
Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis
 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
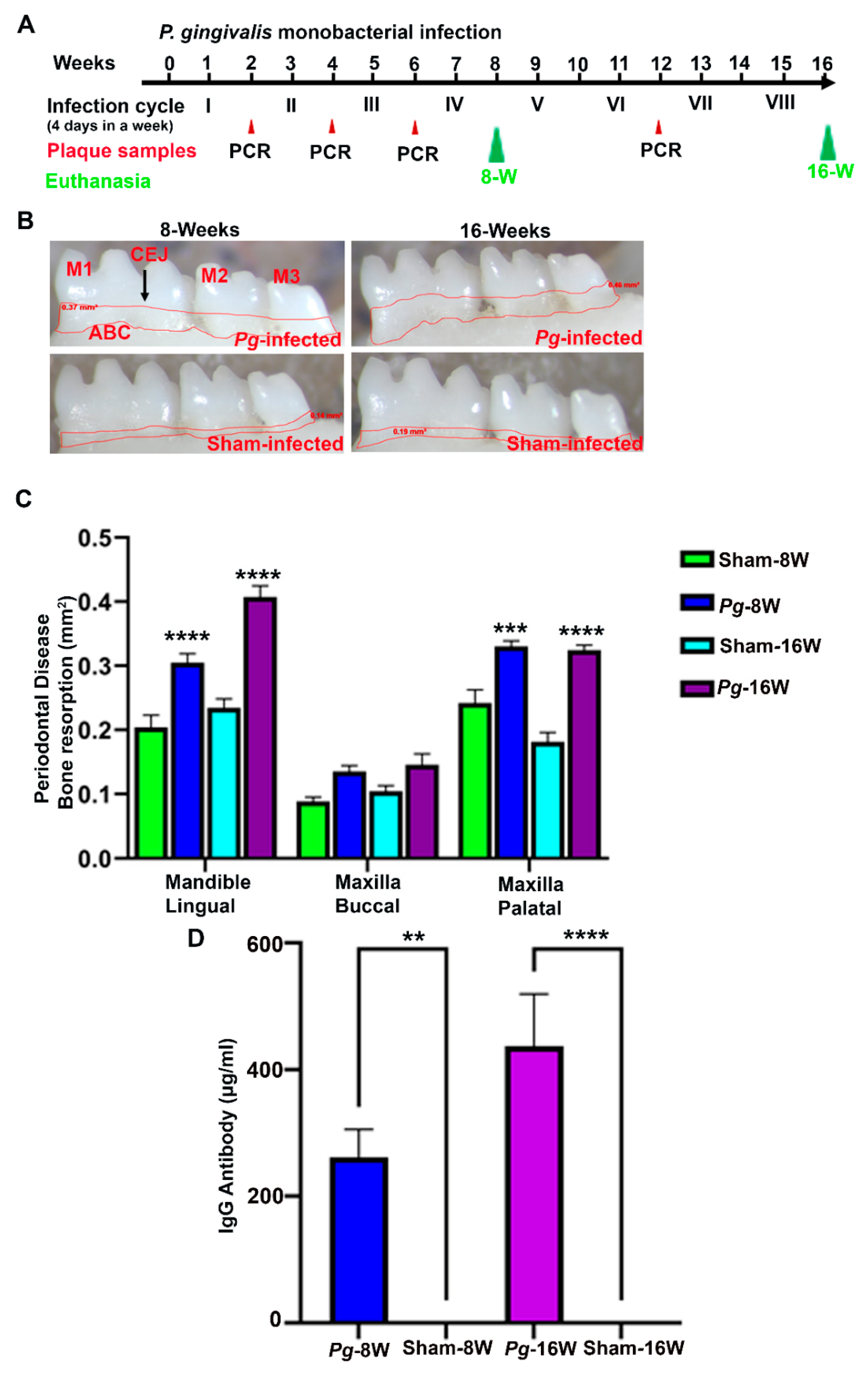
2.1. P. gingivalis Gingival Infection Showed Bacterial Colonization, Induced ABR, IgG Antibody Induction and Dissemination to Distal Organs
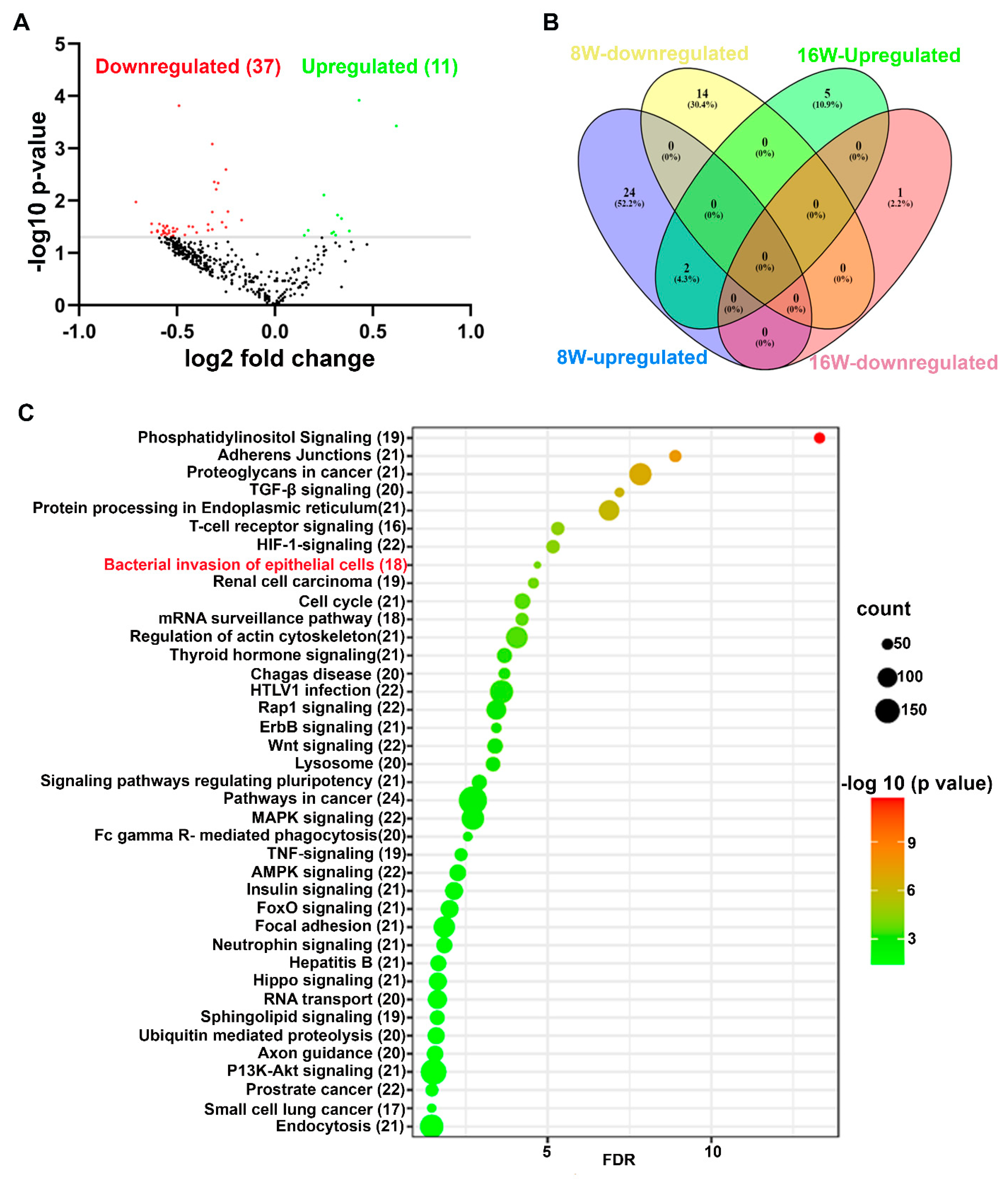
2.2. Unique and Common miRNAs in 8- and 16-Week Time-Points of P. gingivalis-Infected Mice Mandibles
2.3. Predicted Functional Pathway Analysis of DE miRNAs
3. Discussion
4. Materials and Methods
4.1. Topical Gingival Infection of P. gingivalis for the Induction of Periodontitis
4.2. Identification of P. gingivalis Genomic DNA in Gingival Plaques, Bacterial Dissemination and IgG Antibody Quantification
4.3. Mesaurement of Horizontal Alveolar Bone Resorption (ABR)
4.4. miRNA Expression Kinetics Using NanoString nCounter miRNA Panels
4.5. NanoString Data Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nahid, M.A.; Rivera, M.; Lucas, A.; Chan, E.K.; Kesavalu, L. Polymicrobial infection with periodontal pathogens specifically enhances microRNA miR-146a in ApoE-/- mice during experimental periodontal disease. Infect. Immun. 2011, 79, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravindraja, C.; Kashef, M.R.; Vekariya, K.M.; Ghanta, R.K.; Karanth, S.; Chan, E.K.L.; Kesavalu, L. Global Noncoding microRNA Profiling in Mice Infected with Partial Human Mouth Microbes (PAHMM) Using an Ecological Time-Sequential Polybacterial Periodontal Infection (ETSPPI) Model Reveals Sex-Specific Differential microRNA Expression. Int. J. Mol. Sci. 2022, 23, 5107. [Google Scholar] [CrossRef] [PubMed]
- Kebschull, M.; Papapanou, P.N. Mini but mighty: microRNAs in the pathobiology of periodontal disease. Periodontol 2000 2015, 69, 201–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irwandi, R.A.; Vacharaksa, A. The role of microRNA in periodontal tissue: A review of the literature. Arch. Oral Biol. 2016, 72, 66–74. [Google Scholar] [CrossRef]
- Olsen, I.; Singhrao, S.K.; Osmundsen, H. Periodontitis, pathogenesis and progression: miRNA-mediated cellular responses to Porphyromonas gingivalis. J. Oral Microbiol. 2017, 9, 1333396. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.R.; Zhong, S.; Dang, H.; Fordham, J.B.; Nares, S.; Khan, A. Expression Profiling of LPS Responsive miRNA in Primary Human Macrophages. J. Microb. Biochem. Technol. 2016, 8, 136–143. [Google Scholar] [CrossRef]
