
Application of Single-Cell and Spatial Omics in Musculoskeletal Disorder Research


Abstract
:1. Introduction
2. Experimental Design
2.1. Single-Cell and Spatial Transcriptomics Techniques and Downstream Analyses
2.2. Biomarkers Applied in the Single-Cell Characterization of the Musculoskeletal System
2.3. Advanced Protocol for Isolating Qualified Single Cells
2.4. Strategies for Hypothesis Generation and Validation in Single-Cell Omics under Different Sample Collection Situations
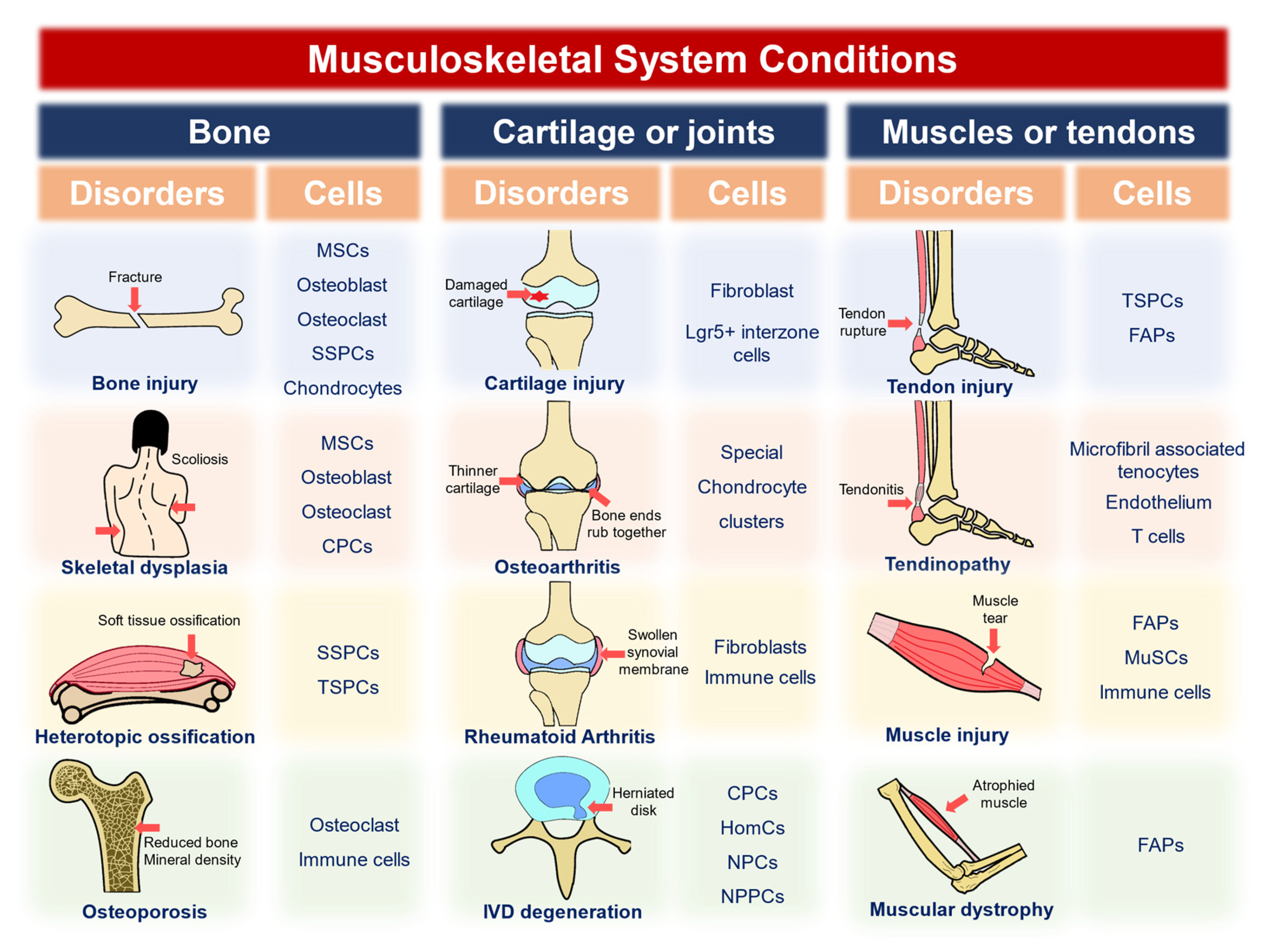
3. Single-Cell and Spatial Omics Were Applied to the Characterization of Various Musculoskeletal Diseases
3.1. Bone-Related Disorders
3.1.1. Bone Injury
3.1.2. Skeletal Dysplasia
3.1.3. Heterotopic Ossification
3.1.4. Osteoporosis
3.2. Cartilage- or Joint-Related Disorders
3.2.1. Cartilage Injury
3.2.2. Osteoarthritis
3.2.3. Rheumatoid Arthritis
3.2.4. IVD Degeneration (IVDD)
3.3. Muscle- or Tendon-Related Disorders
3.3.1. Tendon Injury
3.3.2. Tendinopathy
3.3.3. Muscle Injury
3.3.4. Muscular Dystrophy
4. Treatments
4.1. Implants
4.1.1. Metal Implants
4.1.2. Polymer Implants
4.2. Stem Cell Therapies
4.3. Drugs
5. Emerging Directions for Single-Cell Profiling of Musculoskeletal Diseases
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilliland, K.O.; Kernick, E.T. Musculoskeletal tissues and anatomy. In Clinical Foundations of Musculoskeletal Medicine: A Manual for Medical Students; Esther, R.J., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 11–21. ISBN 978-3-030-42893-8. [Google Scholar]
- Salhotra, A.; Shah, H.N.; Levi, B.; Longaker, M.T. Mechanisms of bone development and repair. Nat. Rev. Mol. Cell Biol. 2020, 21, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Heath, J.R.; Ribas, A.; Mischel, P.S. Single-cell analysis tools for drug discovery and development. Nat. Rev. Drug Discov. 2016, 15, 204–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobasheri, A. Intersection of inflammation and herbal medicine in the treatment of osteoarthritis. Curr. Rheumatol. Rep. 2012, 14, 604–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crevenna, R.; Mickel, M.; Schuhfried, O.; Gesslbauer, C.; Zdravkovic, A.; Keilani, M. Focused extracorporeal shockwave therapy in physical medicine and rehabilitation. Curr. Phys. Med. Rehabil. Rep. 2021, 9, 1–10. [Google Scholar] [CrossRef]
- Richards, J.B.; Zheng, H.-F.; Spector, T.D. Genetics of osteoporosis from genome-wide association studies: Advances and challenges. Nat. Rev. Genet. 2012, 13, 576–588. [Google Scholar] [CrossRef]
- Wang, W.; Ouyang, Y.; Poh, C.K. Orthopaedic implant technology: Biomaterials from past to future. Ann. Acad. Med. Singap. 2011, 40, 237–244. [Google Scholar] [CrossRef]
- Greenblatt, M.B.; Ono, N.; Ayturk, U.M.; Debnath, S.; Lalani, S. The Unmixing Problem: A Guide to Applying Single-Cell RNA Sequencing to Bone. J. Bone Miner. Res. 2019, 34, 1207–1219. [Google Scholar] [CrossRef]
- Ambrosi, T.H.; Longaker, M.T.; Chan, C.K.F. A revised perspective of skeletal stem cell biology. Front. Cell Dev. Biol. 2019, 7, 189. [Google Scholar] [CrossRef] [Green Version]
- Aboul-Soud, M.A.M.; Alzahrani, A.J.; Mahmoud, A. Induced Pluripotent Stem Cells (iPSCs)-Roles in Regenerative Therapies, Disease Modelling and Drug Screening. Cells 2021, 10, 2319. [Google Scholar] [CrossRef]
- Cai, G.; Liu, W.; He, Y.; Huang, J.; Duan, L.; Xiong, J.; Liu, L.; Wang, D. Recent advances in kartogenin for cartilage regeneration. J. Drug Target. 2019, 27, 28–32. [Google Scholar] [CrossRef]
- Tang, F.; Barbacioru, C.; Wang, Y.; Nordman, E.; Lee, C.; Xu, N.; Wang, X.; Bodeau, J.; Tuch, B.B.; Siddiqui, A.; et al. mRNA-Seq whole-transcriptome analysis of a single cell. Nat. Methods 2009, 6, 377–382. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Yu, C.; Xia, K.; Yang, B.; Zhang, Y.; Ying, L.; Wang, C.; Huang, X.; Chen, Q.; et al. Advances in single-cell sequencing and its application to musculoskeletal system research. Cell Prolif. 2022, 55, e13161. [Google Scholar] [CrossRef]
- Chai, R.C. Single-Cell RNA Sequencing: Unravelling the Bone One Cell at a Time. Curr. Osteoporos. Rep. 2022, 20, 356–362. [Google Scholar] [CrossRef]
- Kendal, A.R.; Layton, T.; Al-Mossawi, H.; Appleton, L.; Dakin, S.; Brown, R.; Loizou, C.; Rogers, M.; Sharp, R.; Carr, A. Multi-omic single cell analysis resolves novel stromal cell populations in healthy and diseased human tendon. Sci. Rep. 2020, 10, 13939. [Google Scholar] [CrossRef]
- Stoeckius, M.; Hafemeister, C.; Stephenson, W.; Houck-Loomis, B.; Chattopadhyay, P.K.; Swerdlow, H.; Satija, R.; Smibert, P. Simultaneous epitope and transcriptome measurement in single cells. Nat. Methods 2017, 14, 865–868. [Google Scholar] [CrossRef] [Green Version]
- Sahu, N.; Grandi, F.C.; Bhutani, N. A single-cell mass cytometry platform to map the effects of preclinical drugs on cartilage homeostasis. JCI Insight 2022, 7, e160702. [Google Scholar] [CrossRef]
- Bendall, S.C.; Simonds, E.F.; Qiu, P.; Amir, E.D.; Krutzik, P.O.; Finck, R.; Bruggner, R.V.; Melamed, R.; Trejo, A.; Ornatsky, O.I.; et al. Single-cell mass cytometry of differential immune and drug responses across a human hematopoietic continuum. Science 2011, 332, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Tower, R.J.; Li, Z.; Cheng, Y.-H.; Wang, X.-W.; Rajbhandari, L.; Zhang, Q.; Negri, S.; Uytingco, C.R.; Venkatesan, A.; Zhou, F.-Q.; et al. Spatial transcriptomics reveals a role for sensory nerves in preserving cranial suture patency through modulation of BMP/TGF-β signaling. Proc. Natl. Acad. Sci. USA 2021, 118, e2103087118. [Google Scholar] [CrossRef]
- Carlberg, K.; Korotkova, M.; Larsson, L.; Catrina, A.I.; Ståhl, P.L.; Malmström, V. Exploring inflammatory signatures in arthritic joint biopsies with Spatial Transcriptomics. Sci. Rep. 2019, 9, 18975. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.; Barkley, D.; França, G.S.; Yanai, I. Exploring tissue architecture using spatial transcriptomics. Nature 2021, 596, 211–220. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.M.; Mazutis, L.; Akartuna, I.; Tallapragada, N.; Veres, A.; Li, V.; Peshkin, L.; Weitz, D.A.; Kirschner, M.W. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell 2015, 161, 1187–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.X.Y.; Terry, J.M.; Belgrader, P.; Ryvkin, P.; Bent, Z.W.; Wilson, R.; Ziraldo, S.B.; Wheeler, T.D.; McDermott, G.P.; Zhu, J.; et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 2017, 8, 14049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, K.N. Single-Cell Tagged Reverse Transcription (STRT-Seq). Methods Mol. Biol. 2019, 1979, 133–153. [Google Scholar] [CrossRef]
