
Mexican Colorectal Cancer Research Consortium (MEX-CCRC): Etiology, Diagnosis/Prognosis, and Innovative Therapies

 , , , , ,
, , , , ,  , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. The Problem
2. Creation of the Consortium
3. Background
4. The Mexican Colorectal Cancer Research Consortium Contributions
4.1. Etiology
4.1.1. Microbiota
4.1.2. Obesity
4.1.3. Food Additives
Food-Grade Titanium Dioxide Enhances Tumor Formation in the Colon
4.2. Diagnosis and Prognosis
4.2.1. Early Detection of CRC by Liquid Biopsy
4.2.2. A Role for the Immune Response during CRC
4.2.3. Macrophage Migration Inhibitory Factor (MIF) Is a Crucial Regulator of CRC Initiation
4.2.4. A Role for STAT (Signal Transducer and Activator of Transcription) Signaling on CRC Initiation and Progression
4.2.5. Immunosuppression in the CAC Model
4.3. Novel Therapies Development
4.3.1. Chemical Therapies
4.3.2. Traditional Mexican Medicine: Cyrtocarpa procera
4.3.3. Drug Repositioning
4.3.4. Targeting microRNAs (miRNAs)
4.3.5. Metabolic Blockade Therapy: Autophagy
4.3.6. Excreted-Secreted Products of Helminths
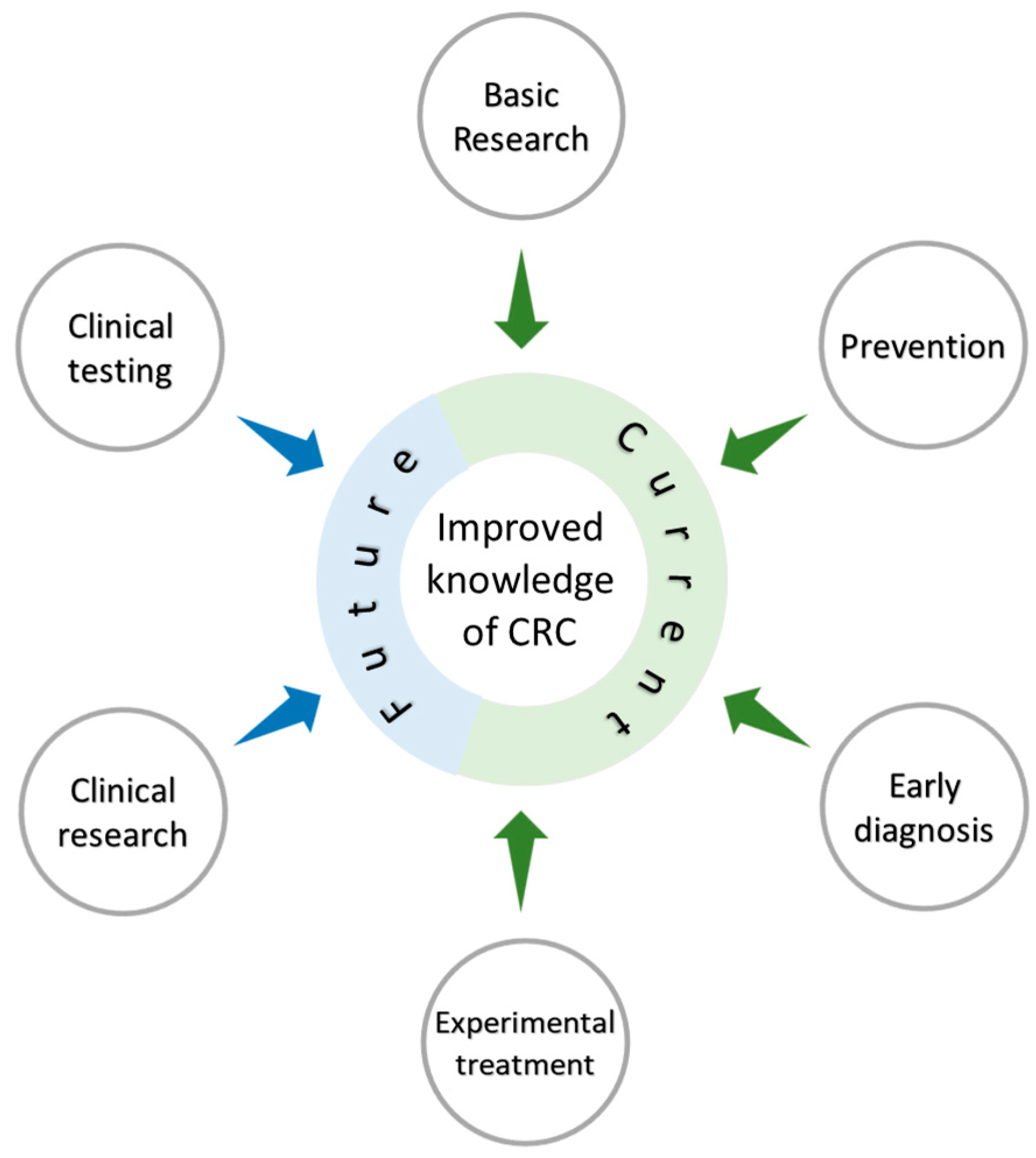
5. Concluding Remarks
6. Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Agency for Research on Cancer, W.H.O. Cancer Over Time 1990–2020. Available online: https://gco.iarc.fr/overtime/en (accessed on 26 August 2022).