- Aravindraja, C.; Sakthivel, R.; Liu, X.; Goodwin, M.; Veena, P.; Godovikova, V.; Fenno, J.C.; Levites, Y.; Golde, T.E.; Kesavalu, L. Intracerebral but Not Peripheral Infection of Live Porphyromonas gingivalis Exacerbates Alzheimer’s Disease like Amyloid Pathology in APP-TgCRND8 Mice. Int. J. Mol. Sci. 2022, 23, 3328. [Google Scholar] [CrossRef]
- Chukkapalli, S.; Rivera-Kweh, M.; Gehlot, P.; Velsko, I.; Bhattacharyya, I.; Calise, S.J.; Satoh, M.; Chan, E.K.; Holoshitz, J.; Kesavalu, L. Periodontal bacterial colonization in synovial tissues exacerbates collagen-induced arthritis in B10.RIII mice. Arthritis Res. Ther. 2016, 18, 161. [Google Scholar] [CrossRef] [Green Version]
- Velsko, I.M.; Chukkapalli, S.S.; Rivera, M.F.; Lee, J.Y.; Chen, H.; Zheng, D.; Bhattacharyya, I.; Gangula, P.R.; Lucas, A.R.; Kesavalu, L. Active invasion of oral and aortic tissues by Porphyromonas gingivalis in mice causally links periodontitis and atherosclerosis. PLoS ONE 2014, 9, e97811. [Google Scholar] [CrossRef] [Green Version]
- Menikdiwela, K.R.; Ramalingam, L.; Abbas, M.M.; Bensmail, H.; Scoggin, S.; Kalupahana, N.S.; Palat, A.; Gunaratne, P.; Moustaid-Moussa, N. Role of microRNA 690 in Mediating Angiotensin II Effects on Inflammation and Endoplasmic Reticulum Stress. Cells 2020, 9, 1327. [Google Scholar] [CrossRef]
- Feng, Y.; Li, Y.; Zhang, Y.; Zhang, B.H.; Zhao, H.; Zhao, X.; Shi, F.D.; Jin, W.N.; Zhang, X.A. miR-1224 contributes to ischemic stroke-mediated natural killer cell dysfunction by targeting Sp1 signaling. J. Neuroinflammation 2021, 18, 133. [Google Scholar] [CrossRef]
- Yu, T.; Ma, P.; Wu, D.; Shu, Y.; Gao, W. Functions and mechanisms of microRNA-31 in human cancers. Biomed. Pharmacother. 2018, 108, 1162–1169. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, P.J.; Li, C.H.; Lv, Z.M.; Zhang, W.W.; Jin, C.H. miRNA-133 augments coelomocyte phagocytosis in bacteria-challenged Apostichopus japonicus via targeting the TLR component of IRAK-1 in vitro and in vivo. Sci. Rep. 2015, 5, 12608. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhong, W.; Zhu, M.; Li, M.; Yang, Z. miR-185 inhibits prostate cancer angiogenesis induced by the nodal/ALK4 pathway. BMC Urol. 2020, 20, 49. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, Y.C.; Chen, Y.; Xiang, Y.; Shen, C.X.; Li, Y.G. The role of miR-19b in the inhibition of endothelial cell apoptosis and its relationship with coronary artery disease. Sci. Rep. 2015, 5, 15132. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Zhan, H.; Zhou, X.Y.; Yao, L.; Yan, M.; Chen, A.; Liu, J.; Ren, X.; Zhang, X.; Liu, J.X.; et al. MicroRNA-22 regulates inflammation and angiogenesis via targeting VE-cadherin. FEBS Lett. 2017, 591, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Alexan, B.; Dennis, D.; Bettina, C.; Christoph, L.I.M.; Tang, Y. microRNA-193-3p attenuates myocardial injury of mice with sepsis via STAT3/HMGB1 axis. J. Transl. Med. 2021, 19, 386. [Google Scholar] [CrossRef]