- Hagemann-Jensen, M.; Ziegenhain, C.; Chen, P.; Ramsköld, D.; Hendriks, G.-J.; Larsson, A.J.M.; Faridani, O.R.; Sandberg, R. Single-cell RNA counting at allele and isoform resolution using Smart-seq3. Nat. Biotechnol. 2020, 38, 708–714. [Google Scholar] [CrossRef]
- Ramsköld, D.; Luo, S.; Wang, Y.-C.; Li, R.; Deng, Q.; Faridani, O.R.; Daniels, G.A.; Khrebtukova, I.; Loring, J.F.; Laurent, L.C.; et al. Full-length mRNA-Seq from single-cell levels of RNA and individual circulating tumor cells. Nat. Biotechnol. 2012, 30, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, W.; Donlin, L.T.; Butler, A.; Rozo, C.; Bracken, B.; Rashidfarrokhi, A.; Goodman, S.M.; Ivashkiv, L.B.; Bykerk, V.P.; Orange, D.E.; et al. Single-cell RNA-seq of rheumatoid arthritis synovial tissue using low-cost microfluidic instrumentation. Nat. Commun. 2018, 9, 791. [Google Scholar] [CrossRef] [Green Version]
- Papalexi, E.; Satija, R. Single-cell RNA sequencing to explore immune cell heterogeneity. Nat. Rev. Immunol. 2018, 18, 35–45. [Google Scholar] [CrossRef]
- Blackburn, D.M.; Lazure, F.; Corchado, A.H.; Perkins, T.J.; Najafabadi, H.S.; Soleimani, V.D. High-resolution genome-wide expression analysis of single myofibers using SMART-Seq. J. Biol. Chem. 2019, 294, 20097–20108. [Google Scholar] [CrossRef]
- Moses, L.; Pachter, L. Museum of spatial transcriptomics. Nat. Methods 2022, 19, 534–546. [Google Scholar] [CrossRef]
- Meier-Ruge, W.; Bielser, W.; Remy, E.; Hillenkamp, F.; Nitsche, R.; Unsöld, R. The laser in the Lowry technique for microdissection of freeze-dried tissue slices. Histochem. J. 1976, 8, 387–401. [Google Scholar] [CrossRef]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.H.; Boettiger, A.N.; Moffitt, J.R.; Wang, S.; Zhuang, X. Spatially resolved, highly multiplexed RNA profiling in single cells. Science 2015, 348, eaaa6090. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Allen, W.E.; Wright, M.A.; Sylwestrak, E.L.; Samusik, N.; Vesuna, S.; Evans, K.; Liu, C.; Ramakrishnan, C.; Liu, J.; et al. Three-dimensional intact-tissue sequencing of single-cell transcriptional states. Science 2018, 361, eaat5691. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Sharon, N.; Bar-Joseph, Z. Temporal modelling using single-cell transcriptomics. Nat. Rev. Genet. 2022, 23, 355–368. [Google Scholar] [CrossRef]
- Cao, J.; Spielmann, M.; Qiu, X.; Huang, X.; Ibrahim, D.M.; Hill, A.J.; Zhang, F.; Mundlos, S.; Christiansen, L.; Steemers, F.J.; et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 2019, 566, 496–502. [Google Scholar] [CrossRef]
- Ji, Z.; Ji, H. TSCAN: Pseudo-time reconstruction and evaluation in single-cell RNA-seq analysis. Nucleic Acids Res. 2016, 44, e117. [Google Scholar] [CrossRef] [Green Version]
- Street, K.; Risso, D.; Fletcher, R.B.; Das, D.; Ngai, J.; Yosef, N.; Purdom, E.; Dudoit, S. Slingshot: Cell lineage and pseudotime inference for single-cell transcriptomics. BMC Genom. 2018, 19, 477. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Bao, E.L.; Wagner, M.; Whitsett, J.A.; Xu, Y. SLICE: Determining cell differentiation and lineage based on single cell entropy. Nucleic Acids Res. 2017, 45, e54. [Google Scholar] [CrossRef]
- Wolf, F.A.; Hamey, F.K.; Plass, M.; Solana, J.; Dahlin, J.S.; Göttgens, B.; Rajewsky, N.; Simon, L.; Theis, F.J. PAGA: Graph abstraction reconciles clustering with trajectory inference through a topology preserving map of single cells. Genome Biol. 2019, 20, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux de Bézieux, H.; Van den Berge, K.; Street, K.N.; Dudoit, S. Trajectory inference across multiple conditions with condiments: Differential topology, progression, differentiation, and expression. bioRxiv 2021. [Google Scholar] [CrossRef]
- Campbell, K.R.; Yau, C. Uncovering pseudotemporal trajectories with covariates from single cell and bulk expression data. Nat. Commun. 2018, 9, 2442. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Almet, A.A.; Cang, Z.; Jin, S.; Nie, Q. The landscape of cell-cell communication through single-cell transcriptomics. Curr. Opin. Syst. Biol. 2021, 26, 12–23. [Google Scholar] [CrossRef]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.-H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef]
- Efremova, M.; Vento-Tormo, M.; Teichmann, S.A.; Vento-Tormo, R. CellPhoneDB: Inferring cell-cell communication from combined expression of multi-subunit ligand-receptor complexes. Nat. Protoc. 2020, 15, 1484–1506. [Google Scholar] [CrossRef]
- Shen, F.; Shi, Y. Recent Advances in Single-Cell View of Mesenchymal Stem Cell in Osteogenesis. Front. Cell Dev. Biol. 2021, 9, 809918. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, M.; Shi, D.; Chen, K.; Zhao, J.; He, S.; Bai, Y.; Shen, P.; Ni, H. Single-cell RNA Seq reveals cellular landscape-specific characteristics and potential etiologies for adolescent idiopathic scoliosis. JOR Spine 2021, 4, e1184. [Google Scholar] [CrossRef]
- He, J.; Yan, J.; Wang, J.; Zhao, L.; Xin, Q.; Zeng, Y.; Sun, Y.; Zhang, H.; Bai, Z.; Li, Z.; et al. Dissecting human embryonic skeletal stem cell ontogeny by single-cell transcriptomic and functional analyses. Cell Res. 2021, 31, 742–757. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Xu, Q.; Li, J.; Zhao, F. Single-cell RNA sequencing analysis dissected the osteo-immunology microenvironment and revealed key regulators in osteoporosis. Int. Immunopharmacol. 2022, 113, 109302. [Google Scholar] [CrossRef]
- Gan, Y.; He, J.; Zhu, J.; Xu, Z.; Wang, Z.; Yan, J.; Hu, O.; Bai, Z.; Chen, L.; Xie, Y.; et al. Spatially defined single-cell transcriptional profiling characterizes diverse chondrocyte subtypes and nucleus pulposus progenitors in human intervertebral discs. Bone Res. 2021, 9, 37. [Google Scholar] [CrossRef]
- Ji, Q.; Zheng, Y.; Zhang, G.; Hu, Y.; Fan, X.; Hou, Y.; Wen, L.; Li, L.; Xu, Y.; Wang, Y.; et al. Single-cell RNA-seq analysis reveals the progression of human osteoarthritis. Ann. Rheum. Dis. 2019, 78, 100–110. [Google Scholar] [CrossRef]
- Chou, C.-H.; Jain, V.; Gibson, J.; Attarian, D.E.; Haraden, C.A.; Yohn, C.B.; Laberge, R.-M.; Gregory, S.; Kraus, V.B. Synovial cell cross-talk with cartilage plays a major role in the pathogenesis of osteoarthritis. Sci. Rep. 2020, 10, 10868. [Google Scholar] [CrossRef]
