
Necrotizing Enterocolitis: The Role of Hypoxia, Gut Microbiome, and Microbial Metabolites

 , , ,
, , ,
Abstract
:1. Introduction
2. Risk Factors for NEC
2.1. Prenatal Risk Factors
2.2. Perinatal and Postnatal Risk Factors of NEC
3. The Role of Hypoxia as a Risk Factor for NEC
3.1. Physiological Niches of Hypoxia: Fetus, Placenta, and Intestines
3.2. Oxidative Stress and Its Consequences for Glycosylation of the Cellular Glycocalyx
3.3. Intestinal Ischemia and NEC
3.4. Epithelial Barrier Dysfunction
3.5. The Role of Fucosylation in the Regulation of TLR-4-Mediated Activation of the Notch Signaling Pathway
4. Fucosylated Glycans and the Risk of Developing NEC
5. Human Milk Oligosaccharides and Formation of the Intestinal Microbiome
6. Maternal Microbiome Dysbiosis as a Risk Factor for NEC
6.1. The Maternal Microbiome in Preterm Birth
6.2. The Maternal Gut Microbiome in Pre-Eclampsia
6.3. The Gut Microbiome of Mothers and Neonates with IUGR
7. The Microbiome of Preterm Infants
Dynamics of Gut Microbial Colonization in Neonates
- Type S (with a prevalence of Staphylococcus epidermidis);
- Type E (with a predominance of Enterobacteriaceae—Klebsiella pneumoniae or Escherichia coli);
- Type O (wide-spectrum anaerobic and optional microorganisms—Bifidobacterium, Lactobacillales, Veillonellales, and Eubacteriales).
- PT-CST1 (Enterococcus faecalis predominance, Actinomyces, Schaalia);
- PT-CST2 (Escherichia coli predominance, Bacteroides fragilis);
- PT-CST3 (Streptococcus agalacticae predominance, Streptococcus vestibularis);
- PT-CST4 (Staphylococcus epidermidis predominance);
- PT-CST5 (Klebsiella pneumoniae predominance, Akkermansia).
8. Features of the Intestinal Microbiota and Metabolome in NEC
8.1. The Role of Gammaproteobacteria
8.2. The Role of Clostridia
8.3. Is the Predominance of Gammaproteobacteria or Clostridia Dependent on GA?
8.4. The Role of Environmental and Nosocomial Microorganisms
8.5. Intestinal Virome in NEC
8.6. Gut Microbiome Metabolic Effects and Microbial Metabolites in NEC in Preterm Infants
9. Prediction of NEC
10. Remote Ischemic Conditioning (RIC) as a Therapy for NEC
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2′-FL | 2-fucosyllactose |
| 3′-FL | 3-fucosyllactose |
| 3′-SL | 3-sialyllactose |
| 6′-SL | 6-sialyllactose |
| BAX | Bcl-2-associated X protein |
| bFGF | basic fibroblast growth factor |
| CHD | congenital heart disease |
| DCDA | dichorionic diamniotic |
| DSS | dextran sulfate sodium |
| EGF | epidermal growth factor |
| eNOS | endothelial NO synthase |
| ET-1 | endothelin-1 |
| FGFR2 | fibroblast growth factor receptor 2 |
| F-Tr | GDP-fucose transporter |
| Fuc | fucose |
| FUK | fucokinase |
| FUT1 | fucosyltransferase 1 |
| FUT2 | fucosyltransferase 2 |
| FUT7 | fucosyltransferase VII |
| FX | GDP-4-keto-6-deoxymannose 3,5- epimerase- 4-reductase |
| GA | gestational age |
| GDM | gestational diabetes mellitus |
| GFPP | GDP-β-l-fucose pyrophosphorylase |
| GL | genus level |
| GlcNAc | N-acetyl-glucosamine |
| GLUT1 | glucose transporter 1 |
| GMD | GDP-d-mannose-4,6-dehydratase |
| HIF | factors induced by hypoxia |
| HMO | human milk oligosaccharides |
| IBD | inflammatory bowel disease |
| IL-1β | interleukin-1β |
| IUGR | intrauterine growth retardation |
| Ley | Lewis Y |
| LPS | lipopolysaccharide |
| MCDA | monochorionic diamniotic twins |
| MUC1 | mucin 1 |
| MUC2 | mucin-2 |
| MUC5AC | mucin-5AC |
| NEC | necrotizing enterocolitis |
| NF-κB | nuclear factor kappa B |
| NO | nitric oxide |
| OTUs | operational taxonomic units |
| PAF | platelet-activating factor |
| PAI-1 | plasminogen activator inhibitor-1 |
| PDA | persistent ductus arteriosus |
| PE | pre-eclampsia |
| PHD3 | HIF-prolyl hydroxylase 3 |
| PI-CST | preterm community state types |
| PIGF | placental growth factor |
| PIH | pregnancy-induced hypertension |
| PL | phylum level |
| POFUT | protein O-fucosyltransferase |
| RIC | remote ischemic conditioning |
| ROS | reactive oxygen species |
| SCFAs | short-chain fatty acids |
| SCT | syncytiotrophoblast |
| sEng | soluble endoglin |
| sFlt-1 | soluble fms-like tyrosine kinase-1 |
| SIRT1 | sirtuin 1 |
| SL | species level |
| ST6GAL1 | beta-galactosamide-alpha-2,6-sialyltransferase 1 |
| TFF2 | trefoil factor 2 |
| TFF3 | trefoil factor 3 |
| TGF-β1 | transforming growth factor beta 1 |
| TLR4 | toll-like receptor 4 |
| TMAO | trimethylamine-N-oxide |
| TNF-α | tumor necrosis factor alpha |
| TSR | thrombospondin--type 1 repeats |
| UDP-galactose | uridine diphosphate galactose |
| UGT1 | uridine diphosphate glucuronosyl transferase |
| VEGF | vascular endothelial growth factor |
| VEGFR | vascular endothelial growth factor receptor |
| VLBW | very low birth weight |
| WAZ | weight-for-age z-score |
| ZO-1 | zonula occludens-1 |
| α1,6-Fuc | α-1,6-fucosyltransferase |
References
- Meister, A.L.; Doheny, K.K.; Travagli, R.A. Necrotizing enterocolitis: It’s not all in the gut. Exp. Biol. Med. 2020, 245, 85–95. [Google Scholar] [CrossRef]
- Alsaied, A.; Islam, N.; Thalib, L. Global incidence of necrotizing enterocolitis: A systematic review and meta-analysis. BMC Pediatr. 2020, 20, 344. [Google Scholar] [CrossRef]
- Robinson, J.R.; Rellinger, E.J.; Hatch, L.D.; Weitkamp, J.-H.; Speck, K.E.; Danko, M.; Blakely, M.L. Surgical necrotizing enterocolitis. Semin. Perinatol. 2017, 41, 70–79. [Google Scholar] [CrossRef] [Green Version]
- van der Heide, M.; Mebius, M.J.; Bos, A.F.; Roofthooft, M.T.R.; Berger, R.M.F.; Hulscher, J.B.F.; Kooi, E.M.W. Hypoxic/ischemic hits predispose to necrotizing enterocolitis in (near) term infants with congenital heart disease: A case control study. BMC Pediatr. 2020, 20, 553. [Google Scholar] [CrossRef]
- Short, S.S.; Papillon, S.; Berel, D.; Ford, H.R.; Frykman, P.K.; Kawaguchi, A. Late onset of necrotizing enterocolitis in the full-term infant is associated with increased mortality: Results from a two-center analysis. J. Pediatr. Surg. 2014, 49, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.P.; Kota, A. Necrotizing Enterocolitis. In Pediatric Surgery, Flowcharts and Clinical Algorithms; Shehata, S., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Schnabl, K.L.; Aerde, J.E.V.; Thomson, A.B.; Clandinin, M.T. Necrotizing enterocolitis: A multifactorial disease with no cure. World J. Gastroenterol. 2008, 14, 2142–2161. [Google Scholar] [CrossRef]
- Ginglen, J.G.; Butki, N. Necrotizing Enterocolitis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Cuna, A.; Sampath, V. Genetic alterations in necrotizing enterocolitis. Semin. Perinatol. 2017, 41, 61–69. [Google Scholar] [CrossRef]
- Magistris, A.D.; Marcialis, M.A.; Puddu, M.; Dessì, A.; Irmesi, R.; Coni, E.; Fanos, V. Embryological development of the intestine and necrotizing enterocolitis. J. Pediatr. Neonat. Individ. Med. 2016, 5, e050213. [Google Scholar] [CrossRef]
- Sampah, M.E.S.; Hackam, D.J. Prenatal immunity and influences on necrotizing enterocolitis and associated neonatal disorders. Front. Immunol. 2021, 12, 650709. [Google Scholar] [CrossRef]
- Kumbhare, S.V.; Patangia, D.V.V.; Patil, R.H.; Shouche, Y.S.; Patil, N.P. Factors influencing the gut microbiome in children: From infancy to childhood. J. Biosci. 2019, 44, 49. [Google Scholar] [CrossRef]
- Khoder-Agha, F.; Kietzmann, T. The glyco-redox interplay: Principles and consequences on the role of reactive oxygen species during protein glycosylation. Redox Biol. 2021, 42, 101888. [Google Scholar] [CrossRef]
- Passaponti, S.; Pavone, V.; Cresti, L.; Ietta, F. The expression and role of glycans at the feto-maternal interface in humans. Tissue Cell 2021, 73, 101630. [Google Scholar] [CrossRef]
- Kononova, S.V. How fucose of blood group glycotopes programs human gut microbiota. Biochemistry 2017, 82, 973–989, Erratum in Biochemistry 2017, 82, 1215. [Google Scholar] [CrossRef] [Green Version]
- Kudelka, M.R.; Stowell, S.R.; Cummings, R.D.; Neish, A.S. Intestinal epithelial glycosylation in homeostasis and gut microbiota interactions in ibd. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 597–617. [Google Scholar] [CrossRef]
- Hackam, D.J.; Sodhi, C.P. Toll-like receptor-mediated intestinal inflammatory imbalance in the pathogenesis of necrotizing enterocolitis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 229–238.e1. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.J.; De Plaen, I.G. Inflammatory signaling in NEC: Role of NF-κB and cytokines. Pathophysiology 2014, 21, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.; Lakshminrusimha, S. Role of NO and other vascular mediators in the etiopathogenesis of necrotizing enterocolitis. Front. Biosci. (School Ed.) 2019, 11, 9–28. [Google Scholar] [CrossRef]
- Duci, M.; Frigo, A.C.; Visentin, S.; Verlato, G.; Gamba, P.; Fascetti-Leon, F. Maternal and placental risk factors associated with the development of necrotizing enterocolitis (NEC) and its severity. J. Pediatr. Surg. 2019, 54, 2099–2102. [Google Scholar] [CrossRef]
- Kamoji, V.M.; Dorling, J.S.; Manktelow, B.; Draper, E.S.; Field, D.J. Antenatal umbilical doppler abnormalities: An independent risk factor for early onset neonatal necrotizing enterocolitis in premature infants. Acta Paediatr. 2008, 97, 327–331. [Google Scholar] [CrossRef]
- Ahle, M.; Drott, P.; Elfvin, A.; Andersson, R.E. Maternal, fetal and perinatal factors associated with necrotizing enterocolitis in Sweden. A national case-control study. PLoS ONE 2018, 13, e0194352. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Zhang, L.; Jiang, S.; Li, M.; Yan, C.; Shen, C.; Yang, Y.; Lee, S.K.; Cao, Y. Epidemiology of necrotizing enterocolitis in preterm infants in China: A multicenter cohort study from 2015 to 2018. J. Pediatr. Surg. 2022, 57, 382–386. [Google Scholar] [CrossRef]
- Rose, A.T.; Patel, R.M. A critical analysis of risk factors for necrotizing enterocolitis. Semin. Fetal Neonatal. Med. 2018, 23, 374–379. [Google Scholar] [CrossRef]
- Wang, K.-G.; Chen, C.-Y.; Chen, Y.-Y. The effects of absent or reversed end-diastolic umbilical artery doppler flow velocity. Taiwan J. Obstet. Gynecol. 2009, 48, 225–231. [Google Scholar] [CrossRef] [Green Version]
- Wardinger, J.E.; Ambati, S. Placental Insufficiency. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Samuels, N.; van de Graaf, R.A.; de Jonge, R.C.J.; Reiss, I.K.M.; Vermeulen, M.J. Risk factors for necrotizing enterocolitis in neonates: A systematic review of prognostic studies. BMC Pediatr. 2017, 17, 105. [Google Scholar] [CrossRef]
- Aouache, R.; Biquard, L.; Vaiman, D.; Miralles, F. Oxidative stress in preeclampsia and placental diseases. Int. J. Mol. Sci. 2018, 19, 1496. [Google Scholar] [CrossRef] [Green Version]
- Ree, I.M.C.; Smits-Wintjens, V.E.H.J.; Rijntjes-Jacobs, E.G.J.; Pelsma, I.C.M.; Steggerda, S.J.; Walther, F.J.; Lopriore, E. Necrotizing Enterocolitis in Small-for-Gestational-Age Neonates: A Matched Case-Control Study. Neonatology 2014, 105, 74–78. [Google Scholar] [CrossRef]
- Yang, C.-C.; Tang, P.-L.; Liu, P.-Y.; Huang, W.-C.; Chen, Y.-Y.; Wang, H.-P.; Chang, J.-T.; Lin, L.-T. Maternal pregnancy-induced hypertension increases subsequent neonatal necrotizing enterocolitis risk. Medicine 2018, 97, e11739. [Google Scholar] [CrossRef]
- Samuel, T.M.; Sakwinska, O.; Makinen, K.; Burdge, G.C.; Godfrey, K.M.; Silva-Zolezzi, I. Preterm Birth: A Narrative Review of the Current Evidence on Nutritional and Bioactive Solutions for Risk Reduction. Nutrients 2019, 11, 1811. [Google Scholar] [CrossRef] [Green Version]
- Watson, S.N.; McElroy, S.J. Potential prenatal origins of necrotizing enterocolitis. Gastroenterol. Clin. N. Am. 2021, 50, 431–444. [Google Scholar] [CrossRef]
- Tan, X.; Zhou, Y.; Xu, L.; Zhang, L.; Wang, J.; Yang, W. The predictors of necrotizing enterocolitis in newborns with low birth weight: A retrospective analysis. Medicine 2022, 101, e28789. [Google Scholar] [CrossRef]
- Gephart, S.M.; Spitzer, A.R.; Effken, J.A.; Dodd, E.; Halpern, M.; McGrath, J.M. Discrimination of GutCheck(NEC): A clinical risk index for necrotizing enterocolitis. J. Perinatol. 2014, 34, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-P.; Zheng, M.-Y.; Xiao, Y.-Y.; Qu, Y.-M.; Wu, H. Risk factors for necrotizing enterocolitis and establishment of prediction model of necrotizing enterocolitis in preterm infants. Zhongguo Dang Dai Er Ke Za Zhi (Chin. J. Contemp. Pediatr.) 2022, 24, 41–48. [Google Scholar] [CrossRef]
- Berkhout, D.J.C.; Klaassen, P.; Niemarkt, H.J.; de Boode, W.P.; Cossey, V.; van Goudoever, J.B.; Hulzebos, C.V.; Andriessen, P.; van Kaam, A.H.; Kramer, B.W.; et al. Risk Factors for Necrotizing Enterocolitis: A Prospective Multicenter Case-Control Study. Neonatology 2018, 114, 277–284. [Google Scholar] [CrossRef]
- Kordasz, M.; Racine, M.; Szavay, P.; Lehner, M.; Krebs, T.; Luckert, C.; Hau, E.-M.; Berger, S.; Kessler, U. Risk factors for mortality in preterm infants with necrotizing enterocolitis: A retrospective multicenter analysis. Eur. J. Pediatr. 2022, 181, 933–939. [Google Scholar] [CrossRef]
- Askie, L.M.; Darlow, B.A.; Davis, P.G.; Finer, N.; Stenson, B.; Vento, M.; Whyte, R. Effects of targeting lower versus higher arterial oxygen saturations on death or disability in preterm infants. Cochrane Database Syst. Rev. 2017, 2017, CD011190. [Google Scholar] [CrossRef] [Green Version]
- Cotten, C.M.; Taylor, S.; Stoll, B.; Goldberg, R.N.; Hansen, N.I.; Sánchez, P.J.; Ambalavanan, N.; Benjamin, D.K., Jr. Prolonged duration of initial empirical antibiotic treatment is associated with increased rates of necrotizing enterocolitis and death for extremely low birth weight infants. Pediatrics 2009, 123, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Raba, A.A.; O’Sullivan, A.; Semberova, J.; Martin, A.; Miletin, J. Are antibiotics a risk factor for the development of necrotizing enterocolitis—Case-control retrospective study. Eur. J. Pediatr. 2019, 178, 923–928. [Google Scholar] [CrossRef]
