
Chrono-Nutrition: Circadian Rhythm and Personalized Nutrition
 ,
,
Abstract
:1. Introduction
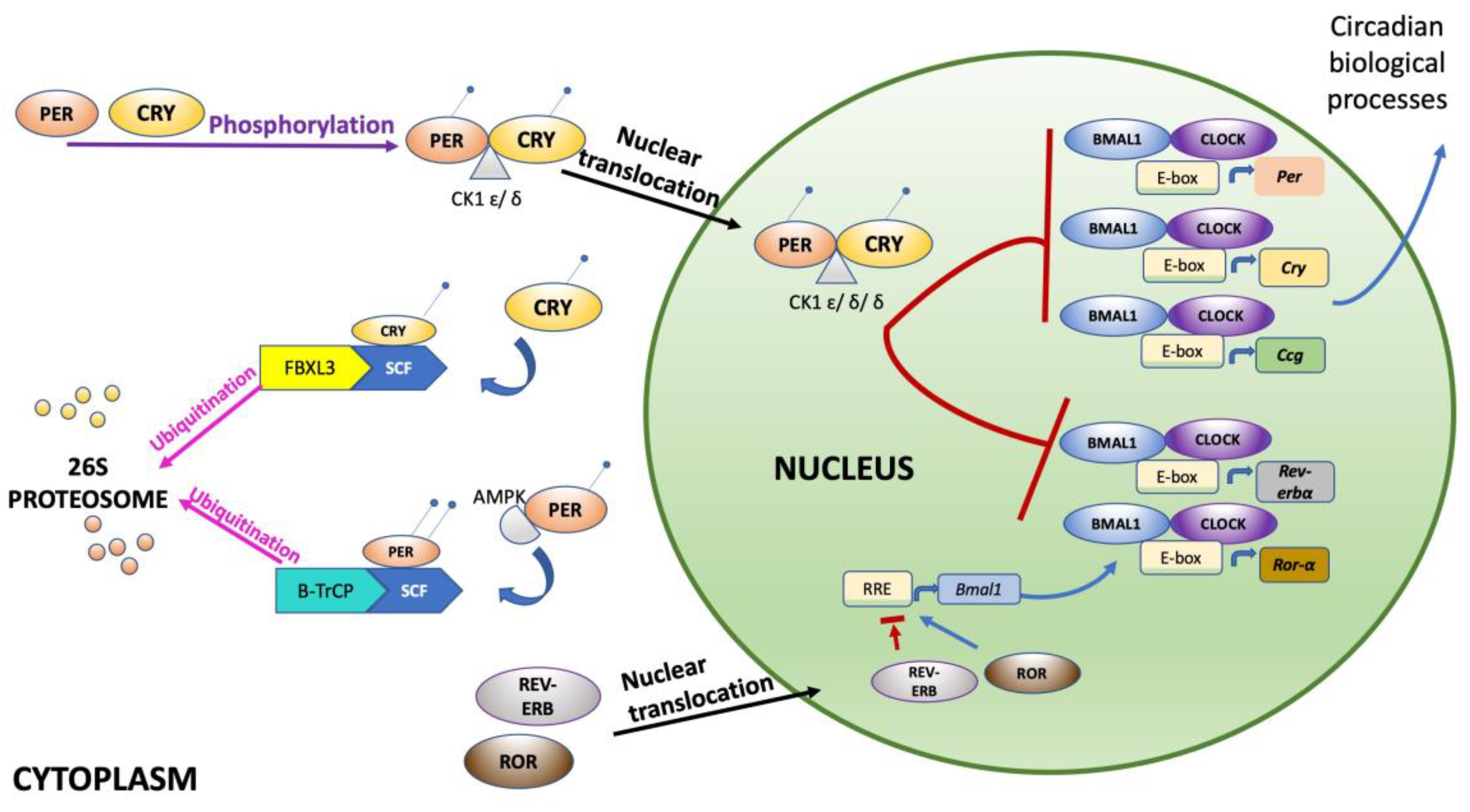
2. Circadian Rhythms
3. Chronotype
4. Chronodisruptions
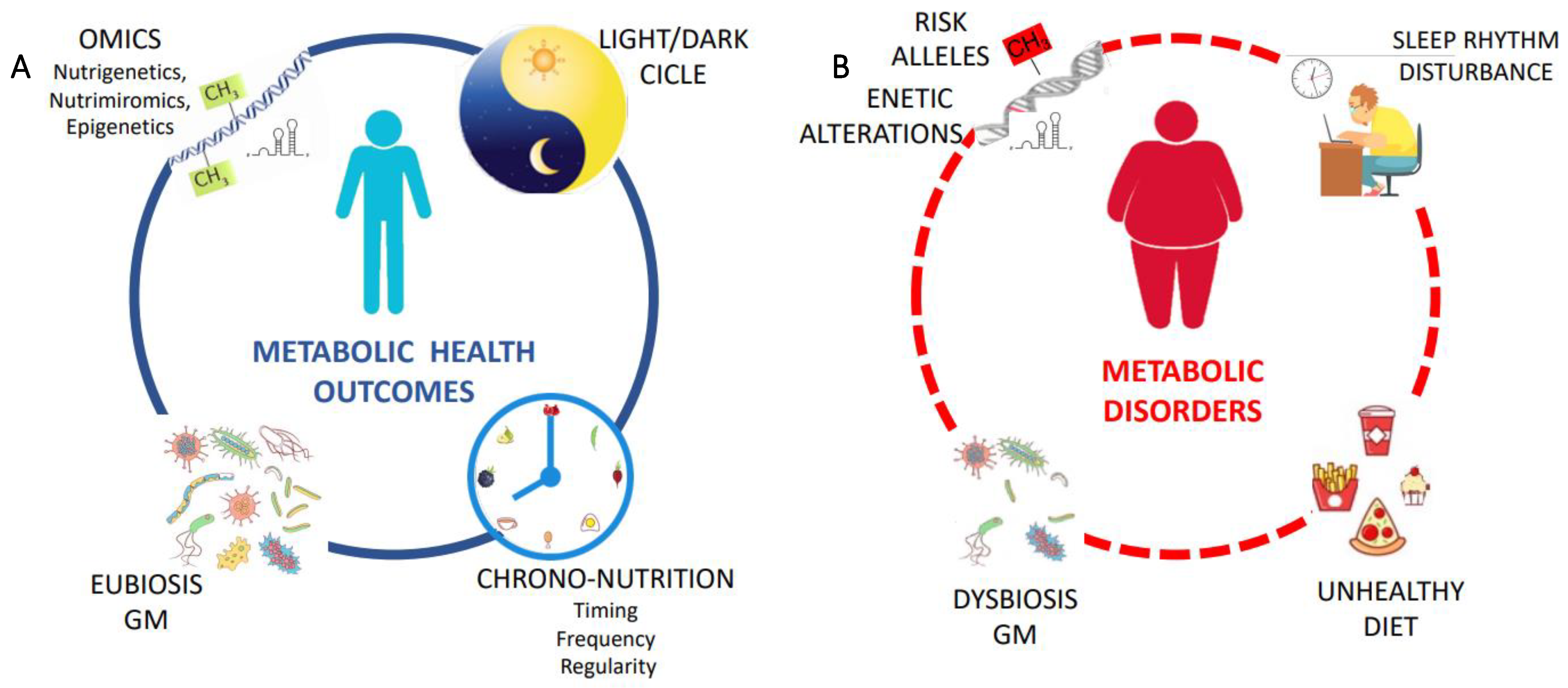
5. Chrono-Nutrition
6. Clock Genes Variants

{kind=link}
{kind=link}
| Authors (Ref) | Sample Size (N) | Study Design | Target Gene (Genetic Variants) | Main Findings |
|---|---|---|---|---|
| Garaulet et al., 2009 [71] | N = 1100 (540 men, 560 women) | Cross-sectional study | CLOCK rs4580704 C > G rs3749474 C > T rs1801260 (3111T→C) rs1464490 C > T rs4864548 G > A | Association with obesity and MetS The minor allele G of rs4580704 showed lower risk of hypertension and diabetes. Protective effect the minor allele G of rs4580704 on insulin sensitivity when MUFA intake was >13.2% of energy. Different effects across CLOCK 3111T→C genotypes for saturated fatty acid intake (% of energy) (p = 0.017). |
| Garaulet et al., 2010 [89] | N = 454 Overweight/obese, aged 20 to 65 | Dietary program based on the Mediterranean diet (28 weeks) | CLOCK rs1801260 (3111T→C) rs3749474 C > T rs4580704 C > G rs1464490 C > T rs4864548 G > A | Relationship between CLOCK SNPs and obesity. CLOCK rs1801260 may predict the outcome of body weight reduction strategies based on low-energy diets. |
| Garaulet et al., 2010a [81] | N = 454 overweight/obese, aged 20 to 65 (380 women, 74 men) | Weight loss intervention based on the Mediterranean diet | PER2 rs230467 C > G rs4663302 C > T rs4663307 G > A | Association with abdominal obesity (p < 0.05) Minor allele carriers G of rs2304672 displayed several obesogenic behaviors. The frequency of the carriers of rs4663307 minor allele who withdrew was greater than in those who successfully completed treatment. |
| Garaulet et al., 2012 [90] | N = 1106 (715 lean and 391 overweight or obese, aged 20 to 65) | Behavioral treatment for obesity based on a Mediterranean diet (30 weeks) | SIRT1-CLOCK rs1467568 G > A 3111T > C | A higher resistance to weight loss and a lower weekly weight loss rate in carriers of minor alleles at SIRT1 and CLOCK loci as compared with homozygotes for both major alleles. |
| Rahati et al., 2022 [75] | N = 403 overweight and/or obese, aged 20 to 50 | Cross-sectional study | CLOCK rs1801260 (3111T→C) | Significant difference between genotypes for physical activity (p = 0.001), waist circumference (p < 0.05), BMI (<0.01), weight (p = 0.001), GLP-1 (p = 0.02), ghrelin (p = 0.04), appetite (p < 0.001), chronotype (p < 0.001), sleep (p < 0.001), food timing (p < 0.001), energy (p < 0.05), carbohydrate (p < 0.05), and fat intake (p < 0.001). |
| Lopez-Guimera et al., 2014 [91] | N = 1272 overweight/obese aged 20 to 65 | Prospective longitudinal study | CLOCK rs1801260 (3111T→C) | SNP interacts with emotional eating behaviors for weight loss. |
| Garaulet et al., 2014a [82] | N = 2414 (1404 Spanish Mediterranean 810 North American populations) | Cross-sectional study | REV-ERB-ALPHA1- rs2314339 G > A | A lower probability of abdominal obesity in minor allele A carriers (OR = 1.5). No significantly association with energy intake but the physical activity was different by genotype. Interaction between the REV-ERB-ALPHA1 variant and MUFA intake for obesity in the Mediterranean population (p = 0.014). |