- De Micheli, A.J.; Spector, J.A.; Elemento, O.; Cosgrove, B.D. A reference single-cell transcriptomic atlas of human skeletal muscle tissue reveals bifurcated muscle stem cell populations. Skelet. Muscle 2020, 10, 19. [Google Scholar] [CrossRef]
- Ding, Y.; Mo, C.; Geng, J.; Li, J.; Sun, Y. Identification of periosteal osteogenic progenitors in jawbone. J. Dent. Res. 2022, 101, 1101–1109. [Google Scholar] [CrossRef]
- Gao, M.; Guo, P.; Liu, X.; Zhang, P.; He, Z.; Wen, L.; Liu, S.; Zhou, Z.; Zhu, W. Systematic study of single-cell isolation from musculoskeletal tissues for single-sell sequencing. BMC Mol. Cell Biol. 2022, 23, 32. [Google Scholar] [CrossRef]
- Debnath, S.; Greenblatt, M.B. Specimen Preparation for Single-Cell Sequencing Analysis of Skeletal Cells. Methods Mol. Biol. 2021, 2221, 89–100. [Google Scholar] [CrossRef]
- Ayturk, U.M.; Scollan, J.P.; Goz Ayturk, D.; Suh, E.S.; Vesprey, A.; Jacobsen, C.M.; Divieti Pajevic, P.; Warman, M.L. Single-Cell RNA Sequencing of Calvarial and Long-Bone Endocortical Cells. J. Bone Miner. Res. 2020, 35, 1981–1991. [Google Scholar] [CrossRef] [PubMed]
- Baryawno, N.; Przybylski, D.; Kowalczyk, M.S.; Kfoury, Y.; Severe, N.; Gustafsson, K.; Kokkaliaris, K.D.; Mercier, F.; Tabaka, M.; Hofree, M.; et al. A cellular taxonomy of the bone marrow stroma in homeostasis and leukemia. Cell 2019, 177, 1915–1932.e16. [Google Scholar] [CrossRef] [PubMed]
- Serowoky, M.A.; Kuwahara, S.T.; Liu, S.; Vakhshori, V.; Lieberman, J.R.; Mariani, F.V. A murine model of large-scale bone regeneration reveals a selective requirement for Sonic Hedgehog. NPJ Regen. Med. 2022, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Z.; Tower, R.J.; Negri, S.; Wang, Y.; Meyers, C.A.; Sono, T.; Qin, Q.; Lu, A.; Xing, X.; et al. NGF-p75 signaling coordinates skeletal cell migration during bone repair. Sci. Adv. 2022, 8, eabl5716. [Google Scholar] [CrossRef] [PubMed]
- Yip, R.K.H.; Chan, D.; Cheah, K.S.E. Mechanistic insights into skeletal development gained from genetic disorders. Curr. Top. Dev. Biol. 2019, 133, 343–385. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, C.; Lu, W.; He, T.; Fan, J.; Wang, C.; Jie, Q.; Chan, D.; Cheah, K.S.E.; Yang, L. Hedgehog signaling orchestrates cartilage-to-bone transition independently of Smoothened. Matrix Biol. 2022, 110, 76–90. [Google Scholar] [CrossRef]
- Mundy, C.; Yao, L.; Sinha, S.; Chung, J.; Rux, D.; Catheline, S.E.; Koyama, E.; Qin, L.; Pacifici, M. Activin A promotes the development of acquired heterotopic ossification and is an effective target for disease attenuation in mice. Sci. Signal. 2021, 14, eabd0536. [Google Scholar] [CrossRef]
- Kozhemyakina, E.; Zhang, M.; Ionescu, A.; Ayturk, U.M.; Ono, N.; Kobayashi, A.; Kronenberg, H.; Warman, M.L.; Lassar, A.B. Identification of a Prg4-expressing articular cartilage progenitor cell population in mice. Arthritis Rheumatol. 2015, 67, 1261–1273. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Lin, Y.-L.; Yan, M.; Li, T.; Wu, E.Y.; Zimmel, K.; Qureshi, O.; Falck, A.; Sherman, K.M.; Huggins, S.S.; et al. Hyaline cartilage differentiation of fibroblasts in regeneration and regenerative medicine. Development 2022, 149, dev200249. [Google Scholar] [CrossRef]
- Wang, X.; Ning, Y.; Zhang, P.; Poulet, B.; Huang, R.; Gong, Y.; Hu, M.; Li, C.; Zhou, R.; Lammi, M.J.; et al. Comparison of the major cell populations among osteoarthritis, Kashin-Beck disease and healthy chondrocytes by single-cell RNA-seq analysis. Cell Death Dis. 2021, 12, 551. [Google Scholar] [CrossRef]
- Lv, Z.; Han, J.; Li, J.; Guo, H.; Fei, Y.; Sun, Z.; Dong, J.; Wang, M.; Fan, C.; Li, W.; et al. Single cell RNA-seq analysis identifies ferroptotic chondrocyte cluster and reveals TRPV1 as an anti-ferroptotic target in osteoarthritis. EBioMedicine 2022, 84, 104258. [Google Scholar] [CrossRef]
- Zhang, F.; Wei, K.; Slowikowski, K.; Fonseka, C.Y.; Rao, D.A.; Kelly, S.; Goodman, S.M.; Tabechian, D.; Hughes, L.B.; Salomon-Escoto, K.; et al. Defining inflammatory cell states in rheumatoid arthritis joint synovial tissues by integrating single-cell transcriptomics and mass cytometry. Nat. Immunol. 2019, 20, 928–942. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, S.; Kong, M.; Tu, Q.; Zhang, L.; Ma, X. Single-cell RNA-seq analysis identifies unique chondrocyte subsets and reveals involvement of ferroptosis in human intervertebral disc degeneration. Osteoarthr. Cartil. 2021, 29, 1324–1334. [Google Scholar] [CrossRef]
- Ling, Z.; Liu, Y.; Wang, Z.; Zhang, Z.; Chen, B.; Yang, J.; Zeng, B.; Gao, Y.; Jiang, C.; Huang, Y.; et al. Single-Cell RNA-Seq Analysis Reveals Macrophage Involved in the Progression of Human Intervertebral Disc Degeneration. Front. Cell Dev. Biol. 2021, 9, 833420. [Google Scholar] [CrossRef]
- Cherif, H.; Mannarino, M.; Pacis, A.S.; Ragoussis, J.; Rabau, O.; Ouellet, J.A.; Haglund, L. Single-Cell RNA-Seq Analysis of Cells from Degenerating and Non-Degenerating Intervertebral Discs from the Same Individual Reveals New Biomarkers for Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2022, 23, 3993. [Google Scholar] [CrossRef]
- Malecova, B.; Gatto, S.; Etxaniz, U.; Passafaro, M.; Cortez, A.; Nicoletti, C.; Giordani, L.; Torcinaro, A.; De Bardi, M.; Bicciato, S.; et al. Dynamics of cellular states of fibro-adipogenic progenitors during myogenesis and muscular dystrophy. Nat. Commun. 2018, 9, 3670. [Google Scholar] [CrossRef] [Green Version]
- De Micheli, A.J.; Laurilliard, E.J.; Heinke, C.L.; Ravichandran, H.; Fraczek, P.; Soueid-Baumgarten, S.; De Vlaminck, I.; Elemento, O.; Cosgrove, B.D. Single-Cell Analysis of the Muscle Stem Cell Hierarchy Identifies Heterotypic Communication Signals Involved in Skeletal Muscle Regeneration. Cell Rep. 2020, 30, 3583–3595.e5. [Google Scholar] [CrossRef] [Green Version]
- Dell’Orso, S.; Juan, A.H.; Ko, K.-D.; Naz, F.; Perovanovic, J.; Gutierrez-Cruz, G.; Feng, X.; Sartorelli, V. Single cell analysis of adult mouse skeletal muscle stem cells in homeostatic and regenerative conditions. Development 2019, 146, dev174177. [Google Scholar] [CrossRef] [Green Version]
- Oprescu, S.N.; Yue, F.; Qiu, J.; Brito, L.F.; Kuang, S. Temporal Dynamics and Heterogeneity of Cell Populations during Skeletal Muscle Regeneration. iScience 2020, 23, 100993. [Google Scholar] [CrossRef]