- International Agency for Research on Cancer, W.H.O. Cancer Tomorrow. Available online: https://gco.iarc.fr/tomorrow/en (accessed on 10 September 2022).
- Lichtenstern, C.R.; Ngu, R.K.; Shalapour, S.; Karin, M. Immunotherapy, inflammation and colorectal cancer. Cells 2020, 9, 618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islami, F.; Goding Sauer, A.; Miller, K.D.; Siegel, R.L.; Fedewa, S.A.; Jacobs, E.J.; McCullough, M.L.; Patel, A.V.; Ma, J.; Soerjomataram, I. Proportion and number of cancer cases and deaths attributable to potentially modifiable risk factors in the United States. CA A Cancer J. Clin. 2018, 68, 31–54. [Google Scholar] [CrossRef] [PubMed]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114.e2105. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Luo, C.; Zhang, H. Early detection of ulcerative colitis-associated colorectal cancer. Gastroenterol. Rep. 2018, 6, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Jess, T.; Rungoe, C.; Peyrin–Biroulet, L. Risk of colorectal cancer in patients with ulcerative colitis: A meta-analysis of population-based cohort studies. Clin. Gastroenterol. Hepatol. 2012, 10, 639–645. [Google Scholar] [CrossRef]
- Becker, W.R.; Nevins, S.A.; Chen, D.C.; Chiu, R.; Horning, A.M.; Guha, T.K.; Laquindanum, R.; Mills, M.; Chaib, H.; Ladabaum, U.; et al. Single-cell analyses define a continuum of cell state and composition changes in the malignant transformation of polyps to colorectal cancer. Nat. Genet. 2022, 54, 985–995. [Google Scholar] [CrossRef]
- Mangifesta, M.; Mancabelli, L.; Milani, C.; Gaiani, F.; de’Angelis, N.; de’Angelis, G.L.; van Sinderen, D.; Ventura, M.; Turroni, F. Mucosal microbiota of intestinal polyps reveals putative biomarkers of colorectal cancer. Sci. Rep. 2018, 8, 13974. [Google Scholar] [CrossRef] [Green Version]
- Van Cutsem, E.; Oliveira, J. Advanced colorectal cancer: ESMO clinical recommendations for diagnosis, treatment and follow-up. Ann. Oncol. 2009, 20, iv61–iv63. [Google Scholar] [CrossRef] [PubMed]
- Van der Stok, E.P.; Spaander, M.C.; Grünhagen, D.J.; Verhoef, C.; Kuipers, E.J. Surveillance after curative treatment for colorectal cancer. Nat. Rev. Clin. Oncol. 2017, 14, 297–315. [Google Scholar] [CrossRef]
- Ladabaum, U.; Dominitz, J.A.; Kahi, C.; Schoen, R.E. Strategies for Colorectal Cancer Screening. Gastroenterology 2020, 158, 418–432. [Google Scholar] [CrossRef]
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- Lizárraga-Verdugo, E.; Ruiz-García, E.; Lopez-Camarillo, C.; Bermúdez, M.; Avendaño-Félix, M.; Ramos-Payán, R.; Romero-Quintana, G.; Ayala-Ham, A.; Villegas-Mercado, C.; Pérez-Plasencia, C. Cell survival is regulated via SOX9/BCL2L1 Axis in HCT-116 colorectal cancer cell line. J. Oncol. 2020, 2020, 5701527. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-H.; Chen, Y.-X.; Fang, J.-Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, C.V.; de Camargo, M.R.; Russo, E.; Amedei, A. Role of diet and gut microbiota on colorectal cancer immunomodulation. World J. Gastroenterol. 2019, 25, 151. [Google Scholar] [CrossRef]
- Weng, Y.J.; Gan, H.Y.; Li, X.; Huang, Y.; Li, Z.C.; Deng, H.M.; Chen, S.Z.; Zhou, Y.; Wang, L.S.; Han, Y.P. Correlation of diet, microbiota and metabolite networks in inflammatory bowel disease. J. Dig. Dis. 2019, 20, 447–459. [Google Scholar] [CrossRef]
- Song, M.; Garrett, W.S.; Chan, A.T. Nutrients, foods, and colorectal cancer prevention. Gastroenterology 2015, 148, 1244–1260.e1216. [Google Scholar] [CrossRef] [Green Version]
- Donovan, M.G.; Selmin, O.I.; Doetschman, T.C.; Romagnolo, D.F. Mediterranean diet: Prevention of colorectal cancer. Front. Nutr. 2017, 4, 59. [Google Scholar] [CrossRef]