| Dashti et al., 2014 [87] | N = 1548 (728 Mediterranean and 820 European origin North American populations) | Cross-sectional study | CRY rs2287161 G > C | Significant interactions between the CRY1 variant and dietary carbohydrates for insulin resistance in both populations (p < 0.05). |
| Garaulet et al., 2011 [92] | N = 1495 overweight/obese, aged 20 to 65 years | Cross-sectional study (weight loss program 12–14 weeks) | CLOCK rs1801260 (3111T→C) | Carriers of the minor C allele were more resistant to weight loss, showed shorter sleep duration, higher plasma ghrelin concentrations, delayed breakfast time, evening preference, and less compliance with a Mediterranean diet pattern than TT individuals. |
7. Epigenetic Alterations in the Clock Genes
8. Gut Microbiome
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Green, C.B.; Takahashi, J.S.; Bass, J. The Meter of Metabolism. Cell 2008, 134, 728–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, J.; Takahashi, J.S. Circadian Integration of Metabolism and Energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, S.M.; Udoh, U.S.; E Young, M. Circadian regulation of metabolism. J. Endocrinol. 2014, 222, R75–R96. [Google Scholar] [CrossRef] [Green Version]
- Oosterman, J.E.; Kalsbeek, A.; la Fleur, S.E.; Belsham, D.D. Impact of nutrients on circadian rhythmicity. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R337–R350. [Google Scholar] [CrossRef] [Green Version]
- Corbalán-Tutau, M.; Gómez-Abellán, P.; Madrid, J.A.; Canteras, M.; Ordovas, J.; Garaulet, M. Toward a chronobiological characterization of obesity and metabolic syndrome in clinical practice. Clin. Nutr. 2015, 34, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Bandín, C.; Martinez-Nicolas, A.; Ordovás, J.M.; A Madrid, J.; Garaulet, M. Circadian rhythmicity as a predictor of weight-loss effectiveness. Int. J. Obes. 2013, 38, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Micó, V.; Díez-Ricote, L.; Daimiel, L. Nutrigenetics and Nutrimiromics of the Circadian System: The Time for Human Health. Int. J. Mol. Sci. 2016, 17, 299. [Google Scholar] [CrossRef] [Green Version]
- Hawley, J.A.; Sassone-Corsi, P.; Zierath, J.R. Chrono-nutrition for the prevention and treatment of obesity and type 2 diabetes: From mice to men. Diabetologia 2020, 63, 2253–2259. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Chaix, A.; Panda, S. Daily Eating Patterns and Their Impact on Health and Disease. Trends Endocrinol. Metab. 2015, 27, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Bishehsari, F.; Voigt, R.M.; Keshavarzian, A. Circadian rhythms and the gut microbiota: From the metabolic syndrome to cancer. Nat. Rev. Endocrinol. 2020, 16, 731–739. [Google Scholar] [CrossRef]
- Oh, E.S.; Petronis, A. Origins of human disease: The chrono-epigenetic perspective. Nat. Rev. Genet. 2021, 22, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Qi, L. Nutrition for precision health: The time is now. Obesity 2022, 30, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulli, G.; Manoogian, E.N.; Taub, P.R.; Panda, S. Training the Circadian Clock, Clocking the Drugs, and Drugging the Clock to Prevent, Manage, and Treat Chronic Diseases. Trends Pharmacol. Sci. 2018, 39, 812–827. [Google Scholar] [CrossRef]
