
Toxicity Effects of Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) on Two Green Microalgae Species

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
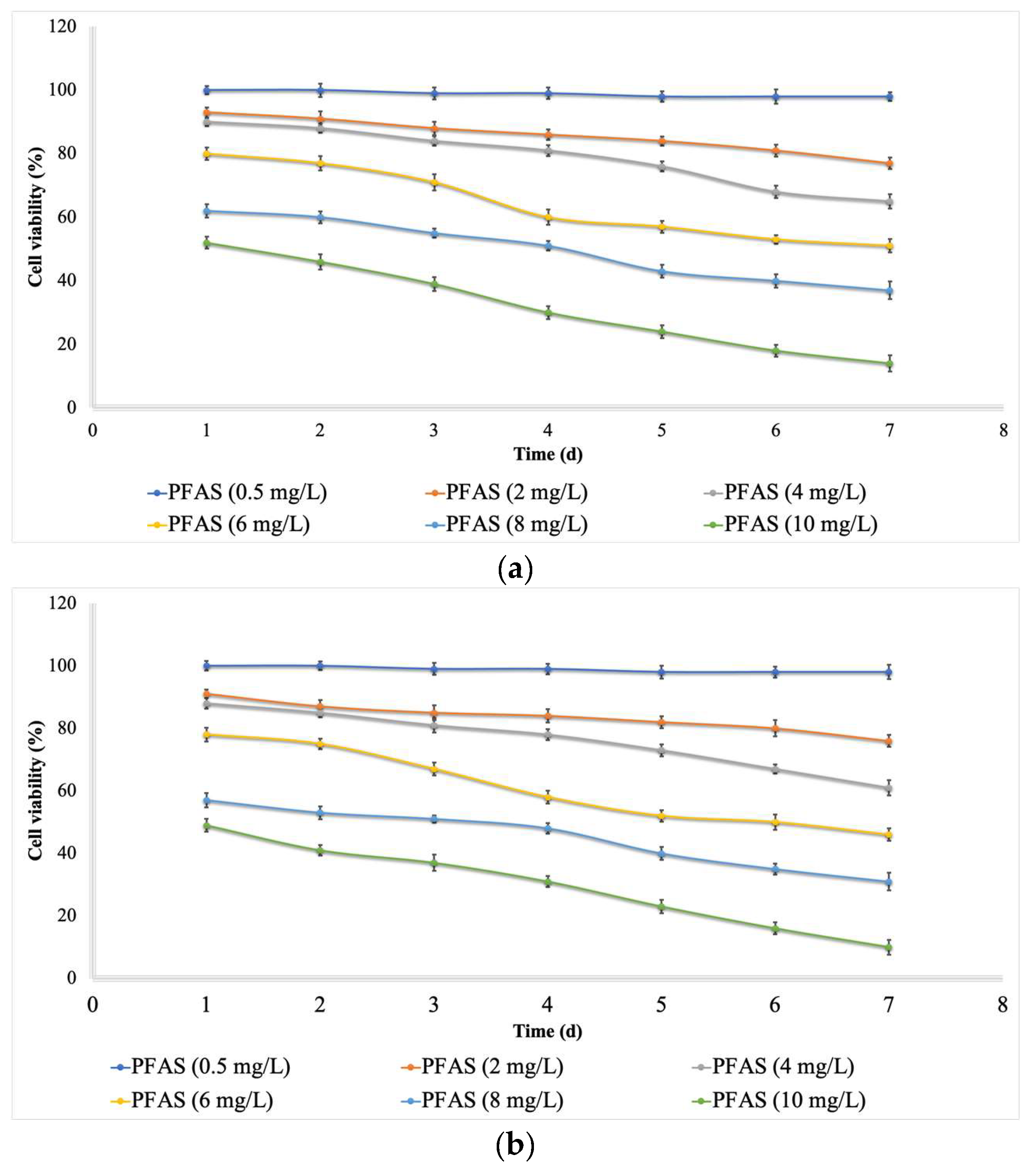
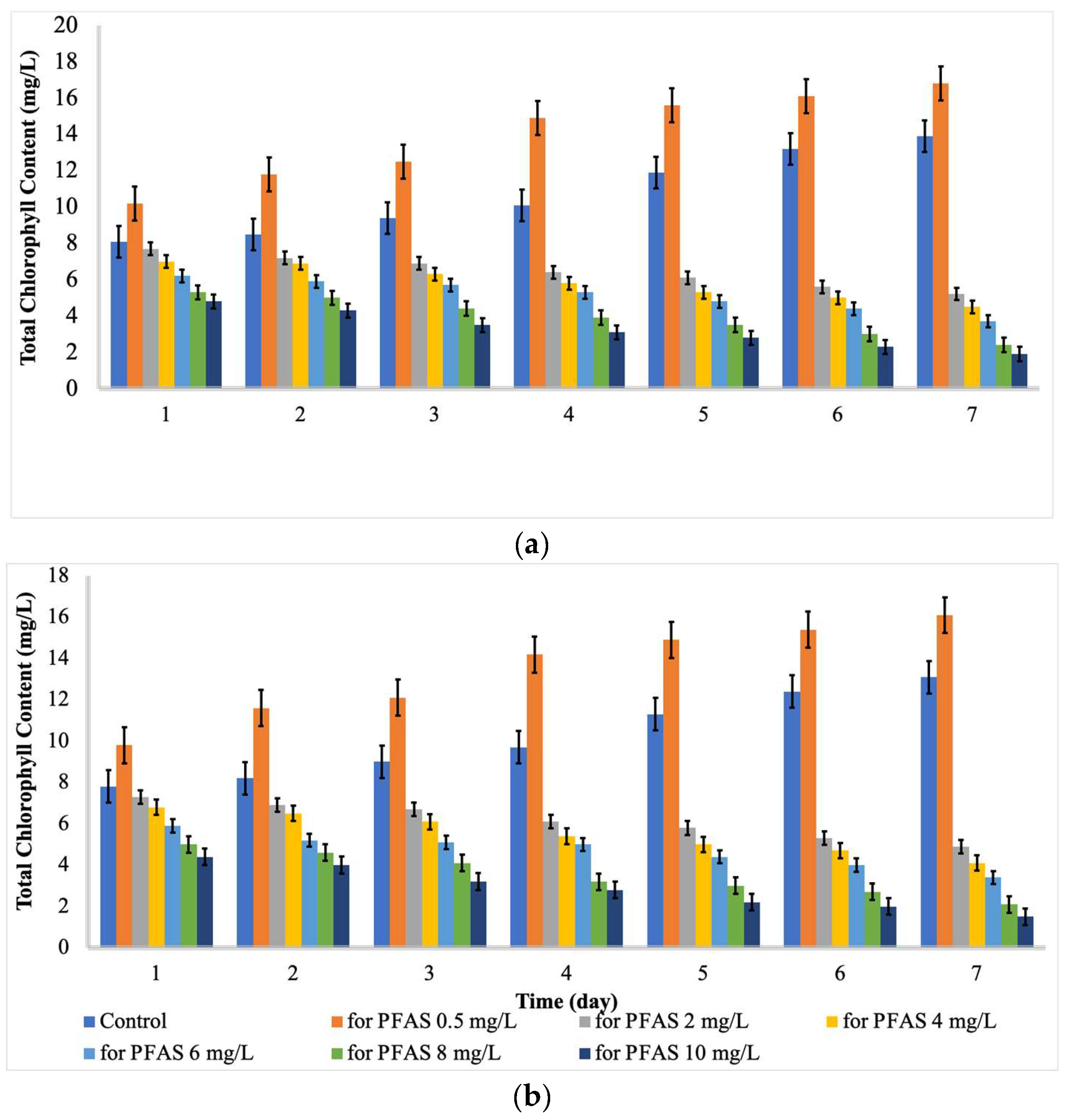