- Harvey, T.; Flamenco, S.; Fan, C.-M. A Tppp3+Pdgfra+ tendon stem cell population contributes to regeneration and reveals a shared role for PDGF signalling in regeneration and fibrosis. Nat. Cell Biol. 2019, 21, 1490–1503. [Google Scholar] [CrossRef]
- Garcia-Melchor, E.; Cafaro, G.; MacDonald, L.; Crowe, L.A.N.; Sood, S.; McLean, M.; Fazzi, U.G.; McInnes, I.B.; Akbar, M.; Millar, N.L. Novel self-amplificatory loop between T cells and tenocytes as a driver of chronicity in tendon disease. Ann. Rheum. Dis. 2021, 80, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- Wubneh, A.; Tsekoura, E.K.; Ayranci, C.; Uludağ, H. Current state of fabrication technologies and materials for bone tissue engineering. Acta Biomater. 2018, 80, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Gartland, A.; Mason-Savas, A.; Yang, M.; MacKay, C.A.; Birnbaum, M.J.; Odgren, P.R. Septoclast deficiency accompanies postnatal growth plate chondrodysplasia in the toothless (tl) osteopetrotic, colony-stimulating factor-1 (CSF-1)-deficient rat and is partially responsive to CSF-1 injections. Am. J. Pathol. 2009, 175, 2668–2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.R.; Lamplugh, L.; Shepard, N.L.; Mort, J.S. The septoclast, a cathepsin B-rich cell involved in the resorption of growth plate cartilage. J. Histochem. Cytochem. 1995, 43, 525–536. [Google Scholar] [CrossRef] [Green Version]
- Bando, Y.; Sakashita, H.; Taira, F.; Miyake, G.; Ogasawara, Y.; Sakiyama, K.; Owada, Y.; Amano, O. Origin and development of septoclasts in endochondral ossification of mice. Histochem. Cell Biol. 2018, 149, 645–654. [Google Scholar] [CrossRef]
- Sivaraj, K.K.; Majev, P.-G.; Jeong, H.-W.; Dharmalingam, B.; Zeuschner, D.; Schröder, S.; Bixel, M.G.; Timmen, M.; Stange, R.; Adams, R.H. Mesenchymal stromal cell-derived septoclasts resorb cartilage during developmental ossification and fracture healing. Nat. Commun. 2022, 13, 571. [Google Scholar] [CrossRef]
- Li, Z.; Meyers, C.A.; Chang, L.; Lee, S.; Li, Z.; Tomlinson, R.; Hoke, A.; Clemens, T.L.; James, A.W. Fracture repair requires TrkA signaling by skeletal sensory nerves. J. Clin. Investig. 2019, 129, 5137–5150. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, R.E.; Li, Z.; Zhang, Q.; Goh, B.C.; Li, Z.; Thorek, D.L.J.; Rajbhandari, L.; Brushart, T.M.; Minichiello, L.; Zhou, F.; et al. NGF-TrkA Signaling by Sensory Nerves Coordinates the Vascularization and Ossification of Developing Endochondral Bone. Cell Rep. 2016, 16, 2723–2735. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Viejo, M.; Menéndez-Menéndez, Y.; Otero-Hernández, J. CD271 as a marker to identify mesenchymal stem cells from diverse sources before culture. World J. Stem Cells 2015, 7, 470–476. [Google Scholar] [CrossRef]
- Aghajanian, P.; Mohan, S. The art of building bone: Emerging role of chondrocyte-to-osteoblast transdifferentiation in endochondral ossification. Bone Res. 2018, 6, 19. [Google Scholar] [CrossRef]
- Breeland, G.; Sinkler, M.A.; Menezes, R.G. Embryology, Bone Ossification. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- El-Farrash, R.A.; Ali, R.H.; Barakat, N.M. Post-natal bone physiology. Semin. Fetal Neonatal Med. 2020, 25, 101077. [Google Scholar] [CrossRef]
- Kelly, N.H.; Huynh, N.P.T.; Guilak, F. Single cell RNA-sequencing reveals cellular heterogeneity and trajectories of lineage specification during murine embryonic limb development. Matrix Biol. 2020, 89, 1–10. [Google Scholar] [CrossRef]
- Cohen, M.; Kicheva, A.; Ribeiro, A.; Blassberg, R.; Page, K.M.; Barnes, C.P.; Briscoe, J. Ptch1 and Gli regulate Shh signalling dynamics via multiple mechanisms. Nat. Commun. 2015, 6, 6709. [Google Scholar] [CrossRef]
- Lv, X.; Xu, J.; Jiang, J.; Wu, P.; Tan, R.; Wang, B. Genetic animal models of scoliosis: A systematical review. Bone 2021, 152, 116075. [Google Scholar] [CrossRef]
- Latalski, M.; Danielewicz-Bromberek, A.; Fatyga, M.; Latalska, M.; Kröber, M.; Zwolak, P. Current insights into the aetiology of adolescent idiopathic scoliosis. Arch. Orthop. Trauma Surg. 2017, 137, 1327–1333. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Chan, W.C.W.; Lam, Y.; Wang, X.; Chen, P.; Niu, B.; Ng, V.C.W.; Yeo, J.C.; Stricker, S.; Cheah, K.S.E.; et al. Lgr5 and Col22a1 Mark Progenitor Cells in the Lineage toward Juvenile Articular Chondrocytes. Stem Cell Rep. 2019, 13, 713–729. [Google Scholar] [CrossRef]
- Bian, Q.; Cheng, Y.-H.; Wilson, J.P.; Su, E.Y.; Kim, D.W.; Wang, H.; Yoo, S.; Blackshaw, S.; Cahan, P. A single cell transcriptional atlas of early synovial joint development. Development 2020, 147, dev185777. [Google Scholar] [CrossRef]
- Meyers, C.; Lisiecki, J.; Miller, S.; Levin, A.; Fayad, L.; Ding, C.; Sono, T.; McCarthy, E.; Levi, B.; James, A.W. Heterotopic ossification: A comprehensive review. JBMR PLUS 2019, 3, e10172. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, E.F.; Sundaram, M. Heterotopic ossification: A review. Skelet. Radiol. 2005, 34, 609–619. [Google Scholar] [CrossRef]
- Łęgosz, P.; Drela, K.; Pulik, Ł.; Sarzyńska, S.; Małdyk, P. Challenges of heterotopic ossification-Molecular background and current treatment strategies. Clin. Exp. Pharmacol. Physiol. 2018, 45, 1229–1235. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, D.; Wang, H.; Tan, J. Heterotopic ossification of tendon and ligament. J. Cell. Mol. Med. 2020, 24, 5428–5437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Li, B.; Yu, Y.; Li, W.; Qiu, G.; Zhao, Y. The relationship between dural ossification and spinal stenosis in thoracic ossification of the ligamentum flavum. J. Bone Jt. Surg. Am. 2019, 101, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Guo, S.; Qiu, G.; Li, W.; Liu, Y.; Zhao, Y. A potential mechanism of dural ossification in ossification of ligamentum flavum. Med. Hypotheses 2016, 92, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yu, L.; Guo, S.; Zhao, Y. Identification of the molecular mechanism and diagnostic biomarkers in the thoracic ossification of the ligamentum flavum using metabolomics and transcriptomics. BMC Mol. Cell Biol. 2020, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, N.; Chijimatsu, R.; Okada, H.; Oichi, T.; Taniguchi, Y.; Maenohara, Y.; Miyahara, J.; Ishikura, H.; Iwanaga, Y.; Arino, Y.; et al. RSPO2 defines a distinct undifferentiated progenitor in the tendon/ligament and suppresses ectopic ossification. Sci. Adv. 2022, 8, eabn2138. [Google Scholar] [CrossRef]
