
Construction of Biocatalysts Using the P450 Scaffold for the Synthesis of Indigo from Indole


Abstract
:1. Introduction
2. Results and Discussion
2.1. Protein Expression and Characterization
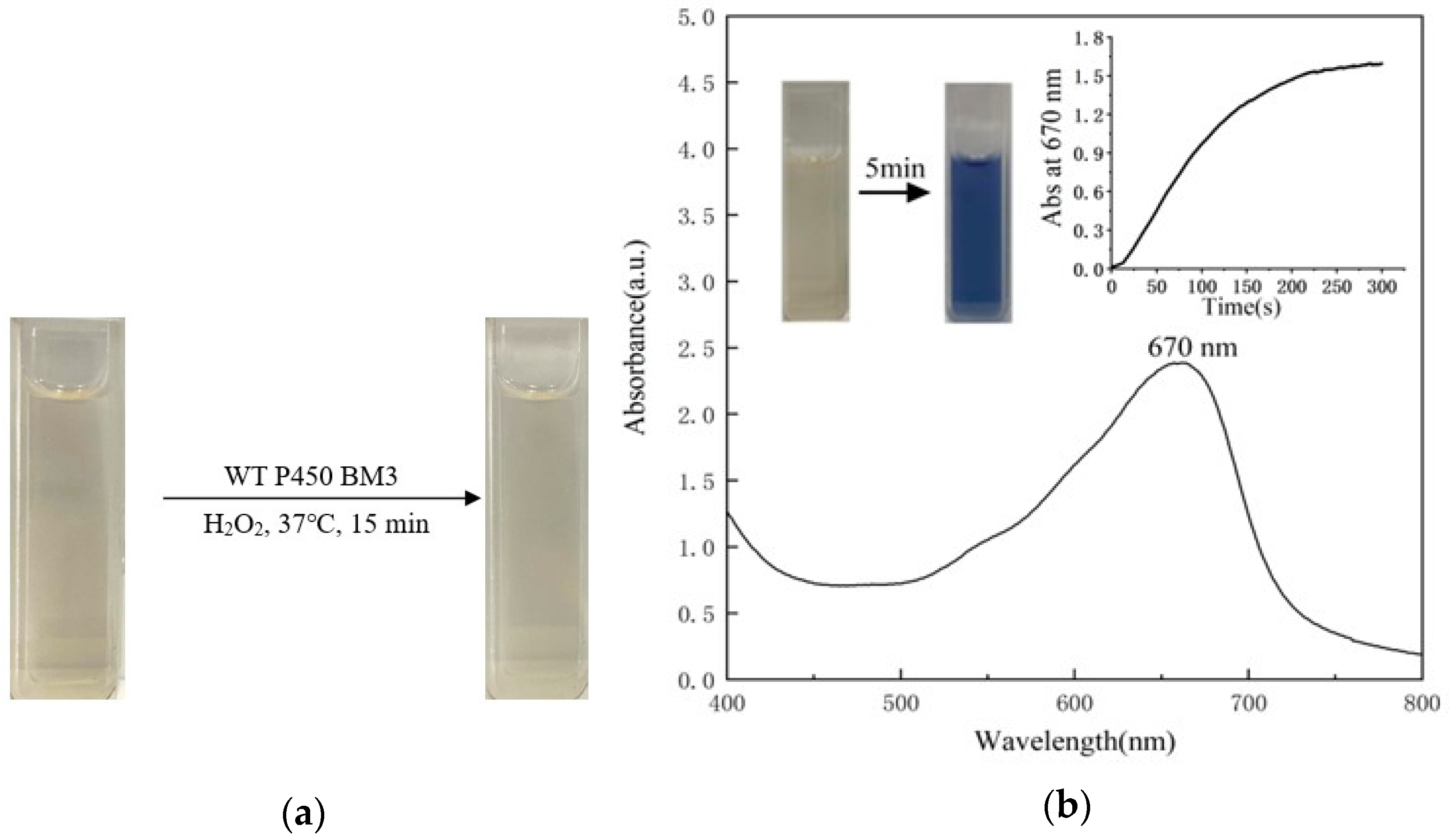
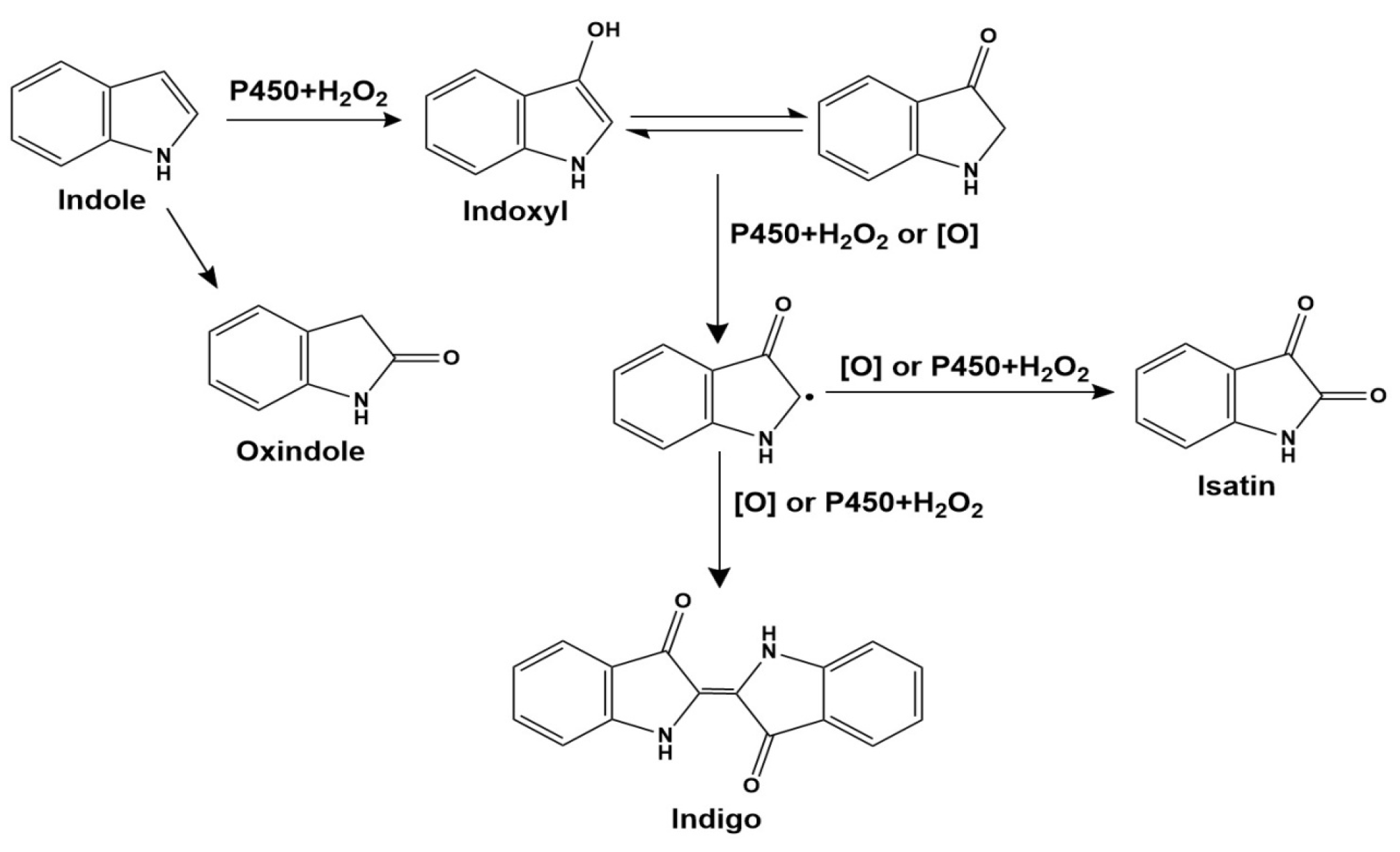
2.2. Indole Oxidation Catalyzed by the P450BM3 Peroxygenase System
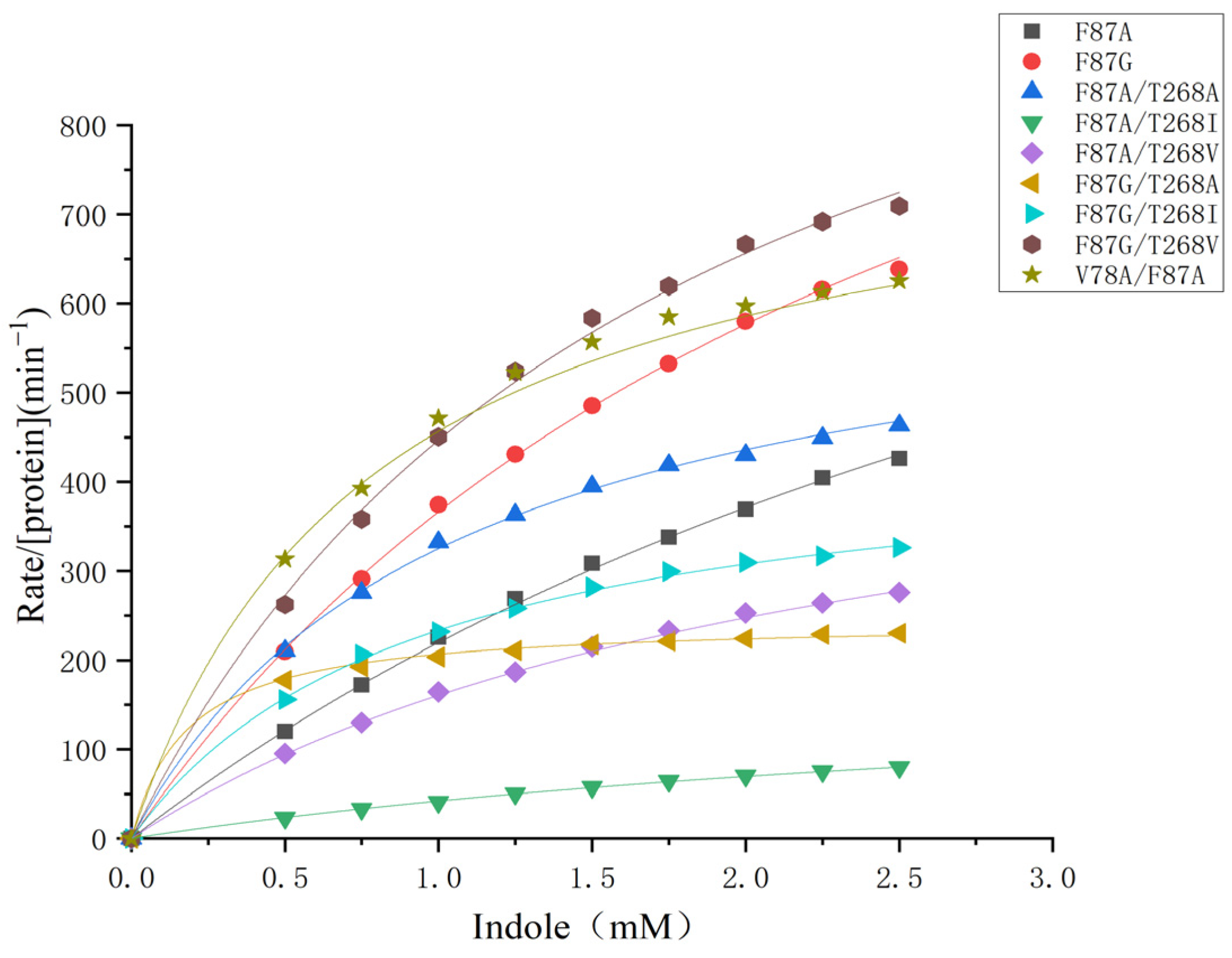
2.3. Kinetic Study on the Indigo Formation and the Site-Directed Mutagenesis
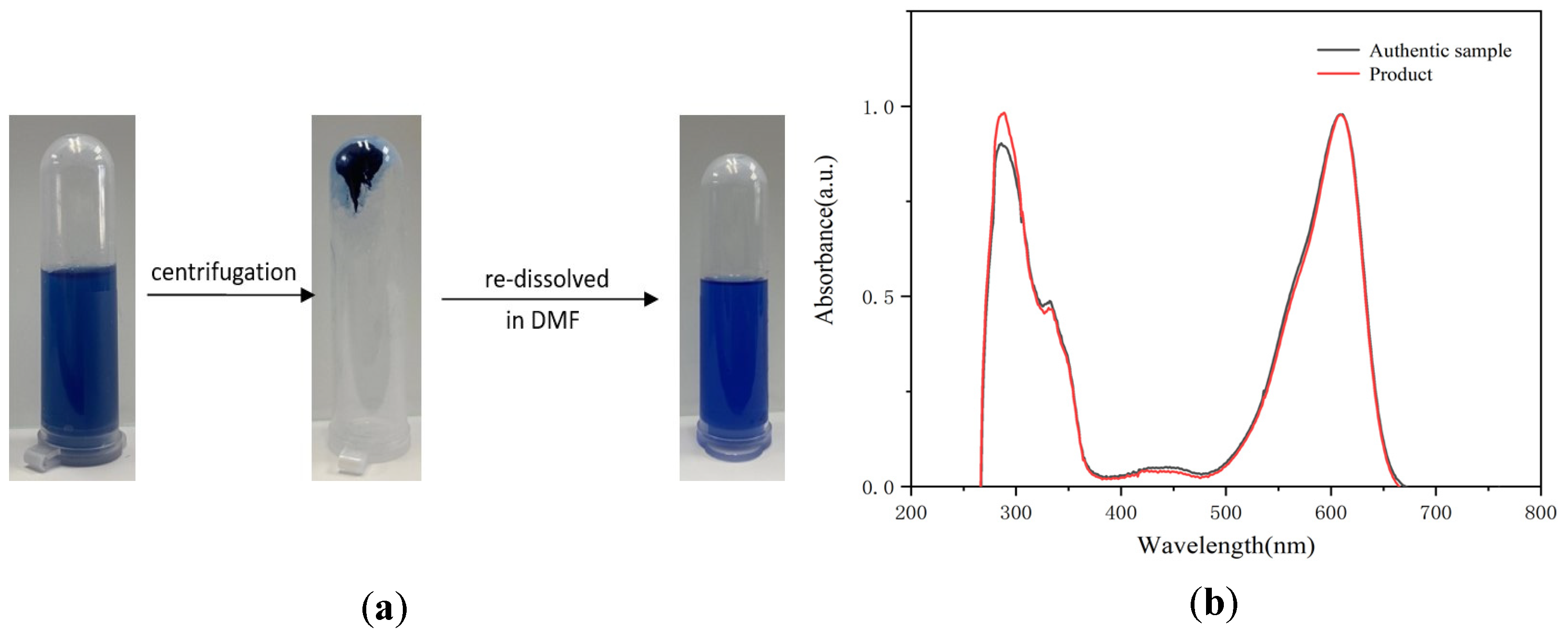
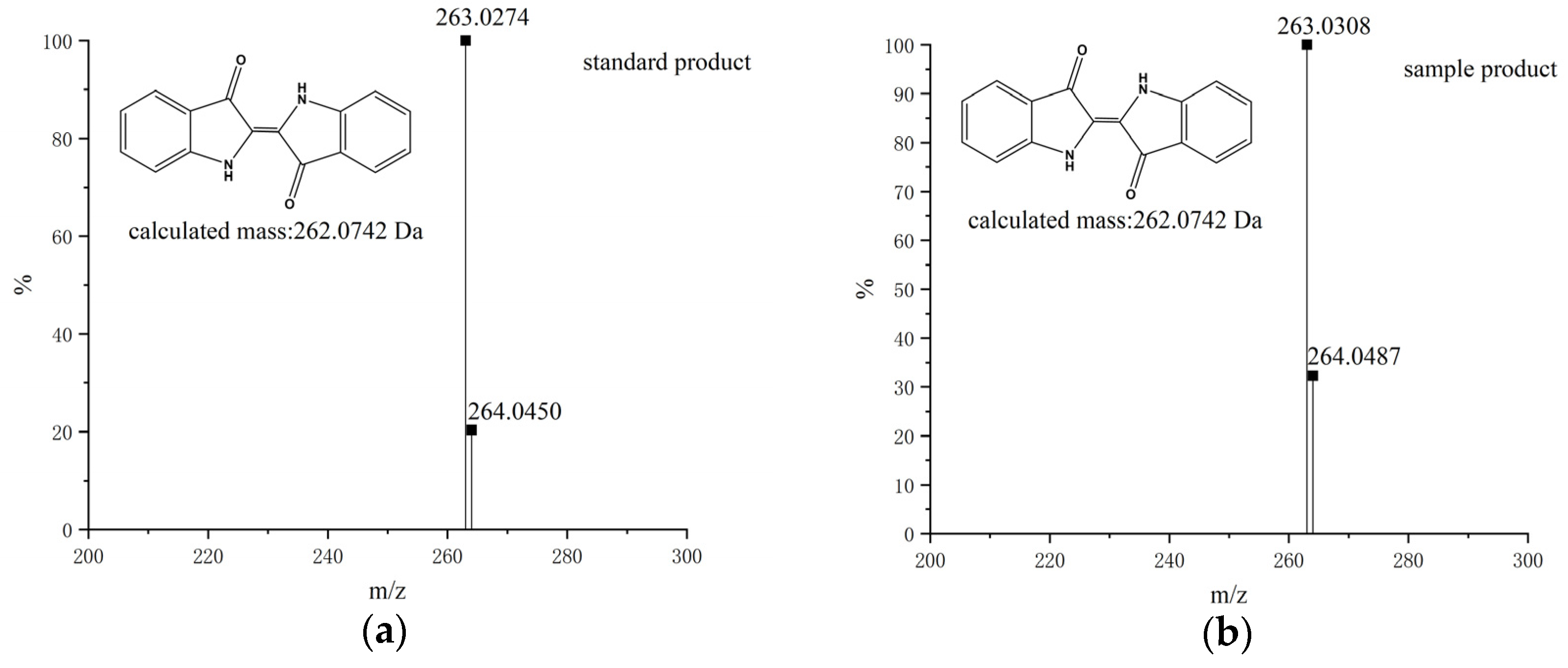
2.4. Analysis of the Mixture after the Oxidation Reaction
2.5. Molecular Modeling Simulation
3. Materials and Methods
3.1. Chemicals
3.2. Instrument
3.3. Site-Directed Mutagenesis, Cultivation, and Purification of P450BM3 Mutants
3.4. Catalytic Activity Assay
3.5. Analysis of the Reaction Mixture after the Oxidation
3.6. Isolation and Characterization of Indigo
3.7. Molecular Modeling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, G.W. Natural dyes in textile applications. Color. Technol. 1986, 16, 53–61. [Google Scholar] [CrossRef]
- Ferreira, E.S.B.; Hulme, A.N.; McNab, H.; Quye, A. The natural constituents of historical textile dyes. Chem. Soc. Rev. 2004, 33, 329–336. [Google Scholar] [CrossRef]
- Gilbert, K.G.; Maule, H.G.; Rudolph, B.; Lewis, M.; Vandenburg, H.; Sales, E.; Tozzi, S.; Cooke, D.T. Quantitative analysis of indigo and indigo precursors in leaves of Isatis spp. and Polygonum tinctorium. Biotechnol. Prog. 2010, 20, 1289–1292. [Google Scholar] [CrossRef]
- Cotson, S.; Holt, S.J. Studies in enzyme cytochemistry. IV. kinetics of aerial oxidation of indoxyl and some of its halogen derivatives. Proc. R. Soc. Biol. Sci. 1958, 148, 506–519. [Google Scholar]
- Ziegler, E.; Kappe, T. A new synthesis of indigo. Angew. Chem. (Int. Ed.) 1964, 3, 754. [Google Scholar] [CrossRef]
- Baeyer, A.D.A.; Drewsen, V.B.a. Darstellung von Indigblau aus Orthonitrobenzaldehyd. Ber. Der Dtsch. Chem. Ges. 1882, 15, 2856–2864. [Google Scholar] [CrossRef] [Green Version]
