


A Comparative Study between Lycorine and Galantamine Abilities to Interact with AMYLOID β and Reduce In Vitro Neurotoxicity




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
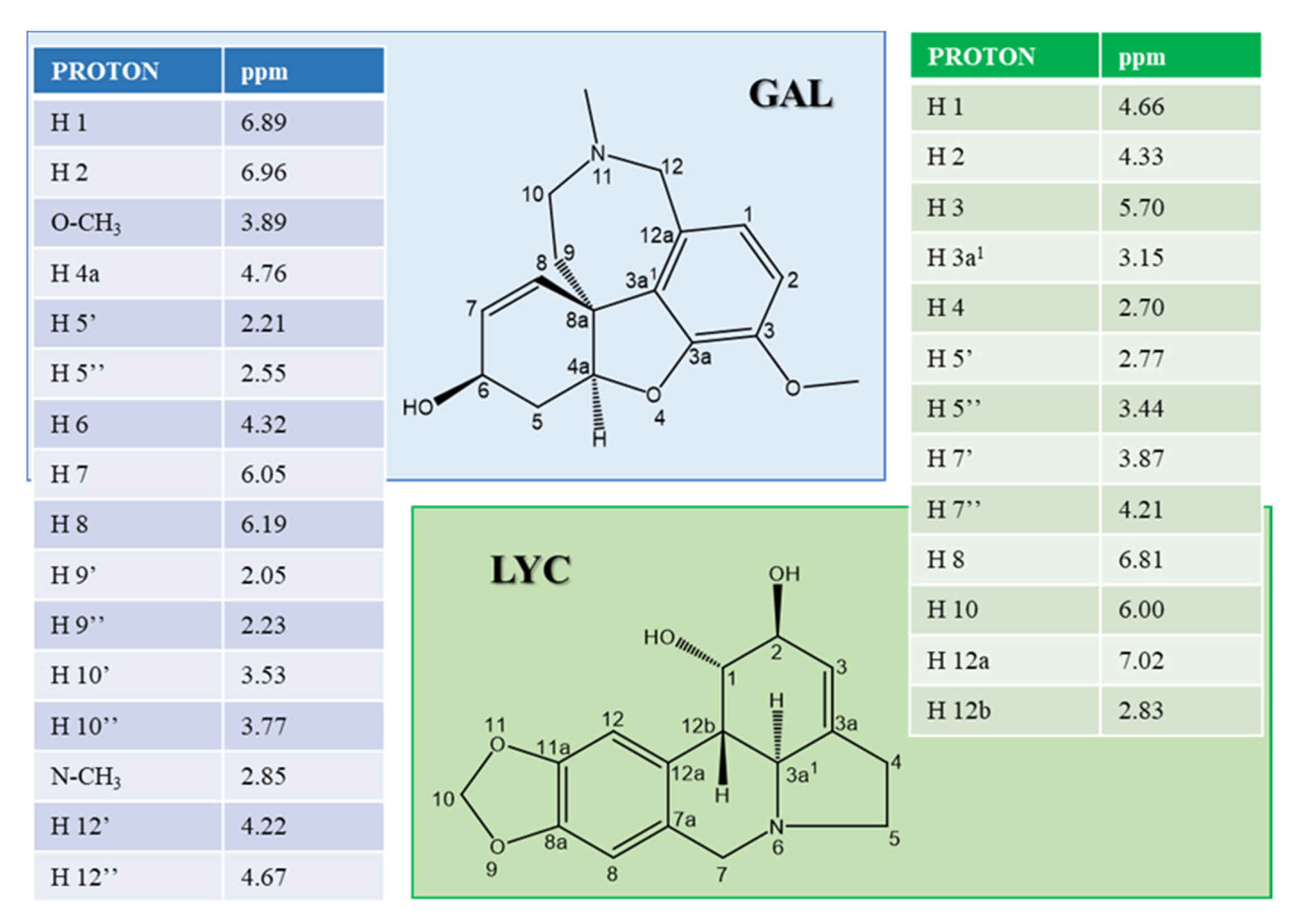
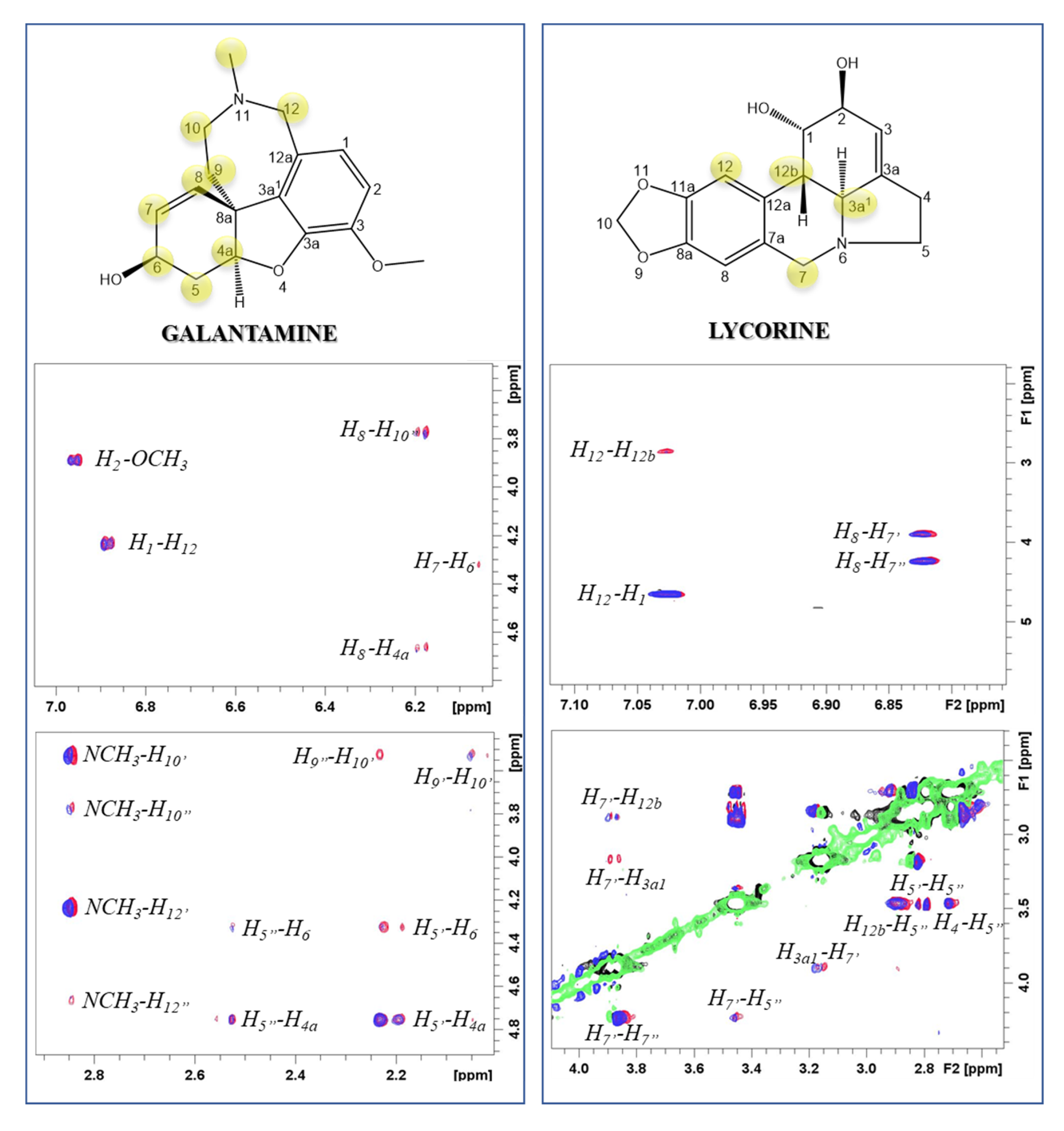
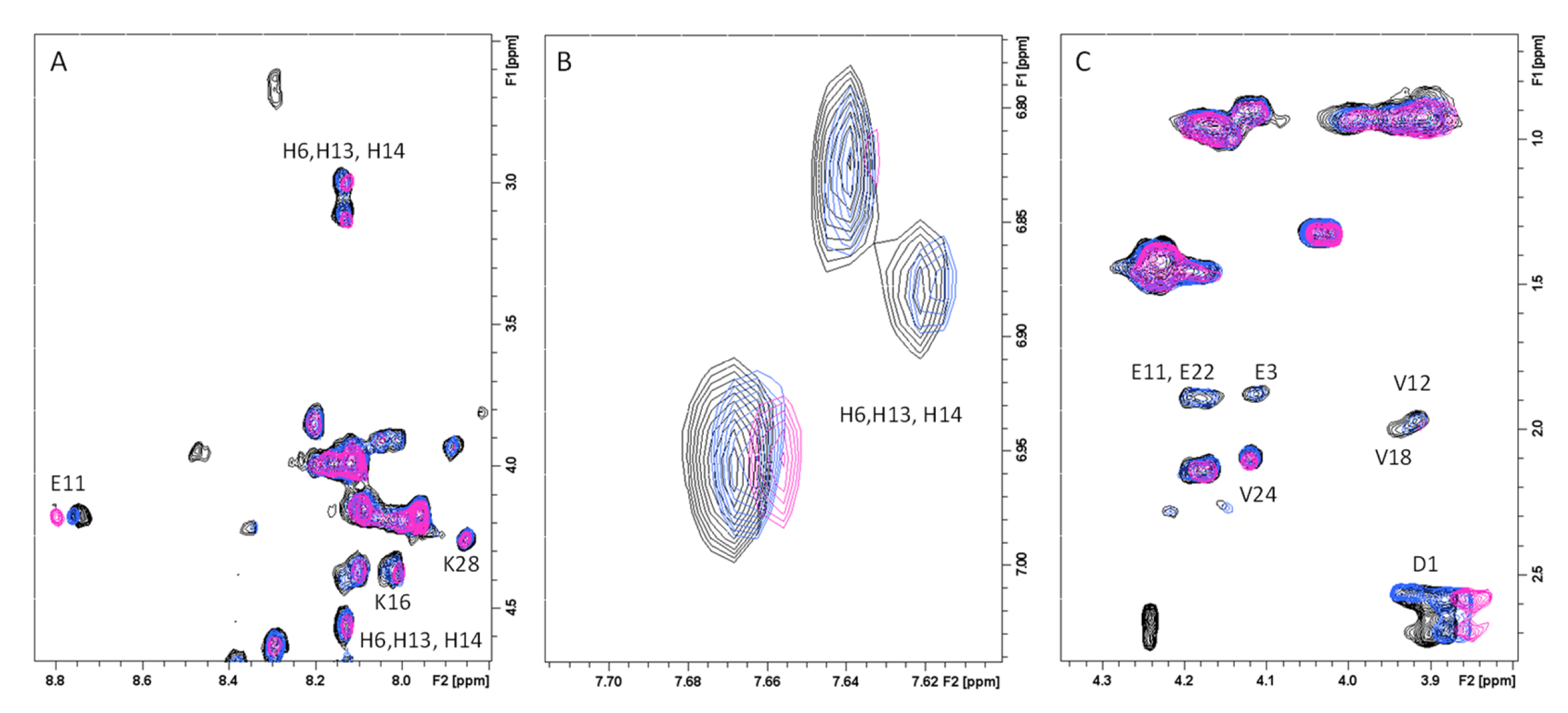
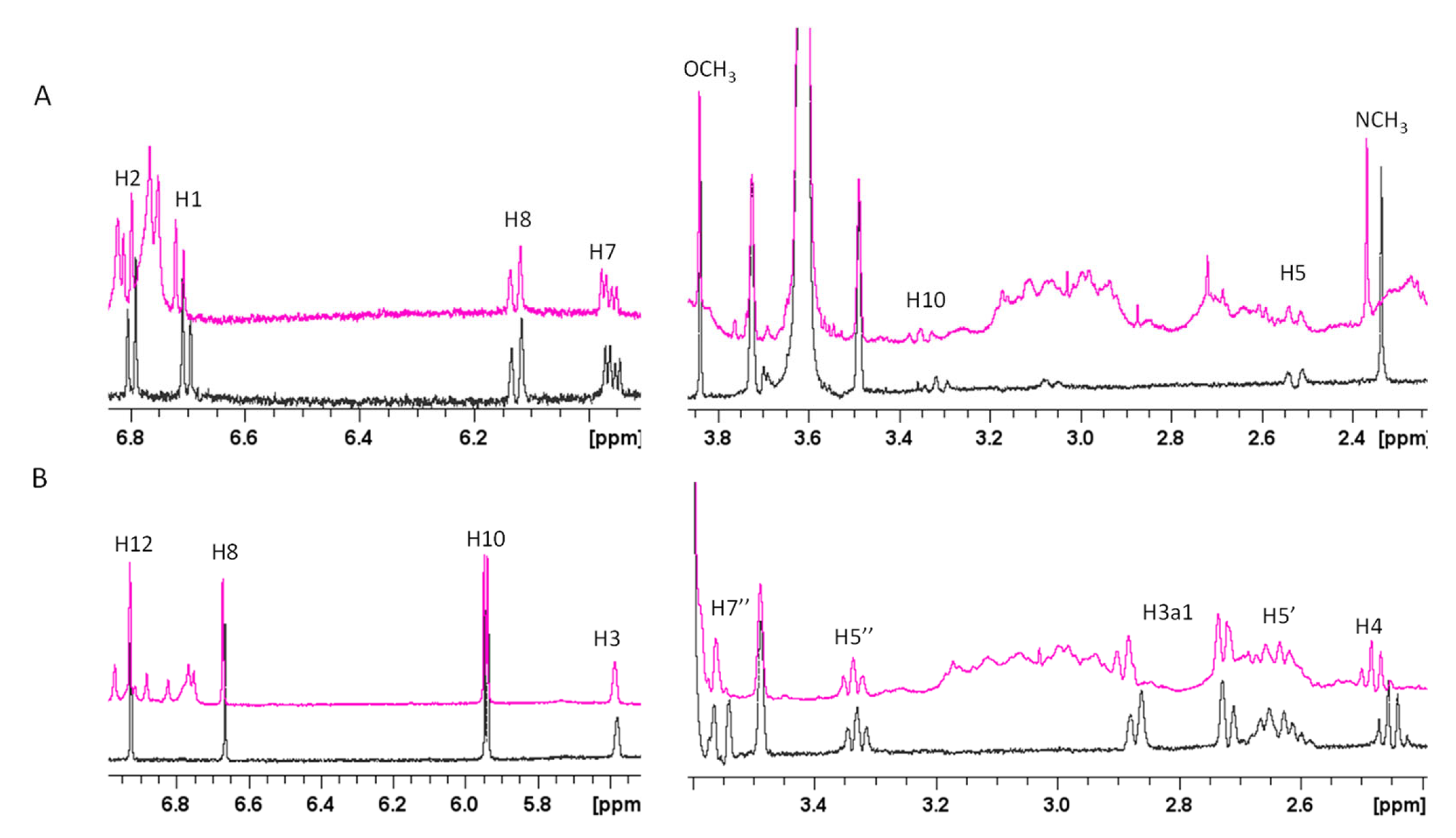