- Zwittink, R.D.; van Zoeren-Grobben, D.; Martin, R.; van Lingen, R.A.; Groot Jebbink, L.J.; Boeren, S.; Renes, I.B.; van Elburg, R.M.; Belzer, C.; Knol, J. Metaproteomics reveals functional differences in intestinal microbiota development of preterm infants. Mol. Cell. Proteomics 2017, 16, 1610–1620. [Google Scholar] [CrossRef] [Green Version]
- Lemme-Dumit, J.M.; Song, Y.; Lwin, H.W.; Hernandez-Chavez, C.; Sundararajan, S.; Viscardi, R.M.; Ravel, J.; Pasetti, M.F.; Ma, B. Altered gut microbiome and fecal immune phenotype in early preterm infants with leaky gut. Front. Immunol. 2022, 13, 815046. [Google Scholar] [CrossRef]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Kling Bäckhed, H.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Dahl, C.; Stanislawski, M.; Iszatt, N.; Mandal, S.; Lozupone, C.; Clemente, J.C.; Knight, R.; Stigum, H.; Eggesbø, M. Gut microbiome of mothers delivering prematurely shows reduced diversity and lower relative abundance of Bifidobacterium and Streptococcus. PLoS ONE 2017, 12, e0184336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-L.; An, Y.; He, Y.; Hu, X.-Y.; Guo, L.; Li, Q.-Y.; Liu, L.; Li, L.-Q. Risk factors of necrotizing enterocolitis in neonates with sepsis: A retrospective case-control study. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420963818. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Heida, F.H.; Kooi, E.M.W.; Wagner, J.; Nguyen, T.-Y.; Hulscher, J.B.F.; van Zoonen, A.G.J.F.; Bos, A.F.; Harmsen, H.J.M.; de Goffau, M.C. Weight shapes the intestinal microbiome in preterm infants: Results of a prospective observational study. BMC Microbiol. 2021, 21, 219. [Google Scholar] [CrossRef]
- Russell, J.T.; Lauren Ruoss, J.; de la Cruz, D.; Li, N.; Bazacliu, C.; Patton, L.; McKinley, K.L.; Garrett, T.J.; Polin, R.A.; Triplett, E.W.; et al. Antibiotics and the developing intestinal microbiome, metabolome and inflammatory environment in a randomized trial of preterm infants. Sci. Rep. 2021, 11, 1943. [Google Scholar] [CrossRef]
- Bowker, R.M.; Yan, X.; De Plaen, I.G. Intestinal microcirculation and necrotizing enterocolitis: The vascular endothelial growth factor system. Semin. Fetal. Neonatal. Med. 2018, 23, 411–415. [Google Scholar] [CrossRef]
- Surmeli Onay, O.; Korkmaz, A.; Yigit, S.; Yurdakok, M. Hypoxic-Ischemic Enterocolitis: A proposal of a new terminology for early NEC or NEC-like disease in preterm infants, a single-center prospective observational study. Eur. J. Pediatr. 2020, 179, 561–570. [Google Scholar] [CrossRef]
- Taylor, C.T.; Colgan, S.P. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat. Rev. Immunol. 2017, 17, 774–785. [Google Scholar] [CrossRef]
- Sagrillo-Fagundes, L.; Laurent, L.; Bienvenue-Pariseault, J.; Vaillancourt, C. In vitro induction of hypoxia/reoxygenation on placental cells: A suitable model for understanding placental diseases. Methods Mol. Biol. 2018, 1710, 277–283. [Google Scholar] [CrossRef]
- Lien, Y.-C.; Zhang, Z.; Cheng, Y.; Polyak, E.; Sillers, L.; Falk, M.J.; Ischiropoulos, H.; Parry, S.; Simmons, R.A. Human placental transcriptome reveals critical alterations in inflammation and energy metabolism with fetal sex differences in spontaneous preterm birth. Int. J. Mol. Sci. 2021, 22, 7899. [Google Scholar] [CrossRef]
- Konjar, Š.; Pavšič, M.; Veldhoen, M. Regulation of oxygen homeostasis at the intestinal epithelial barrier site. Int. J. Mol. Sci. 2021, 22, 9170. [Google Scholar] [CrossRef] [PubMed]
- Belo, A.I.; van Vliet, S.J.; Maus, A.; Laan, L.C.; Nauta, T.D.; Koolwijk, P.; Tefsen, B.; van Die, I. Hypoxia inducible factor 1α down regulates cell surface expression of α1,2-fucosylated glycans in human pancreatic adenocarcinoma cells. FEBS Lett. 2015, 589, 2359–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCracken, S.A.; Seeho, S.K.M.; Carrodus, T.; Park, J.H.; Woodland, N.; Gallery, E.D.M.; Morris, J.M.; Ashton, A.W. Dysregulation of oxygen sensing/response pathways in pregnancies complicated by idiopathic intrauterine growth restriction and early-onset preeclampsia. Int. J. Mol. Sci. 2022, 23, 2772. [Google Scholar] [CrossRef] [PubMed]
- Blois, S.M.; Prince, P.D.; Borowski, S.; Galleano, M.; Barrientos, G. Placental glycoredox dysregulation associated with disease progression in an animal model of superimposed preeclampsia. Cells 2021, 10, 800. [Google Scholar] [CrossRef]
- Kulikova, G.V.; Ziganshina, M.M.; Shchegolev, A.I.; Sukhikh, G.T. Comparative characteristics of the expression of fucosylated glycans and morphometric parameters of terminal placental villi depending on the severity of preeclampsia. Bull. Exp. Biol. Med. 2021, 172, 90–95. [Google Scholar] [CrossRef]
- Ahmadian, E.; Rahbar Saadat, Y.; Hosseiniyan Khatibi, S.M.; Nariman-Saleh-Fam, Z.; Bastami, M.; Zununi Vahed, F.; Ardalan, M.; Zununi Vahed, S. Pre-Eclampsia: Microbiota possibly playing a role. Pharmacol. Res. 2020, 155, 104692. [Google Scholar] [CrossRef]
- Tsou, P.-S.; Ruth, J.H.; Campbell, P.L.; Isozaki, T.; Lee, S.; Marotte, H.; Domino, S.E.; Koch, A.E.; Amin, M.A. A Novel role for inducible Fut2 in angiogenesis. Angiogenesis 2013, 16, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Koike, Y.; Chi, L.; Ahmed, A.; Miyake, H.; Li, B.; Lee, C.; Delgado-Olguín, P.; Pierro, A. Formula feeding and immature gut microcirculation promote intestinal hypoxia, leading to necrotizing enterocolitis. Dis. Model. Mech. 2019, 12, dmm040998. [Google Scholar] [CrossRef] [Green Version]
- Koike, Y.; Li, B.; Ganji, N.; Zhu, H.; Miyake, H.; Chen, Y.; Lee, C.; Janssen Lok, M.; Zozaya, C.; Lau, E.; et al. Remote Ischemic Conditioning Counteracts the Intestinal Damage of Necrotizing Enterocolitis by Improving Intestinal Microcirculation. Nat. Commun. 2020, 11, 4950. [Google Scholar] [CrossRef]
- Henderickx, J.G.E.; Zwittink, R.D.; Renes, I.B.; van Lingen, R.A.; van Zoeren-Grobben, D.; Jebbink, L.J.G.; Boeren, S.; van Elburg, R.M.; Knol, J.; Belzer, C. Maturation of the Preterm Gastrointestinal Tract Can Be Defined by Host and Microbial Markers for Digestion and Barrier Defense. Sci. Rep. 2021, 11, 12808. [Google Scholar] [CrossRef]
- Irons, E.E.; Cortes Gomez, E.; Andersen, V.L.; Lau, J.T.Y. Bacterial colonization and TH17 immunity are shaped by intestinal sialylation in neonatal mice. Glycobiology 2022, 32, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Neal, M.D.; Siggers, R.; Sho, S.; Ma, C.; Branca, M.F.; Prindle, T., Jr.; Russo, A.M.; Afrazi, A.; Good, M.; et al. Intestinal epithelial Toll-like receptor 4 regulates goblet cell development and is required for necrotizing enterocolitis in mice. Gastroenterology 2012, 143, 708–718.e5. [Google Scholar] [CrossRef] [Green Version]
- Iijima, J.; Kobayashi, S.; Kitazume, S.; Kizuka, Y.; Fujinawa, R.; Korekane, H.; Shibata, T.; Saitoh, S.I.; Akashi-Takamura, S.; Miyake, K.; et al. Core fucose is critical for CD14-dependent Toll-like receptor 4 signals. Glycobiology 2017, 27, 1006–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Niño, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The human milk oligosaccharides 2′-fucosyllactose and 6′-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr. Res. 2021, 89, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, V.; Al Assaf, N.; Khan, R. Review of claudin proteins as potential biomarkers for necrotizing enterocolitis. Ir. J. Med. Sci. 2021, 190, 1465–1472. [Google Scholar] [CrossRef]
- Bein, A.; Eventov-Friedman, S.; Arbell, D.; Schwartz, B. Intestinal tight junctions are severely altered in NEC preterm neonates. Pediatr. Neonatol. 2018, 59, 464–473. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Lu, C.; An, L.; Gao, Q.; Xie, W.; Miao, F.; Chen, X.; Pan, Y.; Wang, Q. SIRT1 relieves necrotizing enterocolitis through inactivation of hypoxia-inducible factor (HIF)-1a. Cell Cycle 2020, 19, 2018–2027. [Google Scholar] [CrossRef]
- Högberg, N.; Stenbäck, A.; Carlsson, P.-O.; Wanders, A.; Lilja, H.E. Genes regulating tight junctions and cell adhesion are altered in early experimental necrotizing enterocolitis. J. Pediatr. Surg. 2013, 48, 2308–2312. [Google Scholar] [CrossRef]
- Nolan, L.S.; Rimer, J.M.; Good, M. The Role of Human Milk Oligosaccharides and Probiotics on the Neonatal Microbiome and Risk of Necrotizing Enterocolitis: A Narrative Review. Nutrients 2020, 12, 3052. [Google Scholar] [CrossRef]
- Yan, X.; Managlia, E.; Tan, X.-D.; De Plaen, I.G. Prenatal inflammation impairs intestinal microvascular development through a TNF-dependent mechanism and predisposes newborn mice to necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver. Physiol. 2019, 317, G57–G66. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martínez-Costa, C.; Borsch, C.; Rudloff, S. Influence of gestational age, secretor, and Lewis blood group status on the oligosaccharide content of human milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Urashima, T.; Taufik, E.; Fukuda, K.; Asakuma, S. Recent advances in studies on milk oligosaccharides of cows and other domestic farm animals. Biosci. Biotechnol. Biochem. 2013, 77, 455–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollicone, R.; Cailleau, A.; Oriol, R. Molecular genetics of H, Se, Lewis and other fucosyltransferase genes. Transfus. Clin. Biol. 1995, 2, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Meinzen-Derr, J.; Huang, P.; Schibler, K.R.; Cahill, T.; Keddache, M.; Kallapur, S.G.; Newburg, D.S.; Tabangin, M.; Warner, B.B.; et al. Fucosyltransferase 2 non-secretor and low secretor status predicts severe outcomes in premature infants. J. Pediatr. 2011, 158, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Yu, J. A study on fucosyltransferase 2 gene polymorphism and secretion status related to neonatal necrotizing enterocolitis. J. Healthc. Eng. 2021, 2021, 7219850. [Google Scholar] [CrossRef] [PubMed]
- She, X.; Du, H.; Yi, C.; He, Y.; Ai, Q.; Yu, J. The decrease of fucosylation in intestinal epithelium is related to the development of necrotizing enterocolitis. Mol. Immunol. 2021, 131, 23–32. [Google Scholar] [CrossRef]
- Demmert, M.; Schaper, A.; Pagel, J.; Gebauer, C.; Emeis, M.; Heitmann, F.; Kribs, A.; Siegel, J.; Müller, D.; Keller-Wackerbauer, A.; et al. FUT 2 polymorphism and outcome in very-low-birth-weight infants. Pediatr. Res. 2015, 77, 586–590. [Google Scholar] [CrossRef] [Green Version]
- Nuzzi, G.; Trambusti, I.; DI Cicco, M.E.; Peroni, D.G. Breast milk: More than just nutrition! Minerva Pediatr. 2021, 73, 111–114. [Google Scholar] [CrossRef]
- Sánchez, C.; Franco, L.; Regal, P.; Lamas, A.; Cepeda, A.; Fente, C. Breast Milk: A Source of Functional Compounds with Potential Application in Nutrition and Therapy. Nutrients 2021, 13, 1026. [Google Scholar] [CrossRef]
- Kononova, S.; Litvinova, E.; Vakhitov, T.; Skalinskaya, M.; Sitkin, S. Acceptive immunity: The role of fucosylated glycans in human host–microbiome interactions. Int. J. Mol. Sci. 2021, 22, 3854. [Google Scholar] [CrossRef]
- Cheema, A.S.; Trevenen, M.L.; Turlach, B.A.; Furst, A.J.; Roman, A.S.; Bode, L.; Gridneva, Z.; Lai, C.T.; Stinson, L.F.; Payne, M.S.; et al. Exclusively breastfed infant microbiota develops over time and is associated with human milk oligosaccharide intakes. Int. J. Mol. Sci. 2022, 23, 2804. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yan, J.; Wang, X.; Wang, C.; Chen, L.; Li, Y.; Chen, J.; Guo, H. Maternal fucosyltransferase 2 status associates with the profiles of human milk oligosaccharides and the fecal microbiota composition of breastfed infants. J. Agric. Food Chem. 2021, 69, 3032–3043. [Google Scholar] [CrossRef]
- Cabrera-Rubio, R.; Kunz, C.; Rudloff, S.; García-Mantrana, I.; Crehuá-Gaudiza, E.; Martínez-Costa, C.; Collado, M.C. Association of maternal secretor status and human milk oligosaccharides with milk microbiota: An observational pilot study. J. Pediatr. Gastroenterol. Nutr. 2019, 68, 256–263. [Google Scholar] [CrossRef]
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G.; Van Tassell, M.L.; Miller, M.J.; Jin, Y.S.; German, J.B.; et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015, 3, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldwell, J.; Matson, A.; Mosha, M.; Hagadorn, J.I.; Moore, J.; Brownell, E. Maternal H-antigen secretor status is an early biomarker for potential preterm delivery. J. Perinatol. 2021, 41, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wu, R.Y.; Horne, R.G.; Ahmed, A.; Lee, D.; Robinson, S.C.; Zhu, H.; Lee, C.; Cadete, M.; Johnson-Henry, K.C.; et al. Human Milk Oligosaccharides Protect against Necrotizing Enterocolitis by Activating Intestinal Cell Differentiation. Mol. Nutr. Food Res. 2020, 64, e2000519. [Google Scholar] [CrossRef]
- Becker, D.J.; Lowe, J.B. Fucose: Biosynthesis and biological function in mammals. Glycobiology 2003, 13, 41R–53R. [Google Scholar] [CrossRef]
- Keeley, T.S.; Yang, S.; Lau, E. The diverse contributions of fucose linkages in cancer. Cancers 2019, 11, 1241. [Google Scholar] [CrossRef] [Green Version]
- Honas, B.J.; Glassman, U.M.; Wiese, T.J. Enzymatic activity of alpha-L-fucosidase and L-fucokinase across vertebrate animal species. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 153, 359–364. [Google Scholar] [CrossRef] [Green Version]
- Sosicka, P.; Ng, B.G.; Wong, M.; Xia, Z.J.; Scott, D.; Lebrilla, C.B.; Freeze, H.H. Novel insights into the fucose metabolism–challenging the old dogma. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Park, D.; Ryu, K.S.; Choi, D.; Kwak, J.; Park, C. Characterization and role of fucose mutarotase in mammalian cells. Glycobiology 2007, 17, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Skurska, E.; Szulc, B.; Maszczak-Seneczko, D.; Wiktor, M.; Wiertelak, W.; Makowiecka, A.; Olczak, M. Incorporation of fucose into glycans independent of the GDP-fucose transporter SLC35C1 preferentially utilizes salvaged over de novo GDP-fucose. J. Biol. Chem. 2022, 298, 102206. [Google Scholar] [CrossRef]