- Ensley, B.D.; Gibson, D.T.; Laborde, A.L. Oxidation of naphthalene by a multicomponent enzyme system from Pseudomonas sp. strain NCIB 9816. J. Bacteriol. 1982, 149, 948–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ensley, B.; Ratzkin, B.; Osslund, T.; Simon, M.; Wackett, L. Expression of naphthalene oxidation genes in Escherichia coli results in the biosynthesis of indigo. Science 1983, 222, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Murdock, D.; Ensley, B.D.; Serdar, C.; Thalen, M. Construction of metabolic operons catalyzing the De Novo biosynthesis of indigo in Escherichia coli. Biotechnology 1993, 11, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Bialy, H. Biotechnology, bioremediation, and blue genes. Nat. Biotechnol. 1997, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Gillam, E.M. Formation of indigo by recombinant mammalian cytochrome P450. Biochem. Biophys. Res. Commun. 1999, 265, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Gillam, E.M.J.; Notley, L.M.; Cai, H.; De Voss, J.J.; Guengerich, F.P. Oxidation of indole by cytochrome P450 enzymes. Biochemistry 2000, 39, 13817–13824. [Google Scholar] [CrossRef]
- Li, Q.S.; Schwaneberg, U.; Fischer, P.; Schmid, R.D. Directed evolution of the fatty-acid hydroxylase P450 BM-3 into an indole-hydroxylating catalyst. Chemistry 2015, 6, 1531–1536. [Google Scholar] [CrossRef]
- Nakamura, K.; Martin, M.V.; Guengerich, F.P. Random mutagenesis of human cytochrome p450 2A6 and screening with indole oxidation products. Arch. Biochem. Biophys. 2001, 395, 25–31. [Google Scholar] [CrossRef]
- Li, H.M.; Mei, L.H.; Urlacher, V.B.; Schmid, R.D.l. Cytochrome P450 BM-3 evolved by random and saturation mutagenesis as an effective indole-hydroxylating catalyst. Appl. Biochem. Biotechnol. 2008, 144, 27–36. [Google Scholar] [CrossRef]
- Wolk, J.L.; Frimer, A.A. Preparation of tyrian purple (6,6′-Dibromoindigo): Past and present. Molecules 2010, 15, 5473–5508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.K.; Shoji, O.; Fujishiro, T.; Ohki, T.; Ueno, T.; Watanabe, Y. Construction of biocatalysts using the myoglobin scaffold for the synthesis of indigo from indole. Catal. Sci. Echnol. 2012, 2, 739–744. [Google Scholar] [CrossRef]
