier Co-Culture System for Drug Targeting of Alzheimer’s Disease: Establishment by Using Acitretin as a Model Drug. PLoS ONE 2014, 9, e91003. [Google Scholar] [CrossRef] [PubMed]
- Deli, M.A.; Ábrahám, C.S.; Kataoka, Y.; Niwa, M. Permeability studies on in vitro blood-brain barrier models: Physiology, pathology, and pharmacology. Cell. Mol. Neurobiol. 2005, 25, 59–127. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Gene symbol (RefSeq #) | Relative Expression (2−ΔΔCt) |
|---|---|---|
| TRP ankyrin subfamily | Trpa1 (NM_177781) | n.d. 1 |
| TRP canonical subfamily | Trpc1 (NM_011643) | 3.22 × 10−4 |
| Trpc2 (NM_011644) | 6.23 × 10−4 | |
| Trpc3 (NM_019510) | n.d. 1 | |
| Trpc4 (NM_016984) | n.d. 1 | |
| Trpc5 (NM_009428) | 9.35 × 10−5 | |
| Trpc6 (NM_013838) | n.d. 1 | |
| Trpc7 (NM_012035) | 9.90 × 10−5 | |
| TRP melastatin subfamily | Trpm1 (NM_001039104) | n.d. 1 |
| Trpm2 (NM_138301) | 4.36 × 10−5 | |
| Trpm3 (NM_177341) | n.d. 1 | |
| Trpm4 (NM_175130) | 3.50 × 10−3 | |
| Trpm5 (NM_020277) | 2.82 × 10−4 | |
| Trpm6 (NM_153417) | n.d. 1 | |
| Trpm7 (NM_021450) | 3.48 × 10−2 | |
| Trpm8 (NM_134252) | n.d. 1 | |
| TRP vanilloid subfamily | Trpv1 (NM_001001445) | n.d. 1 |
| Trpv2 (NM_011706) | 1.50 × 10−4 | |
| Trpv3 (NM_145099) | n.d. 1 | |
| Trpv4 (NM_022017) | 1.52 × 10−4 | |
| Trpv5 (NM_001007572) | n.d. 1 | |
| Trpv6 (NM_022413) | n.d. 1 | |
| TRP mucolipin subfamily | Trpml1 (NM_053177) | 1.89 × 10−2 |
| Trpml2 (NM_026656) | 1.14 × 10−3 | |
| Trpml3 (NM_134160) | 3.46 × 10−5 | |
| TRP polycystin subfamily | Trpp1 (NM_013628) | 1.89 × 10−2 |
| Trpp2 (NM_008861) | 1.14 × 10−3 | |
| Trpp3 (NM_181422) | 3.46 × 10−5 | |
| Trpp5 (NM_016927) | 2.77 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mack, M.L.; Huang, W.; Chang, S.L. Involvement of TRPM7 in Alcohol-Induced Damage of the Blood–Brain Barrier in the Presence of HIV Viral Proteins. Int. J. Mol. Sci. 2023, 24, 1910. https://doi.org/10.3390/ijms24031910
Mack ML, Huang W, Chang SL. Involvement of TRPM7 in Alcohol-Induced Damage of the Blood–Brain Barrier in the Presence of HIV Viral Proteins. International Journal of Molecular Sciences. 2023; 24(3):1910. https://doi.org/10.3390/ijms24031910
Chicago/Turabian StyleMack, Michelle L., Wenfei Huang, and Sulie L. Chang. 2023. "Involvement of TRPM7 in Alcohol-Induced Damage of the Blood–Brain Barrier in the Presence of HIV Viral Proteins" International Journal of Molecular Sciences 24, no. 3: 1910. https://doi.org/10.3390/ijms24031910
APA StyleMack, M. L., Huang, W., & Chang, S. L. (2023). Involvement of TRPM7 in Alcohol-Induced Damage of the Blood–Brain Barrier in the Presence of HIV Viral Proteins. International Journal of Molecular Sciences, 24(3), 1910. https://doi.org/10.3390/ijms24031910