- Anand, D.; Al Saai, S.; Shrestha, S.K.; Barnum, C.E.; Chuma, S.; Lachke, S.A. Genome-Wide Analysis of Differentially Expressed miRNAs and Their Associated Regulatory Networks in Lenses Deficient for the Congenital Cataract-Linked Tudor Domain Containing Protein TDRD7. Front. Cell Dev. Biol. 2021, 9, 615761. [Google Scholar] [CrossRef]
- Kim, M.; Jo, H.; Kwon, Y.; Jeong, M.S.; Jung, H.S.; Kim, Y.; Jeoung, D. MiR-154-5p-MCP1 Axis Regulates Allergic Inflammation by Mediating Cellular Interactions. Front. Immunol. 2021, 12, 663726. [Google Scholar] [CrossRef]
- Friedrich, M.; Pracht, K.; Mashreghi, M.F.; Jäck, H.M.; Radbruch, A.; Seliger, B. The role of the miR-148/-152 family in physiology and disease. Eur. J. Immunol. 2017, 47, 2026–2038. [Google Scholar] [CrossRef] [Green Version]
- Dai, T.; Zhao, X.; Li, Y.; Yu, L.; Li, Y.; Zhou, X.; Gong, Q. miR-423 Promotes Breast Cancer Invasion by Activating NF-kappaB Signaling. OncoTargets Ther. 2020, 13, 5467–5478. [Google Scholar] [CrossRef] [PubMed]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Bock, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating miR-125a-5p, miR-125b-5p, and miR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef] [PubMed]
- Du, K.; Zhao, C.; Wang, L.; Wang, Y.; Zhang, K.Z.; Shen, X.Y.; Sun, H.X.; Gao, W.; Lu, X. MiR-191 inhibit angiogenesis after acute ischemic stroke targeting VEZF1. Aging 2019, 11, 2762–2786. [Google Scholar] [CrossRef] [PubMed]
- Foley, N.H.; O’Neill, L.A. miR-107: A toll-like receptor-regulated miRNA dysregulated in obesity and type II diabetes. J. Leukoc. Biol. 2012, 92, 521–527. [Google Scholar] [CrossRef]
- Jiang, L.; Qiao, Y.; Wang, Z.; Ma, X.; Wang, H.; Li, J. Inhibition of microRNA-103 attenuates inflammation and endoplasmic reticulum stress in atherosclerosis through disrupting the PTEN-mediated MAPK signaling. J. Cell. Physiol. 2020, 235, 380–393. [Google Scholar] [CrossRef]
- Jang, J.; Song, G.; Pettit, S.M.; Li, Q.; Song, X.; Cai, C.L.; Kaushal, S.; Li, D. Epicardial HDAC3 Promotes Myocardial Growth Through a Novel MicroRNA Pathway. Circ. Res. 2022, 131, 151–164. [Google Scholar] [CrossRef]