- Xu, Y.X.; Ma, A.; Liu, L. Transforming growth factor β signaling upregulates the expression of human GDP-fucose transporter by activating transcription factor Sp1. PLoS ONE 2013, 8, e74424. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, Y.; Zhang, L.; Qian, W.; Hou, X.; Lin, R. Exogenous l-fucose protects the intestinal mucosal barrier depending on upregulation of FUT2-mediated fucosylation of intestinal epithelial cells. FASEB J. 2021, 35, e21699. [Google Scholar] [CrossRef]
- De Leoz, M.L.; Gaerlan, S.C.; Strum, J.S.; Dimapasoc, L.M.; Mirmiran, M.; Tancredi, D.J.; Smilowitz, J.T.; Kalanetra, K.M.; Mills, D.A.; German, J.B.; et al. Lacto-N-tetraose, fucosylation, and secretor status are highly variable in human milk oligosaccharides from women delivering preterm. J. Proteome Res. 2012, 11, 4662–4672. [Google Scholar] [CrossRef] [Green Version]
- Jilling, T.; Ambalavanan, N.; Cotton, C.M.; Martin, C.A.; Maheshwari, A.; Schibler, K.; Levy, J.; Page, G.P. Surgical necrotizing enterocolitis in extremely premature neonates is associated with genetic variations in an intergenic region of chromosome 8. Pediatr. Res. 2018, 83, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Thomson, P.; Medina, D.A.; Garrido, D. Human milk oligosaccharides and infant gut bifidobacteria: Molecular strategies for their utilization. Food Microbiol. 2018, 75, 37–46. [Google Scholar] [CrossRef]
- Sakanaka, M.; Gotoh, A.; Yoshida, K.; Odamaki, T.; Koguchi, H.; Xiao, J.Z.; Kitaoka, M.; Katayama, T. Varied Pathways of Infant Gut-Associated Bifidobacterium to Assimilate Human Milk Oligosaccharides: Prevalence of the Gene Set and its Correlation with Bifidobacteria-Rich Microbiota Formation. Nutrients 2019, 12, 71. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Díaz, J.; Rubio-del-Campo, A.; Yebra, M.J. Lactobacillus casei ferments the N-Acetylglucosamine moiety of fucosyl-α-1,3-N-acetylglucosamine and excretes L-fucose. Appl. Environ. Microbiol. 2012, 78, 4613–4619. [Google Scholar] [CrossRef] [Green Version]
- Hiltunen, H.; Collado, M.C.; Ollila, H.; Kola, T.; Tölkkö, S.; Isolauri, E.; Salminen, S.; Rautava, S. Spontaneous preterm delivery is reflected in both early neonatal and maternal gut microbiota. Pediatr. Res. 2022, 91, 1804–1811. [Google Scholar] [CrossRef]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, F.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-infant microbial transmission from different body sites shapes the developing infant gut microbiome. Cell Host Microbe 2018, 24, 133–145.e5. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Guo, R.; Li, S.; Liang, F.; Tian, C.; Zhao, X.; Long, Y.; Liu, F.; Jiang, M.; Zhang, Y.; et al. Systematic analysis of gut microbiota in pregnant women and its correlations with individual heterogeneity. Npj Biofilms Microbiomes 2020, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Wacklin, P.; Nakphaichit, M.; Loyttyniemi, E.; Chowdhury, S.; Shouche, Y.; Mättö, J.; Isolauri, E.; Salminen, S. Secretor status is strongly associated with microbial alterations observed during pregnancy. PLoS ONE 2015, 10, e0134623. [Google Scholar] [CrossRef]
- Liu, J.; Yang, H.; Yin, Z.; Jiang, X.; Zhong, H.; Qiu, D.; Zhu, F.; Li, R. Remodeling of the gut microbiota and structural shifts in Preeclampsia patients in South China. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.-J.; Li, S.-H.; Li, S.-C.; Zhong, Z.-C.; Duan, H.-L.; Tian, C.; Li, H.; He, W.; Chen, M.-C.; He, T.-W.; et al. Early-onset preeclampsia is associated with gut microbial alterations in antepartum and postpartum women. Front. Cell. Infect. Microbiol. 2019, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- DiGiulio, D.B.; Callahan, B.J.; McMurdie, P.J.; Costello, E.K.; Lyell, D.J.; Robaczewska, A.; Sun, C.L.; Goltsman, D.S.A.; Wong, R.J.; Shaw, G.; et al. Temporal and spatial variation of the human microbiota during pregnancy. Proc. Natl. Acad. Sci. USA 2015, 112, 11060–11065. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; DiGiulio, D.B.; Goltsman, D.S.A.; Sun, C.L.; Costello, E.K.; Jeganathan, P.; Biggio, J.R.; Wong, R.J.; Druzin, M.L.; Shaw, G.M.; et al. Replication and refinement of a vaginal microbial signature of preterm birth in two racially distinct cohorts of US women. Proc. Natl. Acad. Sci. USA 2017, 114, 9966–9971. [Google Scholar] [CrossRef] [Green Version]
- Zakaria, Z.Z.; Al-Rumaihi, S.; Al-Absi, R.S.; Farah, H.; Elamin, M.; Nader, R.; Bouabidi, S.; Suleiman, S.E.; Nasr, S.; Al-Asmakh, M. Physiological changes and interactions between microbiome and the host during pregnancy. Front. Cell. Infect. Microbiol. 2022, 12, 824925. [Google Scholar] [CrossRef]
- Gough, E.K.; Edens, T.J.; Geum, H.M.; Baharmand, I.; Gill, S.K.; Robertson, R.C.; Mutasa, K.; Ntozini, R.; Smith, L.E.; Chasekwa, B.; et al. Maternal fecal microbiome predicts gestational age, birth weight and neonatal growth in rural Zimbabwe. eBioMedicine 2021, 68, 103421. [Google Scholar] [CrossRef]
- Li, D.; Huang, Y.; Sadykova, A.; Zheng, W.; Lin, L.; Jin, C.; Zhong, W.; Liao, C.; Pan, S. Composition of the microbial communities at different body sites in women with preterm birth and their newborns. Med. Microecol. 2021, 9, 100046. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Z.-H.; Yang, J.; Wei, Y.; Wang, X.-Y.; Zhao, Y.-Y. Gut microbiota dysbiosis in preeclampsia patients in the second and third trimesters. Chin. Med. J. 2020, 133, 1057–1065. [Google Scholar] [CrossRef]
- Qing, W.; Shi, Y.; Zhou, H.; Chen, M. Gut microbiota dysbiosis in patients with preeclampsia: A systematic review. Med. Microecol. 2021, 10, 100047. [Google Scholar] [CrossRef]
- Ishimwe, J.A. Maternal microbiome in preeclampsia pathophysiology and implications on offspring health. Physiol. Rep. 2021, 9, e14875. [Google Scholar] [CrossRef] [PubMed]
- Miao, T.; Yu, Y.; Sun, J.; Ma, A.; Yu, J.; Cui, M.; Yang, L.; Wang, H. Decrease in abundance of bacteria of the genus Bifidobacterium in gut microbiota may be related to pre-eclampsia progression in women from East China. Food. Nutr. Res. 2021, 65. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, P.; Liu, M.; Zheng, H.; He, Y.; Chen, M.-X.; Tang, W.; Yue, X.; Huang, Y.; Zhuang, L.; et al. Gut dysbiosis induces the development of pre-eclampsia through bacterial translocation. Gut 2020, 69, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gu, X.; Yang, J.; Wei, Y.; Zhao, Y. Gut microbiota dysbiosis and increased plasma LPS and TMAO levels in patients with preeclampsia. Front. Cell. Infect. Microbiol. 2019, 9, 409. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Dekker Nitert, M. Increased systolic and diastolic blood pressure is associated with altered gut microbiota composition and butyrate production in early pregnancy. Hypertension 2016, 68, 974–981. [Google Scholar] [CrossRef]
- Huang, L.; Cai, M.; Li, L.; Zhang, X.; Xu, Y.; Xiao, J.; Huang, Q.; Luo, G.; Zeng, Z.; Jin, C.; et al. Gut microbiota changes in preeclampsia, abnormal placental growth and healthy pregnant women. BMC Microbiol. 2021, 21, 265. [Google Scholar] [CrossRef]
- Susic, D.F.; Wang, L.; Roberts, L.M.; Bai, M.; Gia, A.; McGovern, E.; Jiang, X.-T.; Davis, G.K.; El-Omar, E.; Henry, A. The P4 study: Postpartum maternal and infant faecal microbiome 6 months after hypertensive versus normotensive pregnancy. Front. Cell. Infect. Microbiol. 2022, 12, 646165. [Google Scholar] [CrossRef]
- Jin, J.; Gao, L.; Zou, X.; Zhang, Y.; Zheng, Z.; Zhang, X.; Li, J.; Tian, Z.; Wang, X.; Gu, J.; et al. Gut dysbiosis promotes preeclampsia by regulating macrophages and trophoblasts. Circ. Res. 2022, 131, 492–506. [Google Scholar] [CrossRef]
- Li, P.; Wang, H.; Guo, L.; Gou, X.; Chen, G.; Lin, D.; Fan, D.; Guo, X.; Liu, Z. Association between gut microbiota and preeclampsia-eclampsia: A two-sample mendelian randomization study. BMC Med. 2022, 20, 443. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.-J.; Li, S.-H.; Wen, J.-Y.; Wang, G.-Y.; Li, H.; He, T.-W.; Lv, Q.-B.; Xiao, M.-C.; Duan, H.-L.; Chen, M.-C.; et al. Deep metagenomic characterization of gut microbial community and function in preeclampsia. Front. Cell. Infect. Microbiol. 2022, 12, 933523. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Chen, J.; Ma, S.; An, R.; Li, X.; Tan, H. The Association between Gut Microbiome and Pregnancy-Induced Hypertension: A Nested Case–Control Study. Nutrients 2022, 14, 4582. [Google Scholar] [CrossRef]
- Tu, X.; Duan, C.; Lin, B.; Li, K.; Gao, J.; Yan, H.; Wang, K.; Zhao, Z. Characteristics of the gut microbiota in pregnant women with fetal growth restriction. BMC Pregnancy Childbirth 2022, 22, 297. [Google Scholar] [CrossRef]
- Yang, J.; Hou, L.; Wang, J.; Xiao, L.; Zhang, J.; Yin, N.; Yao, S.; Cheng, K.; Zhang, W.; Shi, Z.; et al. Unfavourable intrauterine environment contributes to abnormal gut microbiome and metabolome in twins. Gut 2022, 71, 2451–2462. [Google Scholar] [CrossRef]
- Sun, Y.; Li, L.; Song, J.; Mao, W.; Xiao, K.; Jiang, C. Intrauterine hypoxia changed the colonization of the gut microbiota in newborn rats. Front. Pediatr. 2021, 9, 675022. [Google Scholar] [CrossRef]
- Tarracchini, C.; Milani, C.; Longhi, G.; Fontana, F.; Mancabelli, L.; Pintus, R.; Lugli, G.A.; Alessandri, G.; Anzalone, R.; Viappiani, A.; et al. Unraveling the microbiome of necrotizing enterocolitis: Insights in novel microbial and metabolomic biomarkers. Microbiol. Spectr. 2021, 9, e01176-21. [Google Scholar] [CrossRef]
- Arboleya, S.; Rios-Covian, D.; Maillard, F.; Langella, P.; Gueimonde, M.; Martín, R. Preterm delivery: Microbial dysbiosis, gut inflammation and hyperpermeability. Front. Microbiol. 2022, 12, 806338. [Google Scholar] [CrossRef]
- Hiltunen, H.; Hanani, H.; Luoto, R.; Turjeman, S.; Ziv, O.; Isolauri, E.; Salminen, S.; Koren, O.; Rautava, S. Preterm infant meconium microbiota transplant induces growth failure, inflammatory activation, and metabolic disturbances in germ-free mice. Cell. Rep. Med. 2021, 2, 100447. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Benítez, M.V.; Gámez-Belmonte, R.; Gil-Campos, M.; Hernández-Chirlaque, C.; Bouzas, P.R.; Sánchez de Medina, F.; Martínez-Augustin, O. Premature birth infants present elevated inflammatory markers in the meconium. Front. Pediatr. 2021, 8, 627475. [Google Scholar] [CrossRef]
- Heida, F.H.; van Zoonen, A.G.J.F.; Hulscher, J.B.F.; Te Kiefte, B.J.C.; Wessels, R.; Kooi, E.M.W.; Bos, A.F.; Harmsen, H.J.M.; de Goffau, M.C. A Necrotizing Enterocolitis-Associated Gut Microbiota Is Present in the Meconium: Results of a Prospective Study. Clin. Infect. Dis. 2016, 62, 863–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopp, J.; Ferretti, P.; Meyer, C.U.; Hilbert, K.; Haiß, A.; Marißen, J.; Henneke, P.; Hudalla, H.; Pirr, S.; Viemann, D.; et al. Meconium microbiome of very preterm infants across Germany. mSphere 2022, 7, e00808-21. [Google Scholar] [CrossRef] [PubMed]
- Wandro, S.; Osborne, S.; Enriquez, C.; Bixby, C.; Arrieta, A.; Whiteson, K. The microbiome and metabolome of preterm infant stool are personalized and not driven by health outcomes, including necrotizing enterocolitis and late-onset sepsis. mSphere 2018, 3, e00104-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.-Y.; Chiang Chiau, J.-S.; Ho, Y.-H.; Chang, J.-H.; Tsai, K.-N.; Liu, C.-Y.; Hsu, C.-H.; Lin, C.-Y.; Ko, M.H.-J.; Lee, H.-C. Impact of early empiric antibiotic regimens on the gut microbiota in very low birth weight preterm infants: An observational study. Front. Pediatr. 2021, 9, 651713. [Google Scholar] [CrossRef]
- Zwittink, R.D.; van Zoeren-Grobben, D.; Renes, I.B.; van Lingen, R.A.; Norbruis, O.F.; Martin, R.; Groot Jebbink, L.J.; Knol, J.; Belzer, C. Dynamics of the bacterial gut microbiota in preterm and term infants after intravenous amoxicillin/ceftazidime treatment. BMC Pediatr. 2020, 20, 195. [Google Scholar] [CrossRef]
- La Rosa, P.S.; Warner, B.B.; Zhou, Y.; Weinstock, G.M.; Sodergren, E.; Hall-Moore, C.M.; Stevens, H.J.; Bennett, W.E.; Shaikh, N.; Linneman, L.A.; et al. Patterned progression of bacterial populations in the premature infant gut. Proc. Natl. Acad. Sci. USA 2014, 111, 12522–12527. [Google Scholar] [CrossRef] [Green Version]
- Moles, L.; Gómez, M.; Heilig, H.; Bustos, G.; Fuentes, S.; de Vos, W.; Fernández, L.; Rodríguez, J.M.; Jiménez, E. Bacterial diversity in meconium of preterm neonates and evolution of their fecal microbiota during the first month of life. PLoS ONE 2013, 8, e66986. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Lopez, M.; Dinsmoor, A.M.; Ho, T.T.B.; Donovan, S.M. A Systematic Review of the Factors influencing microbial colonization of the preterm infant gut. Gut Microbes 2021, 13, 1884514. [Google Scholar] [CrossRef]
- Yu, Y.; Lu, L.; Sun, J.; Petrof, E.O.; Claud, E.C. Preterm infant gut microbiota affects intestinal epithelial development in a humanized microbiome gnotobiotic mouse model. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G521–G532. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.T.B.; Groer, M.W.; Kane, B.; Yee, A.L.; Torres, B.A.; Gilbert, J.A.; Maheshwari, A. Dichotomous development of the gut microbiome in preterm infants. Microbiome 2018, 6, 157. [Google Scholar] [CrossRef]
- Rohmer, L.; Hocquet, D.; Miller, S.I. Are pathogenic bacteria just looking for food? Metabolism and microbial pathogenesis. Trends Microbiol. 2011, 19, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, E.R.; Maiden, M.C.; Gupta, S. Metabolic competition as a driver of bacterial population structure. Future Microbiol. 2016, 11, 1339–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosny, M.; Baptiste, E.; Levasseur, A.; La Scola, B. Molecular epidemiology of clostridium neonatale and its relationship with the occurrence of necrotizing enterocolitis in preterm neonates. New Microbes New Infect. 2019, 32, 100612. [Google Scholar] [CrossRef] [PubMed]
