
Targeting Agents in Biomaterial-Mediated Bone Regeneration



Abstract
:1. Introduction
2. Main Features of the Bone Microenvironment
2.1. Bone Structure
2.1.1. Organic Matrix
2.1.2. Inorganic Matrix
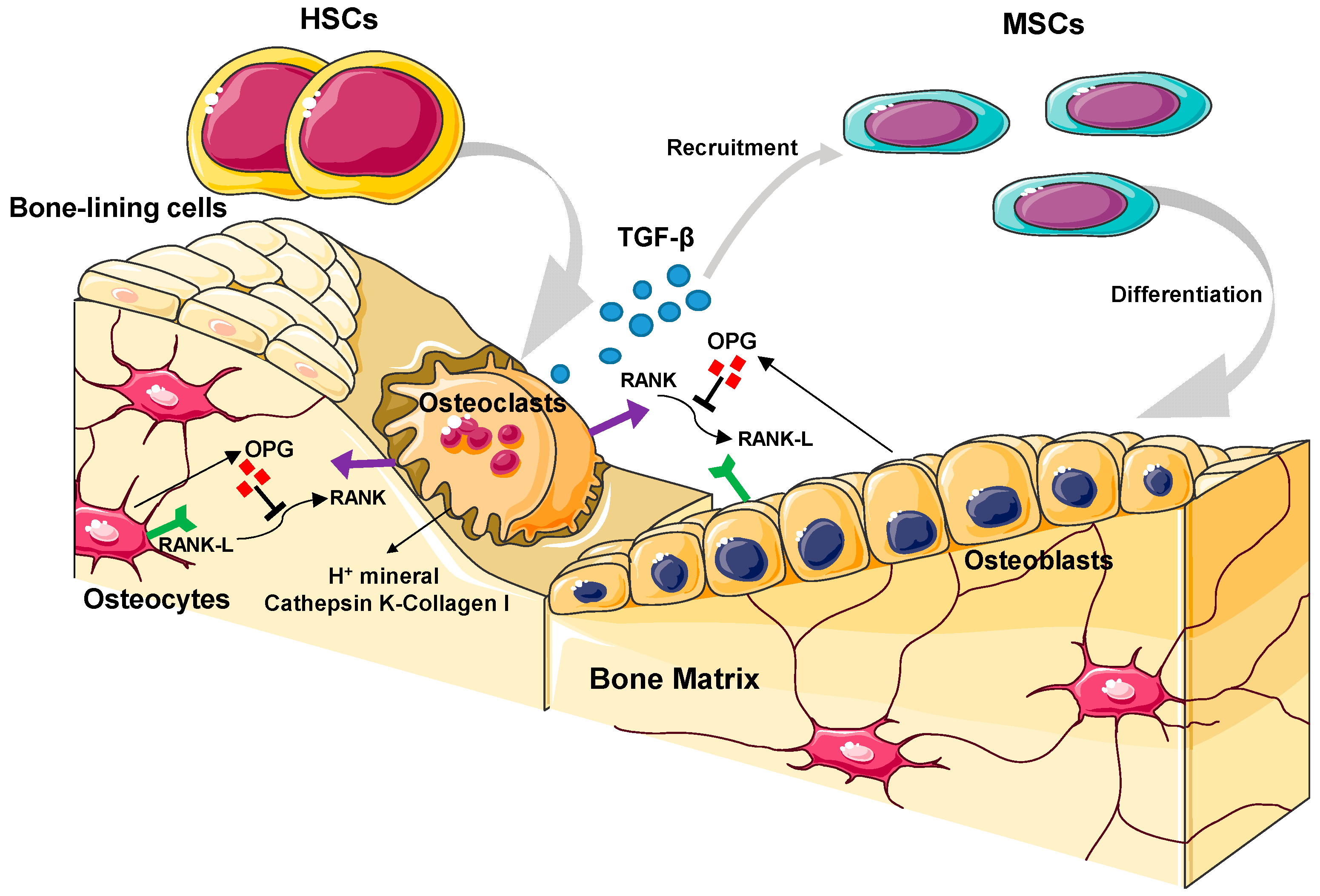
2.2. Bone Cell Biology
2.2.1. Osteoblasts
2.2.2. Osteoclasts
2.2.3. Bone-Lining Cells
2.2.4. Osteocytes
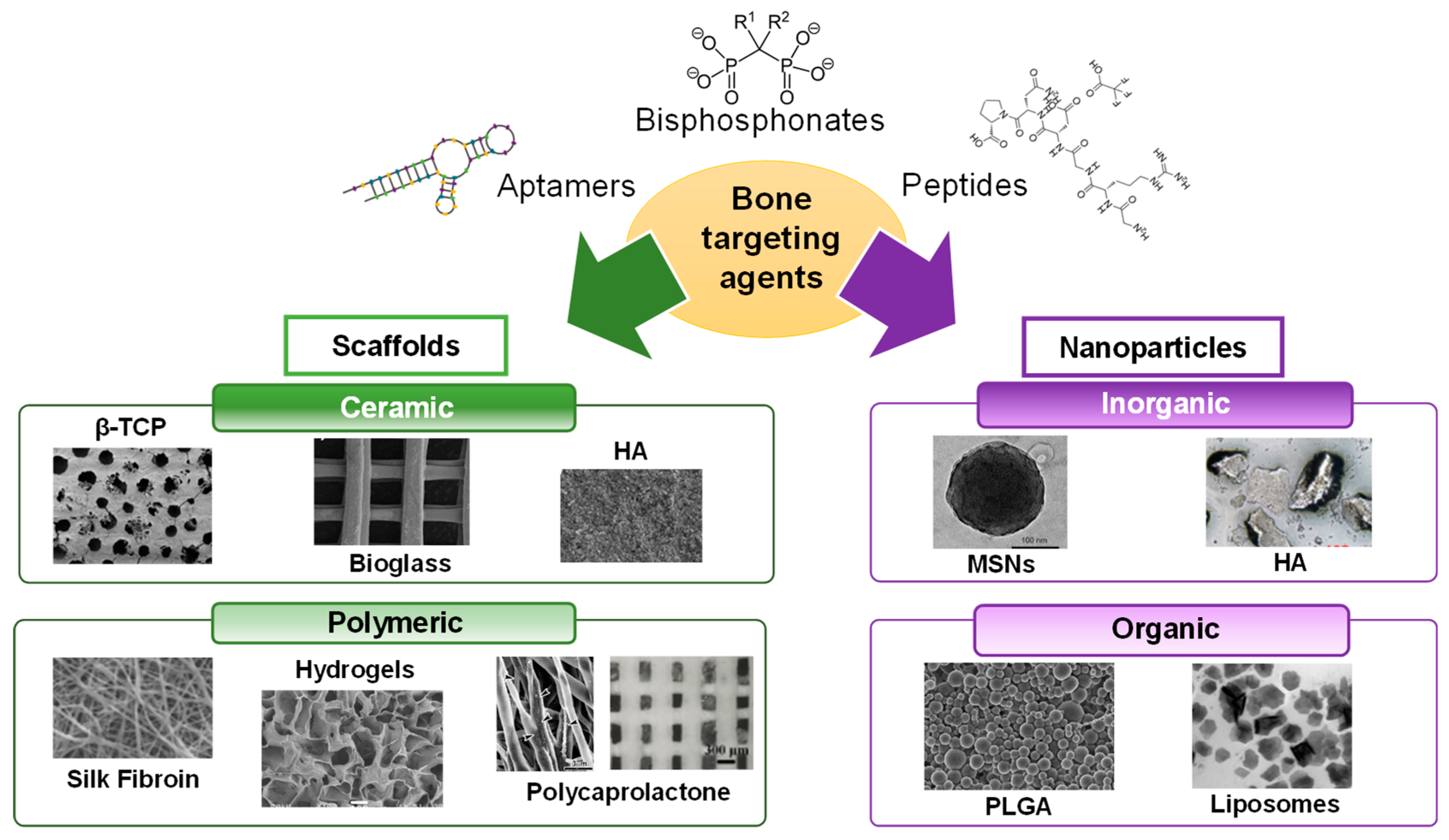
3. Macromolecules Targeting the Bone Regeneration Microenvironment
3.1. Targeting the Bone Surface through Small Molecules and Macromolecules
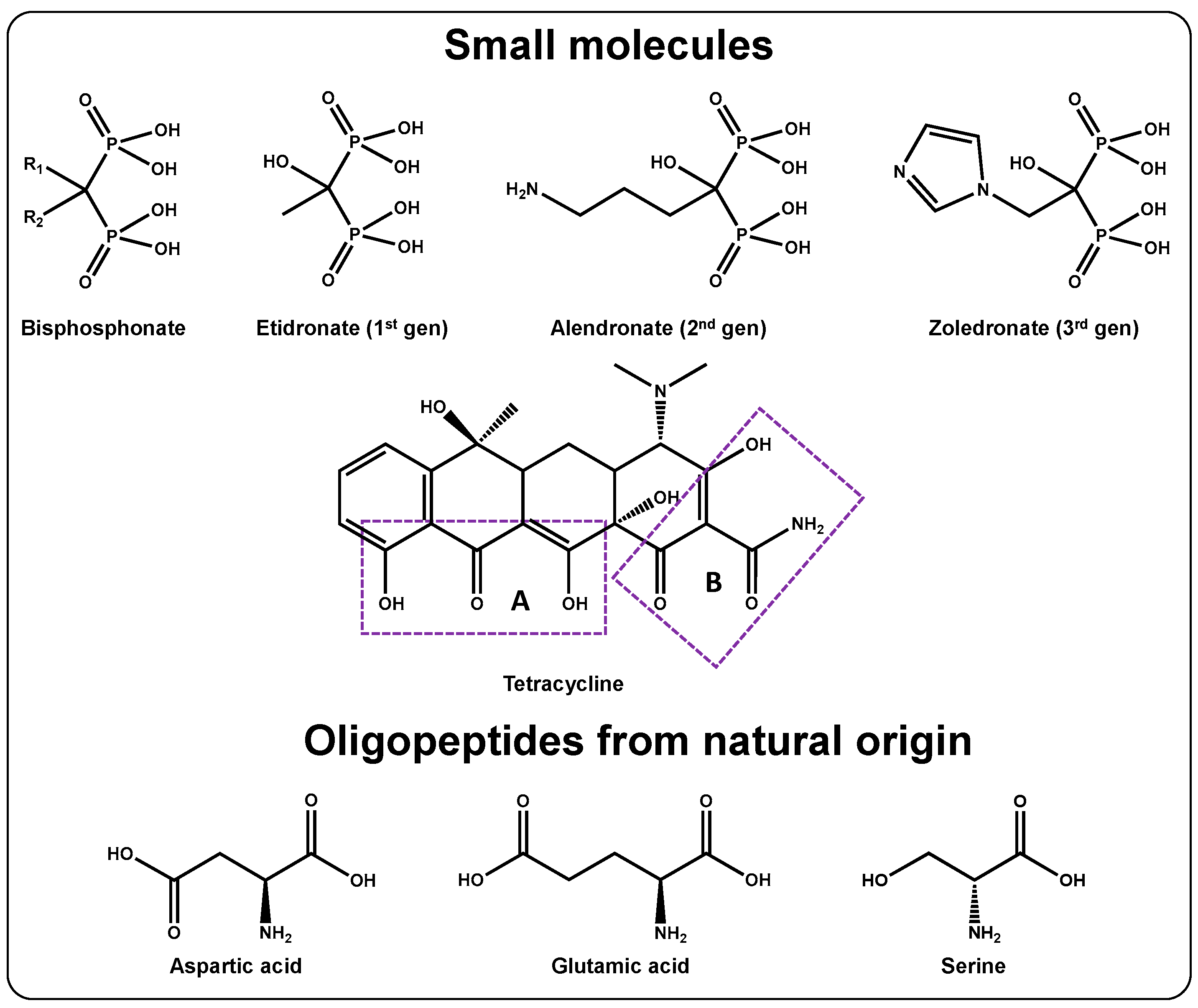
3.1.1. Bisphosphonates
3.1.2. Tetracyclines
3.1.3. Oligopeptides from Natural Origin
3.2. Generating Novel Structures to Target the Bone Microenvironment
3.2.1. Discovery of Novel Structures to Target the Bone Surface
3.2.2. Discovery of Novel Structures to Target Bone-Related Cells
- Aptamers
- Peptides
4. Targeted Nanoparticles in Bone Regeneration
4.1. Alendronate- and Tetracycline-Modified Nanocarriers in Bone Regeneration
4.2. Oligopeptide-Modified Nanocarriers in Bone Regeneration
4.3. Phage-Display-Peptide-Modified Nanocarriers in Bone Regeneration
4.4. Aptamer-Modified Nanocarriers in Bone Regeneration
5. Targeted Scaffolds in Bone Regeneration
5.1. Peptide-Modified Scaffolds in Bone Regeneration
5.2. Aptamer-Modified Scaffolds in Bone Regeneration
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Tu, K.N.; Lie, J.D.; Wan, V.; Cameron, M.; Austel, A.G.; Nguyen, J.K.; Van, K.; Hyun, D. Osteoporosis: A Review of Treatment Options. Pharm. Ther. 2018, 43, 92–104. [Google Scholar]
- Gisbert-Garzarán, M.; Manzano, M.; Vallet-Regí, M. Mesoporous Silica Nanoparticles for the Treatment of Complex Bone Diseases: Bone Cancer, Bone Infection and Osteoporosis. Pharmaceutics 2020, 12, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozen, T.; Ozisik, L.; Calik Basaran, N. An Overview and Management of Osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, N.; Hamlet, S.; Love, R.M.; Nguyen, N.T. Porous Scaffolds for Bone Regeneration. J. Sci. Adv. Mater. Devices 2020, 5, 1–9. [Google Scholar] [CrossRef]
- Albrektsson, T.; Johansson, C. Osteoinduction, Osteoconduction and Osseointegration. Eur. Spine J. 2001, 10, S96–S101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, W.; Gui, X.; Song, P.; Lei, H.; Li, Z.; Zhou, C.; Fan, Y.; Zhang, X. 3D Printing of Customized Key Biomaterials Genomics for Bone Regeneration. Appl. Mater. Today 2022, 26, 101346. [Google Scholar] [CrossRef]
- Gisbert-Garzarán, M.; Lozano, D.; Vallet-Regí, M. Mesoporous Silica Nanoparticles for Targeting Subcellular Organelles. Int. J. Mol. Sci. 2020, 21, 9696. [Google Scholar] [CrossRef]
- Peymani, A.; Shahi, M.; Sahmani, M. Regulation of Bone Metabolism. Rep. Biochem. Mol. Biol. 2017, 5, 73. [Google Scholar]
- Jakob, F.; Ebert, R.; Ignatius, A.; Matsushita, T.; Watanabe, Y.; Groll, J.; Walles, H. Bone Tissue Engineering in Osteoporosis. Maturitas 2013, 75, 118–124. [Google Scholar] [CrossRef]
- Viguet-Carrin, S.; Garnero, P.; Delmas, P.D. The Role of Collagen in Bone Strength. Osteoporos. Int. 2006, 17, 319–336. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Partridge, N.C. Cellular and Molecular Mechanisms of Bone Remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermience, M.; Lognay, G.; Goyens, P. Effects of Thirty Elements on Bone Metabolism. J. Trace Elem. Med. Biol. 2015, 32, 86–106. [Google Scholar] [CrossRef]
- O’Neill, E.; Awale, G.; Daneshmandi, L.; Umerah, O.; Lo, K.W.-H. The Roles of Ions on Bone Regeneration. Drug Discov. Today 2018, 23, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, A.; Mouriño, V.; Boccaccini, A.R. Therapeutic Inorganic Ions in Bioactive Glasses to Enhance Bone Formation and Beyond. Biomater. Sci. 2013, 1, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, C.; Zhang, X.; Chang, J.; Dai, K. Regulation of Immune Response by Bioactive Ions Released from Silicate Bioceramics for Bone Regeneration. Acta Biomater. 2018, 66, 81–92. [Google Scholar] [CrossRef]
- Zhou, R.; Guo, Q.; Xiao, Y.; Guo, Q.; Huang, Y.; Li, C.; Luo, X. Endocrine Role of Bone in the Regulation of Energy Metabolism. Bone Res. 2021, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Minamide, M.; Iwaya, C.; Ogata, K.; Iwata, J. Role of Metabolism in Bone Development and Homeostasis. Int. J. Mol. Sci. 2020, 21, 8992. [Google Scholar] [CrossRef] [PubMed]
- Desiderio, V.; Tirino, V.; Papaccio, G.; Paino, F. Bone Defects: Molecular and Cellular Therapeutic Targets. Int. J. Biochem. Cell Biol. 2014, 51, 75–78. [Google Scholar] [CrossRef]
- Sims, N.A.; Gooi, J.H. Bone Remodeling: Multiple Cellular Interactions Required for Coupling of Bone Formation and Resorption. Semin. Cell Dev. Biol. 2008, 19, 444–451. [Google Scholar] [CrossRef]
- Tamer, A.G.; Hanan, M.F. Biologics for Osteoporosis: Where Do We Stand? J. Musculoskelet Disord Treat 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Appelman-Dijkstra, N.M.; Papapoulos, S.E. Novel Approaches to the Treatment of Osteoporosis. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 843–857. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Magid, A.F. Inhibition of Cathepsin K: A Novel and Promising Treatment for Osteoporosis. ACS Med. Chem. Lett. 2015, 6, 628–629. [Google Scholar] [CrossRef] [PubMed]
- Langdahl, B.; Ferrari, S.; Dempster, D.W. Bone Modeling and Remodeling: Potential as Therapeutic Targets for the Treatment of Osteoporosis. Ther. Adv. Musculoskelet. Dis. 2016, 8, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayakumar, P.; di Silvio, L. Osteoblasts in Bone Tissue Engineering. Proc. Inst. Mech. Eng. H 2010, 224, 1415–1440. [Google Scholar] [CrossRef]
- Zhou, M.; Li, S.; Pathak, J.L. Pro-Inflammatory Cytokines and Osteocytes. Curr. Osteoporos. Rep. 2019, 17, 97–104. [Google Scholar] [CrossRef]