- Li, X.; Du, N.; Zhang, Q.; Li, J.; Chen, X.; Liu, X.; Hu, Y.; Qin, W.; Shen, N.; Xu, C.; et al. MicroRNA-30d regulates cardiomyocyte pyroptosis by directly targeting foxo3a in diabetic cardiomyopathy. Cell Death Dis. 2014, 5, e1479. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Lv, J. MicroRNA-301a-3p increases oxidative stress, inflammation and apoptosis in ox-LDL-induced HUVECs by targeting KLF7. Exp. Ther. Med. 2021, 21, 569. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, K.; Hassan, S.H.; Zhang, X.; Tang, X.; Pu, H.; Stetler, R.A.; Chen, J.; Yin, K.J. Endothelium-Targeted Deletion of microRNA-15a/16-1 Promotes Poststroke Angiogenesis and Improves Long-Term Neurological Recovery. Circ. Res. 2020, 126, 1040–1057. [Google Scholar] [CrossRef]
- Ceolotto, G.; Giannella, A.; Albiero, M.; Kuppusamy, M.; Radu, C.; Simioni, P.; Garlaschelli, K.; Baragetti, A.; Catapano, A.L.; Iori, E.; et al. miR-30c-5p regulates macrophage-mediated inflammation and pro-atherosclerosis pathways. Cardiovasc. Res. 2017, 113, 1627–1638. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Xu, N.; Jin, S. miR-30e-5p attenuates neuronal deficit and inflammation of rats with intracerebral hemorrhage by regulating TLR4. Exp. Ther. Med. 2022, 24, 492. [Google Scholar] [CrossRef]
- Krell, J.; Frampton, A.E.; Jacob, J.; Pellegrino, L.; Roca-Alonso, L.; Zeloof, D.; Alifrangis, C.; Lewis, J.S.; Jiao, L.R.; Stebbing, J.; et al. The clinico-pathologic role of microRNAs miR-9 and miR-151-5p in breast cancer metastasis. Mol. Diagn. Ther. 2012, 16, 167–172. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, Z.; Mijiti, M.; Du, G.; Li, Y.; Dangmurenjiafu, G. MiR-28-5p promotes human glioblastoma cell growth through inactivation of FOXO1. Int. J. Clin. Exp. Pathol. 2019, 12, 2972–2980. [Google Scholar]
- An, L.; Ji, D.; Hu, W.; Wang, J.; Jin, X.; Qu, Y.; Zhang, N. Interference of Hsa_circ_0003928 Alleviates High Glucose-Induced Cell Apoptosis and Inflammation in HK-2 Cells via miR-151-3p/Anxa2. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 3157–3168. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Fan, W.; Lou, W. hsa_circ_0001955 Enhances In Vitro Proliferation, Migration, and Invasion of HCC Cells through miR-145-5p/NRAS Axis. Mol. Ther. Nucleic Acids 2020, 22, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, S.; Sun, M.; Zhang, C.; Wei, C.; Yang, C.; Dou, R.; Liu, Q.; Xiong, B. miR-195-5p/NOTCH2-mediated EMT modulates IL-4 secretion in colorectal cancer to affect M2-like TAM polarization. J. Hematol. Oncol. 2019, 12, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, T.; Lu, Y.; He, Q.X.; Wang, H.; Li, B.F.; Zhu, L.Y.; Xu, Q.Y. The Role of MicroRNA, miR-24, and Its Target CHI3L1 in Osteomyelitis Caused by Staphylococcus aureus. J. Cell. Biochem. 2015, 116, 2804–2813. [Google Scholar] [CrossRef]