- Illescas, O.; Rodríguez-Sosa, M.; Gariboldi, M. Mediterranean diet to prevent the development of colon diseases: A meta-analysis of gut Microbiota studies. Nutrients 2021, 13, 2234. [Google Scholar] [CrossRef]
- Franco-Esquivias, A.P.; Pena, C.; Torres-Lozano, C.; Vaca-Paniagua, F.; Díaz-Velásquez, C.; Ortega-Cisneros, M.; Quintero-Ramos, A. Gut microbiota in Mexican patients with common variable immunodeficiency. Gac. Med. Mex. 2019, 155, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Clinton, S.K.; Giovannucci, E.L.; Hursting, S.D. The World Cancer Research Fund/American Institute for Cancer Research third expert report on diet, nutrition, physical activity, and cancer: Impact and future directions. J. Nutr. 2020, 150, 663–671. [Google Scholar] [CrossRef]
- Bardou, M.; Barkun, A.N.; Martel, M. Obesity and colorectal cancer. Gut 2013, 62, 933–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Yang, Y.; Wang, F.; Zhang, P.; Shi, C.; Zou, Y.; Qin, H. Obesity and risk of colorectal cancer: A systematic review of prospective studies. PLoS ONE 2013, 8, e53916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, Y.; Vaca-Paniagua, F.; Herrera, L.; Oñate, L.; Herrera-Goepfert, R.; Navarro-Martínez, G.; Cerrato, D.; Díaz-Velázquez, C.; Quezada, E.M.; García-Cuellar, C. Nutritional Indexes as Predictors of Survival and Their Genomic Implications in Gastric Cancer Patients. Nutr. Cancer 2021, 73, 1429–1439. [Google Scholar] [CrossRef]
- Bou Malhab, L.J.; Abdel-Rahman, W.M. Obesity and inflammation: Colorectal cancer engines. Curr. Mol. Pharmacol. 2022, 15, 620–646. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kim, C.; Jung, S.; Park, Y.; Lee, Y.; Jo, J.; Hong, M.; Lee, S.; Oh, Y.; Jung, K. Determination and identification of titanium dioxide nanoparticles in confectionery foods, marketed in South Korea, using inductively coupled plasma optical emission spectrometry and transmission electron microscopy. Food Addit. Contam. Part A 2018, 35, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Ortega, I.M.; Garduño-Balderas, L.G.; Delgado-Buenrostro, N.L.; Freyre-Fonseca, V.; Flores-Flores, J.O.; González-Robles, A.; Pedraza-Chaverri, J.; Hernández-Pando, R.; Rodríguez-Sosa, M.; León-Cabrera, S. Food-grade titanium dioxide exposure exacerbates tumor formation in colitis associated cancer model. Food Chem. Toxicol. 2016, 93, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Bettini, S.; Boutet-Robinet, E.; Cartier, C.; Comera, C.; Gaultier, E.; Dupuy, J.; Naud, N.; Tache, S.; Grysan, P.; Reguer, S.; et al. Food-grade TiO2 impairs intestinal and systemic immune homeostasis, initiates preneoplastic lesions and promotes aberrant crypt development in the rat colon. Sci. Rep. 2017, 7, 40373. [Google Scholar] [CrossRef] [Green Version]
- Proquin, H.; Jetten, M.J.; Jonkhout, M.C.; Garduno-Balderas, L.G.; Briede, J.J.; de Kok, T.M.; Chirino, Y.I.; van Loveren, H. Gene expression profiling in colon of mice exposed to food additive titanium dioxide (E171). Food Chem. Toxicol. 2018, 111, 153–165. [Google Scholar] [CrossRef]
- Proquin, H.; Rodriguez-Ibarra, C.; Moonen, C.G.; Urrutia Ortega, I.M.; Briede, J.J.; de Kok, T.M.; van Loveren, H.; Chirino, Y.I. Titanium dioxide food additive (E171) induces ROS formation and genotoxicity: Contribution of micro and nano-sized fractions. Mutagenesis 2017, 32, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Medina-Reyes, E.I.; Delgado-Buenrostro, N.L.; Díaz-Urbina, D.; Rodríguez-Ibarra, C.; Déciga-Alcaraz, A.; González, M.I.; Reyes, J.L.; Villamar-Duque, T.E.; Flores-Sánchez, M.L.; Hernández-Pando, R. Food-grade titanium dioxide (E171) induces anxiety, adenomas in colon and goblet cells hyperplasia in a regular diet model and microvesicular steatosis in a high fat diet model. Food Chem. Toxicol. 2020, 146, 111786. [Google Scholar] [CrossRef]