- Feng, X. Chemical and Biochemical Basis of Cell-Bone Matrix Interaction in Health and Disease. Curr. Chem. Biol. 2009, 3, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Reffitt, D.M.; Ogston, N.; Jugdaohsingh, R.; Cheung, H.F.J.; Evans, B.A.J.; Thompson, R.P.H.; Powell, J.J.; Hampson, G.N. Orthosilicic Acid Stimulates Collagen Type 1 Synthesis and Osteoblastic Differentiation in Human Osteoblast-like Cells in Vitro. Bone 2003, 32, 127–135. [Google Scholar] [CrossRef]
- Sun, H.; Wu, C.; Dai, K.; Chang, J.; Tang, T. Proliferation and Osteoblastic Differentiation of Human Bone Marrow-Derived Stromal Cells on Akermanite-Bioactive Ceramics. Biomaterials 2006, 27, 5651–5657. [Google Scholar] [CrossRef]
- Abbasi, N.; Ivanovski, S.; Gulati, K.; Love, R.M.; Hamlet, S. Role of Offset and Gradient Architectures of 3-D Melt Electrowritten Scaffold on Differentiation and Mineralization of Osteoblasts. Biomater. Res. 2020, 24, 1–16. [Google Scholar] [CrossRef]
- Leyva-Leyva, M.; López-Díaz, A.; Barrera, L.; Camacho-Morales, A.; Hernandez-Aguilar, F.; Carrillo-Casas, E.M.; Arriaga-Pizano, L.; Calderón-Pérez, J.; García-Álvarez, J.; Orozco-Hoyuela, G.; et al. Differential Expression of Adhesion-Related Proteins and MAPK Pathways Lead to Suitable Osteoblast Differentiation of Human Mesenchymal Stem Cells Subpopulations. Stem. Cells Dev. 2015, 24, 2577–2590. [Google Scholar] [CrossRef]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wein, M.N. Bone Lining Cells: Normal Physiology and Role in Response to Anabolic Osteoporosis Treatments. Curr. Mol. Biol. Rep. 2017, 3, 79–84. [Google Scholar] [CrossRef]
- Matic, I.; Matthews, B.G.; Wang, X.; Dyment, N.A.; Worthley, D.L.; Rowe, D.W.; Grcevic, D.; Kalajzic, I. Quiescent Bone Lining Cells Are a Major Source of Osteoblasts During Adulthood. Stem. Cells 2016, 34, 2930–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochefort, G.Y.; Pallu, S.; Benhamou, C.L. Osteocyte: The Unrecognized Side of Bone Tissue. Osteoporos. Int. 2010, 21, 1457–1469. [Google Scholar] [CrossRef]
- Birmingham, E.; Niebur, G.L.; Mchugh, P.E.; Shaw, G.; Barry, F.P.; McNamara, L.M. Osteogenic Differentiation of Mesenchymal Stem Cells Is Regulated by Osteocyte and Osteoblast Cells in a Simplified Bone Niche. Eur. Cell Mater. 2012, 23, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Kattimani, V.S.; Kondaka, S.; Lingamaneni, K.P. Hydroxyapatite–-Past, Present, and Future in Bone Regeneration. Bone Tissue Regen. Insights 2016, 7, BTRI.S36138. [Google Scholar] [CrossRef] [Green Version]
- Papapoulos, S.E. Bisphosphonate Actions: Physical Chemistry Revisited. Bone 2006, 38, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Drake, M.T.; Clarke, B.L.; Khosla, S. Bisphosphonates: Mechanism of Action and Role in Clinical Practice. Mayo Clin. Proc. 2008, 83, 1032–1045. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.G.G.; Rogers, M.J.; Frith, J.C.; Luckman, S.P.; Coxon, F.P.; Benford, H.L.; Croucher, P.I.; Shipman, C.; Fleisch, H.A. The Pharmacology of Bisphosphonates and New Insights into Their Mechanisms of Action. J. Bone Miner. Res. 1999, 14, 53–65. [Google Scholar] [CrossRef]
- Russell, R.G.G.; Watts, N.B.; Ebetino, F.H.; Rogers, M.J. Mechanisms of Action of Bisphosphonates: Similarities and Differences and Their Potential Influence on Clinical Efficacy. Osteoporos. Int. 2008, 19, 733–759. [Google Scholar] [CrossRef]
- Ian, C.; Marilyn, R. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Gomes, P.S.; Fernandes, M.H. Effect of Therapeutic Levels of Doxycycline and Minocycline in the Proliferation and Differentiation of Human Bone Marrow Osteoblastic Cells. Arch. Oral Biol. 2007, 52, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Kinugawa, S.; Koide, M.; Kobayashi, Y.; Mizoguchi, T.; Ninomiya, T.; Muto, A.; Kawahara, I.; Nakamura, M.; Yasuda, H.; Takahashi, N.; et al. Tetracyclines Convert the Osteoclastic-Differentiation Pathway of Progenitor Cells To Produce Dendritic Cell-like Cells. J. Immunol. 2012, 188, 1772–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettany, J.T.; Peet, N.M.; Wolowacz, R.G.; Skerry, T.M.; Grabowski, P.S. Tetracyclines Induce Apoptosis in Osteoclasts. Bone 2000, 27, 75–80. [Google Scholar] [CrossRef]
- Skinner, H.C.; Nalbandian, J. Tetracyclines and Mineralized Tissues: Review and Perspectives. Yale J. Biol. Med. 1975, 48, 377–397. [Google Scholar] [PubMed]
- Tao, S.; Chen, S.Q.; Zhou, W.T.; Yu, F.Y.; Bao, L.; Qiu, G.X.; Qiao, Q.; Hu, F.Q.; Wang, J.W.; Yuan, H. A Novel Biocompatible, Simvastatin-Loaded, Bone-Targeting Lipid Nanocarrier for Treating Osteoporosis More Effectively. RSC Adv. 2020, 10, 20445–20459. [Google Scholar] [CrossRef]
- Kasugai, S.; Fujisawa, R.; Waki, Y.; Miyamoto, K.-I.; Ohya, K. Selective Drug Delivery System to Bone: Small Peptide (Asp)6 Conjugation. J. Bone Miner. Res. 2000, 15, 936–943. [Google Scholar] [CrossRef]
- Sekido, T.; Sakura, N.; Higashi, Y.; Miya, K.; Nitta, Y.; Nomura, M.; Sawanishi, H.; Morito, K.; Masamune, Y.; Kasugai, S.; et al. Novel Drug Delivery System to Bone Using Acidic Oligopeptide: Pharmacokinetic Characteristics and Pharmacological Potential. J. Drug Target. 2001, 9, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Zhang, J.; Pan, J.; Yan, L.; Guo, L. Synthesis and Preliminary Evaluation in Vitro of Novel Naproxen-Dendritic Peptide Conjugates. Drug Deliv. 2009, 16, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.C.; Fu, T.F.; Wang, H.J.; Lin, C.W.; Lee, G.H.; Wu, S.C.; Wang, C.K. Aspartic Acid-Based Modified PLGA-PEG Nanoparticles for Bone Targeting: In Vitro and in Vivo Evaluation. Acta Biomater 2014, 10, 4583–4596. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Miller, S.C.; Shlyakhtenko, L.S.; Portillo, A.M.; Liu, X.M.; Papangkorn, K.; Kopečková, P.; Lyubchenko, Y.; Higuchi, W.I.; Kopeček, J. Osteotropic Peptide That Differentiates Functional Domains of the Skeleton. Bioconjugate Chem. 2007, 18, 1375–1378. [Google Scholar] [CrossRef] [PubMed]