- Yang, T.-L.; Shen, H.; Liu, A.; Dong, S.-S.; Zhang, L.; Deng, F.-Y.; Zhao, Q.; Deng, H.-W. A road map for understanding molecular and genetic determinants of osteoporosis. Nat. Rev. Endocrinol. 2020, 16, 91–103. [Google Scholar] [CrossRef]
- Karasik, D.; Rivadeneira, F.; Johnson, M.L. The genetics of bone mass and susceptibility to bone diseases. Nat. Rev. Rheumatol. 2016, 12, 323–334. [Google Scholar] [CrossRef]
- Komori, T. Animal models for osteoporosis. Eur. J. Pharmacol. 2015, 759, 287–294. [Google Scholar] [CrossRef]
- Windahl, S.H.; Börjesson, A.E.; Farman, H.H.; Engdahl, C.; Movérare-Skrtic, S.; Sjögren, K.; Lagerquist, M.K.; Kindblom, J.M.; Koskela, A.; Tuukkanen, J.; et al. Estrogen receptor-α in osteocytes is important for trabecular bone formation in male mice. Proc. Natl. Acad. Sci. USA 2013, 110, 2294–2299. [Google Scholar] [CrossRef] [Green Version]
- Kondoh, S.; Inoue, K.; Igarashi, K.; Sugizaki, H.; Shirode-Fukuda, Y.; Inoue, E.; Yu, T.; Takeuchi, J.K.; Kanno, J.; Bonewald, L.F.; et al. Estrogen receptor α in osteocytes regulates trabecular bone formation in female mice. Bone 2014, 60, 68–77. [Google Scholar] [CrossRef]
- Melville, K.M.; Kelly, N.H.; Khan, S.A.; Schimenti, J.C.; Ross, F.P.; Main, R.P.; van der Meulen, M.C.H. Female mice lacking estrogen receptor-alpha in osteoblasts have compromised bone mass and strength. J. Bone Miner. Res. 2014, 29, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Määttä, J.A.; Büki, K.G.; Gu, G.; Alanne, M.H.; Vääräniemi, J.; Liljenbäck, H.; Poutanen, M.; Härkönen, P.; Väänänen, K. Inactivation of estrogen receptor α in bone-forming cells induces bone loss in female mice. FASEB J. 2013, 27, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Iyer, S.; Martin-Millan, M.; Bartell, S.M.; Han, L.; Ambrogini, E.; Onal, M.; Xiong, J.; Weinstein, R.S.; Jilka, R.L.; et al. Estrogen receptor-α signaling in osteoblast progenitors stimulates cortical bone accrual. J. Clin. Investig. 2013, 123, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Martin-Millan, M.; Almeida, M.; Ambrogini, E.; Han, L.; Zhao, H.; Weinstein, R.S.; Jilka, R.L.; O’Brien, C.A.; Manolagas, S.C. The estrogen receptor-alpha in osteoclasts mediates the protective effects of estrogens on cancellous but not cortical bone. Mol. Endocrinol. 2010, 24, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Imai, Y.; Matsumoto, T.; Sato, S.; Takeuchi, K.; Igarashi, K.; Harada, Y.; Azuma, Y.; Krust, A.; Yamamoto, Y.; et al. Estrogen prevents bone loss via estrogen receptor alpha and induction of Fas ligand in osteoclasts. Cell 2007, 130, 811–823. [Google Scholar] [CrossRef]
- Chavassieux, P.; Buffet, A.; Vergnaud, P.; Garnero, P.; Meunier, P.J. Short-term effects of corticosteroids on trabecular bone remodeling in old ewes. Bone 1997, 20, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Quarles, L.D. Prednisone-induced osteopenia in beagles: Variable effects mediated by differential suppression of bone formation. Am. J. Physiol. 1992, 263, E136–E141. [Google Scholar] [CrossRef]
- Baofeng, L.; Zhi, Y.; Bei, C.; Guolin, M.; Qingshui, Y.; Jian, L. Characterization of a rabbit osteoporosis model induced by ovariectomy and glucocorticoid. Acta Orthop. 2010, 81, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Grardel, B.; Sutter, B.; Flautre, B.; Viguier, E.; Lavaste, F.; Hardouin, P. Effects of glucocorticoids on skeletal growth in rabbits evaluated by dual-photon absorptiometry, microscopic connectivity and vertebral compressive strength. Osteoporos. Int. 1994, 4, 204–210. [Google Scholar] [CrossRef]
- Ding, M.; Cheng, L.; Bollen, P.; Schwarz, P.; Overgaard, S. Glucocorticoid induced osteopenia in cancellous bone of sheep: Validation of large animal model for spine fusion and biomaterial research. Spine 2010, 35, 363–370. [Google Scholar] [CrossRef]
- Vega, S.L.; Kwon, M.Y.; Burdick, J.A. Recent advances in hydrogels for cartilage tissue engineering. Eur. Cell. Mater. 2017, 33, 59–75. [Google Scholar] [CrossRef]
- Yu, L.; Dawson, L.A.; Yan, M.; Zimmel, K.; Lin, Y.-L.; Dolan, C.P.; Han, M.; Muneoka, K. BMP9 stimulates joint regeneration at digit amputation wounds in mice. Nat. Commun. 2019, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Storer, M.A.; Mahmud, N.; Karamboulas, K.; Borrett, M.J.; Yuzwa, S.A.; Gont, A.; Androschuk, A.; Sefton, M.V.; Kaplan, D.R.; Miller, F.D. Acquisition of a Unique Mesenchymal Precursor-like Blastema State Underlies Successful Adult Mammalian Digit Tip Regeneration. Dev. Cell 2020, 52, 509–524.e9. [Google Scholar] [CrossRef]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef]
- Heinegård, D.; Saxne, T. The role of the cartilage matrix in osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 50–56. [Google Scholar] [CrossRef]
- Saito, T.; Fukai, A.; Mabuchi, A.; Ikeda, T.; Yano, F.; Ohba, S.; Nishida, N.; Akune, T.; Yoshimura, N.; Nakagawa, T.; et al. Transcriptional regulation of endochondral ossification by HIF-2alpha during skeletal growth and osteoarthritis development. Nat. Med. 2010, 16, 678–686. [Google Scholar] [CrossRef]
- St-Jacques, B.; Hammerschmidt, M.; McMahon, A.P. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999, 13, 2072–2086. [Google Scholar] [CrossRef] [Green Version]
- Prein, C.; Warmbold, N.; Farkas, Z.; Schieker, M.; Aszodi, A.; Clausen-Schaumann, H. Structural and mechanical properties of the proliferative zone of the developing murine growth plate cartilage assessed by atomic force microscopy. Matrix Biol. 2016, 50, 1–15. [Google Scholar] [CrossRef]
- Yao, Y.; Pei, F.; Kang, P. Selenium, iodine, and the relation with Kashin-Beck disease. Nutrition 2011, 27, 1095–1100. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Y.; Zeng, G.; Yang, S.; Yang, T.; Ma, M.; Song, W. Single-Cell Profiles of Age-Related Osteoarthritis Uncover Underlying Heterogeneity Associated With Disease Progression. Front. Mol. Biosci. 2021, 8, 748360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Younis, S.; Shi, H.; Hu, S.; Zia, A.; Wong, H.H.; Elliott, E.E.; Chang, T.; Bloom, M.S.; Zhang, W.; et al. RNA-seq characterization of histamine-releasing mast cells as potential therapeutic target of osteoarthritis. Clin. Immunol. 2022, 244, 109117. [Google Scholar] [CrossRef] [PubMed]