2.1. Effects of PFOA and PFOS on Microalgae
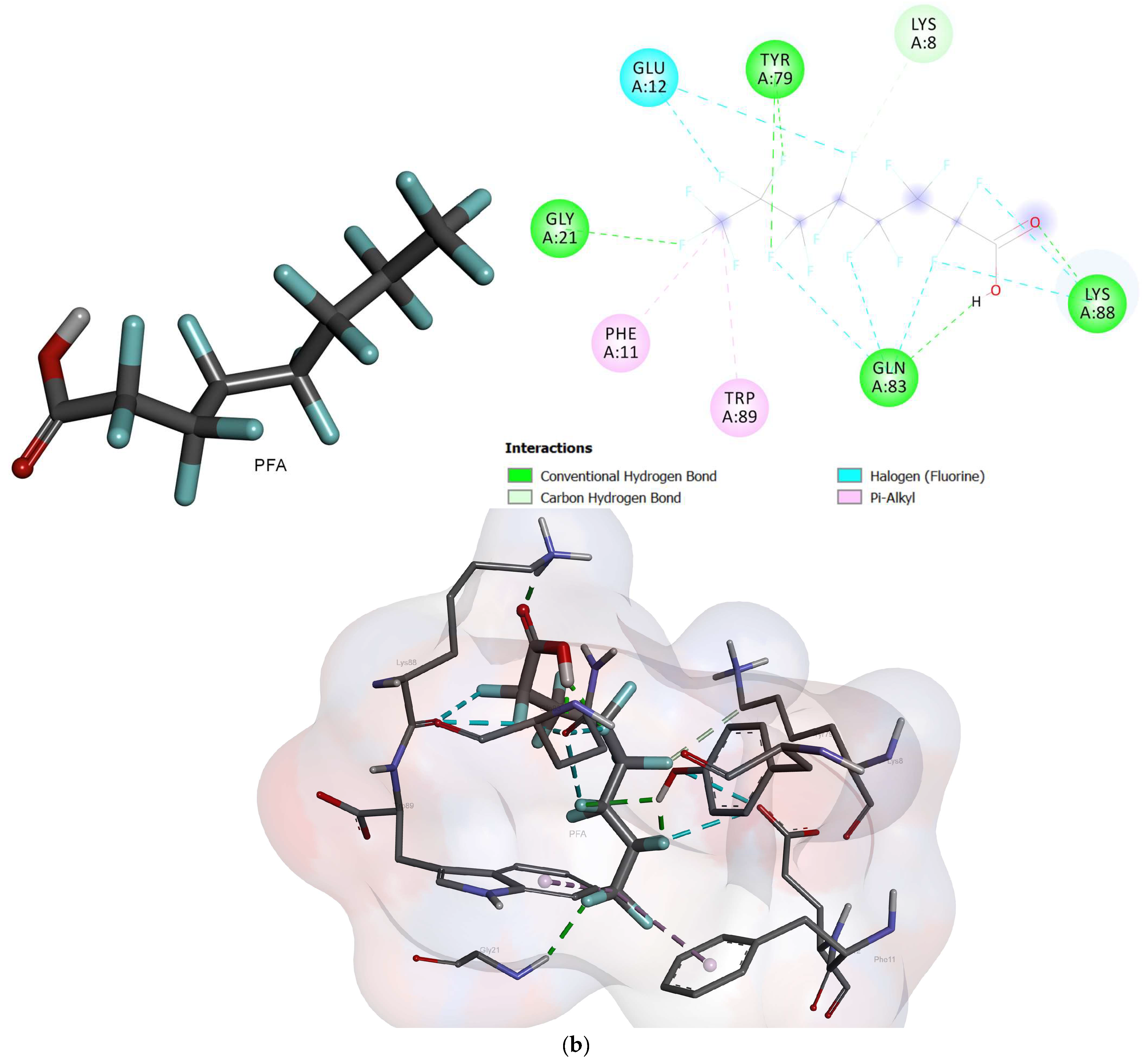
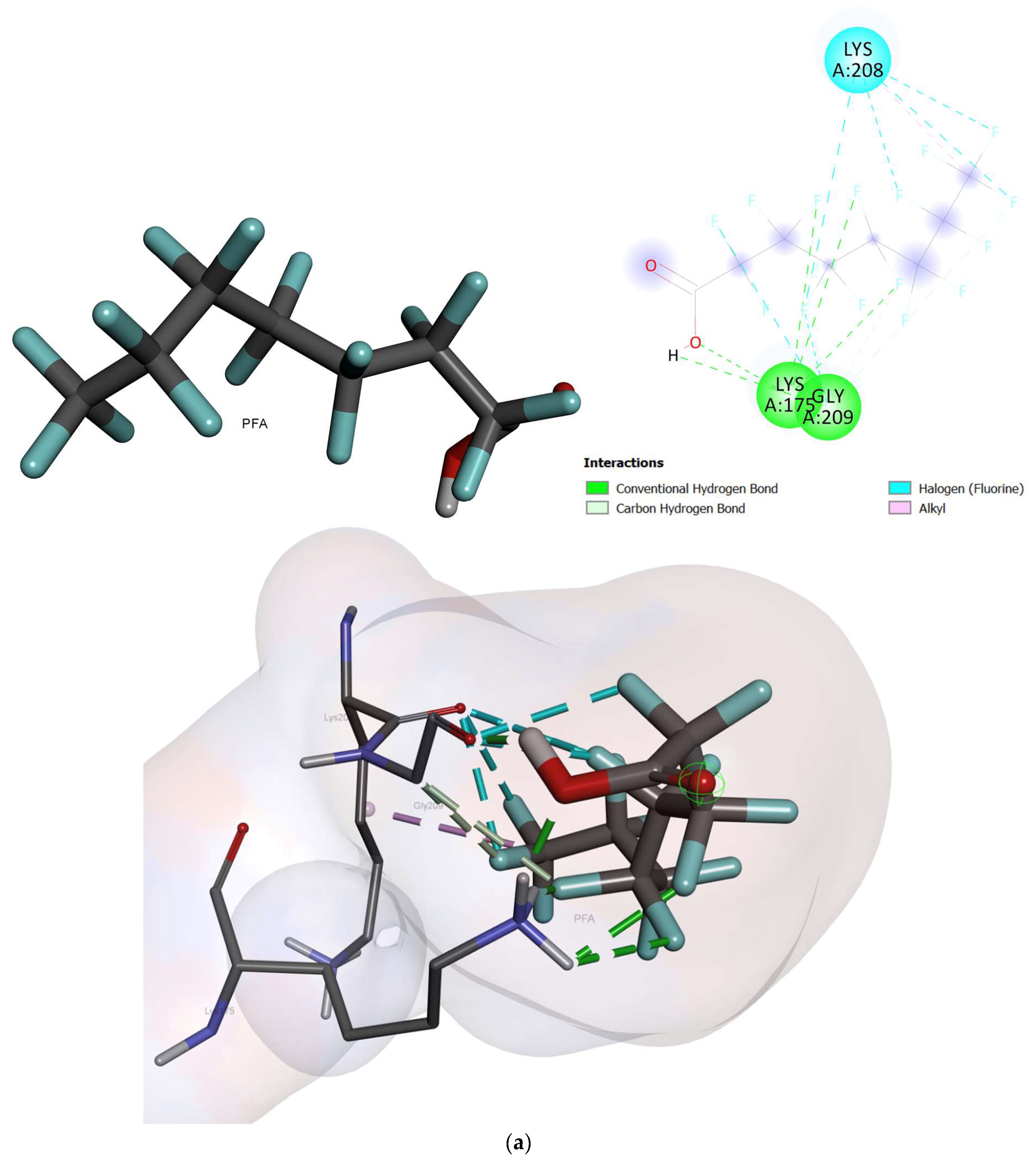
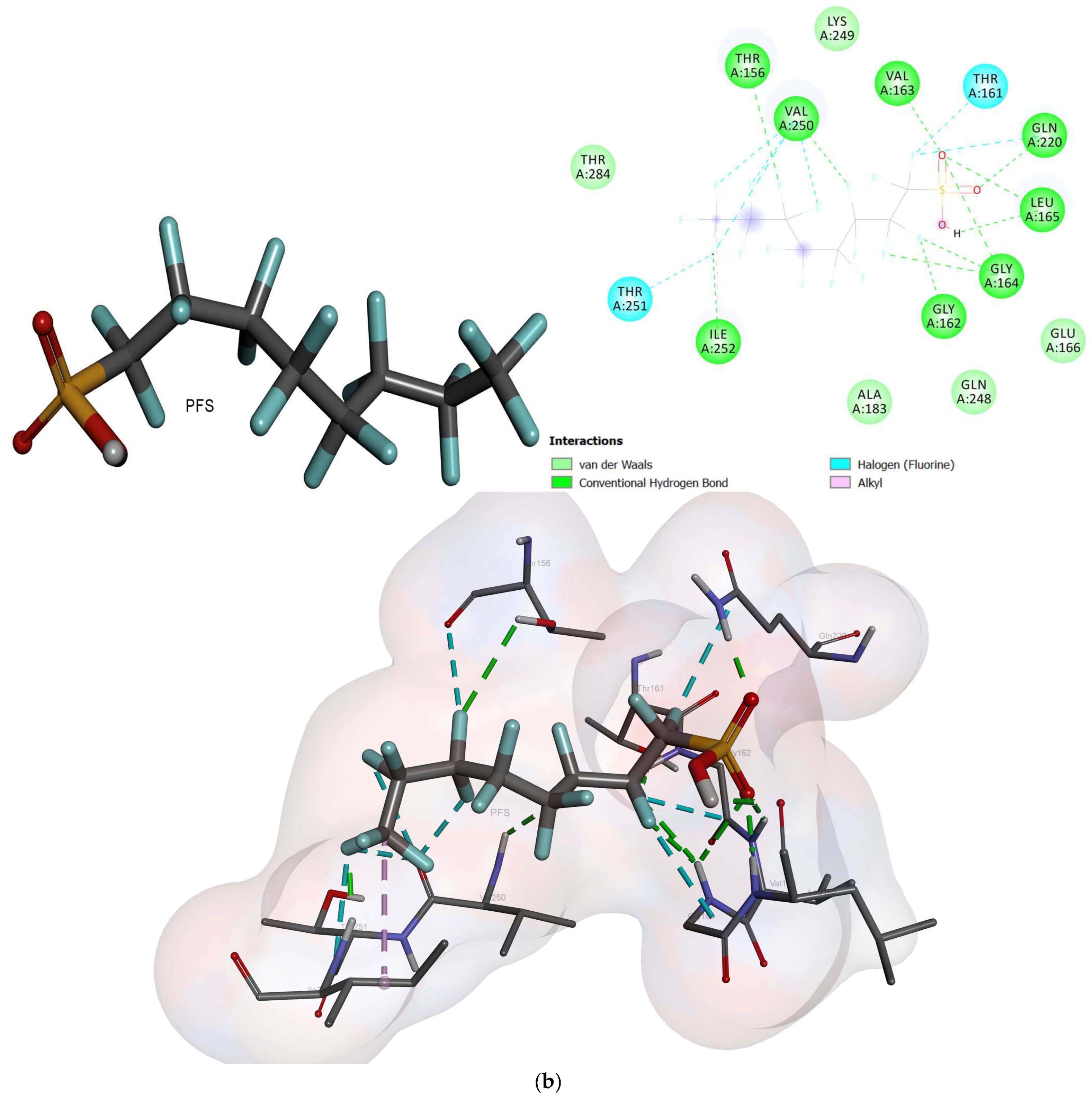
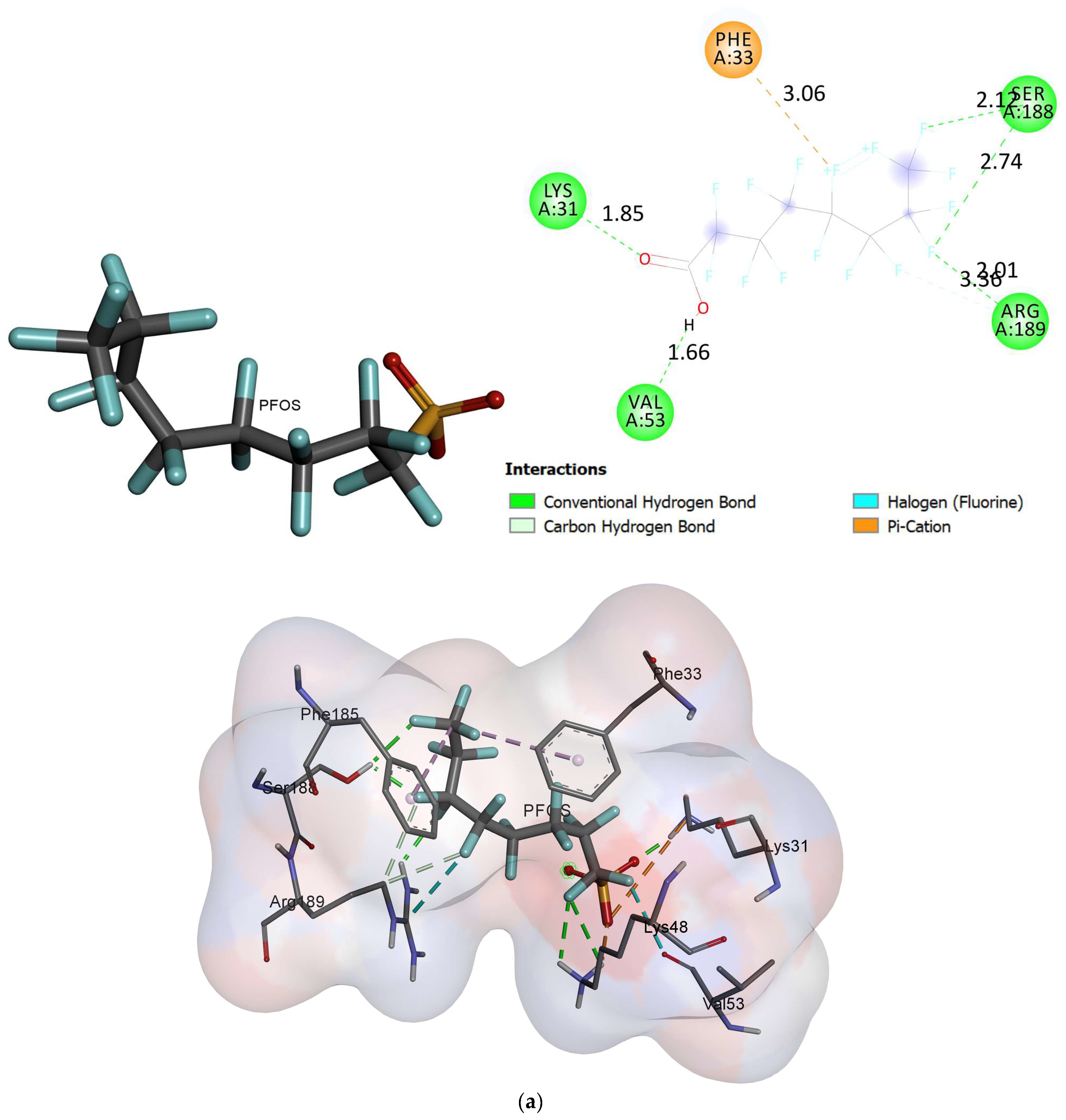