2.1. Atomic and Molecular Interactions between GAL/LYC and Aβ
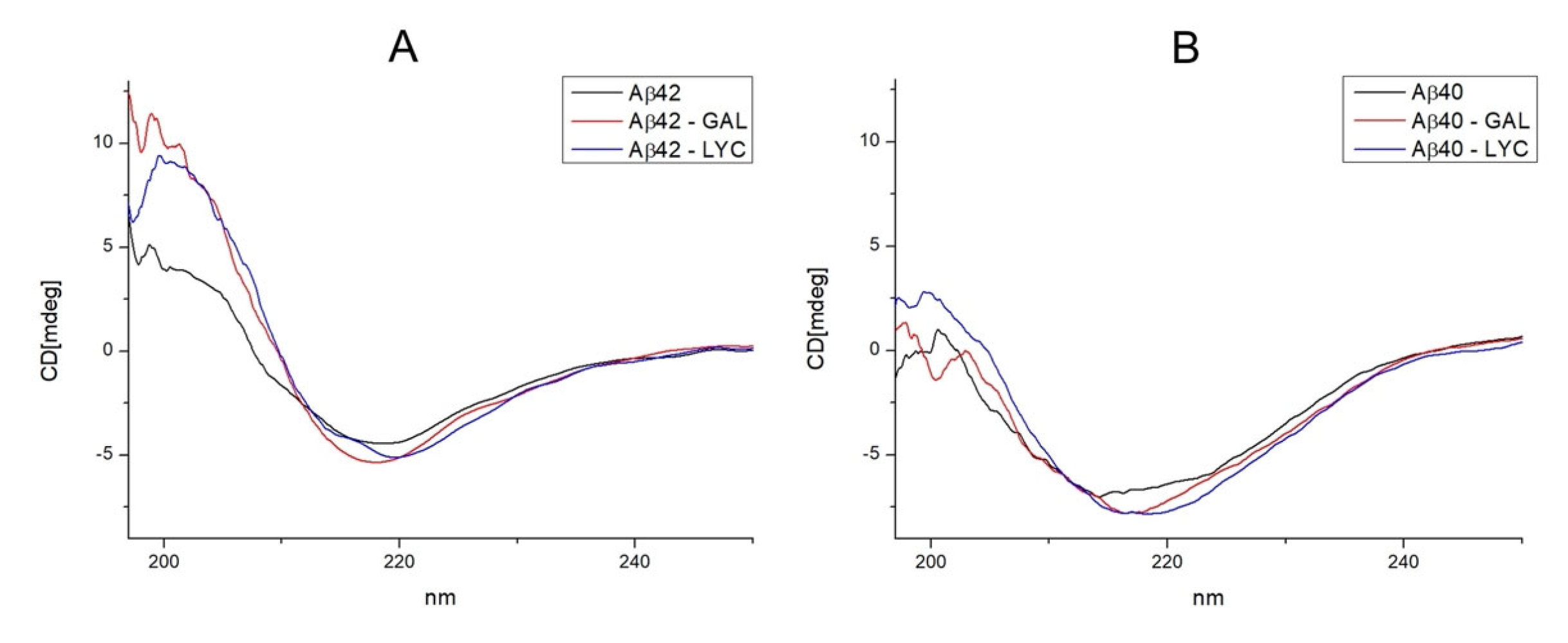
2.2. GAL and LYC Effects on the Secondary Structure of Amyloid-β
2.3. Antioxidant abilities of GAL and LYC
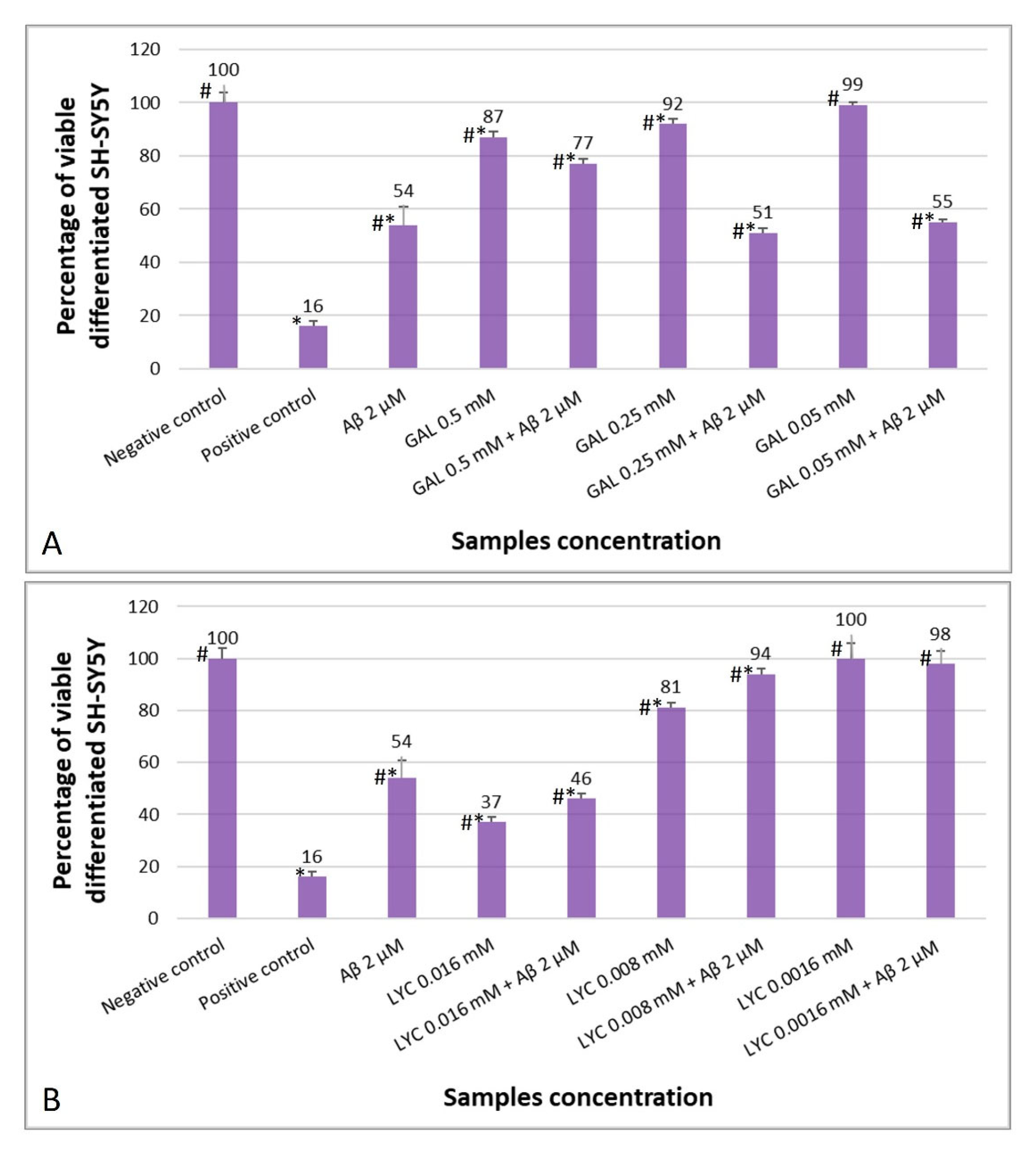
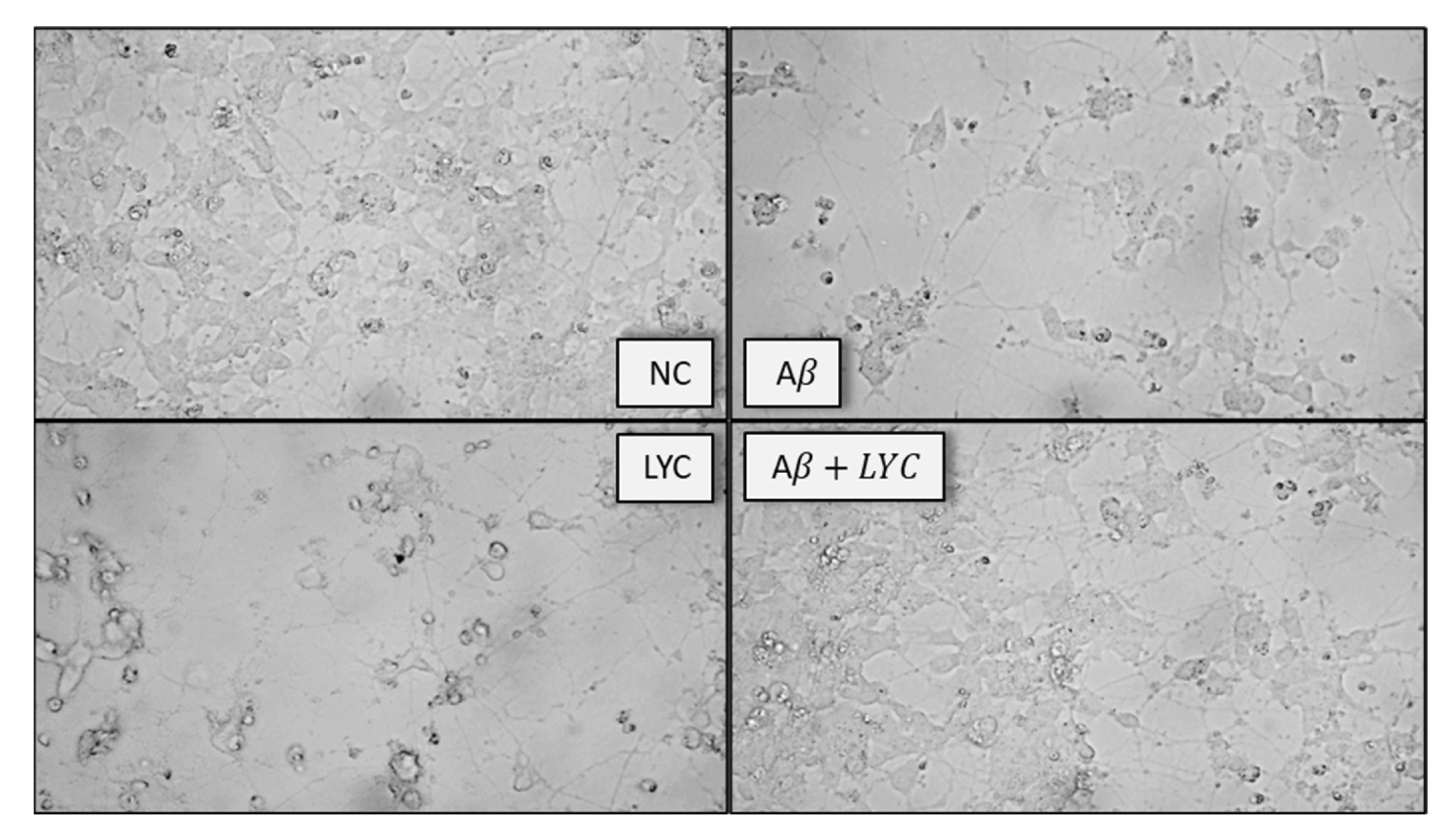
2.4. The Protective Role of GAL and LYC against Aβ-Induced Cytotoxicity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Sample Preparation
5.3. NMR Experiments
5.4. CD Studies
5.5. UV-VIS Studies
5.6. Differentiation of SH-SY5Y Cells
- Day 1: BGM was removed, and Differentiation Media 1 (DM 1) was added to each Petri dish;
- Days 3 and 6: DM 1 was changed;
- Day 7: cells were split as described before, suspended in DM 1 and plated into new 35 mm2 Petri dishes;
- Day 8: DM 1 was removed and Differentiation Medium 2 (DM 2) was added at each dish;
- Day 10: cells were split, suspended in DM 2, seeded in 35 mm2 extracellular matrix (ECM) coated plates and returned to incubator;
- Day 13: DM 2 was removed and Differentiation Medium 3 (DM 3) added;
- Days 14 and 17: DM 3 was changed with fresh medium. At Day 18 neuronal cultures were ready for use.