- Nagayama, H.; Sugawara, T.; Endo, R.; Ono, A.; Kato, H.; Ohtsubo, Y.; Nagata, Y.; Tsuda, M. Isolation of oxygenase genes for indigo-forming activity from an artificially polluted soil metagenome by functional screening using Pseudomonas putida strains as hosts. Appl. Microbiol. Biotechnol. 2015, 99, 4453–4470. [Google Scholar] [CrossRef] [PubMed]
- Brixius-Anderko, S.; Hannemann, F.; Ringle, M.; Khatri, Y.; Bernhardt, R. An indole-deficient Escherichia coli strain improves screening of cytochromes P450 for biotechnological applications. Biotechnol. Appl. Biochem. 2017, 64, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, F.; Hatzl, A.M.; Schmidt, S.; Savino, S.; Glieder, A.; Mattevi, A. The extreme structural plasticity in the CYP153 subfamily of P450s directs development of designer hydroxylases. Biochemistry 2018, 57, 6701–6714. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, P.G.; Jung, E.O.; Kim, B.G. In vitro characterization of CYP102G4 from Streptomyces cattleya: A self-sufficient P450 naturally producing indigo. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Fabara, A.N.; Fraaije, M.W. An overview of microbial indigo-forming enzymes. Appl. Microbiol. Biotechnol. 2020, 104, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xu, J.K.; Gao, S.Q.; He, B.; Wei, C.W.; Wang, X.J.; Wang, Z.H.; Lin, Y.W. Green and efficient biosynthesis of indigo from indole by engineered myoglobins. RSC Adv. 2018, 8, 33325–33330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Xu, J.K.; Li, L.Z.; Gao, S.Q.; Wen, G.B.; Lin, Y.W. Design and engineering of neuroglobin to catalyze the synthesis of indigo and derivatives for textile dyeing. Mol. Syst. Des. Eng. 2022, 7, 239–247. [Google Scholar] [CrossRef]
- Oliver, C.F.; Modi, S.; Sutcliffe, M.J.; Primrose, W.U.; Lian, L.Y.; Roberts, G.C.K. A single mutation in cytochrome P450 BM3 changes substrate orientation in a catalytic intermediate and the regiospecificity of hydroxylation. Biochemistry 1997, 36, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.S.; Schwaneberg, U.; Fischer, M.; Schmitt, J.; Schmid, R.D. Rational evolution of a medium chain-specific cytochrome P-450 BM-3 variant. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2001, 1545, 114–121. [Google Scholar] [CrossRef]