- Martínez-Gregorio, H.; Díaz-Velásquez, C.; Romero-Piña, M.E.; De La Cruz, M.R.; Delgado-Buenrostro, N.L.; De La Cruz-Montoya, A.; Chirino, Y.I.; Ignacio, L.J.J.o.C. Early detection of colorectal cancer somatic mutations using cfDNA liquid biopsies in a murine carcinogenesis model. J. Cancer 2022, 13, 3404–3414. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Xu, C.; Hong, S.; Xia, X.; Cao, Y.; McDermott, J.; Mu, Y.; Han, J.-D.J. Immune cell types and secreted factors contributing to inflammation-to-cancer transition and immune therapy response. Cell Rep. 2019, 26, 1965–1977.e1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, Y.P.; Abadia-Molina, A.C.; Satoskar, A.R.; Clarke, K.; Rietdijk, S.T.; Faubion, W.A.; Mizoguchi, E.; Metz, C.N.; Al Sahli, M.; Ten Hove, T. Development of chronic colitis is dependent on the cytokine MIF. Nat. Immunol. 2001, 2, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Klemke, L.; De Oliveira, T.; Witt, D.; Winkler, N.; Bohnenberger, H.; Bucala, R.; Conradi, L.-C.; Schulz-Heddergott, R. Hsp90-stabilized MIF supports tumor progression via macrophage recruitment and angiogenesis in colorectal cancer. Cell Death Dis. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Choi, S.; Kim, H.-R.; Leng, L.; Kang, I.; Jorgensen, W.L.; Cho, C.-S.; Bucala, R.; Kim, W.-U. Role of macrophage migration inhibitory factor in the regulatory T cell response of tumor-bearing mice. J. Immunol. 2012, 189, 3905–3913. [Google Scholar] [CrossRef] [Green Version]
- Legendre, H.; Decaestecker, C.; Nagy, N.; Hendlisz, A.; Schüring, M.-P.; Salmon, I.; Gabius, H.-J.; Pector, J.-C.; Kiss, R. Prognostic values of galectin-3 and the macrophage migration inhibitory factor (MIF) in human colorectal cancers. Mod. Pathol. 2003, 16, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Fernández, T.; Juárez-Avelar, I.; Illescas, O.; Terrazas, L.I.; Hernández-Pando, R.; Pérez-Plasencia, C.; Gutiérrez-Cirlos, E.B.; Ávila-Moreno, F.; Chirino, Y.I.; Reyes, J.L. Macrophage migration inhibitory factor promotes the interaction between the tumor, macrophages, and T cells to regulate the progression of chemically induced colitis-associated colorectal cancer. Mediat. Inflamm. 2019, 2019, 2056085. [Google Scholar] [CrossRef] [Green Version]
- Juárez-Avelar, I.; Rodríguez, T.; García-García, A.P.; Rodríguez-Sosa, M. Macrophage migration inhibitory factor (MIF): Its role in the genesis and progression of colorectal cancer. In Immunotherapy in Resistant Cancer: From the Lab Bench Work to Its Clinical Perspectives; Elsevier: Amsterdam, The Netherlands, 2021; pp. 173–193. [Google Scholar]
- Yu, H.; Pardoll, D.; Jove, R. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar] [CrossRef]
- Wang, C.G.; Ye, Y.J.; Yuan, J.; Liu, F.F.; Zhang, H.; Wang, S. EZH2 and STAT6 expression profiles are correlated with colorectal cancer stage and prognosis. World J. Gastroenterol. 2010, 16, 2421–2427. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Ramirez, Y.; Colly, V.; Villanueva Gonzalez, G.; Leon-Cabrera, S. Signal transducer and activator of transcription 6 as a target in colon cancer therapy. Oncol. Lett. 2020, 20, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Leon-Cabrera, S.A.; Molina-Guzman, E.; Delgado-Ramirez, Y.G.; Vázquez-Sandoval, A.; Ledesma-Soto, Y.; Pérez-Plasencia, C.G.; Chirino, Y.I.; Delgado-Buenrostro, N.L.; Rodríguez-Sosa, M.; Vaca-Paniagua, F. Lack of STAT6 attenuates inflammation and drives protection against early steps of colitis-associated colon cancer. Cancer Immunol. Res. 2017, 5, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Ramirez, Y.; Ocaña-Soriano, A.; Ledesma-Soto, Y.; Olguín, J.E.; Hernandez-Ruiz, J.; Terrazas, L.I.; Leon-Cabrera, S. STAT6 Is Critical for the Induction of Regulatory T Cells In Vivo Controlling the Initial Steps of Colitis-Associated Cancer. Int. J. Mol. Sci. 2021, 22, 4049. [Google Scholar] [CrossRef]
- Leon-Cabrera, S.; Vázquez-Sandoval, A.; Molina-Guzman, E.; Delgado-Ramirez, Y.; Delgado-Buenrostro, N.L.; Callejas, B.E.; Chirino, Y.I.; Pérez-Plasencia, C.; Rodríguez-Sosa, M.; Olguín, J.E. Deficiency in STAT1 signaling predisposes gut inflammation and prompts colorectal cancer development. Cancers 2018, 10, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Ramirez, Y.; Baltazar-Perez, I.; Martinez, Y.; Callejas, B.E.; Medina-Andrade, I.; Olguín, J.E.; Delgado-Buenrostro, N.L.; Chirino, Y.I.; Terrazas, L.I.; Leon-Cabrera, S. STAT1 Is Required for Decreasing Accumulation of Granulocytic Cells via IL-17 during Initial Steps of Colitis-Associated Cancer. Int. J. Mol. Sci. 2021, 22, 7695. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Rhee, K.J.; Albesiano, E.; Rabizadeh, S.; Wu, X.; Yen, H.R.; Huso, D.L.; Brancati, F.L.; Wick, E.; McAllister, F.; et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat. Med. 2009, 15, 1016–1022. [Google Scholar] [CrossRef]
- Perez-Lopez, A.; Nuccio, S.-P.; Ushach, I.; Edwards, R.A.; Pahu, R.; Silva, S.; Zlotnik, A.; Raffatellu, M. CRTAM shapes the gut microbiota and enhances the severity of infection. J. Immunol. 2019, 203, 532–543. [Google Scholar] [CrossRef]
- Napolitano, S.; Matrone, N.; Muddassir, A.L.; Martini, G.; Sorokin, A.; De Falco, V.; Giunta, E.F.; Ciardiello, D.; Martinelli, E.; Belli, V.; et al. Triple blockade of EGFR, MEK and PD-L1 has antitumor activity in colorectal cancer models with constitutive activation of MAPK signaling and PD-L1 overexpression. J. Exp. Clin. Cancer Res. 2019, 38, 492. [Google Scholar] [CrossRef] [Green Version]
- Pastille, E.; Bardini, K.; Fleissner, D.; Adamczyk, A.; Frede, A.; Wadwa, M.; von Smolinski, D.; Kasper, S.; Sparwasser, T.; Gruber, A.D.; et al. Transient ablation of regulatory T cells improves antitumor immunity in colitis-associated colon cancer. Cancer Res. 2014, 74, 4258–4269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olguin, J.E.; Medina-Andrade, I.; Molina, E.; Vazquez, A.; Pacheco-Fernandez, T.; Saavedra, R.; Perez-Plasencia, C.; Chirino, Y.I.; Vaca-Paniagua, F.; Arias-Romero, L.E.; et al. Early and Partial Reduction in CD4(+)Foxp3(+) Regulatory T Cells during Colitis-Associated Colon Cancer Induces CD4(+) and CD8(+) T Cell Activation Inhibiting Tumorigenesis. J. Cancer 2018, 9, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olguin, J.E.; Medina-Andrade, I.; Rodriguez, T.; Rodriguez-Sosa, M.; Terrazas, L.I. Relevance of Regulatory T Cells during Colorectal Cancer Development. Cancers 2020, 12, 1888. [Google Scholar] [CrossRef] [PubMed]
- Medina-Andrade, I.; Olguín, J.E.; Guerrero-García, S.; Espinosa, J.A.; Garduño-Javier, E.; Hernández-Gómez, V.; Vaca-Paniagua, F.; Rodríguez-Sosa, M.; Terrazas, L.I. Recruitment of M1 Macrophages May Not Be Critical for Protection against Colitis-Associated Tumorigenesis. Int. J. Mol. Sci. 2021, 22, 11204. [Google Scholar] [CrossRef]