5.7. Cytotoxicity Evaluation of Test Compounds towards Differentiated SH-SY5Y Cells
5.8. Evaluation of Cell Viability by Neutral Red Uptake (NRU) Test
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheltens:, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s Disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Patterson, C. World Alzheimer Report 2018: The State of the Art of Dementia Research—New Frontiers; Alzheimer’s Disease International: London, UK, 2018. [Google Scholar]
- Gauthier, S.; Rosa-Neto, P.; Morais, J.A.; Webster, C. World Alzheimer Report 2021: Journey through the Diagnosis of Dementia; Alzheimer’s Disease International: London, UK, 2021. [Google Scholar]
- Oumata, N.; Lu, K.; Teng, Y.; Cavé, C.; Peng, Y.; Galons, H.; Roques, B.P. Molecular Mechanisms in Alzheimer’s Disease and Related Potential Treatments Such as Structural Target Convergence of Antibodies and Simple Organic Molecules. Eur. J. Med. Chem. 2022, 240, 114578. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, D.; Gong, X.; Zheng, J. A Mechanistic Survey of Alzheimer’s Disease. Biophys. Chem. 2022, 281, 106735. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Vassar, R.; Strooper, B.D.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; Vos, A.D.; et al. The β-Secretase BACE1 in Alzheimer’s Disease. Biol. Psychiatry 2021, 89, 745. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.-H.; Alghrably, M.; Dhahri, M.; Sharfalddin, A.; Alsiary, R.; Jaremko, M.; Faa, G.; Campagna, M.; Congiu, T.; Piras, M.; et al. Living with the Enemy: From Protein-Misfolding Pathologies We Know, to Those We Want to Know. Ageing Res. Rev. 2021, 70, 101391. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Guo, J.; Ye, X.-Y.; Xie, Y.; Xie, T. Oxidative Stress: The Core Pathogenesis and Mechanism of Alzheimer’s Disease. Ageing Res. Rev. 2022, 77, 101619. [Google Scholar] [CrossRef] [PubMed]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative Stress and the Amyloid Beta Peptide in Alzheimer’s Disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef]
- Stefaniak, E.; Bal, W. CuII Binding Properties of N-Truncated Aβ Peptides: In Search of Biological Function. Inorg. Chem. 2019, 58, 13561–13577. [Google Scholar] [CrossRef] [Green Version]
- Alsiary, R.A.; Alghrably, M.; Saoudi, A.; Al-Ghamdi, S.; Jaremko, L.; Jaremko, M.; Emwas, A.-H. Using NMR Spectroscopy to Investigate the Role Played by Copper in Prion Diseases. Neurol. Sci. 2020, 41, 2389–2406. [Google Scholar] [CrossRef] [Green Version]
- Gaggelli, E.; Kozlowski, H.; Valensin, D.; Valensin, G. Copper Homeostasis and Neurodegenerative Disorders (Alzheimer’s, Prion and Parkinson’s Diseases and Amyotrophic Lateral Sclerosis). Chem. Rev. 2006, 106, 1995–2044. [Google Scholar] [CrossRef]
- Faller, P.; Hureau, C. Metal Ions in Neurodegenerative Diseases. Coord. Chem. Rev. 2012, 256, 2127–2128. [Google Scholar] [CrossRef]
- Viles, J.H. Metal Ions and Amyloid Fiber Formation in Neurodegenerative Diseases. Copper, Zinc and Iron in Alzheimer’s, Parkinson’s and Prion Diseases. Coord. Chem. Rev. 2012, 256, 2271–2284. [Google Scholar] [CrossRef]
- Valensin, D.; Gabbiani, C.; Messori, L. Metal Compounds as Inhibitors of β-Amyloid Aggregation. Perspectives for an Innovative Metallotherapeutics on Alzheimer’s Disease. Coord. Chem. Rev. 2012, 256, 2357–2366. [Google Scholar] [CrossRef]
- Kozlowski, H.; Luczkowski, M.; Remelli, M.; Valensin, D. Copper, Zinc and Iron in Neurodegenerative Diseases (Alzheimer’s, Parkinson’s and Prion Diseases). Coord. Chem. Rev. 2012, 256, 2129–2141. [Google Scholar] [CrossRef]
- Pardo-Moreno, T.; González-Acedo, A.; Rivas-Domínguez, A.; García-Morales, V.; García-Cozar, F.J.; Ramos-Rodríguez, J.J.; Melguizo-Rodríguez, L. Therapeutic Approach to Alzheimer’s Disease: Current Treatments and New Perspectives. Pharmaceutics 2022, 14, 1117. [Google Scholar] [CrossRef]
- Wang, S.; Kong, X.; Chen, Z.; Wang, G.; Zhang, J.; Wang, J. Role of Natural Compounds and Target Enzymes in the Treatment of Alzheimer’s Disease. Molecules 2022, 27, 4175. [Google Scholar] [CrossRef]
- Tolar, M.; Abushakra, S.; Hey, J.A.; Porsteinsson, A.; Sabbagh, M. Aducanumab, Gantenerumab, BAN2401 and ALZ-801-the First Wave of Amyloid-Targeting Drugs for Alzheimer’s Disease with Potential for near Term Approval. Alzheimer’s Res. Ther. 2020, 12, 95. [Google Scholar] [CrossRef]
- Filling the Void for New Alzheimer’s Disease Therapy. Nat. Aging 2021, 1, 567–568. [CrossRef]
- Heller, G.T.; Aprile, F.A.; Michaels, T.C.T.; Limbocker, R.; Perni, M.; Ruggeri, F.S.; Mannini, B.; Löhr, T.; Bonomi, M.; Camilloni, C.; et al. Small-Molecule Sequestration of Amyloid-β as a Drug Discovery Strategy for Alzheimer’s Disease. Sci. Adv. 2020, 6, eabb5924. [Google Scholar] [CrossRef]
- Sannigrahi, A.; Chowdhury, S.; Das, B.; Banerjee, A.; Halder, A.; Kumar, A.; Saleem, M.; Naganathan, A.N.; Karmakar, S.; Chattopadhyay, K. The Metal Cofactor Zinc and Interacting Membranes Modulate SOD1 Conformation-Aggregation Landscape in an in Vitro ALS Model. eLife 2021, 10, e61453. [Google Scholar] [CrossRef]
- Tacrine Granted Marketing Approval for Alzheimer’s Disease. Clin. Pharm. 1993, 12, 873–876.