- Asher, G.; Sassone-Corsi, P. Time for Food: The Intimate Interplay between Nutrition, Metabolism, and the Circadian Clock. Cell 2015, 161, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.S.; Hong, H.-K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Nguyen, K.M.; Busino, L. The Biology of F-box Proteins: The SCF Family of E3 Ubiquitin Ligases. J. Biomed. Sci. 2020, 1217, 111–122. [Google Scholar] [CrossRef]
- Sakamoto, A.; Terui, Y.; Uemura, T.; Igarashi, K.; Kashiwagi, K. Translational Regulation of Clock Genes BMAL1 and REV-ERBα by Polyamines. Int. J. Mol. Sci. 2021, 22, 1307. [Google Scholar] [CrossRef]
- Maury, E. Off the Clock: From Circadian Disruption to Metabolic Disease. Int. J. Mol. Sci. 2019, 20, 1597. [Google Scholar] [CrossRef] [Green Version]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar]
- Lack, L.; Bailey, M.; Lovato, N.; Wright, H. Chronotype differences in circadian rhythms of temperature, melatonin, and sleepiness as measured in a modified constant routine protocol. Nat. Sci. Sleep 2009, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Yun, C.-H.; Ahn, J.H.; Suh, S.; Cho, H.J.; Lee, S.K.; Yoo, H.J.; Seo, J.A.; Kim, S.G.; Choi, K.M.; et al. Evening Chronotype Is Associated With Metabolic Disorders and Body Composition in Middle-Aged Adults. J. Clin. Endocrinol. Metab. 2015, 100, 1494–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaruli, A.; Castelli, L.; Mulè, A.; Scurati, R.; Esposito, F.; Galasso, L.; Roveda, E. Biological Rhythm and Chronotype: New Perspectives in Health. Biomolecules 2021, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Bhar, D.; Bagepally, B.S.; Rakesh, B. Association between chronotype and cardio-vascular disease risk factors: A systematic review and meta-analysis. Clin. Epidemiol. Glob. Health 2022, 16, 101108. [Google Scholar] [CrossRef]
- Natale, V.; Ballardini, D.; Schumann, R.; Mencarelli, C.; Magelli, V. Morningness–eveningness preference and eating dis-orders. Personal. Individ. Differ. 2008, 45, 549–553. [Google Scholar] [CrossRef]
- Almoosawi, S.; Vingeliene, S.; Gachon, F.; Voortman, T.; Palla, L.; Johnston, J.D.; Van Dam, R.M.; Darimont, C.; Karagounis, L.G. Chronotype: Implications for Epidemiologic Studies on Chrono-Nutrition and Cardiometabolic Health. Adv. Nutr. Int. Rev. J. 2018, 10, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Merikanto, I.; Lahti, T.; Puolijoki, H.; Vanhala, M.; Peltonen, M.; Laatikainen, T.; Vartiainen, E.; Salomaa, V.; Kronholm, E.; Partonen, T. Associations of Chronotype and Sleep With Cardiovascular Diseases and Type 2 Diabetes. Chronobiol. Int. 2013, 30, 470–477. [Google Scholar] [CrossRef]
- Knutson, K.L.; von Schantz, M. Associations between chronotype, morbidity and mortality in the UK Biobank cohort. Chronobiol. Int. 2018, 35, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Reutrakul, S.; Hood, M.M.; Crowley, S.J.; Morgan, M.K.; Teodori, M.; Knutson, K.L. The Relationship Between Breakfast Skipping, Chronotype, and Glycemic Control in Type 2 Diabetes. Chronobiol. Int. 2013, 31, 64–71. [Google Scholar] [CrossRef]
- Kasof, J. Eveningness and bulimic behavior. Personal. Individ. Differ. 2001, 31, 361–369. [Google Scholar] [CrossRef]