- Hollande, F.; Pannequin, J.; Joubert, D. The long road to colorectal cancer therapy: Searching for the right signals. Drug Resist. Updates 2010, 13, 44–56. [Google Scholar] [CrossRef]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment. Int. J. Oncol. 2019, 54, 407–419. [Google Scholar]
- Jacobo-Herrera, N.; Gomez-Quiroz, L.; Pérez-Plasencia, C. Tumor Cell Metabolism and Autophagy as Therapeutic Targets. Front. Oncol. 2020, 10, 2937. [Google Scholar] [CrossRef]
- Hou, J.; Karin, M.; Sun, B. Targeting cancer-promoting inflammation—have anti-inflammatory therapies come of age? Nat. Rev. Clin. Oncol. 2021, 18, 261–279. [Google Scholar] [CrossRef]
- Rodriguez-Canales, M.; Martinez-Galero, E.; Nava-Torres, A.D.; Sanchez-Torres, L.E.; Garduño-Siciliano, L.; Canales-Martinez, M.M.; Terrazas, L.I.; Rodriguez-Monroy, M.A. Anti-Inflammatory and antioxidant activities of the methanolic extract of Cyrtocarpa procera bark reduces the severity of ulcerative colitis in a chemically induced colitis model. Mediat. Inflamm. 2020, 2020, 5062506. [Google Scholar] [CrossRef] [Green Version]
- Calderón-Torres, C.M.; Sarabia-Curz, L.; Ledesma-Soto, Y.; Murguía-Romero, M.; Terrazas, L.I. Denitrase activity of Debaryomyces hansenii reduces the oxidized compound 3-nitrotyrosine in mice liver with colitis. Exp. Ther. Med. 2019, 17, 3748–3754. [Google Scholar] [PubMed]
- Wuerth, R.; Thellung, S.; Bajetto, A.; Mazzanti, M.; Florio, T.; Barbieri, F. Drug-repositioning opportunities for cancer therapy: Novel molecular targets for known compounds. Drug Discov. Today 2016, 21, 190–199. [Google Scholar] [CrossRef] [PubMed]
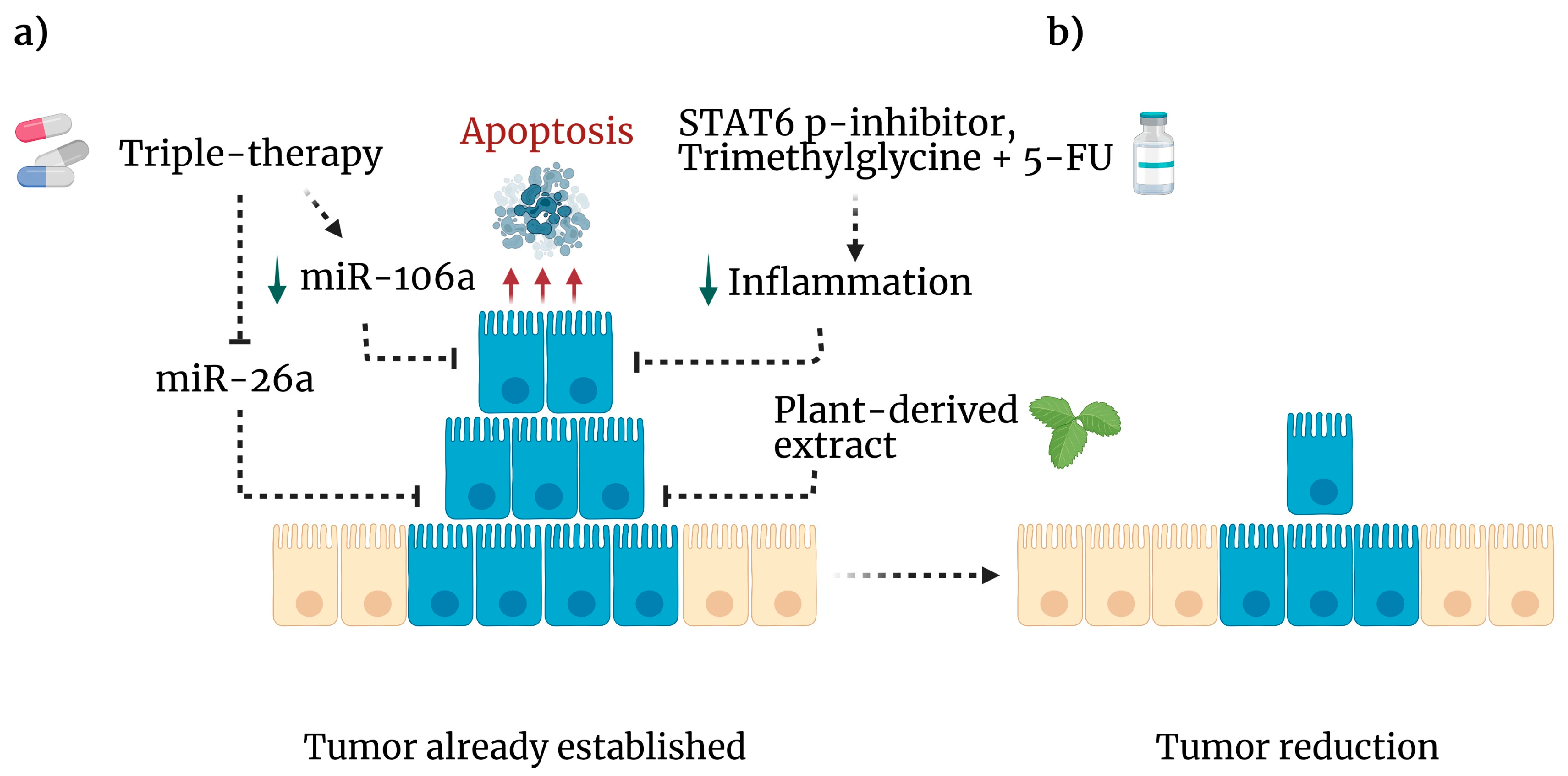
- Figueroa-González, G.; García-Castillo, V.; Coronel-Hernández, J.; López-Urrutia, E.; León-Cabrera, S.; Arias-Romero, L.E.; Terrazas, L.; Rodríguez-Sosa, M.; Campos-Parra, A.D.; Zúñiga-Calzada, E. Anti-inflammatory and antitumor activity of a triple therapy for a colitis-related colorectal cancer. J. Cancer 2016, 7, 1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza-Rodríguez, M.G.; Sánchez-Barrera, C.Á.; Callejas, B.E.; García-Castillo, V.; Beristain-Terrazas, D.L.; Delgado-Buenrostro, N.L.; Chirino, Y.I.; León-Cabrera, S.A.; Rodríguez-Sosa, M.; Gutierrez-Cirlos, E.B. Use of STAT6 phosphorylation inhibitor and trimethylglycine as new adjuvant therapies for 5-fluorouracil in colitis-associated tumorigenesis. Int. J. Mol. Sci. 2020, 21, 2130. [Google Scholar] [CrossRef] [Green Version]
- López-Urrutia, E.; Bustamante Montes, L.P.; Ladrón de Guevara Cervantes, D.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk between long non-coding RNAs, micro-RNAs and mRNAs: Deciphering molecular mechanisms of master regulators in cancer. Front. Oncol. 2019, 9, 669. [Google Scholar] [CrossRef]
- Coronel-Hernández, J.; Salgado-García, R.; Cantú-De León, D.; Jacobo-Herrera, N.; Millan-Catalan, O.; Delgado-Waldo, I.; Campos-Parra, A.D.; Rodríguez-Morales, M.; Delgado-Buenrostro, N.L.; Pérez-Plasencia, C. Combination of Metformin, Sodium Oxamate and Doxorubicin Induces Apoptosis and Autophagy in Colorectal Cancer Cells via Downregulation HIF-1α. Front. Oncol. 2021, 11, 1852. [Google Scholar] [CrossRef]
- López-Urrutia, E.; Coronel-Hernández, J.; García-Castillo, V.; Contreras-Romero, C.; Martínez-Gutierrez, A.; Estrada-Galicia, D.; Terrazas, L.I.; López-Camarillo, C.; Maldonado-Martínez, H.; Jacobo-Herrera, N. MiR-26a downregulates retinoblastoma in colorectal cancer. Tumor Biol. 2017, 39, 1010428317695945. [Google Scholar] [CrossRef] [Green Version]
- Coronel-Hernández, J.; López-Urrutia, E.; Contreras-Romero, C.; Delgado-Waldo, I.; Figueroa-González, G.; Campos-Parra, A.D.; Salgado-García, R.; Martínez-Gutierrez, A.; Rodríguez-Morales, M.; Jacobo-Herrera, N. Cell migration and proliferation are regulated by miR-26a in colorectal cancer via the PTEN–AKT axis. Cancer Cell Int. 2019, 19, 1–14. [Google Scholar] [CrossRef]