- Wichmann, R.; Vasic-Racki, D. Cofactor regeneration at the lab scale. Technol. Transf. Biotechnol. 2005, 92, 225–260. [Google Scholar]
- Whitehouse, C.J.C.; Bell, S.G.; Wong, L.-L. P450(BM3) (CYP102A1): Connecting the dots. Chem. Soc. Rev. 2012, 41, 1218–1260. [Google Scholar] [CrossRef] [PubMed]
- Green, M.T. C-H bond activation in heme proteins: The role of thiolate ligation in cytochrome P450. Curr. Opin. Chem. Biol. 2009, 13, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Meunier, B.; Visser, S.P.d.; Shaik, S. Mechanism of oxidation reactions catalyzed by cytochrome P450 enzymes. Chem. Rev. 2004, 104, 3947–3980. [Google Scholar] [CrossRef] [PubMed]
- Joyce, M.G.; Ekanem, I.S.; Roitel, O.; Dunford, A.J.; Neeli, R.; Girvan, H.M.; Baker, G.J.; Curtis, R.A.; Munro, A.W.; Leys, D. The crystal structure of the FAD/NADPH-binding domain of flavocytochrome P450 BM3. FEBS J. 2012, 279, 1694–1706. [Google Scholar] [CrossRef] [PubMed]
- Modi, A.R.; Dawson, J.H. Oxidizing intermediates in P450 catalysis: A case for multiple oxidants. Adv. Exp. Med. Biol. 2015, 851, 63–81. [Google Scholar] [PubMed]
- Munro, A.W.; McLean, K.J.; Grant, J.L.; Makris, T.M. Structure and function of the cytochrome P450 peroxygenase enzymes. Biochem. Soc. Trans. 2018, 46, 183–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, Y.; Fulco, A.J. Omega-1, Omega-2 and Omega-3 hydroxylation of long-chain fatty acids, amides and alcohols by a soluble enzyme system from Bacillus megaterium. Biochim. Biophys. Acta 1975, 388, 305–317. [Google Scholar] [CrossRef]