- Yarbrough, D.K.; Hagerman, E.; Eckert, R.; He, J.; Choi, H.; Cao, N.; Le, K.; Hedger, J.; Qi, F.; Anderson, M.; et al. Specific Binding and Mineralization of Calcified Surfaces by Small Peptides. Calcif. Tissue Int. 2010, 86, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Guo, B.; Wu, H.; Tang, T.; Zhang, B.T.; Zheng, L.; He, Y.; Yang, Z.; Pan, X.; Chow, H.; et al. A Delivery System Targeting Bone Formation Surfaces to Facilitate RNAi-Based Anabolic Therapy. Nat. Med. 2012, 18, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-H.; Liu, I.-J.; Lu, R.-M.; Wu, H.-C. Advancement and Applications of Peptide Phage Display Technology in Biomedical Science. J. Biomed. Sci. 2016, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, Z.; Yu, Y.; Wang, M.; Li, J.; Zhang, Z.; Liu, J.; Wu, X.; Lu, A.; Zhang, G.; Zhang, B. Recent Advances in SELEX Technology and Aptamer Applications in Biomedicine. Int. J. Mol. Sci. 2017, 18, 2142. [Google Scholar] [CrossRef] [Green Version]
- Duffy, E.; Florek, J.; Colon, S.; Gerdon, A.E. Selected DNA Aptamers as Hydroxyapatite Affinity Reagents. Anal. Chim. Acta 2020, 1110, 115–121. [Google Scholar] [CrossRef]
- Roy, M.D.; Stanley, S.K.; Amis, E.J.; Becker, M.L. Identification of a Highly Specific Hydroxyapatite-Binding Peptide Using Phage Display. Adv. Mater. 2008, 20, 1830–1836. [Google Scholar] [CrossRef]
- Gungormus, M.; Fong, H.; Kim, I.W.; Evans, J.S.; Tamerler, C.; Sarikaya, M. Regulation of in Vitro Calcium Phosphate Mineralization by Combinatorially Selected Hydroxyapatite-Binding Peptides. Biomacromolecules 2008, 9, 966–973. [Google Scholar] [CrossRef]
- Segvich, S.; Biswas, S.; Becker, U.; Kohn, D.H. Identification of Peptides with Targeted Adhesion to Bone-like Mineral via Phage Display and Computational Modeling. Proc. Cells Tissues Organs 2008, 189, 245–251. [Google Scholar] [CrossRef]
- Addison, W.N.; Miller, S.J.; Ramaswamy, J.; Mansouri, A.; Kohn, D.H.; McKee, M.D. Phosphorylation-Dependent Mineral-Type Specificity for Apatite-Binding Peptide Sequences. Biomaterials 2010, 31, 9422–9430. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cao, B.; Modali, S.; Lee, E.M.Y.; Xu, H.; Petrenko, V.; Gray, J.J.; Yang, M.; Mao, C. Understanding the Interactions between Bone Mineral Crystals and Their Binding Peptides Derived from Filamentous Phage. Mater. Today Adv. 2022, 15, 100263. [Google Scholar] [CrossRef]
- Mao, J.; Shi, X.; Wu, Y.B.; Gong, S.Q. Identification of Specific Hydroxyapatite {001} Binding Heptapeptide by Phage Display and Its Nucleation Effect. Materials 2016, 9, 700. [Google Scholar] [CrossRef] [PubMed]
- Bang, J.; Park, H.; Yoo, J.; Lee, D.; Choi, W.; Lee, J.H.; Lee, Y.R.; Kim, C.; Koo, H.; Kim, S. Selection and Identification of a Novel Bone-Targeting Peptide for Biomedical Imaging of Bone. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.T.; Scharnweber, D.; Schwenzer, B.; Ziemer, G.; Wendel, H.P. The Effect of Electrochemical Functionalization of Ti-Alloy Surfaces by Aptamer-Based Capture Molecules on Cell Adhesion. Biomaterials 2007, 28, 468–474. [Google Scholar] [CrossRef]
- Liang, C.; Guo, B.; Wu, H.; Shao, N.; Li, D.; Liu, J.; Dang, L.; Wang, C.; Li, H.; Li, S.; et al. Aptamer-Functionalized Lipid Nanoparticles Targeting Osteoblasts as a Novel RNA Interference-Based Bone Anabolic Strategy. Nat. Med. 2015, 21, 288–294. [Google Scholar] [CrossRef]
- Ren, M.; Li, Y.; Zhang, H.; Li, L.; He, P.; Ji, P.; Yang, S. An Oligopeptide/Aptamer-Conjugated Dendrimer-Based Nanocarrier for Dual-Targeting Delivery to Bone. J. Mater. Chem. B 2021, 9, 2831–2844. [Google Scholar] [CrossRef]
- Huang, S.-S.; Lee, K.-J.; Chen, H.-C.; Putra Prajnamitra, R.; Hsu, C.-H.; Jian, C.-B.; Yu, X.-E.; Chueh, D.-Y.; Wen Kuo, C.; Chiang, T.-C.; et al. Immune cell shuttle for precise delivery of nanotherapeutics for heart disease and cancer. Sci. Adv. 2021, 7, eabf2400. [Google Scholar] [CrossRef]
- Li, C.-J.; Cheng, P.; Liang, M.-K.; Chen, Y.-S.; Lu, Q.; Wang, J.-Y.; Xia, Z.-Y.; Zhou, H.-D.; Cao, X.; Xie, H.; et al. MicroRNA-188 Regulates Age-Related Switch between osteoblast and Adipocyte Differentiation. J. Clin. Investig. 2015, 125, 1509–1522. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Meyer, S.; Propson, N.E.; Nie, J.; Jiang, P.; Stewart, R.; Thomson, J.A. Characterization and Target Identification of a DNA Aptamer That Labels Pluripotent Stem Cells. Cell Res. 2015, 25, 390–393. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wu, H.; Li, Q.; Yang, Y.; Che, F.; Wang, G.; Zhang, L. Novel Aptamer-Functionalized Nanoparticles Enhances Bone Defect Repair by Improving Stem Cell Recruitment. Int. J. Nanomed. 2019, 14, 8707–8724. [Google Scholar] [CrossRef] [Green Version]
- Ardjomandi, N.; Niederlaender, J.; Aicher, W.K.; Reinert, S.; Schweizer, E.; Wendel, H.P.; Alexander, D. Identification of an Aptamer Binding to Human Osteogenic-Induced Progenitor Cells. Nucleic Acid Ther. 2013, 23, 44–61. [Google Scholar] [CrossRef] [Green Version]
- Sheu, T.J.; Schwarz, E.M.; O’Keefe, R.J.; Rosier, R.N.; Puzas, J.E. Use of a Phage Display Technique to Identify Potential Osteoblast Binding Sites within Osteoclast Lacunae. J. Bone Miner. Res. 2002, 17, 915–922. [Google Scholar] [CrossRef] [PubMed]
- AC’T Hoen, P.; Jirka, S.M.G.; ten Broeke, B.R.; Schultes, E.A.; Aguilera, B.; Pang, K.H.; Heemskerk, H.; Aartsma-Rus, A.; van Ommen, G.J.; den Dunnen, J.T. Phage Display Screening without Repetitious Selection Rounds. Anal. Biochem. 2012, 421, 622–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Ye, X.; Cai, M.; Liu, X.; Xiao, J.; Zhang, C.; Wang, Y.; Yang, L.; Liu, J.; Li, S.; et al. Osteoblast-Targeting-Peptide Modified Nanoparticle for SiRNA/MicroRNA Delivery. ACS Nano 2016, 10, 5759–5768. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Zhang, X.; Pi, Y.; Wang, X.; Jia, Z.; Zhu, J.; Dai, L.; Chen, W.; Yin, L.; Chen, H.; et al. Polycaprolactone Electrospun Mesh Conjugated with an MSC Affinity Peptide for MSC Homing in Vivo. Biomaterials 2012, 33, 3375–3387. [Google Scholar] [CrossRef]