- Shintani, E.Y.; Uchida, K.M. Donepezil: An Anticholinesterase Inhibitor for Alzheimer’s Disease. Am. J. Health Syst. Pharm. 1997, 54, 2805–2810. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.L. FDA Approves New Cholinesterase Inhibitor for Alzheimer’s Disease. Am. J. Health Syst. Pharm. 2000, 57, 1026–1028. [Google Scholar] [CrossRef]
- Thompson, C.A. FDA Approves Galantamine for Alzheimer’s Disease. Am. J. Health Syst. Pharm. 2001, 58, 649. [Google Scholar] [CrossRef] [PubMed]
- Tuzimski, T.; Petruczynik, A. Determination of Anti-Alzheimer’s Disease Activity of Selected Plant Ingredients. Molecules 2022, 27, 3222. [Google Scholar] [CrossRef]
- Dhahri, M.; Alghrably, M.; Mohammed, H.A.; Badshah, S.L.; Noreen, N.; Mouffouk, F.; Rayyan, S.; Qureshi, K.A.; Mahmood, D.; Lachowicz, J.I.; et al. Natural Polysaccharides as Preventive and Therapeutic Horizon for Neurodegenerative Diseases. Pharmaceutics 2021, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Kola, A.; Dudek, D.; Valensin, D. Metal Complexation Mechanisms of Polyphenols Associated to Alzheimer’s Disease. Curr. Med. Chem. 2021, 28, 7278–7294. [Google Scholar] [CrossRef]
- Kola, A.; Hecel, A.; Lamponi, S.; Valensin, D. Novel Perspective on Alzheimer’s Disease Treatment: Rosmarinic Acid Molecular Interplay with Copper(II) and Amyloid β. Life 2020, 10, 118. [Google Scholar] [CrossRef]
- Cahlíková, L.; Vrabec, R.; Pidaný, F.; Peřinová, R.; Maafi, N.; Mamun, A.A.; Ritomská, A.; Wijaya, V.; Blunden, G. Recent Progress on Biological Activity of Amaryllidaceae and Further Isoquinoline Alkaloids in Connection with Alzheimer’s Disease. Molecules 2021, 26, 5240. [Google Scholar] [CrossRef]
- Cortes, N.; Posada-Duque, R.A.; Alvarez, R.; Alzate, F.; Berkov, S.; Cardona-Gómez, G.P.; Osorio, E. Neuroprotective Activity and Acetylcholinesterase Inhibition of Five Amaryllidaceae Species: A Comparative Study. Life Sci. 2015, 122, 42–50. [Google Scholar] [CrossRef]
- Ding, Y.; Qu, D.; Zhang, K.-M.; Cang, X.-X.; Kou, Z.-N.; Xiao, W.; Zhu, J.-B. Phytochemical and Biological Investigations of Amaryllidaceae Alkaloids: A Review. J. Asian Nat. Prod. Res. 2017, 19, 53–100. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qu, C.; Gao, O.; Hu, X.; Hong, X. Biological and Pharmacological Activities of Amaryllidaceae Alkaloids. RSC Adv. 2015, 5, 16562–16574. [Google Scholar] [CrossRef]
- Koirala, M.; Karimzadegan, V.; Liyanage, N.S.; Mérindol, N.; Desgagné-Penix, I. Biotechnological Approaches to Optimize the Production of Amaryllidaceae Alkaloids. Biomolecules 2022, 12, 893. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Cimmino, A.; Evidente, A. Advances in the Chemical and Biological Characterization of Amaryllidaceae Alkaloids and Natural Analogues Isolated in the Last Decade. Molecules 2020, 25, 5621. [Google Scholar] [CrossRef]
- Kalola, U.K.; Nguyen, H. Galantamine; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Ezoulin, M.J.M.; Ombetta, J.-E.; Dutertre-Catella, H.; Warnet, J.-M.; Massicot, F. Antioxidative Properties of Galantamine on Neuronal Damage Induced by Hydrogen Peroxide in SK-N-SH Cells. Neurotoxicology 2008, 29, 270–277. [Google Scholar] [CrossRef]
- Romero, A.; Egea, J.; García, A.G.; López, M.G. Synergistic Neuroprotective Effect of Combined Low Concentrations of Galantamine and Melatonin against Oxidative Stress in SH-SY5Y Neuroblastoma Cells. J. Pineal Res. 2010, 49, 141–148. [Google Scholar] [CrossRef]
- Arias, E.; Gallego-Sandín, S.; Villarroya, M.; García, A.G.; López, M.G. Unequal Neuroprotection Afforded by the Acetylcholinesterase Inhibitors Galantamine, Donepezil, and Rivastigmine in SH-SY5Y Neuroblastoma Cells: Role of Nicotinic Receptors. J. Pharmacol. Exp. Ther. 2005, 315, 1346–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Wu, D.; Zhang, L.; Zhang, Y. Effects of Galantamine on β-Amyloid Release and Beta-Site Cleaving Enzyme 1 Expression in Differentiated Human Neuroblastoma SH-SY5Y Cells. Exp. Gerontol. 2010, 45, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Matharu, B.; Gibson, G.; Parsons, R.; Huckerby, T.N.; Moore, S.A.; Cooper, L.J.; Millichamp, R.; Allsop, D.; Austen, B. Galantamine Inhibits Beta-Amyloid Aggregation and Cytotoxicity. J. Neurol. Sci. 2009, 280, 49–58. [Google Scholar] [CrossRef]
- Castillo, W.O.; Aristizabal-Pachon, A.F.; de Lima Montaldi, A.P.; Sakamoto-Hojo, E.T.; Takahashi, C.S. Galanthamine Decreases Genotoxicity and Cell Death Induced by β-Amyloid Peptide in SH-SY5Y Cell Line. Neurotoxicology 2016, 57, 291–297. [Google Scholar] [CrossRef]
- Li, Q.; Fang, J.; Yang, M.; Wu, D.; Zhang, L.; Zhang, Y. Galantamine Inhibits Calpain-Calcineurin Signaling Activated by Beta-Amyloid in Human Neuroblastoma SH-SY5Y Cells. Neurosci. Lett. 2010, 480, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Berkov, S.; Atanasova, M.; Georgiev, B.; Bastida Armengol, J.; Doytchinova, I. The Amaryllidaceae Alkaloids: An Untapped Source of Acetylcholinesterase Inhibitors. Phytochem. Rev. 2021, 21, 1415–1443. [Google Scholar] [CrossRef]
- Cortes, N.; Alvarez, R.; Osorio, E.H.; Alzate, F.; Berkov, S.; Osorio, E. Alkaloid Metabolite Profiles by GC/MS and Acetylcholinesterase Inhibitory Activities with Binding-Mode Predictions of Five Amaryllidaceae Plants. J. Pharm. Biomed. Anal. 2015, 102, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Elgorashi, E.E.; Stafford, G.I.; Van Staden, J. Acetylcholinesterase Enzyme Inhibitory Effects of Amaryllidaceae Alkaloids. Planta Med. 2004, 70, 260–262. [Google Scholar] [CrossRef]