- Tortorella, A.; Monteleone, P.; Martiadis, V.; Perris, F.; Maj, M. The 3111T/C polymorphism of the CLOCK gene confers a predisposition to a lifetime lower body weight in patients with anorexia nervosa and bulimia nervosa: A preliminary study. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2007, 144B, 992–995. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Xu, F.; Storfer-Isser, A.; Thomas, A.; Ievers-Landis, C.E.; Redline, S. The association of sleep duration with ado-lescents’ fat and carbohydrate consumption. Sleep 2010, 33, 1201–1209. [Google Scholar] [CrossRef] [Green Version]
- Arora, T.; Taheri, S. Associations among late chronotype, body mass index and dietary behaviors in young adolescents. Int. J. Obes. 2014, 39, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Rudic, R.D.; McNamara, P.; Curtis, A.-M.; Boston, R.C.; Panda, S.; HogenEsch, J.B.; Fitzgerald, G.A. BMAL1 and CLOCK, Two Essential Components of the Circadian Clock, Are Involved in Glucose Homeostasis. PLoS Biol. 2004, 2, e377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Zhao, X.; Hatori, M.; Yu, R.T.; Barish, G.D.; Lam, M.T.; Chong, L.W.; DiTacchio, L.; Atkins, A.R.; Glass, C.K.; et al. Regulation of circadian behaviour and metabolism by REV-ERB-α and REV-ERB-β. Nature 2012, 485, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Erren, T.C.; Reiter, R.J.; Piekarski, C. Light, timing of biological rhythms, and chronodisruption in man. Sci. Nat. 2003, 90, 485–494. [Google Scholar] [CrossRef]
- Chaput, J.P.; Tremblay, A. Does short sleep duration favor abdominal adiposity in children? Int. J. Pediatr. Obes. 2007, 2, 188–191. [Google Scholar] [CrossRef]
- Chaput, J.P.; Dutil, C.; Featherstone, R.; Ross, R.; Giangregorio, L.; Saunders, T.J.; Janssen, I.; Poitras, V.J.; Kho, M.E.; Ross-White, A.; et al. Sleep duration and health in adults: An overview of systematic reviews. Appl. Physiol. Nutr. Metab. 2020, 45, S218–S231. [Google Scholar] [CrossRef]
- Beihl, D.A.; Liese, A.D.; Haffner, S.M. Sleep Duration as a Risk Factor for Incident Type 2 Diabetes in a Multiethnic Cohort. Ann. Epidemiol. 2009, 19, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Gangwisch, J.E. Epidemiological evidence for the links between sleep, circadian rhythms and metabolism. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2009, 10, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Eckel-Mahan, K.L.; Patel, V.R.; de Mateo, S.; Orozco-Solis, R.; Ceglia, N.J.; Sahar, S.; Dilag-Penilla, S.A.; Dyar, K.A.; Baldi, P.; Sassone-Corsi, P. Reprogramming of the Circadian Clock by Nutritional Challenge. Cell 2013, 155, 1464–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaput, J.-P.; Després, J.-P.; Bouchard, C.; Tremblay, A. Association of sleep duration with type 2 diabetes and impaired glucose tolerance. Diabetologia 2007, 50, 2298–2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchison, A.T.; Wittert, G.A.; Heilbronn, L.K. Matching Meals to Body Clocks—Impact on Weight and Glucose Metabolism. Nutrients 2017, 9, 222. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Feng, W.; Wang, F.; Li, P.; Li, Z.; Li, M.; Tse, G.; Vlaanderen, J.; Vermeulen, R.; Tse, L.A. Meta-analysis on shift work and risks of specific obesity types. Obes. Rev. 2017, 19, 28–40. [Google Scholar] [CrossRef]