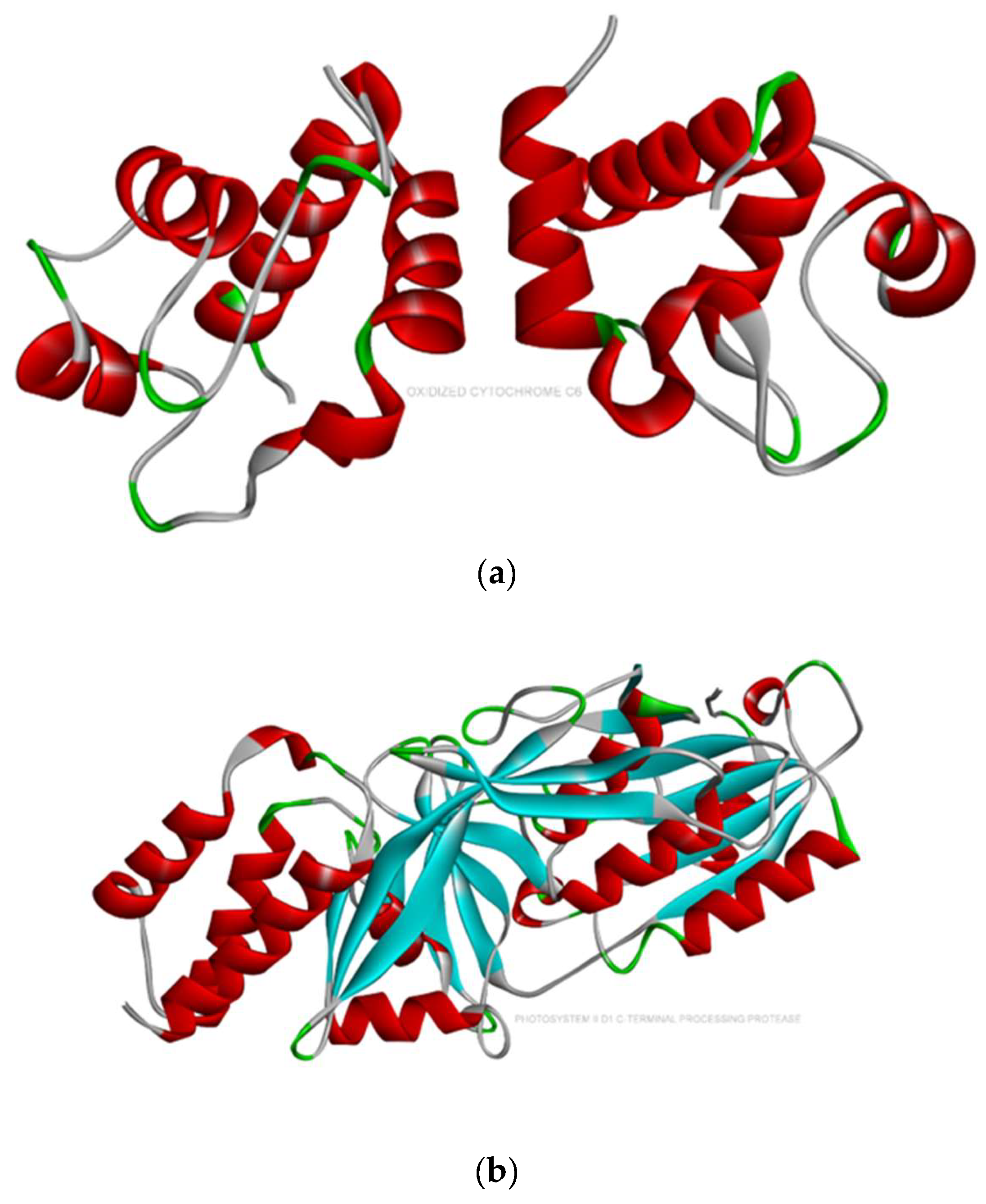
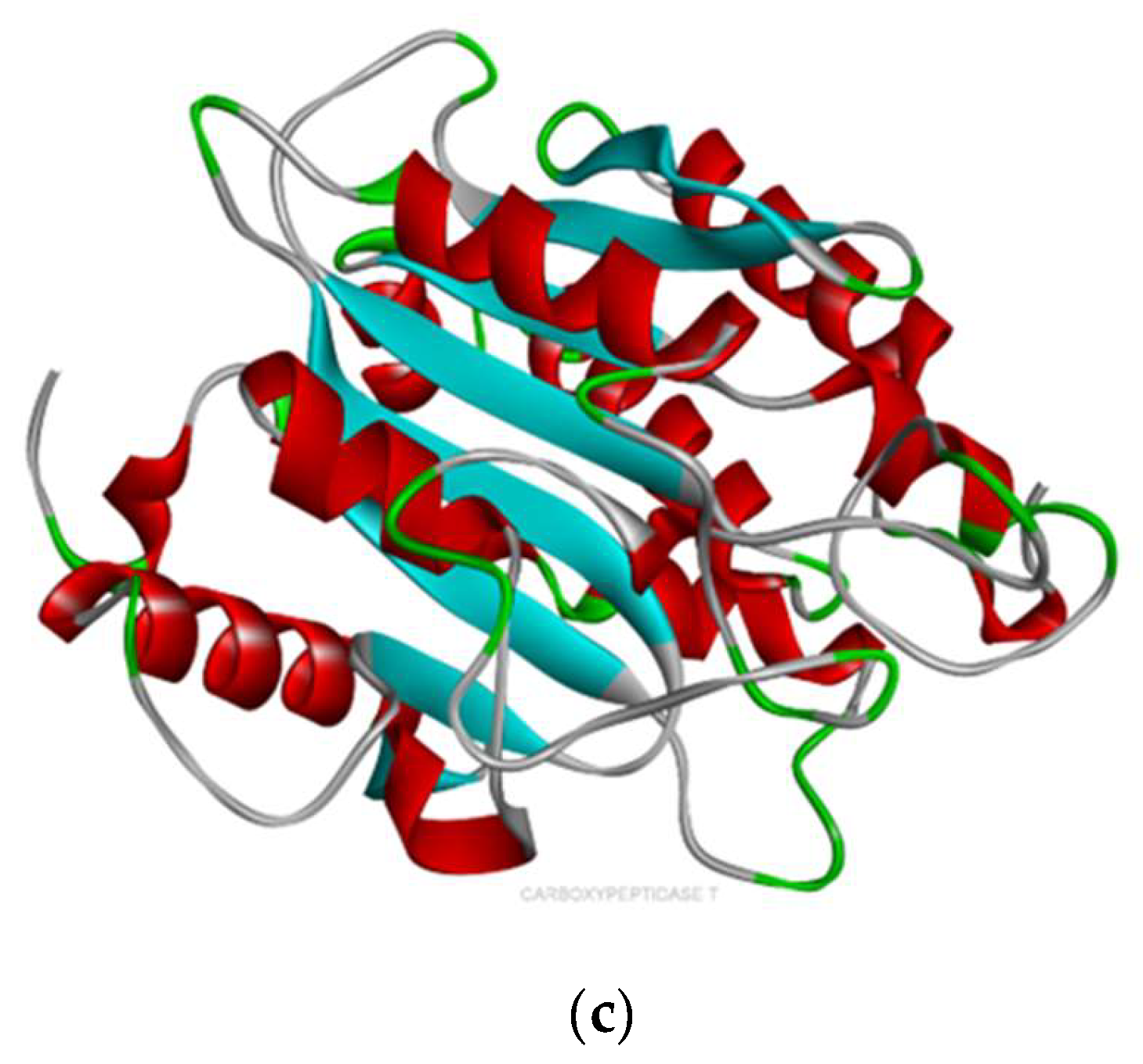
2.2. Molecular Docking
3. Materials and Methods
3.1. Experiments for the Toxicity Effects of PFAS on Microalgae
3.2. Cytotoxicity Rate, and Content of Protein and Chlorophyll
3.3. Analyses the PFAS in Aqueous Solution
3.4. Statistical Analysis and Optimization Process
3.5. Protein Preparation for Bioinformatics Analysis
3.5.1. Software
3.5.2. Methods
3.5.3. Ligand Preparation
4. Conclusions
- The minimum cell viability (16%) and content of chlorophyll (1.9 mg L−1) and protein (9%) for Chlorella vulgaris were detected at high concentrations of the PFOS and PFOA mixture (10 mg L−1) and at a long exposure time (contact time, 7 days).