- Nanus, D.E.; Badoume, A.; Wijesinghe, S.N.; Halsey, A.M.; Hurley, P.; Ahmed, Z.; Botchu, R.; Davis, E.T.; Lindsay, M.A.; Jones, S.W. Synovial tissue from sites of joint pain in knee osteoarthritis patients exhibits a differential phenotype with distinct fibroblast subsets. EBioMedicine 2021, 72, 103618. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S.; Jin, P.; Shang, T.; Sun, R.; Lu, L.; Guo, K.; Liu, J.; Tong, Y.; Wang, J.; et al. Dual functions of microRNA-17 in maintaining cartilage homeostasis and protection against osteoarthritis. Nat. Commun. 2022, 13, 2447. [Google Scholar] [CrossRef]
- Sunkara, V.; Heinz, G.A.; Heinrich, F.F.; Durek, P.; Mobasheri, A.; Mashreghi, M.F.; Lang, A. Combining segmental bulk- and single-cell RNA-sequencing to define the chondrocyte gene expression signature in the murine knee joint. Osteoarthr. Cartil. 2021, 29, 905–914. [Google Scholar] [CrossRef]
- Knights, A.J.; Farrell, E.C.; Ellis, O.M.; Lammlin, L.; Junginger, L.M.; Rzeczycki, P.M.; Bergman, R.F.; Pervez, R.; Cruz, M.; Knight, E.; et al. Synovial fibroblasts assume distinct functional identities and secrete R-spondin 2 in osteoarthritis. Ann. Rheum. Dis. 2022. [Google Scholar] [CrossRef]
- Sebastian, A.; McCool, J.L.; Hum, N.R.; Murugesh, D.K.; Wilson, S.P.; Christiansen, B.A.; Loots, G.G. Single-Cell RNA-Seq Reveals Transcriptomic Heterogeneity and Post-Traumatic Osteoarthritis-Associated Early Molecular Changes in Mouse Articular Chondrocytes. Cells 2021, 10, 1462. [Google Scholar] [CrossRef]
- Sebastian, A.; Hum, N.R.; McCool, J.L.; Wilson, S.P.; Murugesh, D.K.; Martin, K.A.; Rios-Arce, N.D.; Amiri, B.; Christiansen, B.A.; Loots, G.G. Single-cell RNA-Seq reveals changes in immune landscape in post-traumatic osteoarthritis. Front. Immunol. 2022, 13, 938075. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Y.; Wu, R.; Ding, T.; Xue, H.; Gao, C.; Li, X.; Wang, C. New Insights From Single-Cell Sequencing Data: Synovial Fibroblasts and Synovial Macrophages in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 709178. [Google Scholar] [CrossRef]
- Hardt, U.; Carlberg, K.; Af Klint, E.; Sahlström, P.; Larsson, L.; van Vollenhoven, A.; Hernandez Machado, S.; Israelsson, L.; Amara, K.; Chemin, K.; et al. Integrated single cell and spatial transcriptomics reveal autoreactive differentiated B cells in joints of early rheumatoid arthritis. Sci. Rep. 2022, 12, 11876. [Google Scholar] [CrossRef]
- Argyriou, A.; Wadsworth, M.H.; Lendvai, A.; Christensen, S.M.; Hensvold, A.H.; Gerstner, C.; van Vollenhoven, A.; Kravarik, K.; Winkler, A.; Malmström, V.; et al. Single cell sequencing identifies clonally expanded synovial CD4+ TPH cells expressing GPR56 in rheumatoid arthritis. Nat. Commun. 2022, 13, 4046. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Y.; Jin, S.; Wang, M.; Jiao, Y.; Yang, B.; Lu, X.; Ji, X.; Fei, Y.; Yang, H.; et al. Single-cell sequencing of immune cells from anticitrullinated peptide antibody positive and negative rheumatoid arthritis. Nat. Commun. 2021, 12, 4977. [Google Scholar] [CrossRef]
- Gawel, D.R.; Serra-Musach, J.; Lilja, S.; Aagesen, J.; Arenas, A.; Asking, B.; Bengnér, M.; Björkander, J.; Biggs, S.; Ernerudh, J.; et al. A validated single-cell-based strategy to identify diagnostic and therapeutic targets in complex diseases. Genome Med. 2019, 11, 47. [Google Scholar] [CrossRef] [Green Version]
- Francisco, V.; Pino, J.; González-Gay, M.Á.; Lago, F.; Karppinen, J.; Tervonen, O.; Mobasheri, A.; Gualillo, O. A new immunometabolic perspective of intervertebral disc degeneration. Nat. Rev. Rheumatol. 2022, 18, 47–60. [Google Scholar] [CrossRef]
- Howell, K.; Chien, C.; Bell, R.; Laudier, D.; Tufa, S.F.; Keene, D.R.; Andarawis-Puri, N.; Huang, A.H. Novel model of tendon regeneration reveals distinct cell mechanisms underlying regenerative and fibrotic tendon healing. Sci. Rep. 2017, 7, 45238. [Google Scholar] [CrossRef] [Green Version]
- Riley, G. Tendinopathy--from basic science to treatment. Nat. Clin. Pract. Rheumatol. 2008, 4, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Forde, M.S.; Punnett, L.; Wegman, D.H. Prevalence of musculoskeletal disorders in union ironworkers. J. Occup. Environ. Hyg. 2005, 2, 203–212. [Google Scholar] [CrossRef]
- Bayer, M.L.; Järvinen, T.A.H. Basic muscle physiology in relation to hamstring injury and repair. In Prevention and Rehabilitation of Hamstring Injuries; Thorborg, K., Opar, D., Shield, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 31–63. ISBN 978-3-030-31637-2. [Google Scholar]
- Järvinen, T.A.H.; Järvinen, T.L.N.; Kääriäinen, M.; Aärimaa, V.; Vaittinen, S.; Kalimo, H.; Järvinen, M. Muscle injuries: Optimising recovery. Best Pract. Res. Clin. Rheumatol. 2007, 21, 317–331. [Google Scholar] [CrossRef]
- Quintero, A.J.; Wright, V.J.; Fu, F.H.; Huard, J. Stem cells for the treatment of skeletal muscle injury. Clin. Sports Med. 2009, 28, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Blau, H.M.; Cosgrove, B.D.; Ho, A.T.V. The central role of muscle stem cells in regenerative failure with aging. Nat. Med. 2015, 21, 854–862. [Google Scholar] [CrossRef]
- Wosczyna, M.N.; Rando, T.A. A muscle stem cell support group: Coordinated cellular responses in muscle regeneration. Dev. Cell 2018, 46, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabula Muris Consortium; Overall coordination; Logistical coordination; Organ collection and processing; Library preparation and sequencing; Computational data analysis; Cell type annotation; Writing group; Supplemental text writing group; Principal investigators. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 2018, 562, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, A.B.; Smith, G.R.; Raue, U.; Begue, G.; Minchev, K.; Ruf-Zamojski, F.; Nair, V.D.; Wang, X.; Zhou, L.; Zaslavsky, E.; et al. Single-cell transcriptional profiles in human skeletal muscle. Sci. Rep. 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentzinger, C.F.; Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Cellular dynamics in the muscle satellite cell niche. EMBO Rep. 2013, 14, 1062–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, D.; Goemans, N.; Takeda, S.; Mercuri, E.; Aartsma-Rus, A. Duchenne muscular dystrophy. Nat. Rev. Dis. Primers 2021, 7, 13. [Google Scholar] [CrossRef]
- Mercuri, E.; Bönnemann, C.G.; Muntoni, F. Muscular dystrophies. Lancet 2019, 394, 2025–2038. [Google Scholar] [CrossRef]
- Gao, X.; Fraulob, M.; Haïat, G. Biomechanical behaviours of the bone-implant interface: A review. J. R. Soc. Interface 2019, 16, 20190259. [Google Scholar] [CrossRef]