- Ramaraju, H.; Miller, S.J.; Kohn, D.H. Dual-Functioning Peptides Discovered by Phage Display Increase the Magnitude and Specificity of BMSC Attachment to Mineralized Biomaterials. Biomaterials 2017, 134, 1–12. [Google Scholar] [CrossRef]
- Nowakowski, G.S.; Dooner, M.S.; Valinski, H.M.; Mihaliak, A.M.; Quesenberry, P.J.; Becker, P.S. A Specific Heptapeptide from a Phage Display Peptide Library Homes to Bone Marrow and Binds to Primitive Hematopoietic Stem Cells. Stem. Cells 2004, 22, 1030–1038. [Google Scholar] [CrossRef]
- Wang, G.; Man, Z.; Zhang, N.; Xin, H.; Li, Y.; Sun, T.; Sun, S. Biopanning of Mouse Bone Marrow Mesenchymal Stem Cell Affinity for Cyclic Peptides. Mol. Med. Rep. 2019, 19, 407–413. [Google Scholar] [CrossRef]
- Sun, T.; Man, Z.; Peng, C.; Wang, G.; Sun, S. A Specific Affinity Cyclic Peptide Enhances the Adhesion, Expansion and Proliferation of Rat Bone Mesenchymal Stem Cells on Β-tricalcium Phosphate Scaffolds. Mol. Med. Rep. 2019, 20, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zheng, C.; Li, Y.; Bian, S.; Pan, H.; Zhao, X.; Lu, W.W. Bone Targeted Delivery of SDF-1 via Alendronate Functionalized Nanoparticles in Guiding Stem Cell Migration. ACS Appl. Mater. Interfaces 2018, 10, 23700–23710. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, Y.; Shi, C.; Wu, T.; Cui, Y.; Wang, S.; Liu, P.; Feng, X.; He, Y.; Fu, D. Remote-Controllable Bone-Targeted Delivery of Estradiol for the Treatment of Ovariectomy-Induced Osteoporosis in Rats. J. Nanobiotechnology 2021, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cornel, E.J.; Fan, Z.; He, S.; Du, J. Bone-Targeting Polymer Vesicles for Effective Therapy of Osteoporosis. Nano Lett. 2021, 21, 7998–8007. [Google Scholar] [CrossRef] [PubMed]
- Mora-Raimundo, P.; Lozano, D.; Benito, M.; Mulero, F.; Manzano, M.; Vallet-Regí, M. Osteoporosis Remission and New Bone Formation with Mesoporous Silica Nanoparticles. Adv. Sci. 2021, 8, 2101107. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, C.; Huang, H.; Huang, J.; Deng, A.; Zou, P.; Tan, X. Bone-Targeted Delivery of Simvastatin-Loaded PEG-PLGA Micelles Conjugated with Tetracycline for Osteoporosis Treatment. Drug Deliv. Transl. Res. 2018, 8, 1090–1102. [Google Scholar] [CrossRef]
- Tao, S.; Yu, F.; Song, Y.; Zhou, W.; Lv, J.; Zhao, R.; Wang, C.; Hu, F.; Yuan, H. Water/PH Dual Responsive in Situ Calcium Supplement Collaborates Simvastatin for Osteoblast Promotion Mediated Osteoporosis Therapy via Oral Medication. J. Control. Release 2021, 329, 121–135. [Google Scholar] [CrossRef]
- Lin, C.W.; Lee, C.Y.; Lin, S.Y.; Kang, L.; Fu, Y.C.; Chen, C.H.; Wang, C.K. Bone-Targeting Nanoparticles of a Dendritic (Aspartic Acid)3-Functionalized PEG-PLGA Biopolymer Encapsulating Simvastatin for the Treatment of Osteoporosis in Rat Models. Int. J. Mol. Sci. 2022, 23, 10530. [Google Scholar] [CrossRef]
- Huang, L.; Wang, X.; Cao, H.; Li, L.; Chow, D.H.K.; Tian, L.; Wu, H.; Zhang, J.; Wang, N.; Zheng, L.; et al. A Bone-Targeting Delivery System Carrying Osteogenic Phytomolecule Icaritin Prevents Osteoporosis in Mice. Biomaterials 2018, 182, 58–71. [Google Scholar] [CrossRef]
- Gao, Y.; Xin, H.; Cai, B.; Wang, L.; Lv, Q.; Hou, Y.; Liu, F.; Dai, T.; Kong, L. RNA Interference-Based Osteoanabolic Therapy for Osteoporosis by a Bone-Formation Surface Targeting Delivery System. Biochem. Biophys. Res. Commun. 2022, 601, 86–92. [Google Scholar] [CrossRef]
- Sui, B.; Liu, J.; Zheng, C.; Dang, L.; Chen, J.; Cao, Y.; Zhang, K.; Liu, L.; Dang, M.; Zhang, L.; et al. Targeted Inhibition of Osteoclastogenesis Reveals the Pathogenesis and Therapeutics of Bone Loss under Sympathetic Neurostress. Int. J. Oral. Sci. 2022, 14, 1–10. [Google Scholar] [CrossRef]
- Cai, M.; Yang, L.; Zhang, S.; Liu, J.; Sun, Y.; Wang, X. A Bone-Resorption Surface-Targeting Nanoparticle to Deliver Anti-MiR214 for Osteoporosis Therapy. Int. J. Nanomed. 2017, 12, 7469–7482. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hu, Z.; Shi, F.; Dong, J.; Dang, L.; Wang, Y.; Sun, Z.; Zhou, H.; Zhang, S.; Cao, X.; et al. Osteoblast-Targeted Delivery of MiR-33-5p Attenuates Osteopenia Development Induced by Mechanical Unloading in Mice. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Miron, R.J.; Bian, Z.; Zhang, Y.F. A Bone-Targeting Drug-Delivery System Based on Semaphorin 3A Gene Therapy Ameliorates Bone Loss in Osteoporotic Ovariectomized Mice. Bone 2018, 114, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Xie, J.; Wang, D.; Kim, J.M.; Tai, P.W.L.; Gravallese, E.; Gao, G.; Shim, J.H. Bone-Targeting AAV-Mediated Silencing of Schnurri-3 Prevents Bone Loss in Osteoporosis. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.S.; Xie, J.; Chaugule, S.; Wang, D.; Kim, J.M.; Kim, J.H.; Tai, P.W.L.; Seo, S.; Gravallese, E.; Gao, G.; et al. Bone-Targeting AAV-Mediated Gene Silencing in Osteoclasts for Osteoporosis Therapy. Mol. Ther.-Methods Clin. Dev. 2020, 17, 922–935. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Que, K.; Wang, H.; An, R.; Chen, Z.; Qiu, Z.; Lin, M.; Song, J.; Yang, J.; Lu, D.; et al. Rapid Biomimetic Remineralization of the Demineralized Enamel Surface Using Nano-Particles of Amorphous Calcium Phosphate Guided by Chimaeric Peptides. Dent. Mater. 2017, 33, 1217–1228. [Google Scholar] [CrossRef]
- Cui, Y.; Guo, Y.; Kong, L.; Shi, J.; Liu, P.; Li, R.; Geng, Y.; Gao, W.; Zhang, Z.; Fu, D. A Bone-Targeted Engineered Exosome Platform Delivering SiRNA to Treat Osteoporosis. Bioact. Mater. 2022, 10, 207–221. [Google Scholar] [CrossRef]
- Wang, Y.; Newman, M.R.; Ackun-Farmmer, M.; Baranello, M.P.; Sheu, T.J.; Puzas, J.E.; Benoit, D.S.W. Fracture-Targeted Delivery of β-Catenin Agonists via Peptide-Functionalized Nanoparticles Augments Fracture Healing. ACS Nano 2017, 11, 9445–9458. [Google Scholar] [CrossRef] [Green Version]