- Petrova, N.A.; Kaplina, A.V.; Khavkin, A.I.; Pervunina, T.M.; Komlichenko, E.V.; Nikiforov, V.G.; Sitkin, S.I. Necrotizing enterocolitis: Current concepts of etiopathogenesis with an emphasis on microbiome and metabolomics. Vopr. Prakt. Pediatr. (Clin. Pract. Pediatr.) 2021, 16, 98–105. [Google Scholar] [CrossRef]
- Dobbler, P.T.; Procianoy, R.S.; Mai, V.; Silveira, R.C.; Corso, A.L.; Rojas, B.S.; Roesch, L.F.W. Low microbial diversity and abnormal microbial succession is associated with necrotizing enterocolitis in preterm infants. Front. Microbiol. 2017, 8, 2243. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Du, W.; Xiao, S.; Zeng, B.; She, X.; Liu, D.; Du, H.; Li, L.; Li, F.; Ai, Q.; et al. Colonization of fecal microbiota from patients with neonatal necrotizing enterocolitis exacerbates intestinal injury in germfree mice subjected to necrotizing enterocolitis-induction protocol via alterations in butyrate and regulatory T cells. J. Transl. Med. 2021, 19, 510. [Google Scholar] [CrossRef]
- Lindberg, T.P.; Caimano, M.J.; Hagadorn, J.I.; Bennett, E.M.; Maas, K.; Brownell, E.A.; Matson, A.P. Preterm infant gut microbial patterns related to the development of necrotizing enterocolitis. J. Matern. Fetal Neonatal Med. 2020, 33, 349–358. [Google Scholar] [CrossRef]
- Olm, M.R.; Bhattacharya, N.; Crits-Christoph, A.; Firek, B.A.; Baker, R.; Song, Y.S.; Morowitz, M.J.; Banfield, J.F. Necrotizing enterocolitis is preceded by increased gut bacterial replication, Klebsiella, and fimbriae-encoding bacteria. Sci. Adv. 2019, 5, eaax5727. [Google Scholar] [CrossRef] [Green Version]
- Warner, B.B.; Deych, E.; Zhou, Y.; Hall-Moore, C.; Weinstock, G.M.; Sodergren, E.; Shaikh, N.; Hoffmann, J.A.; Linneman, L.A.; Hamvas, A.; et al. Gut bacteria dysbiosis and necrotising enterocolitis in very low birthweight infants: A prospective case-control study. Lancet 2016, 387, 1928–1936. [Google Scholar] [CrossRef] [Green Version]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J.; et al. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.V.; Scholz, M.; Zolfo, M.; Taft, D.H.; Schibler, K.R.; Tett, A.; Segata, N.; Morrow, A.L. Metagenomic sequencing with strain-level resolution implicates uropathogenic E. coli in necrotizing enterocolitis and mortality in preterm infants. Cell. Rep. 2016, 14, 2912–2924. [Google Scholar] [CrossRef] [Green Version]
- Fahey, R.C.; Brown, W.C.; Adams, W.B.; Worsham, M.B. Occurrence of glutathione in bacteria. J. Bacteriol. 1978, 133, 1126–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano-Keeler, J.; Shilts, M.H.; Tovchigrechko, A.; Wang, C.; Brucker, R.M.; Moore, D.J.; Fonnesbeck, C.; Meng, S.; Correa, H.; Lovvorn, H.N.; et al. Distinct mucosal microbial communities in infants with surgical necrotizing enterocolitis correlate with age and antibiotic exposure. PLoS ONE 2018, 13, e0206366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozé, J.-C.; Ancel, P.-Y.; Lepage, P.; Martin-Marchand, L.; Al Nabhani, Z.; Delannoy, J.; Picaud, J.-C.; Lapillonne, A.; Aires, J.; Durox, M.; et al. Nutritional strategies and gut microbiota composition as risk factors for necrotizing enterocolitis in very-preterm infants. Am. J. Clin. Nutr. 2017, 106, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.-Y.; Li, L.-Q.; Yang, T.; She, X.; Ai, Q.; Wang, Z.-L. Autoinducer-2 may be a new biomarker for monitoring neonatal necrotizing enterocolitis. Front. Cell. Infect. Microbiol. 2020, 10, 140. [Google Scholar] [CrossRef]
- Kaiko, G.E.; Ryu, S.H.; Koues, O.I.; Collins, P.L.; Solnica-Krezel, L.; Pearce, E.J.; Pearce, E.L.; Oltz, E.M.; Stappenbeck, T.S. The colonic crypt protects stem cells from microbiota-derived metabolites. Cell 2016, 165, 1708–1720. [Google Scholar] [CrossRef] [Green Version]
- Salvi, P.S.; Cowles, R.A. Butyrate and the intestinal epithelium: Modulation of proliferation and inflammation in homeostasis and disease. Cells 2021, 10, 1775. [Google Scholar] [CrossRef]
- Zhou, Y.; Shan, G.; Sodergren, E.; Weinstock, G.; Walker, W.A.; Gregory, K.E. longitudinal analysis of the premature infant intestinal microbiome prior to necrotizing enterocolitis: A case-control study. PLoS ONE 2015, 10, e0118632. [Google Scholar] [CrossRef]
- Fu, X.; Li, S.; Jiang, Y.; Hu, X.; Wu, H. Necrotizing enterocolitis and intestinal microbiota: The timing of disease and combined effects of multiple species. Front. Pediatr. 2021, 9, 657349. [Google Scholar] [CrossRef]
- Kaelin, E.A.; Rodriguez, C.; Hall-Moore, C.; Hoffmann, J.A.; Linneman, L.A.; Ndao, I.M.; Warner, B.B.; Tarr, P.I.; Holtz, L.R.; Lim, E.S. Longitudinal gut virome analysis identifies specific viral signatures that precede necrotizing enterocolitis onset in preterm infants. Nat. Microbiol. 2022, 7, 653–662. [Google Scholar] [CrossRef]
- Jayasinghe, T.N.; Vatanen, T.; Chiavaroli, V.; Jayan, S.; McKenzie, E.J.; Adriaenssens, E.; Derraik, J.G.B.; Ekblad, C.; Schierding, W.; Battin, M.R.; et al. Differences in compositions of gut bacterial populations and bacteriophages in 5–11 year-olds born preterm compared to full term. Front. Cell. Infect. Microbiol. 2020, 10, 276. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Guo, Q.; Ran, Y.; Lin, L.; Chen, P.; He, J.; Chen, Y.; Wen, J. Multiomics Study Reveals Enterococcus and Subdoligranulum Are Beneficial to Necrotizing Enterocolitis. Front. Microbiol. 2021, 12, 3196. [Google Scholar] [CrossRef] [PubMed]
- Federici, S.; Kredo-Russo, S.; Valdés-Mas, R.; Kviatcovsky, D.; Weinstock, E.; Matiuhin, Y.; Silberberg, Y.; Atarashi, K.; Furuichi, M.; Oka, A.; et al. Targeted suppression of human IBD-associated gut microbiota commensals by phage consortia for treatment of intestinal inflammation. Cell 2022, 185, 2879–2898.e24. [Google Scholar] [CrossRef] [PubMed]
- Renwick, V.L.; Stewart, C.J. Exploring functional metabolites in preterm infants. Acta Paediatr. 2022, 111, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Lagomarcino, A.J.; Schibler, K.R.; Taft, D.H.; Yu, Z.; Wang, B.; Altaye, M.; Wagner, M.; Gevers, D.; Ward, D.V.; et al. Early microbial and metabolomic signatures predict later onset of necrotizing enterocolitis in preterm infants. Microbiome 2013, 1, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomaidou, A.; Chatziioannou, A.C.; Deda, O.; Benaki, D.; Gika, H.; Mikros, E.; Agakidis, C.; Raikos, N.; Theodoridis, G.; Sarafidis, K. A Pilot case-control study of urine metabolomics in preterm neonates with necrotizing enterocolitis. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1117, 10–21. [Google Scholar] [CrossRef]
- Stewart, C.J.; Embleton, N.D.; Marrs, E.C.L.; Smith, D.P.; Nelson, A.; Abdulkadir, B.; Skeath, T.; Petrosino, J.F.; Perry, J.D.; Berrington, J.E.; et al. Temporal bacterial and metabolic development of the preterm gut reveals specific signatures in health and disease. Microbiome 2016, 4, 67. [Google Scholar] [CrossRef] [Green Version]
- Brehin, C.; Dubois, D.; Dicky, O.; Breinig, S.; Oswald, E.; Serino, M. Evolution of gut microbiome and metabolome in suspected necrotizing enterocolitis: A case-control study. J. Clin. Med. 2020, 9, 2278. [Google Scholar] [CrossRef]
- Liu, X.-C.; Du, T.-T.; Gao, X.; Zhao, W.-J.; Wang, Z.-L.; He, Y.; Bao, L.; Li, L.-Q. Gut microbiota and short-chain fatty acids may be new biomarkers for predicting neonatal necrotizing enterocolitis: A pilot study. Front. Microbiol. 2022, 13, 969656. [Google Scholar] [CrossRef]
- Ji, J.; Ling, X.B.; Zhao, Y.; Hu, Z.; Zheng, X.; Xu, Z.; Wen, Q.; Kastenberg, Z.J.; Li, P.; Abdullah, F.; et al. A Data-Driven Algorithm Integrating Clinical and Laboratory Features for the Diagnosis and Prognosis of Necrotizing Enterocolitis. PLoS ONE 2014, 9, e89860. [Google Scholar] [CrossRef] [Green Version]
- Vance, D.; Frese, S.A.; Casaburi, G. Artificial intelligence accurately predicts necrotizing enterocolitis from the healthy preterm infant gut microbiome. Pediatrics 2019, 144, 697. [Google Scholar] [CrossRef]
- Lure, A.C.; Du, X.; Black, E.W.; Irons, R.; Lemas, D.J.; Taylor, J.A.; Lavilla, O.; de la Cruz, D.; Neu, J. Using machine learning analysis to assist in differentiating between necrotizing enterocolitis and spontaneous intestinal perforation: A novel predictive analytic tool. J. Pediatr. Surg. 2021, 56, 1703–1710. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Salleb-Aouissi, A.; Hooven, T.A. Interpretable prediction of necrotizing enterocolitis from machine learning analysis of premature infant stool microbiota. BMC Bioinform. 2022, 23, 104. [Google Scholar] [CrossRef] [PubMed]
- Gephart, S.M.; Underwood, M.A.; Rosito, S.; Kim, J.H.; Caplan, M.S. Grading the evidence to identify strategies to modify risk for necrotizing enterocolitis. Pediatr. Res. 2020, 88, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Maheshwari, A.; Neu, J.; Ei-Saie, A.; Pammi, M. An Overview of Systematic Reviews of Randomized-Controlled Trials for Preventing Necrotizing Enterocolitis in Preterm Infants. Neonatology 2020, 117, 46–56. [Google Scholar] [CrossRef]
- Chandran, S.; Anand, A.J.; Rajadurai, V.S.; Seyed, E.S.; Khoo, P.C.; Chua, M.C. Evidence-Based Practices Reduce Necrotizing Enterocolitis and Improve Nutrition Outcomes in Very Low-Birth-Weight Infants. J. Parenter. Enter. Nutr. 2021, 45, 1408–1416. [Google Scholar] [CrossRef]
- Neu, J. Necrotizing Enterocolitis: The Future. Neonatology 2020, 117, 240–244. [Google Scholar] [CrossRef]
- Gill, E.M.; Jung, K.; Qvist, N.; Ellebæk, M.B. Antibiotics in the medical and surgical treatment of necrotizing enterocolitis. A systematic review. BMC Pediatr. 2022, 22, 66. [Google Scholar] [CrossRef]
- Bering, S.B. Human Milk Oligosaccharides to Prevent Gut Dysfunction and Necrotizing Enterocolitis in Preterm Neonates. Nutrients 2018, 10, 1461. [Google Scholar] [CrossRef] [Green Version]
- Orczyk-Pawiłowicz, M.; Lis-Kuberka, J. The Impact of Dietary Fucosylated Oligosaccharides and Glycoproteins of Human Milk on Infant Well-Being. Nutrients 2020, 12, 1105. [Google Scholar] [CrossRef]
- Vongbhavit, K.; Underwood, M.A. Prevention of Necrotizing Enterocolitis Through Manipulation of the Intestinal Microbiota of the Premature Infant. Clin. Ther. 2016, 38, 716–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C.; Fente, C.; Regal, P.; Lamas, A.; Lorenzo, M.P. Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review. Foods 2021, 10, 1429. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Somatic Factors | Factors Related to Nutrition |
|---|---|
| Lower birth weight [22,23,27] | Formula feeding [36] |
| Gestational age at birth | Breast milk fortification [35] |
| Intrauterine growth retardation [20,29] | |
| Genetic predisposition [9] | |
| Factors associated with hypoxia | Factors associated with tissue perfusion |
| Persistent ductus arteriosus [22] | Arterial hypotension [27] |
| Red-blood-cell transfusion [35] | Hematocrit > 49.65% and mean corpuscular volume > 114.35 fl [35] |
| Apgar score < 7 at 5 min [22] Apgar score < 6 for 1 min, <7 for 5 min, and <8 for 10 min increases the risk of mortality in NEC [37] | Congenital heart disease [33] |
| Resuscitation in the delivery room | Persistent ductus arteriosus [24] |
| Assisted ventilation [27] | Pre-eclampsia (?) |
| Lower target oxygen saturation [38] | Abnormal blood flow in the umbilical artery prenatally [20,21] |
| Isoimmunization [22] | Placental abruption [24] |
| Intracranial hemorrhage ≥ Grade II [35] | |
| Higher blood lactate level [37] | |
| Factors associated with infection | Gut-microbiota-related factors |
| Maternal chorioamnionitis [20] | Long-term antibiotic therapy in children [39,40] Impaired intestinal colonization in the early neonatal period [41,42] |
| Premature rupture of membranes (?) | Features of the intestinal microbiota of the mother depending on the duration of pregnancy [43,44] |
| Sepsis [33,36,45] | Disruption of gut microbial colonization due to prematurity, birth by caesarean section [46,47], or formula feeding [48] |
| Bacterial infection [22,34] | Administration of H2 blockers that suppress acidity [34] |
| № | Authors, PMID/doi | Setting and Time Period | Participants and Study Design | Methods | Features of Microbiome |
|---|---|---|---|---|---|
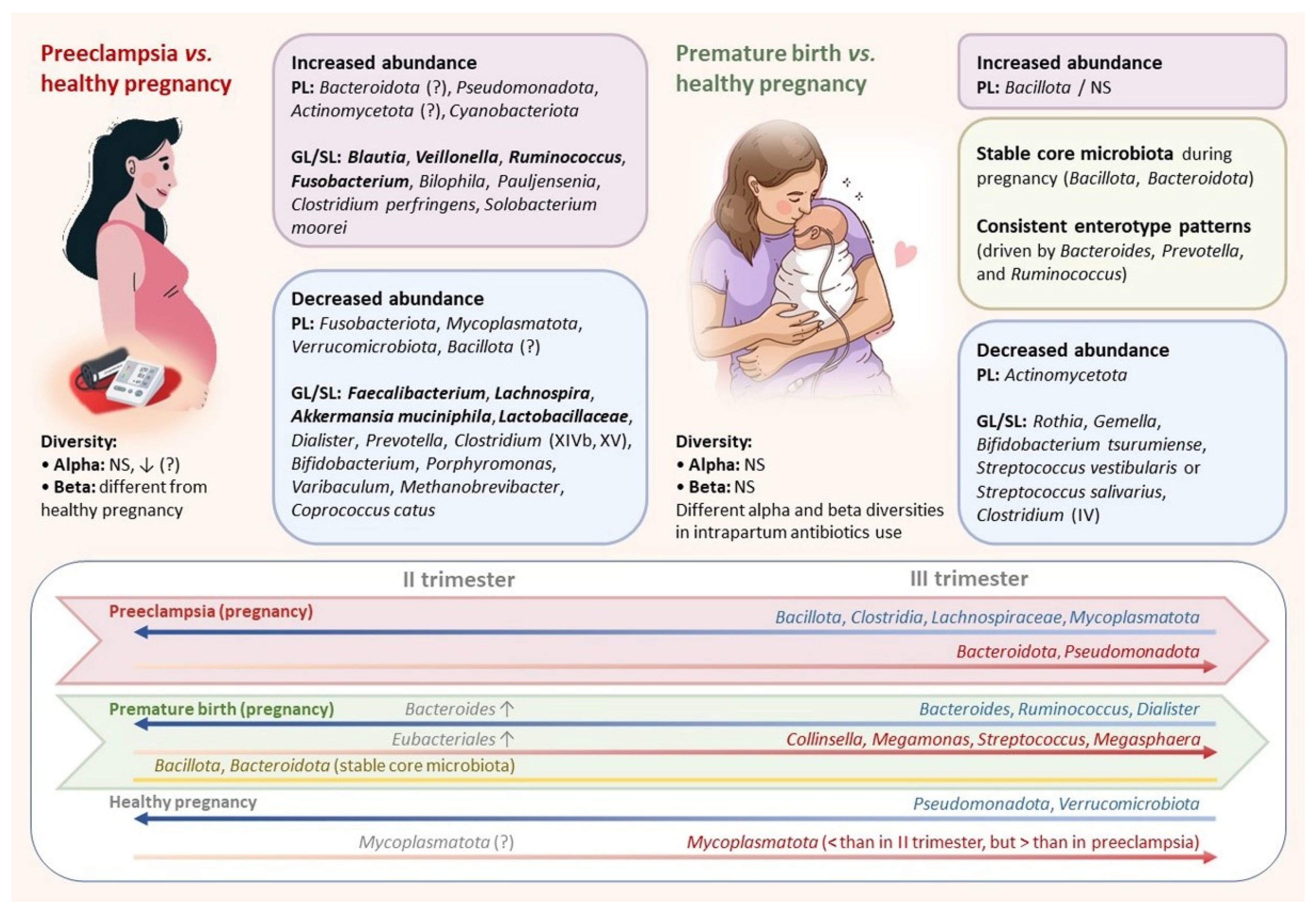
| 1 | Dahl C. (2017) [44] PMID: 29069100 | The Norwegian Microbiota Study (NoMIC) County hospital (Sykehuset Østfold), Norway Study period: 2002–2005. | Cases: 19 mothers delivering prematurely (<259 days of gestation). Controls: 102 mothers of term neonates. Conditions: vaginal delivery; no reported antibiotic use on or after the day of delivery. Fecal samples were collected on the 4th day postpartum. | 16S ribosomal RNA gene (V4 region) sequencing. Bioinformatics analysis: QIIME version 1.7.0. | The maternal-gut-microbiota profiles (4th postpartum day): Phylum level: women delivering prematurely had more Bacillota (87% vs. 81%), but less Actinomycetota (6% vs. 10%). Family level: a lower median abundance of the families Streptococcaceae and Bifidobacteriaceae (NS when applying correction for multiple testing). Four OTUs had a significantly lower abundance in mothers of preterm deliveries compared to term: OTU1142029 (NCBI BLAST: B. tsurumiense), OTU4425214 (NCBI BLAST: S.vestibularis or S. salivarius), OTU4412546 (Oscillospiraceae, Clostridium cluster IV), and OTU208539 (Mogibacteriaceae, Clostridium Family XIII Incertae Sedis). Diversity: No difference in the maternal beta diversity of preterm vs. full-term deliveries. One IQR increase in Shannon diversity was associated 38% (95% CI: 1%, 61%) lower odds of having a spontaneous preterm birth. |