- Scheer, F.A.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian mis-alignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [Green Version]
- Roenneberg, T.; Allebrandt, K.V.; Merrow, M.; Vetter, C. Social Jetlag and Obesity. Curr. Biol. 2012, 22, 939–943. [Google Scholar] [CrossRef] [Green Version]
- Beebe, D.W.; Simon, S.; Summer, S.; Hemmer, S.; Strotman, D.; Dolan, L.M. Dietary Intake Following Experimentally Restricted Sleep in Adolescents. Sleep 2013, 36, 827–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingenberg, L.; Chaput, J.-P.; Holmbäck, U.; Visby, T.; Jennum, P.; Nikolic, M.; Astrup, A.; Sjödin, A. Acute Sleep Restriction Reduces Insulin Sensitivity in Adolescent Boys. Sleep 2013, 36, 1085–1090. [Google Scholar] [CrossRef] [Green Version]
- Shapira, N. The Metabolic Concept of Meal Sequence vs. Satiety: Glycemic and Oxidative Responses with Reference to In-flammation Risk, Protective Principles and Mediterranean Diet. Nutrients 2019, 11, 2373. [Google Scholar] [CrossRef]
- Papakonstantinou, E.; Oikonomou, C.; Nychas, G.; Dimitriadis, G.D. Effects of Diet, Lifestyle, Chrononutrition and Alternative Dietary Interventions on Postprandial Glycemia and Insulin Resistance. Nutrients 2022, 14, 823. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, H.; Genzer, Y.; Cohen, R.; Chapnik, N.; Madar, Z.; Froy, O. Timed high-fat diet resets circadian metabolism and prevents obesity. FASEB J. 2012, 26, 3493–3502. [Google Scholar] [CrossRef]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-Restricted Feeding Is a Preventative and Therapeutic Intervention against Diverse Nutritional Challenges. Cell Metab. 2014, 20, 991–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamovich, Y.; Rousso-Noori, L.; Zwighaft, Z.; Neufeld-Cohen, A.; Golik, M.; Kraut-Cohen, J.; Wang, M.; Han, X.; Asher, G. Circadian Clocks and Feeding Time Regulate the Oscillations and Levels of Hepatic Triglycerides. Cell Metab. 2014, 19, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular Mechanisms and Clinical Applications. Cell Metab. 2014, 19, 181–192. [Google Scholar] [CrossRef] [Green Version]
- De Cabo, R.; Mattson, M.P. Effects of intermittent fasting on health, aging, and disease. N. Engl. J. Med. 2019, 381, 2541–2551. [Google Scholar] [CrossRef]
- Delabos, A.; Rapin, J.R. Mincir sur Mesure: Grâce à la Chrono-Nutrition; Éditions Albin Michel: Paris, France, 2005. [Google Scholar]
- Tarquini, R.; Mazzoccoli, G. Clock Genes, Metabolism, and Cardiovascular Risk. Heart Fail. Clin. 2017, 13, 645–655. [Google Scholar] [CrossRef]
- Thomas, E.A.; Zaman, A.; Cornier, M.-A.; Catenacci, V.A.; Tussey, E.J.; Grau, L.; Arbet, J.; Broussard, J.L.; Rynders, C.A. Later Meal and Sleep Timing Predicts Higher Percent Body Fat. Nutrients 2020, 13, 73. [Google Scholar] [CrossRef]
- Dashti, H.S.; Gómez-Abellán, P.; Qian, J.; Esteban, A.; Morales, E.; Scheer, F.A.J.L.; Garaulet, M. Late eating is associated with cardiometabolic risk traits, obesogenic behaviors, and impaired weight loss. Am. J. Clin. Nutr. 2020, 113, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Crispim, C.A.; Mota, M.C. New perspectives on chrononutrition. Biol. Rhythm. Res. 2018, 50, 63–77. [Google Scholar] [CrossRef]