- García-García, P.; Reyes, R.; García-Sánchez, D.; Pérez-Campo, F.M.; Rodríguez-Rey, J.C.; Évora, C.; Díaz-Rodríguez, P.; Delgado, A. Nanoparticle-Mediated Selective Sfrp-1 Silencing Enhances Bone Density in Osteoporotic Mice. J. Nanobiotechnol. 2022, 20, 1–19. [Google Scholar] [CrossRef]
- Luo, Z.W.; Li, F.X.Z.; Liu, Y.W.; Rao, S.S.; Yin, H.; Huang, J.; Chen, C.Y.; Hu, Y.; Zhang, Y.; Tan, Y.J.; et al. Aptamer-Functionalized Exosomes from Bone Marrow Stromal Cells Target Bone to Promote Bone Regeneration. Nanoscale 2019, 11, 20884–20892. [Google Scholar] [CrossRef]
- Gómez-Cerezo, N.; Arcos, D.; Vallet-Regí, M. Mesoporous Bioactive Glasses for Biomedical Composites. In Materials for Biomedical Engineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 355–391. [Google Scholar] [CrossRef]
- Casarrubios, L.; Gómez-Cerezo, N.; Sánchez-Salcedo, S.; Feito, M.J.; Serrano, M.C.; Saiz-Pardo, M.; Ortega, L.; de Pablo, D.; Díaz-Güemes, I.; Fernández-Tomé, B.; et al. Silicon Substituted Hydroxyapatite/VEGF Scaffolds Stimulate Bone Regeneration in Osteoporotic Sheep. Acta Biomater 2020, 101, 544–553. [Google Scholar] [CrossRef]
- Hamlet, S.M.; Vaquette, C.; Shah, A.; Hutmacher, D.W.; Ivanovski, S. 3-Dimensional Functionalized Polycaprolactone-Hyaluronic Acid Hydrogel Constructs for Bone Tissue Engineering. J. Clin. Periodontol. 2017, 44, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Sánchez, M.; Escobar-Barrios, V.A.; Pozos-Guillén, A.; Escobar-García, D.M. RGD-Functionalization of PLA/Starch Scaffolds Obtained by Electrospinning and Evaluated in Vitro for Potential Bone Regeneration. Mater. Sci. Eng. C 2019, 96, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.R.; Serra, T.; Oliveira, M.I.; Planell, J.A.; Barbosa, M.A.; Navarro, M. Impact of 3-D Printed PLA- and Chitosan-Based Scaffolds on Human Monocyte/Macrophage Responses: Unraveling the Effect of 3-D Structures on Inflammation. Acta Biomater 2014, 10, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Miszuk, J.M.; Xu, T.; Yao, Q.; Fang, F.; Childs, J.D.; Hong, Z.; Tao, J.; Fong, H.; Sun, H. Functionalization of PCL-3D Electrospun Nanofibrous Scaffolds for Improved BMP2-Induced Bone Formation. Appl. Mater. Today 2018, 10, 194–202. [Google Scholar] [CrossRef]
- Siddiqui, N.; Asawa, S.; Birru, B.; Baadhe, R.; Rao, S. PCL-Based Composite Scaffold Matrices for Tissue Engineering Applications. Mol. Biotechnol. 2018, 60, 506–532. [Google Scholar] [CrossRef]
- Gómez-Cerezo, M.N.; Peña, J.; Ivanovski, S.; Arcos, D.; Vallet-Regí, M.; Vaquette, C. Multiscale Porosity in Mesoporous Bioglass 3D-Printed Scaffolds for Bone Regeneration. Mater. Sci. Eng. C 2020, 120, 111706. [Google Scholar] [CrossRef]
- Liu, X.; Ma, P.X. Phase Separation, Pore Structure, and Properties of Nanofibrous Gelatin Scaffolds. Biomaterials 2009, 30, 4094–4103. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vázquez, F.J.; Cabañas, M.V.; Paris, J.L.; Lozano, D.; Vallet-Regí, M. Fabrication of Novel Si-Doped Hydroxyapatite/Gelatine Scaffolds by Rapid Prototyping for Drug Delivery and Bone Regeneration. Acta Biomater. 2015, 15, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Velasco, D.; Benito, L.; Fernández-Gutiérrez, M.; San Román, J.; Elvira, C. Preparation in Supercritical CO2 of Porous Poly(Methyl Methacrylate)–Poly(l-Lactic Acid) (PMMA–PLA) Scaffolds Incorporating Ibuprofen. J. Supercrit. Fluids 2010, 54, 335–341. [Google Scholar] [CrossRef]
- Gorna, K.; Gogolewski, S. Preparation, Degradation, and Calcification of Biodegradable Polyurethane Foams for Bone Graft Substitutes. J. Biomed. Mater. Res. 2003, 67A, 813–827. [Google Scholar] [CrossRef]
- Zhai, P.; Peng, X.; Li, B.; Liu, Y.; Sun, H.; Li, X. The Application of Hyaluronic Acid in Bone Regeneration. Int. J. Biol. Macromol. 2020, 151, 1224–1239. [Google Scholar] [CrossRef] [PubMed]
- García-Alvarez, R.; Izquierdo-Barba, I.; Vallet-Regí, M. 3D Scaffold with Effective Multidrug Sequential Release against Bacteria Biofilm. Acta Biomater 2017, 49, 113–126. [Google Scholar] [CrossRef]
- Prokopowicz, M.; Szewczyk, A.; Skwira, A.; Sądej, R.; Walker, G. Biphasic Composite of Calcium Phosphate-Based Mesoporous Silica as a Novel Bone Drug Delivery System. Drug Deliv. Transl. Res. 2019, 10, 455–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallet-Regí, M.; González-Calbet, J.M. Calcium Phosphates as Substitution of Bone Tissues. Prog. Solid State Chem. 2004, 32, 1–31. [Google Scholar] [CrossRef]
- Yeo, A.; Wong, W.J.; Teoh, S.H. Surface Modification of PCL-TCP Scaffolds in Rabbit Calvaria Defects: Evaluation of Scaffold Degradation Profile, Biomechanical Properties and Bone Healing Patterns. J. Biomed. Mater. Res. A 2010, 93, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-W.; Knowles, J.C.; Kim, H.-E. Hydroxyapatite/Poly(ε-Caprolactone) Composite Coatings on Hydroxyapatite Porous Bone Scaffold for Drug Delivery. Biomaterials 2004, 25, 1279–1287. [Google Scholar] [CrossRef]
- Gómez-Cerezo, M.N.; Lozano, D.; Arcos, D.; Vallet-Regí, M.; Vaquette, C. The Effect of Biomimetic Mineralization of 3D-Printed Mesoporous Bioglass Scaffolds on Physical Properties and in Vitro Osteogenicity. Mater. Sci. Eng. C 2020, 109, 110572. [Google Scholar] [CrossRef]
- Wu, C.; Xiao, Y. Mesoporous Bioglass/Silk Composite Scaffolds for Bone Tissue Engineering. Biomaterial 2011, 2, 269–286. [Google Scholar]
- Xynos, I.D.; Hukkanen, M.V.J.; Batten, J.J.; Buttery, L.D.; Hench, L.L.; Polak, J.M. Bioglass ®45S5 Stimulates Osteoblast Turnover and Enhances Bone Formation In Vitro: Implications and Applications for Bone Tissue Engineering. Calcif. Tissue Int. 2000, 67, 321–329. [Google Scholar] [CrossRef]