| 2 | Yang H. (2020) [105] PMID: 32917878 | Guangzhou Women and Children’s Medical Center, Guangzhou, China Population-level investigation Study period: January 2017–September 2017. | Cases: 1479 pregnant women of Chinese origin from the 9th week of gestation to antepartum (>36th week). Fecal samples were collected at one time point; no longitudinal data. Controls: 1048 nonpregnant women from the Guangdong Gut Microbiome Project (GGMP) cohort. | 16S ribosomal RNA gene (V4 region) sequencing. Bioinformatics analysis: QIIME2; the taxonomy was formed by using the Greengenes v.13.8 database. | Pregnant women exhibited consistent enterotype patterns with normal adults, which were driven by the abundance of dominant genera (Bacteroides, Prevotella, and Ruminococcus). This enterotype composition was relatively stable across all gestational stages, with a slight reduction in Ruminococcus-type in the last stage of pregnancy. Phylum level across pregnancy: Bacillota (70.6%) and Bacteroidota (17.8%)—relatively stable “core microbiota”. Microbial alterations associated with gestational age. Increased with gestational age: [Ruminococcus], Collinsella, Megamonas, and unclassified-Erysipelotrichaceae. Decreased with gestational age: Ruminococcus, Dialister, and unclassified-Lachnospiraceae. The most common taxa in the mid-trimester: Streptococcus, Megasphaera, unclassified-Clostridiales, and Bacteroides. Enriched at 21–28 weeks: Streptococcus and Megasphaera. Enriched at 17–24 weeks: unclassified-Clostridiales. Reduced at 21–28 weeks: Bacteroides. Diversity: no significant differences in alpha and beta diversity during the pregnancy period. |
| 3 | Hiltunen H. (2021) [103] PMID: 34349229 | Turku University Hospital, Turku, Finland Study period: not clear. | Cases: 55 preterm neonates (<35 gestational weeks) and their 51 mothers. Controls: 25 spontaneously born full-term neonates. Fecal samples were collected during the first 3–4 postpartum days. | 16S ribosomal RNA gene (V3–V4 region) sequencing Bioinformatics analysis: QIIME2, the taxonomy was formed by using the Greengenes v.13.8 database | The maternal-gut-microbiota profiles (within 3–4 postpartum days): Phylum level: Bacillota (62.4%) was the dominant taxa, followed by Bacteroidota (29.6%) and Actinomycetota (4.3%). Family level: Bacteroidaceae (22.5%), Lachnospiraceae (22.4%), and Oscillospiraceae (21.6%). Relationship of microbiota with clinical characteristics: - The mothers who had received intrapartum antibiotics had a higher abundance of Bacillota, Fusobacteriota, Pseudomonadota, and Actinomycetota. - The mothers not receiving intrapartum antibiotics had a higher abundance of Porphyromonadaceae. - Mothers with vaginal delivery presented a higher abundance of Roseburia and mothers without antibiotic treatment a higher abundance of Macellibacteroides. Diversity: No differences in alpha or beta diversity in relation to gestational age or mode of delivery. Significant differences with regard to intrapartum antibiotic use were seen in alpha diversity Faith PD and evenness, Bray–Curtis, and unweighted UniFrac beta diversity. Contribution of the maternal to neonatal microbiota was higher in neonates born spontaneously as compared to those born after iatrogenic preterm delivery. Gestational age or mode of delivery did not affect the extent to which the maternal gut microbiota contributed to the preterm gut microbiota. |
| 4 | Li D. (2021) [113] doi:10.1016/j.medmic.2021.100046 | Zhujiang Hospital, Southern Medical University, China Study period: July 2020–January 2021 | Cases: - Preterm group: 15 preterm neonates (after 28th but before 37th gestational week) and their mothers. - Term group: 11 term neonates (37–42 gestational weeks) and their mothers. Controls: blank, distilled water, and air sample swabs. Samples collected: neonates—oral and rectal (within 24 h of birth); mothers—vaginal and rectal (at admittance for delivery). | 16S ribosomal RNA gene (V3–V4 region) sequencing Bioinformatics analysis: QIIME version 1.9.1 | The maternal-gut-microbiota profiles (at admittance for delivery): Phylum level: no significant differences between mothers of preterm and term neonates—Bacillota (38.6% vs. 45.2%, preterm and term, respectively), Bacteroidota (44.8% vs. 36.2%, respectively), Pseudomonadota (10.5% vs. 12.7%), Actinomycetota (1.4% vs. 3.4%, respectively). Genus level: the abundance of Rothia and Gemella was considerably decreased in mothers delivered prematurely. Diversity: No difference in the maternal gut microbiome. The preterm group had higher alpha diversity in the maternal vaginal microbiota than the term group, and there were more species in the maternal vaginal microbiota of the preterm group, as opposed to the maternal gut microbiota. The preterm group had higher beta diversity in vaginal (p = 0.003) microbiomes compared to the term group. Contribution of the maternal to neonatal microbiota: Citrobacter freundii, Escherichia coli, Ralstonia pickettii, Prevotella corporis, Lactobacillus iners, Prevotella disiens, Prevotella timonensis, Enterococcus faecium, Faecalibacterium prausnitzii, Corynebacterium amycolatum, and Ureaplasma parvum were detected in neonatal and maternal gut microbiota. Similar species of neonatal oral and maternal vaginal microbiota: Lactobacillus iners, Lactobacillus johnsonii, Ralstonia pickettii, Neisseria subflava, Ureaplasma parvum, Rothia mucilaginosa, Prevotella timonensis, Escherichia coli, Streptococcus salivarius subsp. thermophilus, Prevotella bivia, Prevotella colorans, and Enterococcus faecium. |
| № | Authors, PMID | Setting and Time Period | Participants and Study Design | Methods | Features of Microbiome |
|---|---|---|---|---|---|
| 1 | Liu J. (2017) [107] PMID: 27988814 | Department of Obstetrics of the First Affiliated Hospital of Jinan University, Guangzhou, China. Study period: 2014. | Cases: 26 women newly diagnosed with PE in the third trimester. Control groups: - I: 24 women in the first trimester (11–14 weeks). - II: 24 women in the second trimester (24–28 weeks). - III: 26 women in the third trimester. | 16S ribosomal DNA gene (V4 region) sequencing. Bioinformatics analysis: FLASH software was used to assemble the reads into tags, and the USEARCH package was used to cluster tags into OTUs. | The maternal-gut-microbiota profiles: Phylum level: women with PE had an increased level of Cyanobacteriota (1.07%), which was the fifth most abundant phylum. Healthy women in the third trimester (compared to women in the first and second trimesters) had significantly more Mycoplasmatota (0.30%), and Verrucomicrobiota almost disappeared. Species level: Clostridium perfringens and Bulleidia moorei had a significantly higher and Coprococcus catus had a significantly lower relative abundance in women with PE compared to healthy women in the third trimester. Diversity: There was no statistical significance in alpha diversity among the four groups. |
| 2 | Lv L.-J. (2019) [108] PMID: 31297341 | Guangdong Women and Children Hospital, Guangzhou, China. Study period: January 2017–December 2017. | Cases: 78 women newly diagnosed with PE with severe effects in their third trimesters. Controls: 72 healthy pregnant women. Fecal-sample collection: in the third trimester and at 1 and 6 weeks postpartum. | 16S ribosomal DNA gene (V4 region) sequencing. Bioinformatics analysis: OTU taxonomically classified using Greengenes database v13.8 by QIIME2. | The maternal-gut-microbiota profiles: Phylum level: Fusobacteriota, Mycoplasmatota, and Verrucomicrobiota were decreased in PE women at the antenatal time point. Genus (and species) level: eight genera were enriched in antenatal PE samples—Blautia (Blautia spp., 84.9%) and Ruminococcus2 (R. gnavus, 54.7%), followed by Bilophila (B. wadsworthia, 100%) and Fusobacterium (F. nucleatum, 100%), represented the major variances in PE microbiomes. Genera depleted in antenatal PE samples: Faecalibacterium, Gemmiger, Akkermansia, Dialister, and Methanobrevibacter (mostly consisted of F. prausnitzii, G. formicilis, A. muciniphila, an unclassified Dialister spp., and M. smithii, respectively). Diversity: no significant differences in alpha and beta diversity were detected during the antepartum and postpartum periods. Relationship of microbiota with clinical characteristics: the systolic and diastolic blood pressure levels were positively correlated with PE-enriched genera (Anaerococcus, Ruminococcus2, Fusobacterium, and Oribacterium). The fetal features (e.g., birth weight) were positively correlated with PE-depleted genera. IL-6 was positively associated with Blautia and Bilophila, and negatively associated with Faecalibacterium. |
| 3 | Wang J. (2019) [119] PMID: 31850241 | Peking University Third Hospital, Beijing, China. Study period: January 2018–December 2018. | Cases: 48 women with PE. Controls: 48 healthy pregnant women. Fecal samples were collected in the third trimester. Analysis of fecal and plasma lipopolysaccharide (LPS) and plasma trimethylamine-N-oxide (TMAO) concentration levels. | 16S ribosomal DNA gene (V4 region) sequencing. Bioinformatics analysis: Sequence analysis was performed using Uparse software. The Silva Database based on the Mothur algorithm was used to annotate the taxonomic information. Alpha and beta diversity metrics were calculated using the QIIME v1.7.0. | The maternal-gut-microbiota profiles (third trimester): Phylum level: the relative abundance of Bacillota was decreased in the PE group (51.6% vs. 59.6%, respectively, p < 0.05). The abundance increased in PE compared with controls: Bacteroidota (40.5% vs. 34.8%, p < 0.05), Pseudomonadota (4.5% vs. 2.5%, p < 0.05), and Actinomycetota (2.9% vs. 1.8%, p < 0.05). The abundances were lower in PE: Bacillota, Clostridia, Eubacteriales, Oscillospiraceae, Rikenellaceae, Faecalibacterium, Alistipes, and Bacteroides_stercoris. The abundances were higher in PE: Bacteroidota, Pseudomonadota, Actinomycetota, Bacteroidia, Gammaproteobacteria, Enterobacterales, Enterobacteriaceae, Bacteroides_coprocola, and Bacteroides_fragilis. Diversity: Alpha diversity was lower in the PE group (but NS). Beta diversity (by UniFrac distance) was different (ANOSIM analysis, p = 0.011). LPS and TMAO levels. The fecal and plasma LPS concentrations and plasma TMAO concentrations were higher in PE women. |
| 4 | Chen X. (2020) [118] PMID: 31900289 | Department of Obstetrics of the Nanfang Hospital, Southern Medical University, Guangzhou, China. Study period: March 2017–March 2018. | Cases: 67 women with PE (40 of them with severe PE)—21 with an early onset of PE and 46 with a late onset of PE. Controls: 85 normotensive pregnant women. Fecal samples were collected in the third trimester. Experimental part of the study: fecal-microbiota transplantation in an antibiotic-treated mouse model. | 16S ribosomal RNA gene sequencing. Taxonomic groups were based on the Greengenes Database v.13.8 using QIIME v.1.9.1. | The maternal-gut-microbiota profiles (third trimester): Genus level: Clostridium, Dialister, Veillonella, and Fusobacterium were significantly increased, whereas Lachnospira, Akkermansia, and Faecalibacterium were depleted in PE, but there were no differences between severe and not-severe PE. Diversity: Alpha diversity was markedly decreased in the PE group. The microbiome of the PE group differed significantly from the normal pregnancy group (using the unweighted UniFrac distance). There were no significant differences between the PE with and without severe features or between the early and late onset of PE subgroups for both alpha diversity and beta diversity. Relationship of microbiota with clinical characteristics: Veillonella and Fusobacterium were correlated for the systolic and diastolic blood pressure, proteinuria, oedema levels, alanine aminotransferase, aspartate aminotransferase, serum creatinine, and albumin. The correlations for Lachnospira, Akkermansia, and Faecalibacterium were opposite. No bacteria were correlated with clinical parameters when testing only on the PE group. PE patients’ fecal microbiota transplantation of induced PE phenotype in mice: At 6 weeks after transplantation, mice had higher pregestational systolic blood pressure, which was further elevated. PE-transplanted mice had increased proteinuria, embryonic resorption, and lower fetal and placental weights. Their T regulatory/helper-17 balance in the small intestine and spleen was disturbed with more severe intestinal leakage. |
| 5 | Wang J. (2020) [114] PMID: 32265423 | Nanjing Maternity and Child Health Care Hospital, Nanjing, China. Study period: January 2018–December 2018. | Cases: 25 women who subsequently developed PE. Controls: 25 healthy pregnant women. Fecal samples were collected in the second trimester (20–24 weeks) and third trimester (32–34 weeks). | 16S ribosomal RNA gene (V4 region) sequencing. Bioinformatics analysis: sequence analysis was performed using Uparse software. The Silva Database based on the Mothur algorithm was used to annotate the taxonomic information. Alpha and beta diversity metrics were calculated using QIIME v1.9.1. | The maternal-gut-microbiota profiles: Phylum level in dynamics (second to third trimester): PE patients had a lower relative abundance of Bacteroidota in second trimester than that of the third trimester. The relative abundances of Pseudomonadota and Mycoplasmatota significantly decreased in controls from the second to third trimester. Family level: Enterobacteriaceae decreased in controls. Phylum level in each trimester: second trimester—the relative abundances of Bacillota, Bacteroidota, Actinomycetota, Pseudomonadota, and Mycoplasmatota showed no significant differences between the PE and the control group. Third trimester—the abundance of Bacillota was significantly lower in the PE group than in controls (mean 60.6% vs. 75.5%, respectively, p < 0.05). Bacteroidota and Pseudomonadota were higher in the PE group than in controls (median 31.09% vs. 18.24%, respectively, p < 0.05; 1.52% vs. 0.64%, respectively, p < 0.05). There were no significant differences in the abundances of Actinomycetota, and Mycoplasmatota between the two groups. LefSe: the relative abundances of the phylum Bacteroidota, class Bacteroidia, and order Bacteroidales were increased in the PE group, Bacillota, class Clostridia, order Eubacteriales, and genus unidentified Lachnospiraceae were decreased in the PE group in the third trimester. Family level at the third trimester: the abundance of Enterobacteriaceae was significantly higher in the PE group than in the control group (median, 0.75% vs. 0.01%, respectively, p < 0.05). Diversity: the Shannon and Simpson indices in the PE group were slightly lower than those in the control group in the second and third trimesters, but NS. |