- Wu, W.; Xuan, Y.; Ge, Y.; Mu, S.; Hu, C.; Fan, R. Plasma miR-146a and miR-365 expression and inflammatory factors in patients with osteoarthritis. Malays. J. Pathol. 2021, 43, 311–317. [Google Scholar]
- Lockhart, P.B.; Bolger, A.F.; Papapanou, P.N.; Osinbowale, O.; Trevisan, M.; Levison, M.E.; Taubert, K.A.; Newburger, J.W.; Gornik, H.L.; Gewitz, M.H.; et al. Periodontal disease and atherosclerotic vascular disease: Does the evidence support an independent association?: A scientific statement from the American Heart Association. Circulation 2012, 125, 2520–2544. [Google Scholar] [CrossRef]
- Chukkapalli, S.S.; Easwaran, M.; Rivera-Kweh, M.F.; Velsko, I.M.; Ambadapadi, S.; Dai, J.; Larjava, H.; Lucas, A.R.; Kesavalu, L. Sequential colonization of periodontal pathogens in induction of periodontal disease and atherosclerosis in LDLRnull mice. Pathog. Dis. 2017, 75, ftx003. [Google Scholar] [CrossRef] [Green Version]
- Luan, X.; Zhou, X.; Trombetta-eSilva, J.; Francis, M.; Gaharwar, A.K.; Atsawasuwan, P.; Diekwisch, T.G.H. MicroRNAs and Periodontal Homeostasis. J. Dent. Res. 2017, 96, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Motedayyen, H.; Ghotloo, S.; Saffari, M.; Sattari, M.; Amid, R. Evaluation of MicroRNA-146a and Its Targets in Gingival Tissues of Patients With Chronic Periodontitis. J. Periodontol. 2015, 86, 1380–1385. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group/Bacteria/Infection (Weeks) | Positive Gingival Plaque Samples (n = 10) a | |||
|---|---|---|---|---|
| 2 Weeks | 4 Weeks | 6 Weeks | 12 Weeks | |
| Group I/ P. gingivalis FDC 381 [8 weeks] | 8/10 a | 10/10 | NC | --- |
| Group II/ P. gingivalis FDC 381 [16 weeks] | 10/10 | 10/10 | NC | 10/10 |
| Group III/ Sham-infection [8 weeks] | 0/10 | NC | NC | --- |
| Group IV/ Sham-infection [16 weeks] | 0/10 | NC | NC | 0/10 |
| Weeks/Infection/Sex | Upregulated miRNAs (p < 0.05) | Downregulated miRNAs (p < 0.05) |
|---|---|---|
| 8W- P. gingivalis infected Vs. 8W- Sham infection (n = 10) | 26 (miR-103, miR-30d) | 14 |
| 8W- P. gingivalis infected Female Vs Male (n = 5) | 3 | 2 |
| 16W- P. gingivalis infected Vs. 16W- Sham infection (n = 10) | 7 (miR-103, miR-30d) | 1 |
| 16W- P. gingivalis infected Female Vs Male (n = 5) | 39 | 9 |
| 8W- P. gingivalis infected Vs. 16W- P. gingivalis infected | 11 | 37 |
| Upregulated miRNAs in 8-Weeks ofP. gingivalisInfection | ||||
| miRNAs | Fold Change | p-Value | Reported Functions | Number of Target Genes |