- Miri, A.K.; Muja, N.; Kamranpour, N.O.; Lepry, W.C.; Boccaccini, A.R.; Clarke, S.A.; Nazhat, S.N. Ectopic Bone Formation in Rapidly Fabricated Acellular Injectable Dense Collagen-Bioglass Hybrid Scaffolds via Gel Aspiration-Ejection. Biomaterials 2016, 85, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Xiao, Y.; Chang, J. Silicate—Based Bioactive Ceramics for Bone Regeneration Application. In Advanced Bioactive Inorganic Materials for Bone Regeneration and Drug Delivery; CRC Press: Boca Raton, FL, USA, 2013; pp. 25–46. [Google Scholar]
- Garot, C.; Bettega, G.; Picart, C. Additive Manufacturing of Material Scaffolds for Bone Regeneration: Toward Application in the Clinics. Adv. Funct. Mater. 2020, 31, 2006967. [Google Scholar] [CrossRef]
- Chang, J.; Zhang, X.; Dai, K. Bioactive Materials for Bone Regeneration; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Cui, Y.; Zhu, T.; Li, D.; Li, Z.; Leng, Y.; Ji, X.; Liu, H.; Wu, D.; Ding, J. Bisphosphonate-Functionalized Scaffolds for Enhanced Bone Regeneration. Adv. Healthc. Mater. 2019, 8, 1901073. [Google Scholar] [CrossRef]
- Barik, D.; Dash, P.; Uma, P.I.; Kumari, S.; Dash, M. A Review on Re-Packaging of Bisphosphonates Using Biomaterials. J. Pharm. Sci. 2021, 110, 3757–3772. [Google Scholar] [CrossRef]
- Dettin, M.; Zamuner, A.; Roso, M.; Iucci, G.; Samouillan, V.; Danesin, R.; Modesti, M.; Conconi, M.T. Facile and Selective Covalent Grafting of an RGD-Peptide to Electrospun Scaffolds Improves HUVEC Adhesion. J. Pept. Sci. 2015, 21, 786–795. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Barreto, J.F.; Sikavitsas, V.I. Improved Mesenchymal Stem Cell Seeding on RGD-Modified Poly(L-Lactic Acid) Scaffolds Using Flow Perfusion. Macromol. Biosci. 2007, 7, 579–588. [Google Scholar] [CrossRef]
- Marletta, G.; Ciapetti, G.; Satriano, C.; Pagani, S.; Baldini, N. The Effect of Irradiation Modification and RGD Sequence Adsorption on the Response of Human Osteoblasts to Polycaprolactone. Biomaterials 2005, 26, 4793–4804. [Google Scholar] [CrossRef] [PubMed]
- Tahlawi, A.; Klontzas, M.E.; Allenby, M.C.; Morais, J.C.F.; Panoskaltsis, N.; Mantalaris, A. RGD-Functionalized Polyurethane Scaffolds Promote Umbilical Cord Blood Mesenchymal Stem Cell Expansion and Osteogenic Differentiation. J. Tissue Eng. Regen. Med. 2019, 13, 232–243. [Google Scholar] [CrossRef] [PubMed]
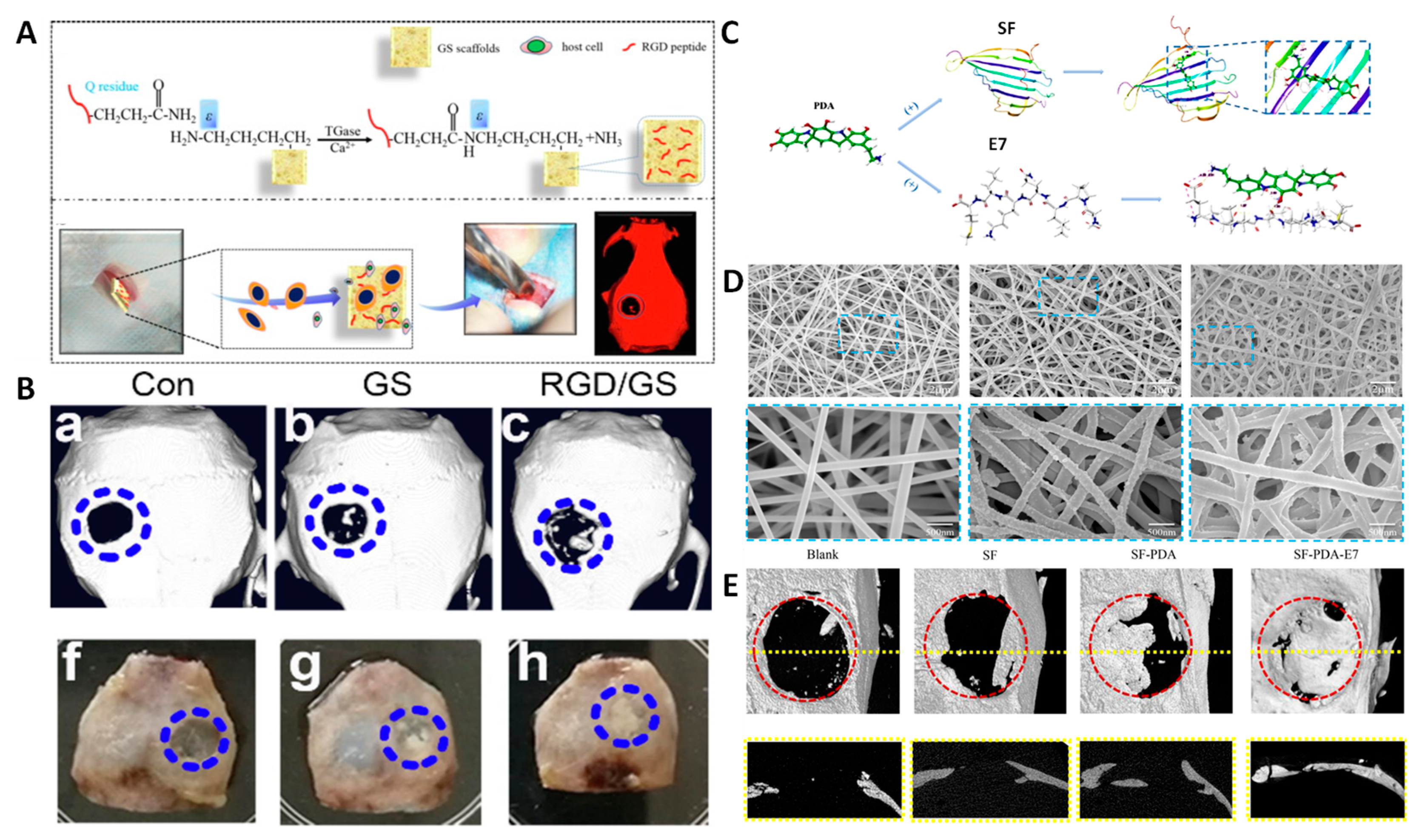
- Li, J.; Zhang, Y.; Zhou, X.; Wang, S.; Hao, R.; Han, J.; Li, M.; Zhao, Y.; Chen, C.; Xu, H. Enzymatically Functionalized RGD-Gelatin Scaffolds That Recruit Host Mesenchymal Stem Cells in Vivo and Promote Bone Regeneration. J. Colloid Interface Sci. 2022, 612, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Salifu, A.A.; Obayemi, J.D.; Uzonwanne, V.O.; Soboyejo, W.O. Mechanical Stimulation Improves Osteogenesis and the Mechanical Properties of Osteoblast-Laden RGD-Functionalized Polycaprolactone/Hydroxyapatite Scaffolds. J. Biomed Mater. Res. A 2020, 108, 2421–2434. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Liu, M.; Xu, T.; Wang, K.; Tan, H.; Lu, X. Chitosan/Biphasic Calcium Phosphate Scaffolds Functionalized with BMP-2-Encapsulated Nanoparticles and RGD for Bone Regeneration. J. Biomed Mater. Res. A 2018, 106, 2613–2624. [Google Scholar] [CrossRef]
- Roy, T.; James, B.D.; Allen, J.B.; Roy, T.; James, B.D.; Allen, J.B. Anti-VEGF-R2 Aptamer and RGD Peptide Synergize in a Bifunctional Hydrogel for Enhanced Angiogenic Potential. Macromol. Biosci. 2021, 21, 2000337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ling, C.; Liu, H.; Zhang, A.; Mao, L.; Wang, J.; Chao, J.; Backman, L.J.; Yao, Q.; Chen, J. Tannic Acid-Mediated Dual Peptide-Functionalized Scaffolds to Direct Stem Cell Behavior and Osteochondral Regeneration. Chem. Eng. J. 2020, 396, 125232. [Google Scholar] [CrossRef]
- Wu, J.; Cao, L.; Liu, Y.; Zheng, A.; Jiao, D.; Zeng, D.; Wang, X.; Kaplan, D.L.; Jiang, X. Functionalization of Silk Fibroin Electrospun Scaffolds via BMSC Affinity Peptide Grafting through Oxidative Self-Polymerization of Dopamine for Bone Regeneration. ACS Appl. Mater. Interfaces 2019, 11, 8878–8895. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fang, J.; Fu, L.; Chen, L.; Dai, W.; Huang, H.; Wang, J.; Zhang, X.; Cai, Q.; Yang, X. Gradient Fibrous Aerogel Conjugated with Chemokine Peptide for Regulating Cell Differentiation and Facilitating Osteochondral Regeneration. Chem. Eng. J. 2021, 422, 130428. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, Y.; Huang, L.; Gao, F.; Sun, H.; Huang, C. Intramembranous Ossification Imitation Scaffold with the Function of Macrophage Polarization for Promoting Critical Bone Defect Repair. ACS Appl. Bio. Mater. 2020, 3, 3569–3581. [Google Scholar] [CrossRef]
- Wang, G.; Xin, H.; Tian, G.; Sheng, K.; Zhang, N.; Sun, S. Core Decompression Combined with Implantation of β-Tricalcium Phosphate Modified by a BMSC Affinity Cyclic Peptide for the Treatment of Early Osteonecrosis of the Femoral Head. Am. J. Transl. Res. 2021, 13, 967. [Google Scholar]
- Lu, J.; Shen, X.; Sun, X.; Yin, H.; Yang, S.; Lu, C.; Wang, Y.; Liu, Y.; Huang, Y.; Yang, Z.; et al. Increased Recruitment of Endogenous Stem Cells and Chondrogenic Differentiation by a Composite Scaffold Containing Bone Marrow Homing Peptide for Cartilage Regeneration. Theranostics 2018, 8, 5039–5058. [Google Scholar] [CrossRef]
- Huang, B.; Li, P.; Chen, M.; Peng, L.; Luo, X.; Tian, G.; Wang, H.; Wu, L.; Tian, Q.; Li, H.; et al. Hydrogel Composite Scaffolds Achieve Recruitment and Chondrogenesis in Cartilage Tissue Engineering Applications. J. Nanobiotechnology 2022, 20, 1–17. [Google Scholar] [CrossRef]