- Xiao, H.; Xu, X.; Du, L.; Li, X.; Zhao, H.; Wang, Z.; Zhao, L.; Yang, Z.; Zhang, S.; Yang, Y.; et al. Lycorine and Organ Protection: Review of Its Potential Effects and Molecular Mechanisms. Phytomedicine 2022, 104, 154266. [Google Scholar] [CrossRef]
- Bendaif, H.; Melhaoui, A.; Ramdani, M.; Elmsellem, H.; Douez, C.; El Ouadi, Y. Antibacterial Activity and Virtual Screening by Molecular Docking of Lycorine from Pancratium foetidum Pom (Moroccan Endemic Amaryllidaceae). Microb Pathog 2018, 115, 138–145. [Google Scholar] [CrossRef]
- Wang, G.; Huang, K.; Dong, Y.; Chen, S.; Zhang, J.; Wang, J.; Xie, Z.; Lin, X.; Fang, X.; Fan, S. Lycorine Suppresses Endplate-Chondrocyte Degeneration and Prevents Intervertebral Disc Degeneration by Inhibiting NF-ΚB Signalling Pathway. Cell. Physiol. Biochem. 2018, 45, 1252–1269. [Google Scholar] [CrossRef]
- Kang, J.; Zhang, Y.; Cao, X.; Fan, J.; Li, G.; Wang, Q.; Diao, Y.; Zhao, Z.; Luo, L.; Yin, Z. Lycorine Inhibits Lipopolysaccharide-Induced INOS and COX-2 up-Regulation in RAW264.7 Cells through Suppressing P38 and STATs Activation and Increases the Survival Rate of Mice after LPS Challenge. Int. Immunopharmacol. 2012, 12, 249–256. [Google Scholar] [CrossRef]
- Ying, X.; Huang, A.; Xing, Y.; Lan, L.; Yi, Z.; He, P. Lycorine Inhibits Breast Cancer Growth and Metastasis via Inducing Apoptosis and Blocking Src/FAK-Involved Pathway. Sci. China Life Sci. 2017, 60, 417–428. [Google Scholar] [CrossRef]
- Li, L.; Dai, H.-J.; Ye, M.; Wang, S.-L.; Xiao, X.-J.; Zheng, J.; Chen, H.-Y.; Luo, Y.-H.; Liu, J. Lycorine Induces Cell-Cycle Arrest in the G0/G1 Phase in K562 Cells via HDAC Inhibition. Cancer Cell Int. 2012, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Lamoral-Theys, D.; Andolfi, A.; Van Goietsenoven, G.; Cimmino, A.; Le Calvé, B.; Wauthoz, N.; Mégalizzi, V.; Gras, T.; Bruyère, C.; Dubois, J.; et al. Lycorine, the Main Phenanthridine Amaryllidaceae Alkaloid, Exhibits Significant Antitumor Activity in Cancer Cells That Display Resistance to Proapoptotic Stimuli: An Investigation of Structure-Activity Relationship and Mechanistic Insight. J. Med. Chem. 2009, 52, 6244–6256. [Google Scholar] [CrossRef] [PubMed]
- McNulty, J.; Nair, J.J.; Little, J.R.L.; Brennan, J.D.; Bastida, J. Structure-Activity Studies on Acetylcholinesterase Inhibition in the Lycorine Series of Amaryllidaceae Alkaloids. Bioorg. Med. Chem. Lett. 2010, 20, 5290–5294. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, W.A.M.; de Andrade, J.P.; Chacon, D.S.; Lucas, C.R.; Mariana, E.; de Santis Ferreira, L.; Guaratini, T.; Barbosa, E.G.; Zuanazzi, J.A.; Hallwass, F.; et al. Isoquinoline Alkaloids Reduce Beta-Amyloid Peptide Toxicity in Caenorhabditis Elegans. Nat. Prod. Res. 2021, 35, 4814–4818. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yang, J.; Chen, Y.; Xue, R.; Mao, Z.; Lu, W.; Jiang, Y. Lycorine Hydrochloride Suppresses Stress-Induced Premature Cellular Senescence by Stabilizing the Genome of Human Cells. Aging Cell 2021, 20, e13307. [Google Scholar] [CrossRef] [PubMed]
- Castillo, W.O.; Aristizabal-Pachon, A.F.; Sakamoto-Hojo, E.; Gasca, C.A.; Cabezas-Fajardo, F.A.; Takahashi, C. Caliphruria subedentata (Amaryllidaceae) Decreases Genotoxicity and Cell Death Induced by β-Amyloid Peptide in SH-SY5Y Cell Line. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2018, 836, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Pagano, K.; Tomaselli, S.; Molinari, H.; Ragona, L. Natural Compounds as Inhibitors of Aβ Peptide Aggregation: Chemical Requirements and Molecular Mechanisms. Front. Neurosci. 2020, 14, 619667. [Google Scholar] [CrossRef]
- Gallego-Villarejo, L.; Wallin, C.; Król, S.; Enrich-Bengoa, J.; Suades, A.; Aguilella-Arzo, M.; Gomara, M.J.; Haro, I.; Wärmlander, S.; Muñoz, F.J.; et al. Big Dynorphin Is a Neuroprotector Scaffold against Amyloid β-Peptide Aggregation and Cell Toxicity. Comput. Struct. Biotechnol. J. 2022, 20, 5672–5679. [Google Scholar] [CrossRef]
- Król, S.; Österlund, N.; Vosough, F.; Jarvet, J.; Wärmländer, S.; Barth, A.; Ilag, L.L.; Magzoub, M.; Gräslund, A.; Mörman, C. The Amyloid-Inhibiting NCAM-PrP Peptide Targets Aβ Peptide Aggregation in Membrane-Mimetic Environments. iScience 2021, 24, 102852. [Google Scholar] [CrossRef]
- Lucas, L.H.; Yan, J.; Larive, C.K.; Zartler, E.R.; Shapiro, M.J. Transferred Nuclear Overhauser Effect in Nuclear Magnetic Resonance Diffusion Measurements of Ligand-Protein Binding. Anal. Chem. 2003, 75, 627–634. [Google Scholar] [CrossRef]
- Carlomagno, T. Ligand-Target Interactions: What Can We Learn from NMR? Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 245–266. [Google Scholar] [CrossRef]
- Ludwig, C.; Guenther, U.L. Ligand Based NMR Methods for Drug Discovery. Front. Biosci. 2009, 14, 4565–4574. [Google Scholar] [CrossRef] [PubMed]
- Ormeño, D.; Romero, F.; López-Fenner, J.; Avila, A.; Martínez-Torres, A.; Parodi, J. Ethanol Reduces Amyloid Aggregation in Vitro and Prevents Toxicity in Cell Lines. Arch. Med. Res. 2013, 44, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Andrade, S.; Loureiro, J.A.; Pereira, M.C. Caffeic Acid for the Prevention and Treatment of Alzheimer’s Disease: The Effect of Lipid Membranes on the Inhibition of Aggregation and Disruption of Aβ Fibrils. Int. J. Biol. Macromol. 2021, 190, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Nedaei, H.; Rezaei-Ghaleh, N.; Giller, K.; Becker, S.; Karami, L.; Moosavi-Movahedi, A.A.; Griesinger, C.; Saboury, A.A. The Calcium-Free Form of Atorvastatin Inhibits Amyloid-β(1–42) Aggregation In Vitro. J. Biol. Chem. 2022, 298, 101662. [Google Scholar] [CrossRef]