- The minimum cell viability (11%) and content of chlorophyll (1.5 mg L−1) and protein (8%) for Scenedesmus obliquus were reported at high concentrations of the PFOS and PFOA mixture (10 mg L−1) and at a long exposure time (contact time, 7 days).
- In comparison to Chlorella vulgaris, Scenedesmus obliquus is more sensitive to exposure to PFAS.
- Based on the molecular docking investigation, C. vulgaris and Scenedesmus obliquus can potentially interact with PFOA and PFOS more efficiently when utilized together due to the presence of the examined enzymes. This may reduce the toxic effects of PFAS in water resources. However, further investigation is needed to confirm this hypothesis.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gagliano, E.; Sgroi, M.; Falciglia, P.P.; Vagliasindi, F.G.A.; Roccaro, P. Removal of poly- and perfluoroalkyl substances (PFAS) from water by adsorption: Role of PFAS chain length, effect of organic matter and challenges in adsorbent regeneration. Water Res. 2020, 171, 115381. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.-J.; Tian, Y.; Sobhani, Z.; Naidu, R.; Fang, C. Synergistic degradation of PFAS in water and soil by dual-frequency ultrasonic activated persulfate. Chem. Eng. J. 2020, 388, 124215. [Google Scholar] [CrossRef]
- Scheringer, M.; Trier, X.; Cousins, I.T.; de Voogt, P.; Fletcher, T.; Wang, Z.; Webster, T.F. Helsingør Statement on poly- and perfluorinated alkyl substances (PFASs). Chemosphere 2014, 114, 337–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, N.; Prabakar, S.; Choi, H. Dependency of the photocatalytic and photochemical decomposition of per- and polyfluoroalkyl substances (PFAS) on their chain lengths, functional groups, and structural properties. Water Sci. Technol. 2021, 84, 3738–3754. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Simcik, M.F.; Halbach, T.R.; Gulliver, J.S. Perfluorooctane sulfonate (PFOS) and perfluorooctanoate (PFOA) in soils and groundwater of a U.S. metropolitan area: Migration and implications for human exposure. Water Res. 2015, 72, 64–74. [Google Scholar] [CrossRef]
- Pauletto, P.S.; Florent, M.; Bandosz, T.J. Insight into the mechanism of perfluorooctanesulfonic acid adsorption on highly porous media: Sizes of hydrophobic pores and the extent of multilayer formation. Carbon 2022, 191, 535–545. [Google Scholar] [CrossRef]
- Bernardini, I.; Matozzo, V.; Valsecchi, S.; Peruzza, L.; Rovere, G.D.; Polesello, S.; Iori, S.; Marin, M.G.; Fabrello, J.; Ciscato, M.; et al. The new PFAS C6O4 and its effects on marine invertebrates: First evidence of transcriptional and microbiota changes in the Manila clam Ruditapes philippinarum. Environ. Int. 2021, 152, 106484. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Zheng, X.; Yang, M.; Gao, X.; Huang, J.; Zhang, L.; Fan, Z. Toxic effects and mechanisms of PFOA and its substitute GenX on the photosynthesis of Chlorella pyrenoidosa. Sci. Total Environ. 2021, 765, 144431. [Google Scholar] [CrossRef]
- Hu, C.; Luo, Q.; Huang, Q. Ecotoxicological effects of perfluorooctanoic acid on freshwater microalgae Chlamydomonas reinhardtii and Scenedesmus obliquus. Environ. Toxicol. Chem. 2014, 33, 1129–1134. [Google Scholar] [CrossRef]
- Baharlooeian, M.; Kerdgari, M.; Shimada, Y. Ecotoxicological effects of TiO2 nanoparticulates and bulk Ti on microalgae Chaetoceros muelleri. Environ. Technol. Innov. 2021, 23, 101720. [Google Scholar] [CrossRef]
- Keller, A.A.; McFerran, S.; Lazareva, A.; Suh, S. Global life cycle releases of engineered nanomaterials. J. Nanopart. Res. 2013, 15, 1692. [Google Scholar] [CrossRef]
- Niu, Z.; Na, J.; Xu, W.; Wu, N.; Zhang, Y. The effect of environmentally relevant emerging per- and polyfluoroalkyl substances on the growth and antioxidant response in marine Chlorella sp. Environ. Pollut. 2019, 252, 103–109. [Google Scholar] [CrossRef]
- González-Naranjo, V.; Boltes, K. Toxicity of ibuprofen and perfluorooctanoic acid for risk assessment of mixtures in aquatic and terrestrial environments. Int. J. Environ. Sci. Technol. 2014, 11, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Marrez, D.A.; Naguib, M.M.; Sultan, Y.Y.; Higazy, A.M. Antimicrobial and anticancer activities of Scenedesmus obliquus metabolites. Heliyon 2019, 5, e01404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, Y.; Zheng, X.; Zhang, L.; Lyu, H.; Huang, H.; Fan, Z. Anti-oxidant mechanisms of Chlorella pyrenoidosa under acute GenX exposure. Sci. Total Environ. 2021, 797, 149005. [Google Scholar] [CrossRef] [PubMed]
- Wirth, R.; Pap, B.; Böjti, T.; Shetty, P.; Lakatos, G.; Bagi, Z.; Kovács, K.L.; Maróti, G. Chlorella vulgaris and Its Phycosphere in Wastewater: Microalgae-Bacteria Interactions During Nutrient Removal. Front. Bioeng. Biotechnol. 2020, 8, 557572. [Google Scholar] [CrossRef]