- Wang, G.; Man, Z.; Xin, H.; Li, Y.; Wu, C.; Sun, S. Enhanced Adhesion and Proliferation of Bone Marrow Mesenchymal Stem Cells on β-Tricalcium Phosphate Modified by an Affinity Peptide. Mol. Med. Rep. 2019, 19, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, Y.; Sun, T.; Wang, C.; Qiao, L.; Wang, Y.; Dong, K.; Yuan, T.; Chen, J.; Chen, G.; et al. BMSC Affinity Peptide-Functionalized β-Tricalcium Phosphate Scaffolds Promoting Repair of Osteonecrosis of the Femoral Head. J. Orthop. Surg. Res. 2019, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ramaraju, H.; Kohn, D.H. Cell and Material-Specific Phage Display Peptides Increase IPS-MSC Mediated Bone and Vasculature Formation In Vivo. Adv. Healthc. Mater. 2019, 8, 1801356. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, X.; Dai, J.; Wang, J.; Tan, Y.; Yang, X.; Yang, S.; Yuan, Q.; Zhang, Y. A 3D Graphene Coated Bioglass Scaffold for Bone Defect Therapy Based on the Molecular Targeting Approach. J. Mater. Chem. B 2017, 5, 6794–6800. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhao, T.; Gao, C.; Cao, F.; Li, H.; Liao, Z.; Fu, L.; Li, P.; Chen, W.; Sun, Z.; et al. 3D-Bioprinted Difunctional Scaffold for in Situ Cartilage Regeneration Based on Aptamer-Directed Cell Recruitment and Growth Factor-Enhanced Cell Chondrogenesis. ACS Appl. Mater. Interfaces 2021, 13, 23369–23383. [Google Scholar] [CrossRef] [PubMed]
- Ardjomandi, N.; Huth, J.; Stamov, D.R.; Henrich, A.; Klein, C.; Wendel, H.P.; Reinert, S.; Alexander, D. Surface Biofunctionalization of β-TCP Blocks Using Aptamer 74 for Bone Tissue Engineering. Mater. Sci. Eng. C 2016, 67, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Chen, M.; Li, M.; Wang, D.; Cai, K.; Luo, Z.; Hu, Y. Aptamer/Hydroxyapatite-Functionalized Titanium Substrate Promotes Implant Osseointegration via Recruiting Mesenchymal Stem Cells. ACS Appl Mater Interfaces 2022, 14, 42915–42930. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, Y.; Tan, Y.; Wang, J.; Liu, H.; Wang, Y.; Yang, S.; Shi, M.; Zhao, S.; Zhang, Y.; et al. A Difunctional Regeneration Scaffold for Knee Repair Based on Aptamer-Directed Cell Recruitment. Adv. Mater. 2017, 29, 1605235. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Li, T.; Chen, J.; Cheng, G.; Yang, L.; Chen, C. Aptamer-Functionalized Bioscaffold Enhances Cartilage Repair by Improving Stem Cell Recruitment in Osteochondral Defects of Rabbit Knees. Am. J. Sport. Med. 2019, 47, 2316–2326. [Google Scholar] [CrossRef]
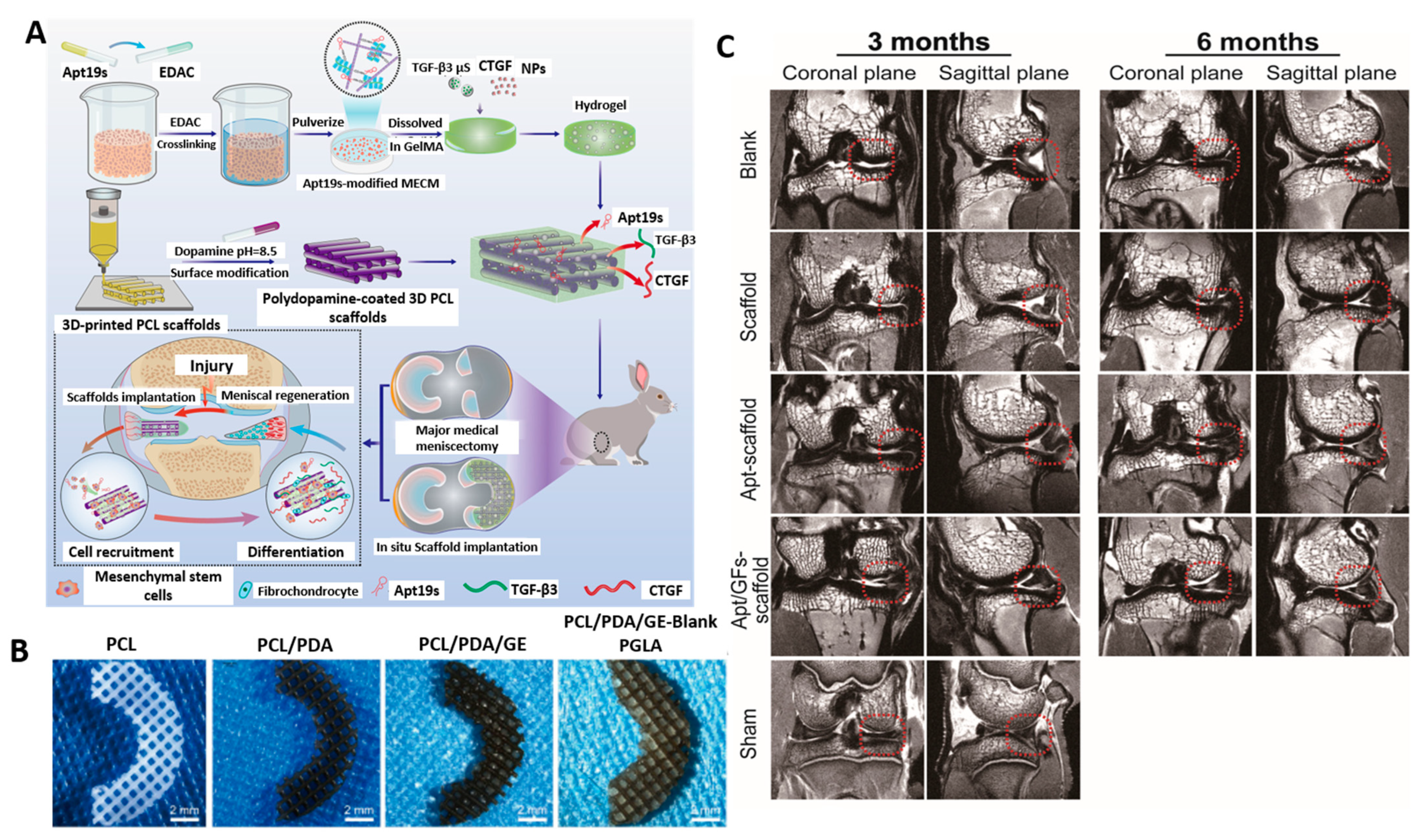
- Li, H.; Zhao, T.; Cao, F.; Deng, H.; He, S.; Li, J.; Liu, S.; Yang, Z.; Yuan, Z.; Guo, Q. Integrated Bioactive Scaffold with Aptamer-targeted Stem Cell Recruitment and Growth Factor-induced Pro-differentiation Effects for Anisotropic Meniscal Regeneration. Bioeng. Transl. Med. 2022, 7, e10302. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeting Agent | Target | Ref. |
|---|---|---|
| Oligopeptides from natural origin | ||
| D6 | HA | [47] |
| D8 | Bone resorption areas | [51] |
| (DSS)6 | Bone formation areas | [53] |
| Peptides generated through phage display | ||
| SVSVGMKPSPRP | HA | [57] |
| MLPHHGA | HA | [58] |
| VTKHLNQISQSY | HA | [59] |
| DSSTPSST | HA | [61] |
| NNHYLPR | HA ({001} face) | [62] |
| KNFQSRSH | HA | [63] |
| TPLSYLKGLVTV | TRAP | [72] |
| YRAPWPP | KS483 osteoblasts | [73] |
| SDSSD | MC3T3-E1 preosteoblastshFOB human osteoblasts | [74] |
| EPLQLKM | Human BMMSCs | [75] |
| DPIYALSWSGMA | Human bone marrow stromal cells | [76] |
| STFTKSP | Hematopoietic stem cells/Mouse BMSCs | [77] |
| CDNVAQSVC | Mouse BMMSCs | [78] |
| CTTNPFSLC | Rat BMMSCs | [79] |
| Aptamers generated through SELEX | ||
| 5′-CAGGGCGCTACGGTATGTGTTGGGTCTGGCGTAGGGCTGGC-3′ | HA | [56] |
| 5′-GAATTCAGTCGGACAGCGCACACGGAACCTCGGAACACAGCTAGCGGGGCTCACTGGATGGACGAATATCGTCTCCC-3′ | SAOS-2 | [64] |
| 5′-AGTCTGTTGGACCGAATCCCGTGGACGCACCCTTTGGACG-3′ | Rat primary osteoblasts | [65] |
| 5′-ACGCTCGGATGCCACTACAGGGATGGGAGGGAGGGGGCTCGTGGCGGCTAGGGGGTATAACTCATGGACGTGCTGGTGAC-3′ | Raw264.7J774A.1 | [67] |
| 5′-GAATTCAGTCGGACAGCGACGACGGTGATATGTCAAGGTCGTATGCACGAGTCAGAGGGATGGACGAATATCGTCTCCC-3′ | Mouse BMMSCs | [68] |
| 5′-AGGTCAGATGAGGAGGGGGACTTAGGACTGGGTTTATGACCTATGCGTG-3′ | Human PSCs | [69] |
| 5′-TGCGTGTGTAGTGTGTCTGCATGCCCCTGTAATCGCCCATGGGTAGCCTCTTAGGGATTTGGGCGG-3′ | Human PSCs | [70] |
| 5′- GGGAGACAAGAATAAACGCTCAACAAATGGGTGGGTGTGGTGGGTGTGAAGGTGCGAGTTGATTCGACAGGAGGCTCACAACAGGC-3′ | Human jaw periosteal cells | [71] |
| Description | In Vivo Model | Ref. |
|---|---|---|
| Alendronate | ||
| Liposomes that transfect the SDF-1 into osteoblastic cells to trigger the migration of MSCs to the bone marrow | C57BL/6 mice | [80] |
| PLGA NPs loaded with 17β estradiol that is released upon heat generation after application of a magnetic field | OVX SD rats | [81] |
| Polymer vesicles carrying β estradiol that acts synergistically with the targeting bisphosphonate | OVX SD rats | [82] |
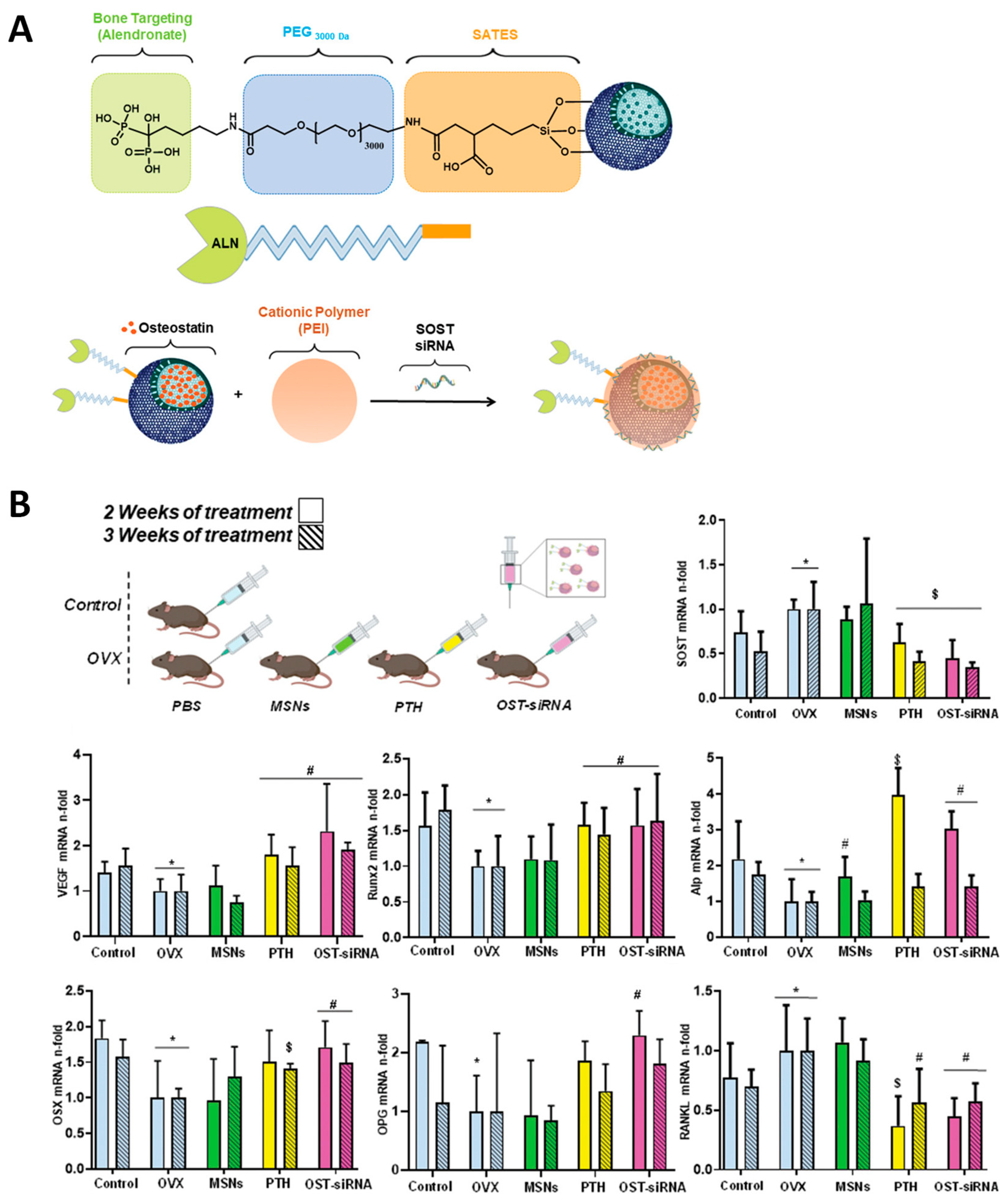
| MSNs carrying an osteogenic peptide and an SOST siRNA that exert synergistic osteogenic effect | OVX C57BL/6 mice | [83] |
| Tetracycline | ||
| Polymeric NPs that deliver simvastatin locally in the bone | OVX SD rats | [84] |
| Lipid-coated nanocarrier for the delivery of Ca2+ ions and simvastatin | OVX ICR mice | [85] |
| (Asp)n | ||
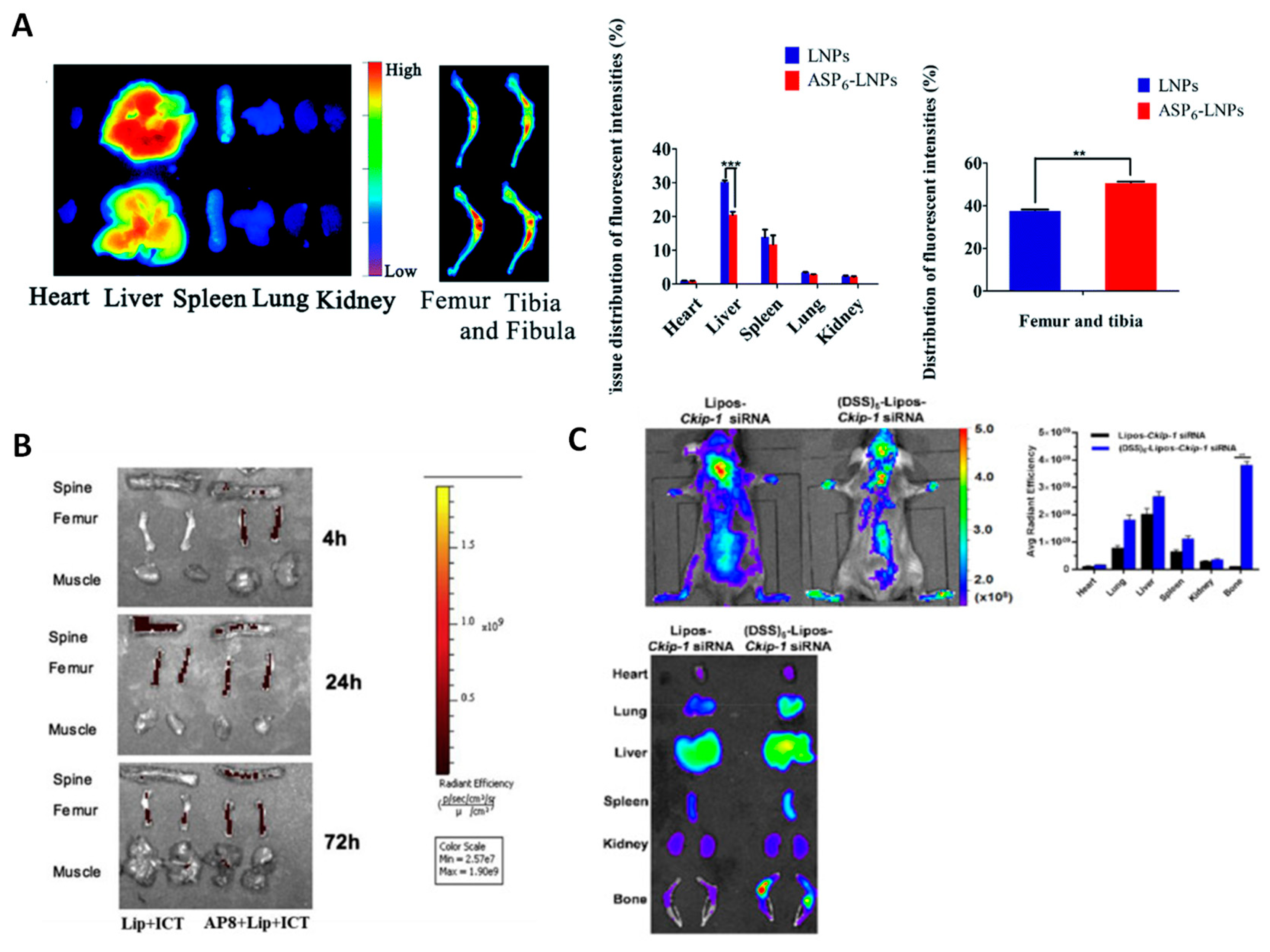
| (Asp)6-coated lipid NPs loaded with simvastatin | OVX ICR mice | [46] |
| Dendritic (Asp)3-PLGA NPs loaded with simvastatin | OVC SD rats and Disuse SD rats | [86] |
| (Asp)8-coated liposomes carrying icaritin that promote osteogenic, rather than adipogenic differentiation of BMSCs | OVX C57/BL6 mice | [87] |
| (Asp)8-coated lipid NPs carrying an miR-21 inhibitor to inhibit osteoclastogenesis | OVX C57/BL6 mice (WT and miR-21-defficient) | [89] |
| (Asp)8-coated polyurethane nanomicelles for the delivery of antimiRNA-214 to inhibit osteoclastogenesis | OVX C57/BL6 mice | [90] |
| (AspSerSer)6 | ||
| Liposomes loaded with an miR-33-5p mimic to promote activity and mineralization of osteoblasts | Hindlimb unloading C57/BL6 mice | [91] |
| Transfecting agents carrying a plasmid encoding the Semaphorin 3A gene to increase the number of osteoblasts and reduce that of osteoclasts simultaneously | OVX Kunming mice | [92] |