- Li, C.; Pisignano, D.; Zhao, Y.; Xue, J. Advances in medical applications of additive manufacturing. Engineering 2020, 6, 1222–1231. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, K. Review on titanium and titanium based alloys as biomaterials for orthopaedic applications. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 102, 844–862. [Google Scholar] [CrossRef]
- Vesprey, A.; Suh, E.S.; Göz Aytürk, D.; Yang, X.; Rogers, M.; Sosa, B.; Niu, Y.; Kalajzic, I.; Ivashkiv, L.B.; Bostrom, M.P.; et al. Tmem100- and Acta2-Lineage Cells Contribute to Implant Osseointegration in a Mouse Model. J. Bone Miner. Res. 2021, 36, 1000–1011. [Google Scholar] [CrossRef]
- Li, J.; Zhao, C.; Xu, Y.; Song, L.; Chen, Y.; Xu, Y.; Ma, Y.; Wang, S.; Xu, A.; He, F. Remodeling of the osteoimmune microenvironment after biomaterials implantation in murine tibia: Single-cell transcriptome analysis. Bioact. Mater. 2023, 22, 404–422. [Google Scholar] [CrossRef]
- Stuart, M.A.C.; Huck, W.T.S.; Genzer, J.; Müller, M.; Ober, C.; Stamm, M.; Sukhorukov, G.B.; Szleifer, I.; Tsukruk, V.V.; Urban, M.; et al. Emerging applications of stimuli-responsive polymer materials. Nat. Mater. 2010, 9, 101–113. [Google Scholar] [CrossRef]
- Wuisman, P.I.J.M.; Smit, T.H. Bioresorbable polymers: Heading for a new generation of spinal cages. Eur. Spine J. 2006, 15, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Konttinen, Y.T.; Pajarinen, J. Adverse reactions to metal-on-metal implants. Nat. Rev. Rheumatol. 2013, 9, 5–6. [Google Scholar] [CrossRef]
- Cherry, C.; Maestas, D.R.; Han, J.; Andorko, J.I.; Cahan, P.; Fertig, E.J.; Garmire, L.X.; Elisseeff, J.H. Computational reconstruction of the signalling networks surrounding implanted biomaterials from single-cell transcriptomics. Nat. Biomed. Eng. 2021, 5, 1228–1238. [Google Scholar] [CrossRef]
- Moore, E.M.; Maestas, D.R.; Cherry, C.C.; Garcia, J.A.; Comeau, H.Y.; Davenport Huyer, L.; Kelly, S.H.; Peña, A.N.; Blosser, R.L.; Rosson, G.D.; et al. Biomaterials direct functional B cell response in a material-specific manner. Sci. Adv. 2021, 7, eabj5830. [Google Scholar] [CrossRef]
- Robins, H. Immunosequencing: Applications of immune repertoire deep sequencing. Curr. Opin. Immunol. 2013, 25, 646–652. [Google Scholar] [CrossRef]
- Agarwal, R.; García, A.J. Biomaterial strategies for engineering implants for enhanced osseointegration and bone repair. Adv. Drug Deliv. Rev. 2015, 94, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Tian, J.; Li, C.; Chen, L.; Zhao, Y. A hydrogel spinal dural patch with potential anti-inflammatory, pain relieving and antibacterial effects. Bioact. Mater. 2022, 14, 389–401. [Google Scholar] [CrossRef]
- Petrigliano, F.A.; Liu, N.Q.; Lee, S.; Tassey, J.; Sarkar, A.; Lin, Y.; Li, L.; Yu, Y.; Geng, D.; Zhang, J.; et al. Long-term repair of porcine articular cartilage using cryopreservable, clinically compatible human embryonic stem cell-derived chondrocytes. NPJ Regen. Med. 2021, 6, 77. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-L.; Dicks, A.; Steward, N.; Tang, R.; Katz, D.B.; Choi, Y.-R.; Guilak, F. Single cell transcriptomic analysis of human pluripotent stem cell chondrogenesis. Nat. Commun. 2021, 12, 362. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Nakahata, A.; Yamada, N.; Yoshizawa, K.; Kato, T.M.; Iwasaki, M.; Zhao, C.; Kuroki, H.; Ikeya, M. Grafting of iPS cell-derived tenocytes promotes motor function recovery after Achilles tendon rupture. Nat. Commun. 2021, 12, 5012. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Amiri, K.I.; Burke, J.R.; Schmid, J.A.; Richmond, A. BMS-345541 targets inhibitor of kappaB kinase and induces apoptosis in melanoma: Involvement of nuclear factor kappaB and mitochondria pathways. Clin. Cancer Res. 2006, 12, 950–960. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.H.-C. Kartogenin induces cartilage-like tissue formation in tendon-bone junction. Bone Res. 2014, 2, 14008. [Google Scholar] [CrossRef] [Green Version]
- Corridoni, D.; Chapman, T.; Antanaviciute, A.; Satsangi, J.; Simmons, A. Inflammatory Bowel Disease Through the Lens of Single-cell RNA-seq Technologies. Inflamm. Bowel Dis. 2020, 26, 1658–1668. [Google Scholar] [CrossRef]
- Cusanovich, D.A.; Daza, R.; Adey, A.; Pliner, H.A.; Christiansen, L.; Gunderson, K.L.; Steemers, F.J.; Trapnell, C.; Shendure, J. Multiplex single cell profiling of chromatin accessibility by combinatorial cellular indexing. Science 2015, 348, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Buenrostro, J.D.; Wu, B.; Litzenburger, U.M.; Ruff, D.; Gonzales, M.L.; Snyder, M.P.; Chang, H.Y.; Greenleaf, W.J. Single-cell chromatin accessibility reveals principles of regulatory variation. Nature 2015, 523, 486–490. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Heidrich-O’Hare, E.; Chen, W.; Duerr, R.H. Comprehensive benchmarking of CITE-seq versus DOGMA-seq single cell multimodal omics. Genome Biol. 2022, 23, 135. [Google Scholar] [CrossRef]
- Peterson, V.M.; Zhang, K.X.; Kumar, N.; Wong, J.; Li, L.; Wilson, D.C.; Moore, R.; McClanahan, T.K.; Sadekova, S.; Klappenbach, J.A. Multiplexed quantification of proteins and transcripts in single cells. Nat. Biotechnol. 2017, 35, 936–939. [Google Scholar] [CrossRef]
- Cao, J.; Cusanovich, D.A.; Ramani, V.; Aghamirzaie, D.; Pliner, H.A.; Hill, A.J.; Daza, R.M.; McFaline-Figueroa, J.L.; Packer, J.S.; Christiansen, L.; et al. Joint profiling of chromatin accessibility and gene expression in thousands of single cells. Science 2018, 361, 1380–1385. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Lake, B.B.; Zhang, K. High-throughput sequencing of the transcriptome and chromatin accessibility in the same cell. Nat. Biotechnol. 2019, 37, 1452–1457. [Google Scholar] [CrossRef]
- Ma, S.; Zhang, B.; LaFave, L.M.; Earl, A.S.; Chiang, Z.; Hu, Y.; Ding, J.; Brack, A.; Kartha, V.K.; Tay, T.; et al. Chromatin Potential Identified by Shared Single-Cell Profiling of RNA and Chromatin. Cell 2020, 183, 1103–1116.e20. [Google Scholar] [CrossRef]
- Mimitou, E.P.; Lareau, C.A.; Chen, K.Y.; Zorzetto-Fernandes, A.L.; Hao, Y.; Takeshima, Y.; Luo, W.; Huang, T.-S.; Yeung, B.Z.; Papalexi, E.; et al. Scalable, multimodal profiling of chromatin accessibility, gene expression and protein levels in single cells. Nat. Biotechnol. 2021, 39, 1246–1258. [Google Scholar] [CrossRef]