| 6 | Miao T. (2021) [117] PMID: 34262418 | Department of Obstetrics and Gynecology in Changzhou Maternity and Child Health Care Hospital (which is affiliated to Nanjing Medical University), Guangzhou, China. Study period: October 2017–April 2018. | Groups (periods of fecal-sample collection): Cases: 12 women with PE (35.2 ± 2.0 weeks). Controls: eight women without PE (34.8 ± 4.6 weeks). Fecal samples were collected in the third trimester. | 16S ribosomal RNA gene (V4 region) sequencing. Bioinformatics analysis: OTU taxonomically classified using Greengenes database v201305 by QIIME v1.8.0. | The maternal-gut-microbiota profiles (third trimester): Phylum level: women with PE had decreased abundance of Actinomycetota compared to the control group (p = 0.042). The control group had an increased abundance of Mycoplasmatota. Family level: The relative abundance of Bifidobacteriaceae was lower in the PE group (3.75%) than in the control group (12.76%) (p = 0.039). The relative abundance of Oscillospiraceae, Prevotellaceae, and Coriobacteriaceae in the PE group was decreased compared to the control group (NS). Genus level: Blautia was increased significantly in the PE group in comparison with that in the control group (19.13% vs. 9.71%, respectively, p = 0.026). Ruminococcus was increased in the PE group compared to the control group (10.32% vs. 6.11%, respectively, p = 0.048). There was a reduction in Bifidobacterium in the PE group compared to the control group (p = 0.038). Downward trend in Faecalibacterium, Roseburia, and Prevotella in the PE group compared with that in the control group (NS). Diversity: There was no statistical significance in the alpha diversity among groups or the differences in beta diversity between groups. Relationship of microbiota with clinical characteristics: The relative abundance of Blautia was positively correlated with maternal age, pregestational weight, hematocrit, levels of C-reactive protein, triglyceride (p < 0.05 for all), and low-density lipoprotein cholesterol (p < 0.01). The relative abundance of Ruminococcus was positively correlated with the pregestational weight, pregestational BMI, antepartum weight, antepartum BMI, lipopolysaccharide-binding protein, and triglyceride (p < 0.05 for all). The relative abundance of Bifidobacterium was significantly negatively correlated with the systolic and diastolic blood pressure, levels of cholesterol and aspartate aminotransferase (p < 0.05 for all), and triglyceride level (p < 0.01). |
| 7 | Huang L. (2021) [121] PMID: 34607559 | Changsha Hospital for Maternal and Child Health Care, Hunan, China. Study period: not clear. | Cases: - 26 pregnant women with PE. - 25 pregnant women with abnormal placental growth. Controls: - 28 healthy pregnant women. - 21 healthy women. Fecal samples were collected in the third trimester. | 16S ribosomal RNA gene (V4 region) sequencing. Bioinformatics analysis: qualified paired-end reads were matched, dereplicated, clustered, and chimera-filtered using VSEARCH (v2.4.4) against the SILVA138 database and then OTUs were assembled using QIIME2. | The maternal-gut-microbiota profiles (the third trimester): Phylum level: the percentage of TM7 (candidatus Saccharibacteria) was significantly increased in the abnormal-placental-growth group. Genus level: The relative abundances of Prevotella, g_WAL_1855D, g_1_68, Porphyromonas, Varibaculum, and Lactobacillaceae were significantly decreased in the PE group compared with the normal-pregnancy group. Prevotella, g_1_68, Porphyromonas, Lactobacillaceae, Mobiluncus, Campylobacter, and Peptostreptococcus were decreased in the abnormal-placental-growth group compared with the normal-pregnancy group. The ratio of number of subject Lactobacillaceae/all and the relative abundance of Lactobacillaceae were significantly higher in the normal-pregnancy group. Diversity: Alpha diversity exhibited no statistical differences. There was a significant difference in beta diversity (Bray–Curtis distance and Adonis) in the bacterial composition only in the abnormal-placental-growth group compared with the nonpregnant group (p = 0.043). The gut microbiota compositions of the abnormal-placental-growth group, the PE group, or both groups of abnormal pregnancies were significantly shifted compared with that of the normal pregnancy group (Adonis, p = 0.002, p = 0.015, p = 0.001, respectively). The PE group was significantly different from the normal-pregnancy group. The abnormal-placental-growth group had the highest number of unique OTUs. Relationship of microbiota with clinical characteristics: Lactobacillaceae (OTU 255) was significantly negatively related to diastolic blood pressure in the PE and normal-pregnancy groups. |
| 8 | Jin J. (2022) [123] PMID: 35950704 | Liaocheng, China Study period: 2017–2022. Samples collected: | Cases: 92 women with PE (37.69 ± 2.4 weeks). Controls: 86 healthy pregnant women (37.05 ± 3.2 weeks). Fecal samples were collected in the third trimester. Analysis of SCFA in feces, serum, and placentas. Experimental part of the study: fecal-microbiota transplantation in mice. | 16S ribosomal RNA gene (V3–V4 regions) sequencing. Bioinformatics analysis: The dereplicated sequence reads were denoised into ASVs. The phylogenetic affiliation was analyzed using the USEARCH 10 SINTAX algorithm against the RDP training-set database. | The maternal-gut-microbiota profiles: Genus and species level: the abundances of many intestinal SCFA-producing bacteria were significantly reduced in PE (Alistripes, Fusicatenibacter, Coprobacter, Oscillibacter, Clostridium_XIVb, and Clostridium_XV). PE and controls could be distinguished only based on the abundances of Akkermansia and Oscillibacter with 89.7% accuracy. Diversity: the alpha and beta diversity and Bacillota/Bacteroidota ratio of the gut microbiota in PEs were significantly changed, suggesting gut dysbiosis. Relationship of microbiota with clinical characteristics: Akkermansia was reduced in PE and negatively correlated with blood pressure and urine protein. A. muciniphila (SCFA-producing bacteria) was significantly reduced in the feces of PE women, and was significantly positively correlated with the levels of propionate and butyrate in the placenta. Akkermansia abundance was positively correlated with the levels of fecal 2-Arachidonoylglycerol and serum IL-10 but negatively correlated with the serum levels of lipopolysaccharide and IL-17. Fecal, placental, and serum levels of propionic and butyric acids were significantly reduced in PE and positively correlated with each other. Placental levels of propionic and butyric acids negatively correlated with blood pressure and urine protein levels. The abundances of bacteria were very low in the placentas, did not differ and are likely to have originated from contamination during the experimental procedure. The peripheral blood Treg/Th17 ratio was significantly decreased in PE. The abnormalities of peripheral Treg and Th17 cells in PEs were closely related to the intestine. The intestinal-barrier damage in PE was revealed. The serum level of lipopolysaccharide was considerably increased in PE. The fecal level of 2-Arachidonoylglycerol, a protector of the intestinal barrier, was decreased. PE patients’ fecal microbiota transplantation of induced pre-eclamptic phenotype in rats. Akkermansia muciniphila, propionate, or butyrate significantly alleviated the symptoms of pre-eclamptic rats. Propionate promoted trophoblast invasion, thereby improving spiral arterial remodeling. |
| 9 | Li P. (2022) [124] PMID: 36380372 | A two-sample Mendelian randomization study. | Analysis of gut microbiota from GWAS meta-analysis conducted by the MiBioGen consortium (n = 18,340 individuals) and the summary statistics of PE from the FinnGen consortium R7 release data (5731 cases and 160,670 controls). | 16S ribosomal RNA gene sequencing targeting variable regions V4, V3–V4, and V1–V2 (MiBioGen consortium). | Maternal microbial profiles: Genus level:Collinsella (OR 0.77), Enterorhabdus (OR 0.76), Eubacterium (ventriosum group) (OR 0.76), Lachnospiraceae (NK4A136 group) (OR 0.77), and Tyzzerella 3 (OR 0.85) were found to be associated with PE. Bifidobacterium had a protective effect against PE. |
| 10 | Lv L.-J. (2022) [125] PMID: 36189343 | Guangdong Women and Children Hospital, Guangzhou, China. Study period: not clear. | Cases: 40 women with severe PE (246 ± 25 gestational days). Controls: 37 healthy pregnant women (273 ± 11 gestational days). Fecal samples were collected in the third trimester. | Shotgun metagenomic sequencing. Taxonomic classification of the bins was realized based on the GTDB-Tk toolkit (v1.4.0) by assigning the sequences of each bin to the Genome Taxonomy Database (v. r95). The taxonomic name of the bins was manually modified to accord with traditional nomenclatures following the National Center for Biotechnology Information (NCBI) taxonomy. | The maternal-gut-microbiota profiles (the third trimester): Family level: Lachnospiraceae and Coriobacteriaceae were significantly enriched in PE patients compared with healthy controls; Bacteroidaceae were markedly depleted in the PE patients. Genus and species levels: - PE-enriched species: Blautia (unknown at the species level), five members of Pauljensenia (P. bouchesdurhonensis and four uncultivated species), five members of Ruminococcus (containing R. gnavus and four uncultivated species), and Fusobacterium ulcerans. - Control-enriched species: 14 members of Bacteroidaceae, containing 7 Bacteroides spp., 4 Phocaeicola spp., Prevotella bivia, Paraprevotella clara, Barnesiella intestinihominis, and also Akkermansia muciniphila and Bilophila wadsworthia. PE-enriched species (Olsenella sp. M220, Ruminococcus sp. M094, Blautia sp. M090, and Senegalimassilia anaerobia M253), as well as several control-enriched species (Flavonifractor plautii M314, Bacteroides uniformis M403, and Bacteroides sp. M398), featured the highest score for the discrimination of PE patients and healthy controls (AUC = 0.805). Diversity: Alpha diversity exhibited no statistical differences. Beta diversity (Bray–Curtis distance) revealed an alteration of the gut microbial structure between PE patients and the control group. The PE status explained 1.4% of the microbial variations (permutated p = 0.005). |
| 11 | Lin H. (2022) [126] PMID: 36364844 | Hunan Provincial Maternal and Child Health Hospital, Changsha, China. Study period: March 2017–March 2018. | Cases: 35 women with pregnancy-induced hypertension (PIH) (15 of them with PE) (13.15 ± 3.26 weeks, initially). Controls: 35 healthy pregnant women (12.70 ± 0.86 weeks, initially). Fecal samples were collected in the first, second, and third trimesters, at delivery and during postpartum period. | Metagenomic sequencing. Taxonomic annotation and functional annotation realized by MetaPhlAn (v. 2.1.0) and HUMAnN (v. 2-0.11.0). | The maternal-gut-microbiota profiles: Phylum level: PIH patients had a higher abundance of Bacillota and a lower abundance of Bacteroidota and Verrucomicrobiota. Species level: - Higher abundance in PIH: Eubacterium rectale and Ruminococcus bromii. - Lower abundance in PIH: Alistipes putredinis, Bacteroides vulgatus, Ruminococcus torques, Oscillibacter unclassified, Akkermansia muciniphila, Clostridium citroniae, Parasutterella excrementihominis, and Burkholderiales bacterium_1_1_47. Diversity: Alpha diversity exhibited no statistical differences. Beta diversity between the PIH and control groups differed statistically. Relationship of microbiota with clinical characteristics: The abundance of A. putredinis was negatively correlated with early uric acid and early systolic and diastolic blood pressure, and was positively correlated with low-density lipoprotein cholesterol. The abundance of B. vulgatus was negatively correlated with early BMI, waist, early ALT, early uric acid, early weight, early AST, insulin, γ-glutamyltransferase (GGT), and early systolic and diastolic blood pressure. The abundance of O. unclassified and A. muciniphila was negatively correlated with early systolic and diastolic blood pressure, and A. muciniphila was positively correlated with GGT. The abundance of C. citroniae was negatively correlated with early diastolic blood pressure. |
| № | Authors, PMID | Setting and Time Period | Participants and Study Design | Methods | Features of Microbiome |
|---|---|---|---|---|---|
| 1 | Zhou Y. (2015) [161] PMID: 25741698 | Brigham and Women’s Hospital, Boston, MA, USA. Study period: not clear. NEC stage: II–III. | Cases: 12 neonates with NEC (GA of 27.8 (24–31) weeks). Age of NEC: early (≤DOL 22) vs. late (>DOL 22). Controls: 26 neonates without NEC (GA of 27.9 (24–31) weeks). Fecal samples were collected with a median sampling interval of 3 days. | 16S ribosomal RNA gene (V3–V5 regions) sequencing. Taxonomic groups were based on the Ribosomal Database Project (RDP) naive Bayesian classifier (version 2.5, training set 9) | Gut microbiota profiles: Early-onset NEC (≤DOL 22):Clostridium sensu stricto (Clostridia) were significantly higher in proximity to NEC onset. Late-onset NEC (>DOL 22): Escherichia/Shigella (Gammaproteobacteria) was significantly higher in cases than controls six days before NEC onset. Cronobacter (Gammaproteobacteria) was significantly higher 1–3 days prior to NEC onset. Diversity: The richness and Shannon diversity from the samples before NEC onset increased significantly over the 2 months of life. Neonates without NEC had a higher richness (but not Shannon diversity). NEC samples tended to have lower microbial diversity than controls, and they also had marginally more antibiotic usage than the controls. |