| Cationic liposomes for the delivery of Plekho1 siRNA to osteoblasts | OVX SD rats | [53] |
| Liposomes encapsulating Ckip-1 siRNA to deliver it to osteoblasts | OVX C57/BL6 mice (WT and Ckip-1 knockdown) | [88] |
| Adeno-associated virus 9 loaded with a miRNA targeting shn3 in osteoblasts | OVX BALB/cJ (Shn3−/−) and C57BL/6J (Shn3fl/fl) mice | [93] |
| Adeno-associated virus 9 delivering an miRNA able to silence RANK and cathepsin K expression | OVX BALB/cJ and C57BL/6J mice | [94] |
| Peptides generated through phage display | ||
| SVSVGMKPSPRP-coated amorphous calcium phosphate NPs to target enamel HA surface | - | [95] |
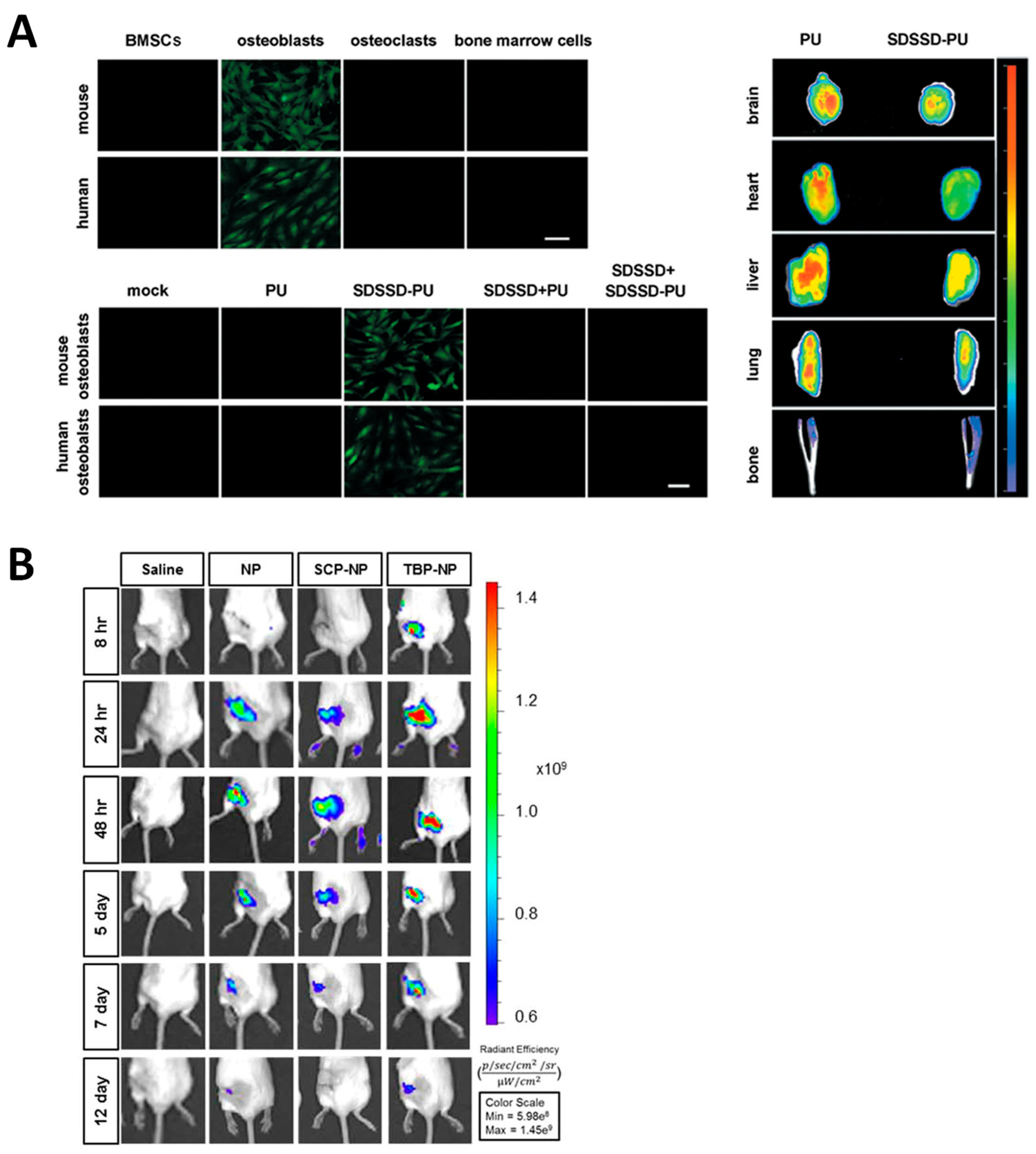
| SDSSD-coated polyurethane nanomicelles carrying antimiR-214 to silence it in osteoblasts | OVX mice | [74] |
| SDSSD-coated exosomes derived from pluripotent stem cells to deliver Shn3 siRNA to osteoblasts | OVX C57BL/6J mice | [96] |
| TRAP peptide-coated polymeric NPs to deliver a β-catenin agonist able to inhibit the glycogen synthase kinase 3 beta | Fracture model in BALB/c mice | [97] |
| Aptamers generated through SELEX | ||
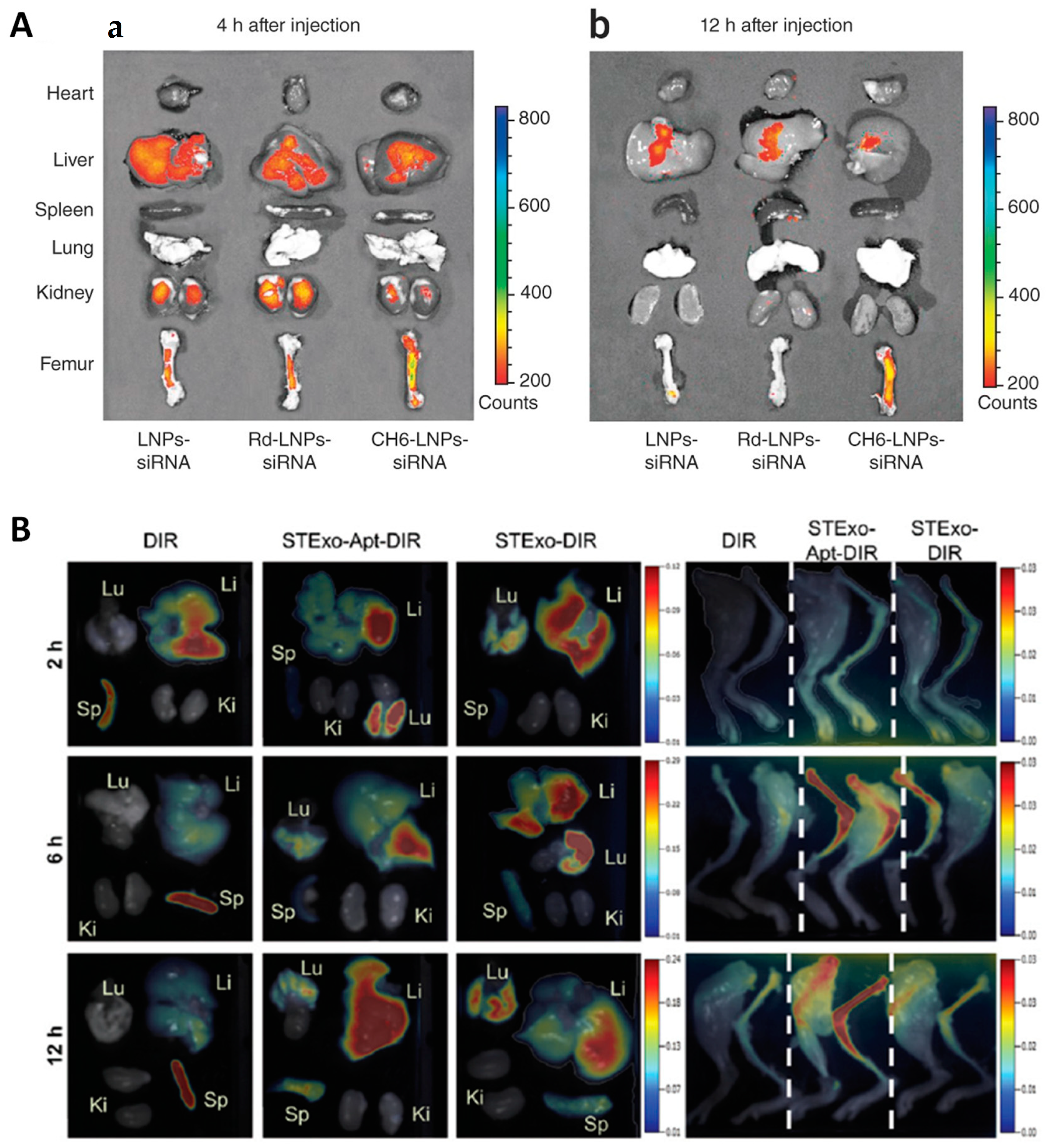
| CH6-targeted lipid NPs for the delivery of Plekho1 siRNA to osteoblasts | OVX SD rats | [65] |
| Assembly of HM69 aptamer into nanoballs to trigger the recruitment of BMSCs | Defect created in SD rats | [70] |
| Lipid NPs modified with the aptamer 5′-GAATTCAGTCGGACAGCGCACACGGAACCTCGGAACACAGCTAGCGGGGCTCACTGGATGGACGAATATCGTCTCCC-3′ to deliver a Sfrp-1 silencing GapmeR to mouse BMSCs | OVX FVB mice | [98] |
| Bone marrow-derived exosomes with antiosteoporotic features functionalized with the aptamer 5′-ACGACGGTGATATGTCAAGGTCGTATGCACGAGTCAGAGG-3′ | OVX C57BL/6J mice | [99] |
| Description | In Vivo Model | Ref. |
|---|---|---|
| EPLQLKM | ||
| Scaffolds functionalized with this peptide and with the peptide P15 that achieve simultaneous cartilage and subchondral bone regeneration in rabbit osteochondral defect model | NZ rabbit | [135] |
| Silk fibroin electrospun scaffolds coated with PDA to induce osteogenic differentiation of BMSCs | SD rats | [136] |
| PLA/gelatin scaffolds containing glycosaminoglycan for improved BMSC migration and differentiation to either chondrogenic or osteogenic phenotypes in a knee osteochondral defect | NZ rabbit | [137] |
| CTTNPFSLC | ||
| β-tricalcium phosphate scaffolds that enhance the adhesion, expansion, and proliferation of BMSCs | SD rats | [76] |
| CDNVAQSVC | ||
| β-TCP scaffolds that enhance BMMSC recruitment with potential application in osteonecrosis treatment | ONFH/NZ rabbit | [139] |
| DPIYALSWSGMA | ||
| β-TCP scaffolds that demonstrate enhanced BMMSC adhesion and proliferation | - | [142] |
| β-TCP scaffolds that recruit BMMSCs and improve osteonecrosis of the femoral head | NZ rabbit | [143] |
| Bone-like mineral functionalized with a dual-peptide containing also the mineral binding sequence VTKHLNQISQSY, which improves bone regeneration thanks to recruiting iPS cells | NIH-Lystbg-JFoxn1nuBtkxid, Charles Rivers mice | [141] |
| SDSSD | ||
| Chitosan scaffolds bearing this peptide grafted and loaded for OB recruitment and promotion of M2 macrophage polarization | SD rats | [138] |
| PFSSTKT | ||
| Hydrogel combining an oriented acellular cartilage matrix with a self-assembling peptide containing the mentioned sequence for specific migration of endogenous stem cells and subsequent chondrogenic differentiation | NZ rabbit | [137] |
| GelMA hydrogel containing chondrocyte extracellular matrix particles decorated with this peptide for the recruitment of BMMSCs and chondrogenic differentiation | NZ rabbit | [138] |
| Description | In Vivo Model | Ref. |
|---|---|---|
| CH6 | ||
| Macro-mesoporous bioactive glass scaffolds functionalized with reduced graphene oxide for specific osteoblast recruitment and scaffold-mediated differentiation for bone defect regeneration | SD rats | [145] |
| HM69 | ||
| GelMA/ PCL scaffolds for recruitment and chondrogenic differentiation of MSCs for cartilage repair | C57/BL6 mice | [146] |
| 74 | ||
| β-TCP scaffolds’ potential enhances JPC cell adhesion on 3D constructs and mineralization on 2D surfaces | - | [147] |
| 19S | ||
| Bioactive titanium implants for recruitment and differentiation of BMMSCs for formation of new bone | SD rats | [148] |
| Graphene-oxide-based scaffold engineered in two layers of different composition, one targeting the cartilage and the other one targeting the subchondral bone defect for efficient repair of osteochondral defect | SD rats | [149] |
| Silk-fibroin-based scaffolds containing hyaluronic acid to improve the chondrogenic capacity for efficient repair of osteochondral defect. | NZ rabbits | [150] |
| GelMA/PCL scaffold containing PLGA microparticles and nanoparticles loaded with different factors for improved meniscus regeneration | SD rats/NZ rabbits | [151] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gisbert-Garzarán, M.; Gómez-Cerezo, M.N.; Vallet-Regí, M. Targeting Agents in Biomaterial-Mediated Bone Regeneration. Int. J. Mol. Sci. 2023, 24, 2007. https://doi.org/10.3390/ijms24032007
Gisbert-Garzarán M, Gómez-Cerezo MN, Vallet-Regí M. Targeting Agents in Biomaterial-Mediated Bone Regeneration. International Journal of Molecular Sciences. 2023; 24(3):2007. https://doi.org/10.3390/ijms24032007
Chicago/Turabian StyleGisbert-Garzarán, Miguel, María Natividad Gómez-Cerezo, and María Vallet-Regí. 2023. "Targeting Agents in Biomaterial-Mediated Bone Regeneration" International Journal of Molecular Sciences 24, no. 3: 2007. https://doi.org/10.3390/ijms24032007
APA StyleGisbert-Garzarán, M., Gómez-Cerezo, M. N., & Vallet-Regí, M. (2023). Targeting Agents in Biomaterial-Mediated Bone Regeneration. International Journal of Molecular Sciences, 24(3), 2007. https://doi.org/10.3390/ijms24032007