- Joyce, M.G.; Girvan, H.M.; Munro, A.W.; Leys, D. A single mutation in cytochrome P450 BM3 induces the conformational rearrangement seen upon substrate binding in the wild-type enzyme. J. Biol. Chem. 2004, 279, 23287–23293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.S.; Ogawa, J.; Schmid, R.D.; Shimizu, S. Indole hydroxylation by bacterial cytochrome P450 BM-3 and modulation of activity by cumene hydroperoxide. Biosci. Biotechnol. Biochem. 2005, 69, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, S.Y.; Fan, S.X.; Jiang, F.J.; Cong, Z.Q. A unique P450 peroxygenase system facilitated by a dual-functional small molecule: Concept, application, and perspective. Antioxidants 2022, 11, 529. [Google Scholar] [CrossRef]
- Shoji, O.; Watanabe, Y. Peroxygenase reactions catalyzed by cytochromes P450. J. Biol. Inorg. Chem. 2014, 19, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.N.; Chen, Z.F.; Chen, J.; Chen, J.F.; Cong, Z. Dual-functional small molecules for generating an efficient cytochrome P450BM3 peroxygenase. Angew. Chem. Int. Ed. 2018, 57, 7628–7633. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kong, F.H.; Ma, N.N.; Zhao, P.X.; Liu, C.; Wang, X.L.; Cong, Z.Q. Peroxide-driven hydroxylation of small alkanes catalyzed by an artificial P450BM3 peroxygenase system. ACS Catal. 2019, 9, 7350–7355. [Google Scholar] [CrossRef]
- Xu, J.K.; Wang, C.L.; Cong, Z.Q. Strategies for substrate-regulated P450 catalysis: From substrate engineering to co-catalysis. Chemistry 2019, 25, 6853–6863. [Google Scholar] [CrossRef] [PubMed]
- Willot, S.J.P.; Tieves, F.; Girhard, M.; Urlacher, V.B.; Hollmann, F.; de Gonzalo, G. P450BM3-catalyzed oxidations employing dual functional small molecules. Catalysts 2019, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.D.; Peng, Y.Q.; Zhao, J.; Li, Q.; Li, A.T. Bacterial cytochrome P450-catalyzed regio- and stereoselective steroid hydroxylation enabled by directed evolution and rational design. Bioresour. Bioprocess. 2020, 7, 2. [Google Scholar] [CrossRef]
- Acevedo-Rocha, C.G.; Gamble, C.; Lonsdale, R.; Li, A.; Nett, N.; Hoebenreich, S.; Lingnau, J.B.; Wirtz, C.; Farès, C.; Hinrichs, H. P450-catalyzed regio- and diastereoselective steroid hydroxylation: Efficient directed evolution enabled by mutability landscaping. ACS Catal. 2018, 8, 3395–3410. [Google Scholar] [CrossRef] [Green Version]