- Flanagan, A.; Bechtold, D.A.; Pot, G.K.; Johnston, J.D. Chrono-nutrition: From molecular and neuronal mechanisms to human epidemiology and timed feeding patterns. J. Neurochem. 2020, 157, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Pot, G.K. Chrono-nutrition—An emerging, modifiable risk factor for chronic disease? Nutr. Bull. 2021, 46, 114–119. [Google Scholar] [CrossRef]
- Tahara, Y.; Shibata, S. Chrono-biology, chrono-pharmacology, and chrono-nutrition. J. Pharmacol. Sci. 2014, 124, 320–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyssenko, V.; Nagorny, C.L.; Erdos, M.R.; Wierup, N.; Jonsson, A.; Spégel, P.; Bugliani, M.; Saxena, R.; Fex, M.; Pulizzi, N.; et al. Common variant in MTNR1B associated with increased risk of type 2 diabetes and impaired early insulin secretion. Nat. Genet. 2009, 41, 82–88. [Google Scholar] [CrossRef]
- Prokopenko, I.; Langenberg, C.; Florez, J.C.; Saxena, R.; Soranzo, N.; Thorleifsson, G.; Loos, R.J.F.; Manning, A.K.; Jackson, A.U.; Aulchenko, Y.; et al. Variants in MTNR1B influence fasting glucose levels. Nat. Genet. 2008, 41, 77–81. [Google Scholar] [CrossRef]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; The Meta-Analysis of Glucose and Insulin-Related Traits Consortium (MAGIC); Clément, N.; Fawcett, K.; Yengo, L.; Vaillant, E.; Guillaume, J.-L.; Dechaume, A.; Payne, F.; Roussel, R.; et al. Rare MTNR1B variants impairing melatonin receptor 1B function contribute to type 2 diabetes. Nat. Genet. 2012, 44, 297–301. [Google Scholar] [CrossRef]
- Garaulet, M.; Lee, Y.-C.; Shen, J.; Parnell, L.D.; Arnett, D.K.; Tsai, M.Y.; Lai, C.-Q.; Ordovas, J.M. CLOCK genetic variation and metabolic syndrome risk: Modulation by monounsaturated fatty acids. Am. J. Clin. Nutr. 2009, 90, 1466–1475. [Google Scholar] [CrossRef] [Green Version]
- Scott, E.M.; Carter, A.M.; Grant, P.J. Association between polymorphisms in the Clock gene, obesity and the metabolic syndrome in man. Int. J. Obes. 2007, 32, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Gemma, C.; Gianotti, T.F.; Burgueño, A.; Castaño, G.; Pirola, C.J. Genetic variants of Clock transcription factor are associated with individual susceptibility to obesity. Am. J. Clin. Nutr. 2008, 87, 1606–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Minguez, J.; Gómez-Abellán, P.; Garaulet, M. Circadian rhythms, food timing and obesity. Proc. Nutr. Soc. 2016, 75, 501–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahati, S.; Qorbani, M.; Naghavi, A.; Nia, M.H.; Pishva, H. Association between CLOCK 3111 T/C polymorphism with ghrelin, GLP-1, food timing, sleep and chronotype in overweight and obese Iranian adults. BMC Endocr. Disord. 2022, 22, 147. [Google Scholar] [CrossRef] [PubMed]
- Grotenfelt, N.E.; Wasenius, N.S.; Rönö, K.; Laivuori, H.; Stach-Lempinen, B.; Orho-Melander, M.; Schulz, C.-A.; Kautiainen, H.; Koivusalo, S.B.; Eriksson, J.G. Interaction between rs10830963 polymorphism in MTNR1B and lifestyle intervention on occurrence of gestational diabetes. Diabetologia 2016, 59, 1655–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alharbi, K.K.; Al-Sulaiman, A.M.; Bin Shedaid, M.K.; Al-Shangiti, A.M.; Marie, M.; Al-Sheikh, Y.A.; Khan, I.A. MTNR1B genetic polymorphisms as risk factors for gestational diabetes mellitus: A case-control study in a single tertiary care center. Ann. Saudi Med. 2019, 39, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Woon, P.Y.; Kaisaki, P.J.; Bragança, J.; Bihoreau, M.