- Salgado-García, R.; Coronel-Hernández, J.; Delgado-Waldo, I.; Cantú de León, D.; García-Castillo, V.; López-Urrutia, E.; Gutiérrez-Ruiz, M.; Pérez-Plasencia, C.; Jacobo-Herrera, N. Negative Regulation of Ulk1 by Microrna-106a in Autophagy Induced by a Triple Drug Combination in Colorectal Cancer Cells in Vitro. Genes 2021, 12, 245. [Google Scholar] [CrossRef]
- López-Rosas, I.; López-Camarillo, C.; Salinas-Vera, Y.M.; HernaNdez-De La Cruz, O.N.; Palma-Flores, C.; Chávez-Munguía, B.; Resendis-Antonio, O.; Guillen, N.; Pérez-Plasencia, C.; Álvarez-Sánchez, M.E. Entamoeba histolytica up-regulates microRNA-643 to promote apoptosis by targeting XIAP in human epithelial colon cells. Front. Cell. Infect. Microbiol. 2019, 8, 437. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Josse, C.; Bours, V. MicroRNAs and Inflammation in Colorectal Cancer. Adv Exp Med Biol 2016, 937, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, S.; Ma, B. Autophagy and autophagy-related proteins in cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Lee, S.H. The roles of autophagy in cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Yao, J.; Xie, J.; Liu, Z.; Zhou, Y.; Pan, H.; Han, W. The role of autophagy in colitis-associated colorectal cancer. Signal Transduct. Target. Ther. 2018, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Lizárraga-Verdugo, E.; Avendaño-Félix, M.; Bermúdez, M.; Ramos-Payán, R.; Pérez-Plasencia, C.; Aguilar-Medina, M. Cancer stem cells and its role in angiogenesis and vasculogenic mimicry in gastrointestinal cancers. Front. Oncol. 2020, 10, 413. [Google Scholar] [CrossRef]
- Xie, Q.; Liu, Y.; Li, X. The interaction mechanism between autophagy and apoptosis in colon cancer. Transl. Oncol. 2020, 13, 100871. [Google Scholar] [CrossRef]
- Pérez-Plasencia, C.; López-Urrutia, E.; García-Castillo, V.; Trujano-Camacho, S.; López-Camarillo, C.; Campos-Parra, A.D. Interplay between autophagy and Wnt/β-catenin signaling in cancer: Therapeutic potential through drug repositioning. Front. Oncol. 2020, 10, 1037. [Google Scholar] [CrossRef]
- Perwez Hussain, S.; Harris, C.C. Inflammation and cancer: An ancient link with novel potentials. Int. J. Cancer 2007, 121, 2373–2380. [Google Scholar] [CrossRef]
- Wanders, L.K.; Dekker, E.; Pullens, B.; Bassett, P.; Travis, S.P.; East, J.E. Cancer risk after resection of polypoid dysplasia in patients with longstanding ulcerative colitis: A meta-analysis. Clin. Gastroenterol. Hepatol. 2014, 12, 756–764. [Google Scholar] [CrossRef]
- Leon-Cabrera, S.; Schwertfeger, K.L.; Terrazas, L.I. Inflammation as a target in cancer therapy. Mediat. Inflamm. 2019, 2019, 1971698. [Google Scholar] [CrossRef] [PubMed]
- Greene, E.R.; Huang, S.; Serhan, C.N.; Panigrahy, D. Regulation of inflammation in cancer by eicosanoids. Prostaglandins Other Lipid Mediat. 2011, 96, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrazas, C.A.; Alcántara-Hernández, M.; Bonifaz, L.; Terrazas, L.I.; Satoskar, A.R. Helminth-excreted/secreted products are recognized by multiple receptors on DCs to block the TLR response and bias Th2 polarization in a cRAF dependent pathway. FASEB J. 2013, 27, 4547–4560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machicado, C.; Marcos, L.A. Carcinogenesis associated with parasites other than Schistosoma, Opisthorchis and Clonorchis: A systematic review. Int. J. Cancer 2016, 138, 2915–2921. [Google Scholar] [CrossRef] [PubMed]