- Omura, T.; Sato, R. The carbon monoxide-binding pigment of liver microsomes. II. solubilization, purification, and properties. J. Biol. Chem. 1964, 239, 2379–2385. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Renata, H.; Peck, N.E.; Farwell, C.C.; Coelho, P.S.; Arnold, F.H. Improved cyclopropanation activity of histidine-ligated cytochrome P450 enables the enantioselective formal synthesis of levomilnacipran. Angew. Chem. Int. Ed. 2014, 126, 6928–6931. [Google Scholar]
- Guengerich, F.P.; Sorrells, J.L.; Schmitt, S.; Krauser, J.A.; Aryal, P.; Meijer, L. Generation of new protein kinase inhibitors utilizing Cytochrome P450 mutant enzymes for indigoid synthesis. J. Med. Chem. 2004, 47, 3236–3241. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yu, Q.; Mei, L.H.; Yao, S.J.; Jin, Z.H. Semi-rational directed evolution in improving indole-hydroxylation ability of cytochrome P450 BM-3. CIESC J. 2009, 60, 2869–2875. [Google Scholar]
- Li, H.M.; Mei, L.H.; Urlacher, V.; Schmid, R. Cytochrome P450BM-3 mutants with improved catalytic properties of hydroxylating indole to indigo by error-prone PCR. Prog. Biochem. Biophys. 2005, 32, 630–635. [Google Scholar]
- Li, H.M.; Mei, L.H.; Gao, S.Y. Amino acid mutation effect on indole-hydroxylating ability of cytochrome P450 BM-3. J. Chem. Eng. Chin. Univ. 2008, 22, 282–287. [Google Scholar]
- Zhang, P.P.; Hu, S.; Mei, L.H.; Lei, Y.L.; Jin, Z.H.; Yao, S.J. Improving indole-hydroxylation ability of cytochrome P450 BM-3 by site-directed mutagenesis. CIESC J. 2013, 64, 3331–3337. [Google Scholar]
- Zhang, P.P.; Hu, S.; Mei, L.H.; Lei, Y.L.; Jin, Z.H.; Yao, S.J. A triple mutant improving activity of cytochrome P450 BM-3 to catalye indole hydroxylation: D168L/E435T/V445A. CIESC J. 2014, 65, 1374–1380. [Google Scholar]
- Reinhard, F.G.C.; Lin, Y.-T.; Stańczak, A.; Visser, S.P.d. Bioengineering of cytochrome P450 OleTJE: How does substrate positioning affect the product distributions? Molecules 2020, 25, 2675. [Google Scholar] [CrossRef]