-T.; Levy, J.C.; Farrall, M.; Gauguier, D. Aryl hydrocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 14412–14417. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.A.; Rees, S.D.; Hydrie, M.Z.I.; Shera, A.S.; Bellary, S.; O’Hare, J.P.; Kumar, S.; Taheri, S.; Basit, A.; Barnett, A.H.; et al. Circadian Gene Variants and Susceptibility to Type 2 Diabetes: A Pilot Study. PLoS ONE 2012, 7, e32670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappa, K.I.; Gazouli, M.; Anastasiou, E.; Iliodromiti, Z.; Antsaklis, A.; Anagnou, N.P. The major circadian pacemaker ARNT-like protein-1 (BMAL1) is associated with susceptibility to gestational diabetes mellitus. Diabetes Res. Clin. Pract. 2012, 99, 151–157. [Google Scholar] [CrossRef]
- Garaulet, M.; Corbalán-Tutau, M.D.; Madrid, J.A.; Baraza, J.C.; Parnell, L.D.; Lee, Y.-C.; Ordovas, J.M. PERIOD2 Variants Are Associated with Abdominal Obesity, Psycho-Behavioral Factors, and Attrition in the Dietary Treatment of Obesity. J. Am. Diet. Assoc. 2010, 110, 917–921. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Smith, C.E.; Gomez-Abellán, P.; Ordovás-Montañés, M.; Lee, Y.C.; Parnell, L.D.; Arnett, D.K.; Ordovás, J.M. REV-ERB-ALPHA circadian gene variant associates with obesity in two independent populations: Mediterranean and North American. Mol. Nutr. Food Res. 2014, 58, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Molina-Montes, E.; Rodríguez-Barranco, M.; Ching-López, A.; Artacho, R.; Huerta, J.M.; Amiano, P.; Lasheras, C.; Moreno-Iribas, C.; Jimenez-Zabala, A.; Chirlaque, M.D.; et al. Circadian clock gene variants and their link with chronotype, chrononutrition, sleeping patterns and obesity in the European prospective investigation into cancer and nutrition (EPIC) study. Clin. Nutr. 2022, 41, 1977–1990. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Lee, Y.-C.; Shen, J.; Parnell, L.D.; Arnett, D.K.; Tsai, M.Y.; Lai, C.-Q.; Ordovas, J.M. Genetic variants in human CLOCK associate with total energy intake and cytokine sleep factors in overweight subjects (GOLDN population). Eur. J. Hum. Genet. 2009, 18, 364–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzago, M.; Santurbano, D.; Vitacolonna, E.; Stuppia, L. Genes and Diet in the Prevention of Chronic Diseases in Future Generations. Int. J. Mol. Sci. 2020, 21, 2633. [Google Scholar] [CrossRef] [PubMed]
- Franzago, M.; Di Nicola, M.; Fraticelli, F.; Marchioni, M.; Stuppia, L.; Vitacolonna, E. Nutrigenetic variants and response to diet/lifestyle intervention in obese subjects: A pilot study. Acta Diabetol. 2022, 59, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Dashti, H.S.; Smith, C.E.; Lee, Y.-C.; Parnell, L.D.; Lai, C.-Q.; Arnett, D.K.; Ordovás, J.M.; Garaulet, M. CRY1 circadian gene variant interacts with carbohydrate intake for insulin resistance in two independent populations: Mediterranean and North American. Chronobiol. Int. 2014, 31, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Uemura, H.; Katsuura-Kamano, S.; Yamaguchi, M.; Arisawa, K.; Hamajima, N.; Hishida, A.; Kawai, S.; Oze, I.; Shinchi, K.; Takashima, N.; et al. Variant of the clock circadian regulator (CLOCK) gene and related haplotypes are associated with the prevalence of type 2 diabetes in the Japanese population. J. Diabetes 2015, 8, 667–676. [Google Scholar] [CrossRef]