| 2 | Heida F.H. (2016) [134] PMID: 26787171 | University Medical Center of Groningen, Groningen, The Netherlands. Study period: October 2013–February 2014. NEC stage: II–III. | Cases: 11 neonates with NEC (GA of 27 (24–29) weeks). Age of NEC: 12.5 (4–43). Controls: 22 neonates without NEC (GA of 26 (24–29) weeks). Fecal-sample collection: at 1 (0–4) day of life (meconium); then, the two samples were collected in the week prior to the onset of NEC (when available). | 16S ribosomal RNA gene (V3–V4 regions) sequencing. Taxonomic identification down to the family and genus levels with QIIME. Sequence identification to the species level with ARB (SILVA rRNA database). | Gut microbiota profiles: NEC group: There was a higher abundance of Clostridium perfringens and Bacteroides dorei, and a lower abundance of Clostridioides difficile in the meconium. Prior to disease onset, NEC cases remained dominated by Enterobacteriaceae or developed an aberrant microbiota composition that formed a distinct cluster, consisting of C. perfringens and B. dorei. Controls: There was a shift from a more Enterobacteriaeceae-dominated microbiota into one with more staphylococci (19%) and other lactate-producing bacilli. Diversity: This was not associated with NEC development. Maternal factors: The amount of breast milk as a percentage of the total amount of feeding was correlated with an increase in lactate-producing bacilli and a decrease in Gram-negative species, which include the Enterobacteriaceae and Bacteroidaceae. |
| 3 | Warner B.B. (2016) [152] PMID: 26969089 | - St. Louis Children’s Hospital; - Children’s Hospital at Oklahoma University Medical Center; - Kosair Children’s Hospital, USA. Study period: July 2009–September 2013. NEC stage: II–III. | Cases:28 neonates with NEC (GA of 26.0 (24.7–27.9) weeks) Age of NEC: 24 (19–48). Controls: 94 Neonates without NEC (GA of 27.0 (25.9–28.7) weeks) Fecal-sample collection: all stools of neonates were collected up to and including the day before NEC was diagnosed or 60 days of age. | 16S ribosomal RNA gene (V3–V5 regions) sequencing Read classification using the Ribosomal Database Project naive Bayesian classifier version 2.5, training set 9. | Gut microbiota profiles: NEC group: A significant increase in Gammaproteobacteria over time, and a decrease in Negativicutes and the combined Clostridia–Negativicutes class (in infants with a GA < 27 weeks). Controls: Associated with an abundance of Negativicutes, and the combined Clostridia–Negativicutes class (in infants with a GA < 27 weeks). The Clostridia–Negativicutes class increased over time (in infants with a GA > 27 weeks). Diversity: The alpha diversity significantly increased in the stools of controls, but not in NEC patients (the diversity decreased over time). Associations with clinical features: A greater GA at birth was associated with higher proportions of Negativicutes and the combined Clostridia–Negativicutes class, and lower proportions of Bacilli. Vaginal birth was associated with lower proportions of Bacilli. Greater antibiotic exposure was associated with higher proportions of Bacilli, and lower proportions of Clostridia, and the combined Clostridia–Negativicutes. |
| 4 | Ward B.V. (2016) [154] PMID: 26997279 | Two level III neonatal intensive care units (NICUs), Cincinnati, OH, USA. One level III NICU in Birmingham, AL, USA. Study period: December 2009–July 2012. NEC stage: II–III. | Cases: 27 neonates with NEC (GA of 26 (23–28) weeks). Age of NEC: 26 (10–39). Preterm controls: 117 neonates without NEC (GA of 26 (23–29) weeks). Term controls: 22 neonates without NEC (GA 39 (38–41) weeks. Fecal-sample collection in three collection periods (days 3–9, 10–16, and 17–22). | Shotgun metagenomic sequencing. Relative taxonomic abundances were determined with MetaPhlAn version 2.0. Pangenome-based strain-level profiling of E. coli from metagenomes, and multilocus sequence type (MLST) analysis. | Preterm gut microbiome analysis (overall): Days 3–9: Median relative abundance (MRL): E. coli (0.92), Serratia marcescens (0.24), Klebsiella spp. (0.14), Streptococcus sp. GMD4S (0.12), and E. faecalis (0.11). Days 10–16: E. coli (0.93), S. marcescens (0.39), Veillonella parvula (0.33), Klebsiella spp. (0.19), K. oxytoca (0.11), and Streptococcus sp. GMD4S (0.12). Days 17–22: E. coli (0.81), Klebsiella spp. (0.48), V. atypica (0.21), E. cloacae (0.14), Citrobacter freundii (0.13), and Streptococcus sp. GMD4S (0.12). Infants with high antibiotic treatment were specifically enriched in E. coli relative to low-treatment infants who were enriched in the order Eubacteriales, genus Veillonella, and Klebsiella spp. Infants who developed NEC had less Veillonella and were specifically enriched in E. coli. NEC group: Colonization by uropathogenic E. coli (UPEC) was a highly significant risk factor for the development of NEC. UPEC was even more strongly correlated with mortality as an outcome. NEC cases with Klebsiella dominance were also observed. Diversity was lower among infants receiving a high level of antibiotic administration. Alpha diversity was higher at days 17–22 in preterm controls but decreased in infants who developed NEC. |
| 5 | Dobbler P.T. (2017) [148] PMID: 29187842 | Neonatology Section of Hospital de Clínicas de Porto Alegre, Brazil. Study period: not clear. NEC stage: not clear. | Preterm neonates with a GA ≤ 32 weeks. Cases: 11 neonates with NEC Age of NEC: 8.0 (5.0–13.0). Controls: 29 neonates without NEC. Fecal-sample collection: with the first stool (meconium) weekly up to the 5th week of life. | 16S ribosomal RNA gene (V4 region) sequencing. Taxonomic classification was carried out in QIIME based on the UCLUST method against the Greengenes 13.5 database. | NEC group: There was an indeterminate pattern of microbial succession (called here as ‘chaotic’ or ‘abnormal’ pattern). There was a higher abundance of Pseudomonadota and lower abundance of Bacillota during week 3. There was an overall higher average abundance of Actinomycetota than in controls during weeks 1 and 2. Days 0–4: dominance of Pseudomonadota (44.66%) and lower abundance of Bacteroidota (35.23%), Bacillota (15%), and Actinomycetota (1.7%). Days 5–7: an abrupt decrease in Pseudomonadota in the NEC group. Weeks 2–3: a bloom of Pseudomonadota; from then, a steady decline through weeks 4 and 5 that coincided with an increase in Bacillota. The early detection of a high dominance of Enterobacteriaceae, especially Citrobacter koseri and Klebsiella pneumoniae, a lack of Lactobacillaceae, low diversity, and altered microbial–microbial associations during the first days of life could be indicators of NEC risk in preterm infants. Controls: a steady increase in Bacillota overlapping with a decline in Pseudomonadota, Bacteroidota, and Actinomycetota until week 4 followed by a sudden re-emergence of Pseudomonadota. Days 0–4: a high abundance of Pseudomonadota (40.07%) and Bacteroidota (36.35%), and a low abundance of Bacillota (13.14%) and Actinomycetota (2.47%). Days 5–7: a dominance of Bacillota (52.64%) and Pseudomonadota (31.43%) with a low abundance of Bacteroidota (13.47%) and Actinomycetota (0.54%). Weeks 2–3: a steady increase in Bacillota overlapping with a decline in Pseudomonadota, Bacteroidota, and Actinomycetota until week 4 followed by a sudden re-emergence of Pseudomonadota. A community of obligate anaerobes was highly influential in the intestine of controls during the first four days of life and appeared to control the proliferation of Enterobacteriaceae. Diversity: patients with NEC tended to have low alpha diversity and high dominance compared to the controls (significant during the 3rd week). Microbial dominance at the 3rd week was higher in the NEC cases than in the controls. |
| 6 | Rozé J.-C. (2017) [157] PMID: 28659297 | EPIFLORE Study, France Study period: 2011. NEC stage: II–III. | Cases: 14 neonates with NEC (GA of 28.4 ± 1.9 weeks). Age of samples collected at onset: 25.8 ± 16.7. Controls: 73 neonates without NEC (GA of 28.5 ± 1.7 weeks). Fecal-sample collection at DOL 7, 28, and at NEC onset (3 (0–7) days after onset)/at discharge in controls. | 16S ribosomal RNA gene (V3–V4 regions) sequencing. Culture methods. | NEC group (culture methods): The Clostridium genus was significantly associated with NEC: (86% vs. 35%, OR: 11.3). At the species level, C. neonatale (50% vs. 11%, OR 5.5) and Staphylococcus aureus (57% vs. 13%, OR 7.1) were significantly associated with NEC. NEC group (16S rRNA sequencing): Colonization with bacteria from the Clostridium sensu stricto genus tended to be associated with NEC (20.6% vs. 11.7%, p = 0.08) and with a higher proportion of C. neonatale, together with C. butyricum. Gammaproteobacteria were also differentially represented with a trend toward lower proportions of Klebsiella and Citrobacter and an association with some specific bacterial operational taxonomic units related to either clostridia or Gammaproteobacteria in NEC infants. |
| 7 | Wandro S. (2018) [136] PMID: 29875143 | Children’s Hospital, Orange County, Orange, CA, USA. Study period: 2011–2014. NEC stage: not clear. | 32 preterm neonates with birth weights of 620–1570 g. Cases: three neonates with NEC; eight neonates with late-onset sepsis. Age of NEC: 27, 31, and 41. Controls: 21 healthy neonates. Fecal-sample collection: between days 7 and 75 of life. The sampling times and numbers of fecal samples varied. Gas chromatography–mass spectrometry was used. | 16S ribosomal RNA gene (V3–V4 regions) sequencing Taxonomy was assigned using QIIME and the Greengenes 13_8 database. | There were no clear signatures of microbiome composition linked to NEC or sepsis. Longitudinal samples from individual infants remained highly personalized over several weeks. Preterm infant microbiomes were shaped by shared exposures, especially to antibiotics, leading to the dominance of antibiotic-resistant facultative anaerobes (Enterococcus spp.). The anaerobic, milk-degrading bifidobacteria were largely absent, even in preterm infants with access to breast milk. Diversity: The difference in alpha diversity was associated with antibiotic use. Only vaginally born infants were colonized by Bacteroides (4 out of 9 infants), while none of the 22 infants born via C-section were colonized. Metabolite profiles: the metabolite profiles varied over time (individually) and were not associated with NEC or late-onset sepsis. |
| 8 | Romano-Keeler J. (2018) [156] PMID: 30365522 | Monroe Carell Jr. Children’s Hospital, Vanderbilt, TN, USA. Study period: not clear. NEC stage: surgical NEC. | Cases: 12 surgical patients with NEC (GA of 25–33 weeks). Controls: 14 surgical patients without NEC (GA of 24–39 weeks). Age at surgery: 5–46. Sample collection: intestinal tissue during surgery, patient’s first post-operative stool, or by scraping surgical tissue. | 16S ribosomal RNA gene (V1–V3 regions) sequencing. Sequences were aligned to the SILVA database release 123 and taxonomically classified with the Ribosomal Database Project (RDP) classifier 11. | NEC group: Tissue samples—higher abundances of Staphylococcus and Clostridium sensu stricto I. Fecal samples—a higher abundance of Staphylococcus and lower abundances of Actinomyces and Corynebacterium. Diversity: The fecal microbial richness and diversity tended to be lower in NEC patients (p = 0.078). The tissue microbial richness was lower (p < 0.05); the alpha diversity tended to be lower than in controls (p = 0.081). There was distinct beta diversity in the tissue samples of NEC patients vs. controls. Metabolic pathways in NEC patients were related to signatures of infectious diseases (bacterial toxins) and Staphylococcus aureus infection was enriched in NEC tissue samples compared to non-NEC tissues. |
| 9 | Olm M.R. (2019) [151] PMID: 31844663 | University of Pittsburgh Medical Center Magee-Womens Hospital, Pittsburgh, PA, USA. Study period: 5-year period. NEC stage: not clear. Pre-NEC samples were analyzed. | Cases: 34 neonates with NEC (GA of 28 ± 5.5 weeks). Age of NEC: DOL 9 ± 9.8. Controls: 126 neonates without NEC (GA of 29 ± 2.2 weeks). Fecal-sample collection: longitudinal (average 7.2 samples per patient) until NEC onset within 2 days before NEC diagnosis— “pre-NEC” samples. | Shotgun metagenomic sequencing. To determine the taxonomy of bins, the amino acid sequences of all predicted genes were searched against the UniProt database using the USEARCH ublast command. tRep was used to convert the list of identified taxIDs into taxonomic levels. | The gut microbiomes of all infants were dominated by Pseudomonadota, regardless of NEC development. The premature infants had increased Enterobacteriaceae and notably low abundances of Actinomycetota and Bacteroidota. NEC group: A low abundance of Bacillota and a higher abundance of Enterobacteriaceae. However, in pre-NEC samples, the gut microbiomes were not significantly enriched in Enterobacteriaceae (the association of Enterobacteriaceae and NEC infants may be after the administration of antibiotics to treat NEC). The K. pneumoniae strain 242_2 was the most associated with NEC. iRep (bacterial replication rates) values of bacteria were significantly higher in pre-NEC versus control samples. Bacterial replication was stable 4 or more days before NEC, increased daily in the 3 days before diagnosis, and crashed following diagnosis (probably due to subsequent antibiotic administration). The genomes of Enterobacteriaceae displayed higher pre-NEC iRep values than bacteria overall. Diversity: not clear Metabolic pathways: Four factors were significantly associated with pre-NEC samples (taken within 2 days before NEC diagnosis): iRep values overall, genomes encoding specific types of secondary metabolite gene clusters (sactipeptides, bacteriocins, and butyrolactones), Klebsiella, and fimbriae cluster 49. |