- Lin, Y.T.; de Visser, S.P. Product distributions of cytochrome P450 OleTJE with phenyl-substituted fatty acids: A computational study. Int. J. Mol. Sci. 2021, 22, 7172. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, L.; Liu, Q.; Zhang, H.; Cao, Y.; Xian, M.; Liu, H. High-specificity synthesis of novel monomers by remodeled alcohol hydroxylase. BMC Biotechnol. 2016, 16, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.F.; Chen, J.; Ma, N.N.; Zhou, H.F.; Cong, Z.Q. Selective hydroxylation of naphthalene using the H2O2-dependent engineered P450BM3 driven by dual-functional small molecules. J. Porphyr. Phthalocyanines 2018, 22, 831–836. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated Docking Using a Lamarckian Genetic Algorithm and Empirical Binding Free Energy Function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System; Schrödinger, LLC: New York, NY, USA, 2010.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | DFSM | kcat (min−1) | Km (mM) | kcat/Km (mM−1min−1) |
|---|---|---|---|---|
| WT P450BM3 | Im-C6-Phe a | nd | nd | nd |
| F87A P450BM3 | Im-C6-Phe | 1007 ± 39.96 | 3.4 ± 0.20 | 296 |
| F87G P450BM3 | Im-C6-Phe | 1304 ± 34.09 | 2.8 ± 0.11 | 466 |
| V78A/F87A P450BM3 | Im-C6-Phe | 1179 ± 18.66 | 1.59 ± 0.05 | 742 |
| V78A/F87G P450BM3 | Im-C6-Phe | nd | nd | nd |
| F87A/T268A P450BM3 | Im-C6-Phe | 687 ± 10.43 | 1.1 ± 0.04 | 625 |
| F87A/T268I P450BM3 | Im-C6-Phe | 223 ± 6.80 | 4.4 ± 0.18 | 50.68 |
| F87A/T268V P450BM3 | Im-C6-Phe | 536 ± 13.81 | 2.33 ± 0.10 | 230 |
| F87G/T268A P450BM3 | Im-C6-Phe | 236 ± 1.47 | 0.17 ± 0.01 | 1388 |
| F87G/T268I P450BM3 | Im-C6-Phe | 478 ± 6.62 | 1.07 ± 0.03 | 447 |
| F87G/T268V P450BM3 | Im-C6-Phe | 1402 ± 44.90 | 2.1 ± 0.12 | 668 |
| A74G/F78V/L188Q/D168N/A255V/K440N P450BM3 [49] | - | 187 | 1.5 | 124 |
| A74G/ F87V/L188Q/D168L [50] | - | 387 | 0.90 | 420 |
| A74G/ F87V/L188Q/D168H [50] | - | 600 | 1.20 | 500 |
| A74G/ F87V/L188Q/E435T [50] | - | 450 | 0.78 | 577 |
| A74G/F87V/L188Q/E435T/D168R P450BM3 [48] | - | 189 | 0.68 | 278 |
| A74G/ F87V/L188Q/D168L/E435T P450BM3 [51] | - | 28.15 | 1.72 | 16.37 |
| D168L/E435T/V445A P450BM3 [52] | - | 73.81 | 2.99 | 24.77 |
| H64D/V68I/I107V Mb b [17] | - | 71.9 | 11.2 | 6.42 |