- Alam, P.; Siddiqi, M.K.; Chaturvedi, S.K.; Zaman, M.; Khan, R.H. Vitamin B12 Offers Neuronal Cell Protection by Inhibiting Aβ-42 Amyloid Fibrillation. Int. J. Biol. Macromol. 2017, 99, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Shishido, N.; Nunomura, A.; Smith, M.A.; Perry, G.; Hayashi, Y.; Nakayama, K.; Hayashi, T. Three Histidine Residues of Amyloid-β Peptide Control the Redox Activity of Copper and Iron. Biochemistry 2007, 46, 12737–12743. [Google Scholar] [CrossRef]
- Guilloreau, L.; Combalbert, S.; Sournia-Saquet, A.; Mazarguil, H.; Faller, P. Redox Chemistry of Copper–Amyloid-β: The Generation of Hydroxyl Radical in the Presence of Ascorbate Is Linked to Redox-Potentials and Aggregation State. ChemBioChem 2007, 8, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Atrián-Blasco, E.; del Barrio, M.; Faller, P.; Hureau, C. Ascorbate Oxidation by Cu(Amyloid-β) Complexes: Determination of the Intrinsic Rate as a Function of Alterations in the Peptide Sequence Revealing Key Residues for Reactive Oxygen Species Production. Anal. Chem. 2018, 90, 5909–5915. [Google Scholar] [CrossRef] [PubMed]
- Petry, F.D.S.; Coelho, B.P.; Gaelzer, M.M.; Kreutz, F.; Guma, F.T.C.R.; Salbego, C.G.; Trindade, V.M.T. Genistein Protects against Amyloid-Beta-Induced Toxicity in SH-SY5Y Cells by Regulation of Akt and Tau Phosphorylation. Phytother. Res. 2020, 34, 796–807. [Google Scholar] [CrossRef]
- Roche, J.; Shen, Y.; Lee, J.H.; Ying, J.; Bax, A. Monomeric Aβ1-40 and Aβ1-42 Peptides in Solution Adopt Very Similar Ramachandran Map Distributions That Closely Resemble Random Coil. Biochemistry 2016, 55, 762–775. [Google Scholar] [CrossRef] [Green Version]
- Meloun, M.; Bordovská, S.; Galla, L. The Thermodynamic Dissociation Constants of Four Non-Steroidal Anti-Inflammatory Drugs by the Least-Squares Nonlinear Regression of Multiwavelength Spectrophotometric PH-Titration Data. J. Pharm. Biomed. Anal. 2007, 45, 552–564. [Google Scholar] [CrossRef] [PubMed]
- T3DB: Lycorine. Available online: http://www.t3db.ca/toxins/T3D4064 (accessed on 20 September 2022).
- Hernández-Rodríguez, M.; Correa-Basurto, J.; Benitez-Cardoza, C.G.; Resendiz-Albor, A.A.; Rosales-Hernández, M.C. In Silico and In Vitro Studies to Elucidate the Role of Cu2+ and Galanthamine as the Limiting Step in the Amyloid Beta (1–42) Fibrillation Process. Protein Sci. 2013, 22, 1320–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, P.P.N.; Mohamed, T.; Osman, W. Investigating the Binding Interactions of Galantamine with β-Amyloid Peptide. Bioorganic Med. Chem. Lett. 2013, 23, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Kovalevich, J.; Langford, D. Considerations for the Use of SH-SY5Y Neuroblastoma Cells in Neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Påhlman, S.; Ruusala, A.I.; Abrahamsson, L.; Mattsson, M.E.; Esscher, T. Retinoic Acid-Induced Differentiation of Cultured Human Neuroblastoma Cells: A Comparison with Phorbolester-Induced Differentiation. Cell Differ. 1984, 14, 135–144. [Google Scholar] [CrossRef]
- Adem, A.; Mattsson, M.E.; Nordberg, A.; Påhlman, S. Muscarinic Receptors in Human SH-SY5Y Neuroblastoma Cell Line: Regulation by Phorbol Ester and Retinoic Acid-Induced Differentiation. Brain Res. 1987, 430, 235–242. [Google Scholar] [CrossRef]
- Shipley, M.M.; Mangold, C.A.; Szpara, M.L. Differentiation of the SH-SY5Y Human Neuroblastoma Cell Line. J. Vis. Exp. 2016, 108, 53193. [Google Scholar] [CrossRef]
- Guarire con le Piante Medicinali—Volume 1. Available online: https://www.aseq.it/medicina-e-discipline-corporee/43936-guarire-con-le-piante-medicinali-volume-1.html (accessed on 30 November 2022).
- Fezoui, Y.; Hartley, D.M.; Harper, J.D.; Khurana, R.; Walsh, D.M.; Condron, M.M.; Selkoe, D.J.; Lansbury, P.T.; Fink, A.L.; Teplow, D.B. An Improved Method of Preparing the Amyloid β-Protein for Fibrillogenesis and Neurotoxicity Experiments. Amyloid 2000, 7, 166–178. [Google Scholar] [CrossRef]
- Hwang, T.L.; Shaka, A.J. Multiple-Pulse Mixing Sequences That Selectively Enhance Chemical Exchange or Cross-Relaxation Peaks in High-Resolution NMR Spectra. J. Magn. Reson. 1998, 135, 280–287. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Stroud, J.C.; Liu, C.; Teng, P.K.; Eisenberg, D. Toxic Fibrillar Oligomers of Amyloid-β Have Cross-β Structure. Proc. Natl. Acad. Sci. USA 2012, 109, 7717–7722. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kola, A.; Lamponi, S.; Currò, F.; Valensin, D. A Comparative Study between Lycorine and Galantamine Abilities to Interact with AMYLOID β and Reduce In Vitro Neurotoxicity. Int. J. Mol. Sci. 2023, 24, 2500. https://doi.org/10.3390/ijms24032500
Kola A, Lamponi S, Currò F, Valensin D. A Comparative Study between Lycorine and Galantamine Abilities to Interact with AMYLOID β and Reduce In Vitro Neurotoxicity. International Journal of Molecular Sciences. 2023; 24(3):2500. https://doi.org/10.3390/ijms24032500
Chicago/Turabian StyleKola, Arian, Stefania Lamponi, Francesco Currò, and Daniela Valensin. 2023. "A Comparative Study between Lycorine and Galantamine Abilities to Interact with AMYLOID β and Reduce In Vitro Neurotoxicity" International Journal of Molecular Sciences 24, no. 3: 2500. https://doi.org/10.3390/ijms24032500
APA StyleKola, A., Lamponi, S., Currò, F., & Valensin, D. (2023). A Comparative Study between Lycorine and Galantamine Abilities to Interact with AMYLOID β and Reduce In Vitro Neurotoxicity. International Journal of Molecular Sciences, 24(3), 2500. https://doi.org/10.3390/ijms24032500