- Swanson, E.; Lord, C.; Reading, J.; Heubeck, A.T.; Genge, P.C.; Thomson, Z.; Weiss, M.D.; Li, X.-J.; Savage, A.K.; Green, R.R.; et al. Simultaneous trimodal single-cell measurement of transcripts, epitopes, and chromatin accessibility using TEA-seq. eLife 2021, 10, e63632. [Google Scholar] [CrossRef]
- Deng, Y.; Bartosovic, M.; Kukanja, P.; Zhang, D.; Liu, Y.; Su, G.; Enninful, A.; Bai, Z.; Castelo-Branco, G.; Fan, R. Spatial-CUT&Tag: Spatially resolved chromatin modification profiling at the cellular level. Science 2022, 375, 681–686. [Google Scholar] [CrossRef]


{kind=link}
| Cell Type | Transcript Markers | Reference |
|---|---|---|
| MSC | ITGB1, VACAM1, THY-1, NT5E, ENG | Yang et al., 2021 [52] |
| Embryonic SSC | PDPN, CADM1 | He et al., 2021 [53] |
| Cranial neural crest cells | PDPN, CADM1 | He et al., 2021 [53] |
| Osteoblast | RUNX2, COLA1, SPP1, ANO5, CDH11 | Yang et al., 2021 [52], Wang et al., 2022 [54] |
| Chondrocyte | COL2A1, SOX9, COL10A1, Aggrecan, COMP, | Yang et al., 2021 [52], Gan et al., 2021 [55] |
| CPC | BIRC5, UBE2C, DHFR, CENPU, STMN1 | Yang et al., 2021 [52], Ji et al., 2019 [56] |
| Prehypertrophic/hypertrophic chondrocytes | COL10A1, IBSP | Chou et al., 2022 [57] |
| Osteoclast | NFKB-1, NFKB-2, ATP6V0D1, NFATC1, OSCAR, MMP9, MMP8 | Yang et al., 2021 [52], Wang et al., 2022 [54] |
| Tendocyte | COL1A1/2 | Kendal et al., 2020 [15] |
| MuSC | APOE, PAX7, MYF5, APOC | De Micheli et al., 2020 [58] |
| Mature skeletal muscle cells | TTN, MYLPF, CKM, TNNC, ACTA1 | De Micheli et al., 2020 [58] |
| Adipocytes | APOD, CXCL14 | De Micheli et al., 2020 [58] |
| Periosteal Osteogenic Progenitors | LEPR, PRRX1, GREM1 | Ding et al. 2022 [59] |
| Disease | Animal | Location | Phenotype Driving Gene/Materials in Mice Models | Important Affected Cell Types | Reference |
|---|---|---|---|---|---|
| Bone injury and regeneration | Mice | Rib | Smo | Cxcl12-expressing SSPCs | Serowoky et al., 2022 [64] |
| Mice | Frontal bones | p75 (Ngfr) | Itgb1-expressing mesenchymal, Il1a-, Il10-, and Tnf-expressing mesenchymal, and immune cells | Xu et al., 2022 [65] | |
| Mice | Long bone | - | Osteoblast lineage cells, chondrocytes, fibroblasts, Fabp5+ Mmp9+ septoclasts | Sivaraj et al., 2022 [66] | |
| Congenital Skeletal dysplasia | Mice | Hind limb | - | HC | Wang et al., 2022 [67] |
| Mice | Calvarium | TrkA | Mesenchymal progenitor cells | Tower et al., 2021 [19] | |
| Adolescent idiopathic scoliosis | Human | Spinal cancellous bone tissues | - | MSC-IGFBP5, CPC-PCNA, and OC-BIRC3 | Yang et al., 2021 [52] |
| Heterotopic ossification | Mice | Muscle | rhBMP2–Matrigel mixtures pre-immune antibody (referred to as BMP2/IgG) or neutralizing activin A antibody (BMP2/nActA.Ab) | Sox9-expressing skeletal progenitors, and Acan- and Col2a1-expressing clusters | Mundy et al., 2021 [68] |
| Mice | Achilles tendon | - | Prg4+ TSPC | Tachibana et al., 2022 [69] | |
| Osteoporosis | Human | Femoral head | - | Osteoclasts and immune cells | Wang et al., 2022 [54] |
| Disease | Animal | Location | Phenotype Driving Gene/Materials in Mice Models | Important Affected Cell Types | Reference |
|---|---|---|---|---|---|
| Chondrogenic regeneration | Mice | Digit tip | Ectopic BMP9 | Fibroblasts | Yu et al., 2022 [70] |
| Osteoarthritis | Human | Cartilage | - | ProC, FC, preHTC, and CPC | Ji et al., 2019 [56] |
| Human | Cartilage | - | FC, preFC, RegC, RepC, and preHTC | Chou et al., 2020 [57] | |
| Human | Cartilage | - | RegC | Wang et al., 2021 [71] | |
| Human | Cartilage | - | StrC (containing a ferropotic cluster), RegC | Lv et al., 2022 [72] | |
| Human | Synovitis | - | Immune cells and fibroblasts | Chou et al., 2020 [57] | |
| Kashin–Beck disease | Human | Cartilage | - | HomC and MTC | Wang et al., 2021 [71] |
| Rheumatoid arthritis | Human | Synovitis | - | CD55+ fibroblast and CD90+ fibroblast | Stephenson et al., 2018 [28] |
| Human | Synovitis | - | THY1(CD90)+HLA-DRAhi sublining fibroblasts | Zhang et al., 2019 [73] | |
| Human | Synovitis | - | Immune cells fibroblasts | Carlberg et al., 2019 [20] | |
| IVD degeneration | Human | IVD | - | CPC, HomC, and other chondrocyte subsets | Zhang et al., 2021 [74] |
| Human | IVD | - | IR NPC, FC NPC | Ling et al., 2021 [75] | |
| Human | IVD | - | - | Cherif et al.,2022 [76] | |
| Human | IVD | - | NP progenitor cells | Gan et al., 2021 [55] |
| Disease | Animal | Location | Phenotype Driving Gene/Materials in Mice Models | Important Affected Cell Types | Reference |
|---|---|---|---|---|---|
| Muscular dystrophy | Mice | Single-cell analysis: hind limb; further experiment: diaphragms | Mdx | Tie2high FAP and Vcam+ FAP | Malecova et al., 2018 [77] |
| Injury of muscle | Mice | Hind limb muscle | Notexin | MuSC, myogenic progenitors, FAP, and tendocytes | De Micheli et al., 2020 [78] |
| Mice | Hind limb muscle | Notexin | MuSC, PM | Dell’Orso et al., 2019 [79] | |
| Mice | Tibialis anterior (TA) muscles | Notexin | MuSC-expressing immune genes, immune cells, and activated FAP | Oprescu et al., 2020 [80] | |
| Mice | Patellar tendons | PDGF-AA protein | Tppp3+Pdgfra+ tendon stem cells. Tppp3−Pdgfra+ FAP | Harvey et al., 2019 [81] | |
| Injury of tendon | Mice | Diseased chilles, toe extensor or diseased peroneus longus | - | IL33-expressing endothelium and microfibril-associated tenocytes | Kendal et al., 2020 [15] |
| Tendinopathy | Human | Supraspinatus and subscapularis tendon | EGF and GAS pathway | T-cells | Garcia-Melchor et al., 2021 [82] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, S.; Li, J.; Tian, J.; Lu, S.; Zhao, Y. Application of Single-Cell and Spatial Omics in Musculoskeletal Disorder Research. Int. J. Mol. Sci. 2023, 24, 2271. https://doi.org/10.3390/ijms24032271
Feng S, Li J, Tian J, Lu S, Zhao Y. Application of Single-Cell and Spatial Omics in Musculoskeletal Disorder Research. International Journal of Molecular Sciences. 2023; 24(3):2271. https://doi.org/10.3390/ijms24032271
Chicago/Turabian StyleFeng, Site, Jiahao Li, Jingjing Tian, Sheng Lu, and Yu Zhao. 2023. "Application of Single-Cell and Spatial Omics in Musculoskeletal Disorder Research" International Journal of Molecular Sciences 24, no. 3: 2271. https://doi.org/10.3390/ijms24032271
APA StyleFeng, S., Li, J., Tian, J., Lu, S., & Zhao, Y. (2023). Application of Single-Cell and Spatial Omics in Musculoskeletal Disorder Research. International Journal of Molecular Sciences, 24(3), 2271. https://doi.org/10.3390/ijms24032271