- Ferreira, A.; Ribeiro, B.; Marques, P.A.S.S.; Ferreira, A.F.; Dias, A.P.; Pinheiro, H.M.; Reis, A.; Gouveia, L. Scenedesmus obliquus mediated brewery wastewater remediation and CO 2 biofixation for green energy purposes. J. Clean. Prod. 2017, 165, 1316–1327. [Google Scholar] [CrossRef]
- Phong, N.V.; Oanh, V.T.; Yang, S.Y.; Choi, J.S.; Min, B.S.; Kim, J.A. PTP1B inhibition studies of biological active phloroglucinols from the rhizomes of Dryopteris crassirhizoma: Kinetic properties and molecular docking simulation. Int. J. Biol. Macromol. 2021, 188, 719–728. [Google Scholar] [CrossRef]
- Qin, X.; Zhong, J.; Wang, Y. A mutant T1 lipase homology modeling, and its molecular docking and molecular dynamics simulation with fatty acids. J. Biotechnol. 2021, 337, 24–34. [Google Scholar] [CrossRef]
- Oukarroum, A.; Samadani, M.; Dewez, D. Influence of pH on the Toxicity of Silver Nanoparticles in the Green Alga Chlamydomonas acidophila. Water Air Soil Pollut. 2014, 225, 2038. [Google Scholar] [CrossRef]
- Nagai, T.; Ishihara, S.; Yokoyama, A.; Iwafune, T. Effects of four rice paddy herbicides on algal cell viability and the relationship with population recovery. Environ. Toxicol. Chem. 2011, 30, 1898–1905. [Google Scholar] [CrossRef] [PubMed]
- Marchetto, F.; Roverso, M.; Righetti, D.; Bogialli, S.; Filippini, F.; Bergantino, E.; Sforza, E. Bioremediation of Per- and Poly-Fluoroalkyl Substances (PFAS) by Synechocystis sp. PCC 6803: A Chassis for a Synthetic Biology Approach. Life 2021, 11, 1300. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, X.; Zhang, L.; Li, J.; Li, Y.; Huang, H.; Fan, Z. Joint toxicity mechanisms of binary emerging PFAS mixture on algae (Chlorella pyrenoidosa) at environmental concentration. J. Hazard. Mater. 2022, 437, 129355. [Google Scholar] [CrossRef] [PubMed]
- Seoane, M.; Cid, Á.; Esperanza, M. Toxicity of bisphenol A on marine microalgae: Single- and multispecies bioassays based on equivalent initial cell biovolume. Sci. Total Environ. 2021, 767, 144363. [Google Scholar] [CrossRef] [PubMed]
- El-Sheekh, M.M.; Hamouda, R.A.; Nizam, A.A. Biodegradation of crude oil by Scenedesmus obliquus and Chlorella vulgaris growing under heterotrophic conditions. Int. Biodeterior. Biodegrad. 2013, 82, 67–72. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, Y.; Li, R.; Yin, Q.; Song, C. Removal of cefradine by Chlorella sp. L166 and Scenedesmus quadricauda: Toxicity investigation, degradation mechanism and metabolic pathways. Process Saf. Environ. Prot. 2022, 160, 632–640. [Google Scholar] [CrossRef]
- Xiong, J.-Q.; Kurade, M.B.; Jeon, B.-H. Ecotoxicological effects of enrofloxacin and its removal by monoculture of microalgal species and their consortium. Environ. Pollut. 2017, 226, 486–493. [Google Scholar] [CrossRef]
- Nong, Q.-Y.; Liu, Y.-A.; Qin, L.-T.; Liu, M.; Mo, L.-Y.; Liang, Y.-P.; Zeng, H.-H. Toxic mechanism of three azole fungicides and their mixture to green alga Chlorella pyrenoidosa. Chemosphere 2021, 262, 127793. [Google Scholar] [CrossRef]
- Hu, J.; Wang, D.; Zhang, N.; Tang, K.; Bai, Y.; Tian, Y.; Li, Y.; Zhang, X. Effects of perfluorooctanoic acid on Microcystis aeruginosa: Stress and self-adaptation mechanisms. J. Hazard. Mater. 2023, 445, 130396. [Google Scholar] [CrossRef]
- Mojiri, A.; Baharlooeian, M.; Zahed, M.A. The Potential of Chaetoceros muelleri in Bioremediation of Antibiotics: Performance and Optimization. Int. J. Environ. Res. Public Health 2021, 18, 977. [Google Scholar] [CrossRef]
- Novotný, J.; Bazzi, S.; Marek, R.; Kozelka, J. Lone-pair–π interactions: Analysis of the physical origin and biological implications. Phys. Chem. Chem. Phys. 2016, 18, 19472–19481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahraman, A.; Morris, R.J.; Laskowski, R.A.; Favia, A.D.; Thornton, J.M. On the diversity of physicochemical environments experienced by identical ligands in binding pockets of unrelated proteins. Proteins Struct. Funct. Bioinform. 2010, 78, 1120–1136. [Google Scholar] [CrossRef] [PubMed]
- Mojiri, A.; Zhou, J.L.; Nazari, V.M.; Rezania, S.; Farraji, H.; Vakili, M. Biochar enhanced the performance of microalgae/bacteria consortium for insecticides removal from synthetic wastewater. Process Saf. Environ. Prot. 2022, 157, 284–296. [Google Scholar] [CrossRef]
- Fan, L.; Tang, J.; Zhang, D.; Ma, M.; Wang, Y.; Han, Y. Investigations on the phytotoxicity of perfluorooctanoic acid in Arabidopsis thaliana. Environ. Sci. Pollut. Res. 2020, 27, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Namasivayam, S.K.; Jayakumar, D.; Kumar, R.; Bharani, R.A. Antibacterial and anticancerous biocompatible silver nanoparticles synthesised from the cold-tolerant strain of Spirulina platensis. J. Coast. Life Med. 2015, 3, 265–272. [Google Scholar] [CrossRef]