| 10 | Lindberg T. (2020) [150] PMID: 29909714 | IV neonatal intensive care unit (NICU), Hartford, CT, USA. Study period: September 2013–September 2015. NEC stage: II–III. | Preterm neonates with a GA of 25.2 (23–27) weeks. Cases: five neonates with NEC. Age of NEC: not clear. Controls: five neonates without NEC. Fecal-sample collection was on a weekly basis beginning with the first bowel movement until patient discharge. | 16S ribosomal RNA gene (V4 region) sequencing. Indicator value analysis using the Indicspecies Package was performed to identify microbial species. | NEC group: A dominance of Pseudomonadota (65.5%), followed by Bacillota (28.1%), Actinomycetota (5.8%), and other bacteria (0.5%). Enterobacteriaceae and Trabulsiella were more abundant in NEC patients. Controls: There was a dominance of Bacillota (55.9%), followed by Pseudomonadota (40.8%), Actinomycetota (2.9%), and other bacteria (0.5%). Veillonella and Enterococcus were more abundant in controls. There was a significant reduction in Pseudomonadota in early samples compared to those collected at later time points. Diversity: Alpha diversity was associated with the day of life (↑ by 0.02 with each day) and antibiotics use (↓ of 0.01 for each additional day). |
| 11 | Brehin C. (2020) [171] PMID: 32709038 | Purpan Hospital in Toulouse, France. Study period: not clear. NEC stage: I (suspected NEC). | Cases: 11 neonates with NEC (GA of 28.4 (26–31) weeks). Age of NEC I: 12 (4–60) Controls: 21 neonates without NEC (GA 30 (26.4–32) weeks). Fecal samples were collected at the time points of 1–10 days; 11–20 d; 21–30 d; and >30 d. Fecal metabolome analysis. | 16S ribosomal RNA gene (V3–V4 regions) sequencing Diseases and host genetic variation linked to NEC-1 associated gut microbiota were identified via MicrobiomeAnalyst. | Gut microbiota profiles in NEC stage I: Days 0–10: had a divergent and more homogenous gut microbiota and lower alpha diversity (Chao1). Days 11–20: had a higher abundance of Streptococcus species and bacteria from the Micrococcales order, lower levels of serine in the fecal metabolome and a higher Chao1 index. Days 21–30: had increased Staphylococcus and Streptococcus species, high intragroup variance, no difference in the Chao1 index, and no difference in the fecal metabolome. The NEC-1 gut microbiota profile was associated with multiple diseases and was found to be significantly increased in ulcerative colitis and host genetic variation and significantly related to ANP32E, a gene involved in ulcerative colitis. Over 30 days: had an increase in Raoultella species. There was no change in the overall microbial diversity indices, but significantly lower levels of ethanol and leucine in the fecal metabolome. |
| 12 | Fu X. (2021) [162] PMID: 34012949 | First Hospital of Jilin University, Changchun, China. Study period: February 2018–April 2019. NEC stage: II–III. | Cases: 15 preterm neonates with NEC (GA 30.2 ± 1.2 weeks). Controls: 15 preterm neonates without NEC (GA 30.1 ± 1.9 weeks). Fecal samples were collected within 48 h after birth, once per week until the NEC diagnosis, 1–2 weeks after treatment, or 28 days after birth. | 16S ribosomal RNA gene (V3–V4 regions) sequencing. Taxonomic groups were based on the Greengenes Database using QIIME2. | Gut microbiota profiles: NEC group: There was a higher abundance of Bacteroidota, and Actinomycetota at birth was much higher than that in controls (which continued until NEC occurred). Higher abundance of Alphaproteobacteria, Betaproteobacteria, Sphingomonas, Lactobacillaceae at NEC onset. In children who subsequently developed NEC, environmental bacteria were detected in the first 48 h of life: Dysgonomonas (lives on surfaces), Hyphomicrobiales, Ralstonia, and Pelomonas (in water sources and hospital ventilators). Controls: had a higher abundance of Gammaproteobacteria, Enterobacteriaceae, and Clostridiaceae. Diversity: at birth—the Chao1 index of the NEC group was higher, but the Shannon index, Good’s coverage, and Pielou’s evenness index were NS; at NEC onset—the Chao1 index, Shannon index, and Pielou’s evenness index of the NEC group were higher. Metabolic pathways: in the NEC group at birth, the pathways involved in the degradation of L-tryptophan and aromatic compounds were upregulated (the species composition was mainly Ralstonia). |
| 13 | Tarracchini C. (2021) [130] PMID: 34704805 | Meta-analysis of metagenomics data from repositories. Data from Croix Rousse University Hospital, Lyon, France were included additionally. Study period: September 2014–November 2014. NEC stage: II–III. | Data from repositories: GA of 23–39 weeks. Cases: 67 neonates with NEC. Age of NEC: not clear. Controls: 57 neonates without NEC. Fecal samples were collected at NEC onset (n = 53) and before NEC (n = 14). Data from Croix Rousse University Hospital: GA of 25–30 weeks. Cases: seven neonates with NEC. Controls: 11 neonates without NEC. Fecal samples were collected weekly during the first 30 days of life. | Shotgun metagenomic sequencing data sets from four different studies retrieved from publicly available repositories. Shotgun metagenomic sequencing. Taxonomic profiling of the retained reads was performed with the METAnnotatorX bioinformatics platform. | Gut microbiota profiles: The microbial profiles of metagenomics samples showed high interindividual variability. NEC group: had a higher abundance of Escherichia coli and Enterococcus faecalis were the main taxa. There was a preterm community state types (PT-CST)-specific increase in opportunistic pathogens, such as E. faecalis, E. coli, Staphylococcus epidermidis, Clostridioides difficile, Ureaplasma parvum, Pseudomonas aeruginosa, Pseudomonas nosocomialis, and members of the Klebsiella genus. There was a reduction in/absence of common early-infant gut commensals (Actinomyces, Schaalia, Veillonella, Bacteroides, and Streptococcus). Clostridium neonatale and Clostridium perfringens species could be potential biomarkers for the predictive early diagnosis of NEC. Controls: had a higher abundance of Streptococcus agalactiae, which was the dominant taxon in premature control subjects. Controls had a much lower relative abundance/absence of K. pneumonia, Proteus mirabilis, Clostridium perfringens, Clostridium neonatale, Pantoea dispersa, and Staphylococcus aureus. Diversity: There was a reduction in the gut microbial biodiversity in the NEC group. Metabolic pathways: in the NEC group, enzymes related to glycosylated protein degradation, i.e., α-fucosidase and sialidase, were almost entirely absent. α-fucosidase and sialidase were positively associated with members of the Blautia, Cutibacterium, and Enterobacter genera and negatively with E. coli. In the NEC group, bacterial tryptophan degradation pathways (tryptophanase enzymes (and indolepyruvate decarboxylase)) were increased. There was gastrointestinal DL-lactate accumulation among NEC patients. |
| 14 | He Y. (2021) [149] PMID: 34922582 | Children’s Hospital of Chongqing Medical University, Chongqing, China. Study period: January 2015–October 2018. NEC stage: II–III. | Cases: 81 neonates with NEC (GA 31.0 (29.4–33.7) weeks); included 19 surgical NEC patients. Age of NEC: 15 (12–19). Controls: 81 neonates without NEC (GA 31.1 (29.3–33.2) weeks); included 19 surgical patients. Fecal samples were collected once the diagnosis of NEC was complete. SCFAs were measured in the fecal samples. Flow cytometry of T cells in ileum lamina propria was performed. The transcription of inflammatory cytokines by qRT-PCR. FMT to germ-free mice before NEC induction. Butyric acid administration. | 16S ribosomal RNA gene (V3–V4 regions) sequencing The phylogenetic affiliation of each 16S rRNA gene sequence was analyzed using the Ribosomal Database Project. | Gut microbiota profiles: NEC group: had a higher abundance of Pseudomonadota, with reduced proportions of Bacillota and Bacteroidota. Diversity: There was decreased α-phylogenetic diversity in NEC patients. SCFA measurement: The levels of butyric acid were significantly lower in the NEC group. Flow cytometry of T cells: the proportion of Treg (Foxp3+) cells compared with T helper cells in the control group was significantly higher than that of the NEC group. Cytokine expression: The surgical NEC cases had significantly higher levels of IL-1β, IL-8, and TNF-α transcripts. NEC cases also had lower expression levels of IL-10 and TGF-β related to the induction and function of Treg cells. Fecal-microbiota transplantation of NEC patients contributed to NEC-like injury in mice. Mice had a decreased Treg/T helper cell ratio. The NEC mice had a similar microbial composition as NEC patients had a higher level of Psudomonadota but lower levels of Bacillota; the concentration of butyrate in the fecal samples was lower. Butyrate administration to mice increased the Treg/T helper cell ratio; mice had lower NEC scores. Butyrate administration did not influence the microbial composition. |
| 15 | Kaelin E. (2022) [163] PMID: 35449461 | St. Louis Children’s Hospital, St. Louis, MO, USA. Study period: not clear. NEC stage: II or higher. | Preterm neonates with a birth weight ≤ 1500 g and GA < 27 weeks. Cases: nine neonates with NEC (GA of 25.5 (24.9–26.0) weeks). Age of NEC: not clear. Controls: 14 neonates without NEC (GA of 25.0 (23.1–25.4) weeks). Fecal samples were collected before NEC onset over the first 11 weeks of life. | Metagenomic sequencing of gut virome DNA. | There was high inter- and intra-individual variation in the infant gut virome. NEC group: A large proportion of the virome could not be assigned family-level taxonomy (unclassified viruses) (84.5%). Classifiable viral contigs included the bacteriophage families Myoviridae, Podoviridae, and Siphoviridae. There was high variability in virus family proportions at each time point and also within individuals over time. A total of 137 contigs were associated with the period 0–10 days before NEC (NEC-associated contigs). Of the NEC-associated contigs with at least five open reading frames (ORFs), 31.7% were predicted to have temperate lifestyles and 68.3% were predicted to be lytic. Controls: A large proportion of the virome was unclassified viruses (85.3%). The identified viral contigs belonged to bacteriophage families including Myoviridae, Podoviridae, and Siphoviridae. The relative abundances of bacteriophage and eukaryotic virus families varied between the control infants in each week of the study. Diversity: There was convergence towards reduced viral beta diversity over the 10 days before NEC onset, driven by specific viral signatures and viral–bacterial interactions. The bacterial beta diversity in NEC patients was stable in windows spanning the 25 days before NEC. The virome diversity in the control group varied between individuals. |
| 16 | Liu X.-C. (2022) [172] PMID: 36060739 | Children’s Hospital of Chongqing Medical University, Chongqing, China. Study period: April 2021–October 2021. NEC stage: not clear. | Cases: 17 neonates with NEC (GA 30.5 ± 2.1 weeks). Controls: 17 neonates without NEC (GA 30.5 ± 1.9 weeks). Age of NEC: 30.2 ± 15.9. Fecal-sample collection: two time points (before NEC (7.0 ± 7.64 days before), and at the diagnosis of NEC). SCFAs were measured in feces. | 16S ribosomal RNA gene (V3–V4 regions) sequencing Data processing with QIIME v1.9.1. Species classification was performed using silva138/16s_bacteria taxonomic data. | Pre-NEC group vs. controls:Pseudomonadota increased while Bacillota, Actinomycetota, and Bacteroidota decreased (NS). Clostridioides, Blautia, and Clostridium sensu stricto I increased, while unclassified_c_Bacilli, Lactobacillaceae, and Bifidobacterium decreased at the genus level (p < 0.05). At the species level, unclassified_g_Clostridioides, Streptococcus salivarius, and Rothia mucilaginosa increased, while unclassified_c_Bacilli, unclassified species of Lactobacillaceae, and Bifidobacterium animals subsp. lactis decreased (p < 0.05). Acetic, propanoic, butyric, and isovaleric acids decreased. NEC-group vs. controls: Bacillota decreased. Stenotrophomonas, Streptococcus, and Prevotella increased at the genus level. Acetic, propanoic, butyric, and isobutyric acids decreased. NEC vs. pre-NEC group: Pseudomonadota and Bacillota decreased, while Actinomycetota and Bacteroidota increased (NS). At the genus level, Faecalibacterium, Microbacterium, and Solobacterium increased (p < 0.05). There were no differences in any of the SCFAs. Diversity: The alpha diversity (Ace, Chao1) was higher in controls than in pre-NEC patients, but the Simpson and Shannon indices showed no difference. There was no difference between NEC patients and controls. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaplina, A.; Kononova, S.; Zaikova, E.; Pervunina, T.; Petrova, N.; Sitkin, S. Necrotizing Enterocolitis: The Role of Hypoxia, Gut Microbiome, and Microbial Metabolites. Int. J. Mol. Sci. 2023, 24, 2471. https://doi.org/10.3390/ijms24032471
Kaplina A, Kononova S, Zaikova E, Pervunina T, Petrova N, Sitkin S. Necrotizing Enterocolitis: The Role of Hypoxia, Gut Microbiome, and Microbial Metabolites. International Journal of Molecular Sciences. 2023; 24(3):2471. https://doi.org/10.3390/ijms24032471
Chicago/Turabian StyleKaplina, Aleksandra, Svetlana Kononova, Ekaterina Zaikova, Tatiana Pervunina, Natalia Petrova, and Stanislav Sitkin. 2023. "Necrotizing Enterocolitis: The Role of Hypoxia, Gut Microbiome, and Microbial Metabolites" International Journal of Molecular Sciences 24, no. 3: 2471. https://doi.org/10.3390/ijms24032471
APA StyleKaplina, A., Kononova, S., Zaikova, E., Pervunina, T., Petrova, N., & Sitkin, S. (2023). Necrotizing Enterocolitis: The Role of Hypoxia, Gut Microbiome, and Microbial Metabolites. International Journal of Molecular Sciences, 24(3), 2471. https://doi.org/10.3390/ijms24032471