| F43Y/H64D Mb [23] | - | 5.22 | 0.44 | 11.86 |
| A51C/H64D/F49Y Ngb c [24] | - | 6.92 | 0.16 | 43.25 |
| Enzymes | Indigo (μM) | Indole (μM) | Indigo Yield (%) | Chemoselectivity (%) |
|---|---|---|---|---|
| F87A | 185 | 241 | 37 ± 1.62 | 49 ± 1.92 |
| F87A/T268A | 83 | 635 | 17 ± 1.85 | 46 ± 2.26 |
| F87A/T268I | 69 | 672 | 14 ± 0.94 | 42 ± 1.48 |
| F87A/T268V | 299 | 252 | 60 ± 2.77 | 80 ± 3.62 |
| F87G | 292 | 50 | 58 ± 2.58 | 62 ± 2.67 |
| F87G/T268A | 41 | 811 | 8 ± 0.95 | 44 ± 1.78 |
| F87G/T268I | 156 | 543 | 31 ± 0.10 | 78 ± 2.20 |
| F87G/T268V | 365 | 26 | 73 ± 1.47 | 75 ± 1.69 |
| V78A/F87A | 289 | 24 | 58 ± 3.10 | 59 ± 1.31 |
| Mutation | Sequence (5′-3′) |
|---|---|
| F87A-F | GCGACAAGCTGGACGCATGAAAAAAATTG |
| F87G-F | GGTACAAGCTGGACGCATGAAAAAAATTGGAA |
| F87-R | TAACCCGTCTCCTGCAAAATCACGTACAA |
| T268A-F | GCGACAAGTGGTCTTTTATCATTTGC |
| T268V-F | GTGACAAGTGGTCTTTTATCATTTGC |
| T268I-F | ATCACAAGTGGTCTTTTATCATTTGC |
| T268-R | TTCGTGTCCCGCAATTAAGAATG |
| V78A-F | GCACGTGATTTTGCAG |
| V78-R | CGCTTGACTTAAGTTTTTATC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Lin, Y.; Wang, F.; Wang, J.; Shoji, O.; Xu, J. Construction of Biocatalysts Using the P450 Scaffold for the Synthesis of Indigo from Indole. Int. J. Mol. Sci. 2023, 24, 2395. https://doi.org/10.3390/ijms24032395
Li Y, Lin Y, Wang F, Wang J, Shoji O, Xu J. Construction of Biocatalysts Using the P450 Scaffold for the Synthesis of Indigo from Indole. International Journal of Molecular Sciences. 2023; 24(3):2395. https://doi.org/10.3390/ijms24032395
Chicago/Turabian StyleLi, Yanqing, Yingwu Lin, Fang Wang, Jinghan Wang, Osami Shoji, and Jiakun Xu. 2023. "Construction of Biocatalysts Using the P450 Scaffold for the Synthesis of Indigo from Indole" International Journal of Molecular Sciences 24, no. 3: 2395. https://doi.org/10.3390/ijms24032395
APA StyleLi, Y., Lin, Y., Wang, F., Wang, J., Shoji, O., & Xu, J. (2023). Construction of Biocatalysts Using the P450 Scaffold for the Synthesis of Indigo from Indole. International Journal of Molecular Sciences, 24(3), 2395. https://doi.org/10.3390/ijms24032395