- Pastille, E.; Frede, A.; McSorley, H.J.; Grab, J.; Adamczyk, A.; Kollenda, S.; Hansen, W.; Epple, M.; Buer, J.; Maizels, R.M.; et al. Intestinal helminth infection drives carcinogenesis in colitis-associated colon cancer. PLoS Pathog. 2017, 13, e1006649. [Google Scholar] [CrossRef]
- Hayes, K.S.; Cliffe, L.J.; Bancroft, A.J.; Forman, S.P.; Thompson, S.; Booth, C.; Grencis, R.K. Chronic Trichuris muris infection causes neoplastic change in the intestine and exacerbates tumour formation in APC min/+ mice. PLoS Negl. Trop. Dis. 2017, 11, e0005708. [Google Scholar] [CrossRef] [Green Version]
- León-Cabrera, S.; Callejas, B.E.; Ledesma-Soto, Y.; Coronel, J.; Pérez-Plasencia, C.; Gutiérrez-Cirlos, E.B.; Ávila-Moreno, F.; Rodríguez-Sosa, M.; Hernández-Pando, R.; Marquina-Castillo, B. Extraintestinal helminth infection reduces the development of colitis-associated tumorigenesis. Int. J. Biol. Sci. 2014, 10, 948. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.A.; Prince, S.; Smith, K.A. Gastrointestinal Nematode-Derived Antigens Alter Colorectal Cancer Cell Proliferation and Migration through Regulation of Cell Cycle and Epithelial-Mesenchymal Transition Proteins. Int. J. Mol. Sci. 2020, 21, 7845. [Google Scholar] [CrossRef]
- Callejas, B.E.; Mendoza-Rodríguez, M.G.; Villamar-Cruz, O.; Reyes-Martínez, S.; Sánchez-Barrera, C.A.; Rodríguez-Sosa, M.; Delgado-Buenrostro, N.L.; Martínez-Saucedo, D.; Chirino, Y.I.; León-Cabrera, S.A. Helminth-derived molecules inhibit colitis-associated colon cancer development through NF-κB and STAT3 regulation. Int. J. Cancer 2019, 145, 3126–3139. [Google Scholar] [CrossRef]
- Callejas, B.E.; Martinez-Saucedo, D.; Terrazas, L.I. Parasites as negative regulators of cancer. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade-Meza, A.; Arias-Romero, L.E.; Armas-López, L.; Ávila-Moreno, F.; Chirino, Y.I.; Delgado-Buenrostro, N.L.; García-Castillo, V.; Gutiérrez-Cirlos, E.B.; Juárez-Avelar, I.; Leon-Cabrera, S.; et al. Mexican Colorectal Cancer Research Consortium (MEX-CCRC): Etiology, Diagnosis/Prognosis, and Innovative Therapies. Int. J. Mol. Sci. 2023, 24, 2115. https://doi.org/10.3390/ijms24032115
Andrade-Meza A, Arias-Romero LE, Armas-López L, Ávila-Moreno F, Chirino YI, Delgado-Buenrostro NL, García-Castillo V, Gutiérrez-Cirlos EB, Juárez-Avelar I, Leon-Cabrera S, et al. Mexican Colorectal Cancer Research Consortium (MEX-CCRC): Etiology, Diagnosis/Prognosis, and Innovative Therapies. International Journal of Molecular Sciences. 2023; 24(3):2115. https://doi.org/10.3390/ijms24032115
Chicago/Turabian StyleAndrade-Meza, Antonio, Luis E. Arias-Romero, Leonel Armas-López, Federico Ávila-Moreno, Yolanda I. Chirino, Norma L. Delgado-Buenrostro, Verónica García-Castillo, Emma B. Gutiérrez-Cirlos, Imelda Juárez-Avelar, Sonia Leon-Cabrera, and et al. 2023. "Mexican Colorectal Cancer Research Consortium (MEX-CCRC): Etiology, Diagnosis/Prognosis, and Innovative Therapies" International Journal of Molecular Sciences 24, no. 3: 2115. https://doi.org/10.3390/ijms24032115
APA StyleAndrade-Meza, A., Arias-Romero, L. E., Armas-López, L., Ávila-Moreno, F., Chirino, Y. I., Delgado-Buenrostro, N. L., García-Castillo, V., Gutiérrez-Cirlos, E. B., Juárez-Avelar, I., Leon-Cabrera, S., Mendoza-Rodríguez, M. G., Olguín, J. E., Perez-Lopez, A., Pérez-Plasencia, C., Reyes, J. L., Sánchez-Pérez, Y., Terrazas, L. I., Vaca-Paniagua, F., Villamar-Cruz, O., & Rodríguez-Sosa, M. (2023). Mexican Colorectal Cancer Research Consortium (MEX-CCRC): Etiology, Diagnosis/Prognosis, and Innovative Therapies. International Journal of Molecular Sciences, 24(3), 2115. https://doi.org/10.3390/ijms24032115