| miR-804 | 1.57 | 0.009465 | Not identified | -- |
| miR-690 | 1.53 | 0.007031 | Inflammation and endoplasmic reticulum stress in obese animal models [10]. | 157 (Srrm2, Fkbp1a, Glu1, Brd8, Cyb5r1, Slc16a12, Cdip1, Tob, Megf9, Hc) |
| miR-1224 | 1.4 | 0.013375 | Ischemic stroke by acting as negative regulator of Natural Killer cells [11]. | -- |
| miR-31 | 1.37 | 0.034075 | Tumor suppressor in many cancers such as Ovarian, HCC, Prostate, Glioma, and Breast Cancer [12]. | 1176 (Mul1, Nipsnap3b, Cyc1, Srrm2, Fubp1, Wipi2, Gcnt4, Kat6b, Arid3a, Bace1) |
| miR-133b | 1.36 | 0.03893 | Regulates TLR component of IRAK-1 and promotes phagocytosis [13] | 1430 (Med13, Rrp8, Cnp, Srrm2, Wipi2, Sh3pxd2b, Dlc1, Tgfb2, Rsbn1l, Kat6b) |
| miR-185 | 1.32 | 0.000534 | Regulates angiogenesis in Prostate Cancer [14] | 332 |
| miR-19b | 1.31 | 0.000122 | Attenuates TNF-induced endothelial cell apoptosis in coronary heart disease [15] | 1901 |
| miR-22 | 1.31 | 0.009126 | Regulates endothelial inflammation, tissue injury and angiogenesis [16]. | 2150 |
| miR-193 | 1.29 | 0.006863 | Attenuates myocardial injury of mice with sepsis [17]. | 0 |
| miR-1198 | 1.27 | 0.001943 | Reported to be downregulated in Juvenile cataracts mouse model [18]. | 3 |
| miR-154 | 1.27 | 0.04589 | Mediates allergic reactions that include passive cutaneous and systemic anaphylaxis [19]. | 541 |
| miR-152 | 1.26 | 0.031135 | Inhibition of cell proliferation and progression in breast cancer, glioma and gastric cancer [20]. | 1115 |
| miR-423-3p | 1.26 | 0.044217 | Breast cancer invasion by activating NF-κB signaling pathway [21]. | 4 |
| miR-125b-5p | 1.24 | 0.02166 | Associated with acute Ischemic stroke [22]. | 1624 |
| miR-191 | 1.23 | 0.013163 | Promotes ischemic brain injury by inhibiting angiogenesis [23]. | 129 |
| miR-107 | 1.22 | 0.022686 | Insulin resistance in Type II Diabetes, Inflammation and Obesity [24]. | 311 |
| miR-103 | 1.21 | 0.004192 | Highly expressed in ApoE-/- mice and linked with atherosclerosis [25]. | 2 |
| miR-322 | 1.19 | 0.019203 | Over expressed in Hdac3 KO mouse epicardial cells and hearts [26]. | 2761 |
| miR-30d | 1.17 | 0.042862 | Promotes cardiomyocyte pyroptosis in diabetic cardiomyopathy [27]. | 10 |
| miR-301a | 1.15 | 0.041072 | Promotes oxidative stress, inflammation and apoptosis in arteriosclerotic cardiovascular disease [28]. | 1220 |
| miR-15a | 1.15 | 0.046356 | Negative regulator of postischemic cerebral angiogenesis and long-term neurological recovery [29]. | 1368 |
| miR-30c | 1.15 | 0.049249 | Regulate macrophage mediated inflammation and pro-atherosclerosis pathways [30]. | 1297 |