- Xiong, J.-Q.; Kurade, M.B.; Kim, J.R.; Roh, H.-S.; Jeon, B.-H. Ciprofloxacin toxicity and its co-metabolic removal by a freshwater microalga Chlamydomonas mexicana. J. Hazard. Mater. 2017, 323, 212–219. [Google Scholar] [CrossRef]
- Damergi, E.; Schwitzguébel, J.-P.; Refardt, D.; Sharma, S.; Holliger, C.; Ludwig, C. Extraction of carotenoids from Chlorella vulgaris using green solvents and syngas production from residual biomass. Algal Res. 2017, 25, 488–495. [Google Scholar] [CrossRef]
- Jabir, T.F.; Noor Abbood, H.A.; Salman, F.S.; Hafit, A.Y. Influence of pH, pesticide and radiation interactions on the chemical composition of Chlorella vulgaris algae. IOP Conf. Ser. Earth Environ. Sci. 2021, 722, 12046. [Google Scholar] [CrossRef]
- Yang, B.; Han, Y.; Deng, Y.; Li, Y.; Zhuo, Q.; Wu, J. Highly efficient removal of perfluorooctanoic acid from aqueous solution by H 2 O 2 -enhanced electrocoagulation-electroflotation technique. Emerg. Contam. 2016, 2, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Mojiri, A.; Baharlooeian, M.; Kazeroon, R.A.; Farraji, H.; Lou, Z. Removal of Pharmaceutical Micropollutants with Integrated Biochar and Marine Microalgae. Microorganisms 2021, 9, 4. [Google Scholar] [CrossRef]
- Fang, S.; Li, L.; Cui, B.; Men, S.; Shen, Y.; Yang, X. Structural insight into plant programmed cell death mediated by BAG proteins in Arabidopsis thaliana. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.; Nazari, V.M.; Yaseen, M.; Iqbal, M.A.; Ahamed, M.B.K.; Majid, A.S.A.; Bhatti, H.N. Green synthesis of selenium-N-heterocyclic carbene compounds: Evaluation of antimicrobial and anticancer potential. Bioorg. Chem. 2019, 90, 103042. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.; Nazari, M.; Iqbal, M.A.; Bhatti, H.N.; Ahmed, M.B.K.; Majid, A.M.S.A. Unsymmetrically substituted benzimidazolium based Silver(I)-N-heterocyclic carbene complexes: Synthesis, characterization and in vitro anticancer study against human breast cancer and colon cancer. J. Saudi Chem. Soc. 2019, 23, 795–808. [Google Scholar] [CrossRef]
- Hayat, K.; Tariq, U.; Wong, Q.A.; Quah, C.K.; Majid, A.S.A.; Nazari, V.M.; Ahamed, M.B.K.; Iqbal, M.A.; Tirmizi, S.A. Green synthesis of selenium based N-heterocyclic carbene compounds; structural, in-vitro anticancer and molecular docking studies. Comput. Biol. Chem. 2021, 94, 107567. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reponses | R2 * | Adj. R2 | Adec. P | SD | CV |
|---|---|---|---|---|---|
| Cell viability | 0.987 | 0.985 | 78.94 | 2.98 | 4.37 |
| Protein content | 0.900 | 0.891 | 29.8 | 3.91 | 8.5 |
| Total chlorophyll | 0.900 | 0.892 | 29.4 | 4.03 | 9.3 |
| Reponses | R2 * | Adj. R2 | Adec. P | SD | CV |
|---|---|---|---|---|---|
| Cell viability | 0.985 | 0.983 | 74.93 | 3.23 | 4.92 |
| Protein content | 0.907 | 0.902 | 28.8 | 6.31 | 6.31 |
| Total chlorophyll | 0.907 | 0.901 | 29.83 | 1.73 | 8.60 |
| FbE(Kcal/mol) | Vug | Cyt6 | Protease |
|---|---|---|---|
| PFOA | −2.96 | −1.98 | −2.83 |
| PFOS | −3.07 | −2.42 | −3.22 |
| Ki | Vug | Cyt6 | Protease |
| PFOA | 6.75 mM | 35.58 mM | 8.42 mM |
| PFOS | 5.60 mM | 16.90 mM | 4.38 mM |
| Compounds | Chemical Formula | Chemical Structure | Molecular Weight (g/mol) | CAS Number |
|---|---|---|---|---|
| PFOA | CF3(CF2)6COOH |  | 414.07 | 335-67-1 |
| PFOS | CF3(CF2)7SO3H |  | 500.13 | 1763-23-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mojiri, A.; Nazari Vishkaei, M.; Ansari, H.K.; Vakili, M.; Farraji, H.; Kasmuri, N. Toxicity Effects of Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) on Two Green Microalgae Species. Int. J. Mol. Sci. 2023, 24, 2446. https://doi.org/10.3390/ijms24032446
Mojiri A, Nazari Vishkaei M, Ansari HK, Vakili M, Farraji H, Kasmuri N. Toxicity Effects of Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) on Two Green Microalgae Species. International Journal of Molecular Sciences. 2023; 24(3):2446. https://doi.org/10.3390/ijms24032446
Chicago/Turabian StyleMojiri, Amin, Mansoureh Nazari Vishkaei, Hanieh Khoshnevis Ansari, Mohammadtaghi Vakili, Hossein Farraji, and Norhafezah Kasmuri. 2023. "Toxicity Effects of Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) on Two Green Microalgae Species" International Journal of Molecular Sciences 24, no. 3: 2446. https://doi.org/10.3390/ijms24032446
APA StyleMojiri, A., Nazari Vishkaei, M., Ansari, H. K., Vakili, M., Farraji, H., & Kasmuri, N. (2023). Toxicity Effects of Perfluorooctanoic Acid (PFOA) and Perfluorooctane Sulfonate (PFOS) on Two Green Microalgae Species. International Journal of Molecular Sciences, 24(3), 2446. https://doi.org/10.3390/ijms24032446