| miR-30e | 1.13 | 0.00518 | Protective role against neuronal deficit and inflammation in intracerebral hemorrhage [31]. | 1324 |
| miR-151-5p | 1.13 | 0.02108 | Breast cancer metastasis [32] | 113 |
| miR-28 | 1.13 | 0.041907 | Oncogene that promotes human glioblastoma cell growth [33]. | 0 |
| miR-151-3p | 1.11 | 0.030216 | Regulates inflammation and apoptosis [34]. | 81 |
| Upregulated miRNAs in 16-Weeks of P. gingivalis Infection | ||||
| miRNAs | Fold Change | p-Value | Reported Functions | Number of Target Genes |
| miR-30d | 1.11 | 0.046212 | Promotes cardiomyocyte pyroptosis in diabetic cardiomyopathy [27]. | 10 Pik3r1, Ctnna1, Cblb, Sept8, Was1, Sept2, Rac1, Crk, Itgb1, Pik3cd |
| miR-103 | 1.13 | 0.044262 | Highly expressed in ApoE-/- mice and linked with atherosclerosis [25]. | 2 (Cltc, Rac1) |
| miR-145 | 1.14 | 0.018223 | Oncogene that enhances migration and invasion of Hepatocellular carcinoma [35]. | 0 |
| miR-195 | 1.18 | 0.028636 | Modulates cell proliferation in colorectal cancers [36]. | 0 |
| miR-24 | 1.19 | 0.020169 | Overexpression of miR-24 reduced the effect of S. aureus in osteomyelitis patients [37]. | 359 (Srrm2, Rsbn1l, Axin1, Sla, Bc030336, Mef2, Fyco1, Wipf1, Kidins220, Mhc) |
| miR-365 | 1.22 | 0.045178 | Highly expressed in patients with Osteoarthritis [38]. | 0 |
| miR-99b | 1.24 | 0.001738 | Predicts clinical outcome of osteosarcoma. | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aravindraja, C.; Vekariya, K.M.; Botello-Escalante, R.; Rahaman, S.O.; Chan, E.K.L.; Kesavalu, L. Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis. Int. J. Mol. Sci. 2023, 24, 2327. https://doi.org/10.3390/ijms24032327
Aravindraja C, Vekariya KM, Botello-Escalante R, Rahaman SO, Chan EKL, Kesavalu L. Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis. International Journal of Molecular Sciences. 2023; 24(3):2327. https://doi.org/10.3390/ijms24032327
Chicago/Turabian StyleAravindraja, Chairmandurai, Krishna Mukesh Vekariya, Ruben Botello-Escalante, Shaik O. Rahaman, Edward K. L. Chan, and Lakshmyya Kesavalu. 2023. "Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis" International Journal of Molecular Sciences 24, no. 3: 2327. https://doi.org/10.3390/ijms24032327
APA StyleAravindraja, C., Vekariya, K. M., Botello-Escalante, R., Rahaman, S. O., Chan, E. K. L., & Kesavalu, L. (2023). Specific microRNA Signature Kinetics in Porphyromonas gingivalis-Induced Periodontitis. International Journal of Molecular Sciences, 24(3), 2327. https://doi.org/10.3390/ijms24032327