- Garaulet, M.; Corbalán, M.D.; A Madrid, J.; Morales, E.; Baraza, J.C.; Lee, Y.C.; Ordovas, J.M. CLOCK gene is implicated in weight reduction in obese patients participating in a dietary programme based on the Mediterranean diet. Int. J. Obes. 2010, 34, 516–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garaulet, M.; Esteban Tardido, A.; Lee, Y.C.; Smith, C.E.; Parnell, L.D.; Ordovás, J.M. SIRT1 and CLOCK 3111T> C combined genotype is associated with evening preference and weight loss resistance in a behavioral therapy treatment for obesity. Int. J. Obes. 2012, 36, 1436–1441. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Guimera, G.; Dashti, H.S.; Smith, C.E.; Sanchez-Carracedo, D.; Ordovas, J.M.; Garaulet, M. CLOCK 3111 T/C SNP in-teracts with emotional eating behavior for weight-loss in a Mediterranean population. PLoS ONE 2014, 9, e99152. [Google Scholar] [CrossRef] [Green Version]
- Garaulet, M.; Sánchez-Moreno, C.; Smith, C.E.; Lee, Y.-C.; Nicolás, F.; Ordovás, J.M. Ghrelin, Sleep Reduction and Evening Preference: Relationships to CLOCK 3111 T/C SNP and Weight Loss. PLoS ONE 2011, 6, e17435. [Google Scholar] [CrossRef] [PubMed]
- Franzago, M.; La Rovere, M.; Franchi, P.G.; Vitacolonna, E.; Stuppia, L. Epigenetics and human reproduction: The primary prevention of the noncommunicable diseases. Epigenomics 2019, 11, 1441–1460. [Google Scholar] [CrossRef] [PubMed]
- Franzago, M.; Pilenzi, L.; Di Rado, S.; Vitacolonna, E.; Stuppia, L. The epigenetic aging, obesity, and lifestyle. Front. Cell Dev. Biol. 2022, 10, 985274. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, P.; Buss, C.; Entringer, S.; Swanson, J. Developmental Origins of Health and Disease: Brief History of the Approach and Current Focus on Epigenetic Mechanisms. Semin. Reprod. Med. 2009, 27, 358–368. [Google Scholar] [CrossRef] [Green Version]
- Rigamonti, A.E.; Bollati, V.; Favero, C.; Albetti, B.; Caroli, D.; De Col, A.; Cella, S.G.; Sartorio, A. Changes in DNA Methylation of Clock Genes in Obese Adolescents after a Short-Term Body Weight Reduction Program: A Possible Metabolic and Endocrine Chrono-Resynchronization. Int. J. Environ. Res. Public Health 2022, 19, 15492. [Google Scholar] [CrossRef] [PubMed]
- Azzi, A.; Dallmann, R.; Casserly, A.; Rehrauer, H.; Patrignani, A.; Maier, B.; Kramer, A.; A Brown, S. Circadian behavior is light-reprogrammed by plastic DNA methylation. Nat. Neurosci. 2014, 17, 377–382. [Google Scholar] [CrossRef]
- Welberg, L. Circadian rhythms: Methylation mediates clock plasticity. Nat. Rev. Neurosci. 2014, 15, 206. [Google Scholar] [CrossRef]
- Bollati, V.; Baccarelli, A.; Sartori, S.; Tarantini, L.; Motta, V.; Rota, F.; Costa, G. Epigenetic Effects of Shiftwork on Blood DNA Methylation. Chronobiol. Int. 2010, 27, 1093–1104. [Google Scholar] [CrossRef] [Green Version]
- Manenschijn, L.; Koper, J.W.; Lamberts, S.W.; van Rossum, E.F. Evaluation of a method to measure long term cortisol levels. Steroids 2011, 76, 1032–1036. [Google Scholar] [CrossRef]
- Milagro, F.I.; Gómez-Abellán, P.; Campión, J.; Martínez, J.A.; Ordovás, J.M.; Garaulet, M. CLOCK, PER2 and BMAL1 DNA Methylation: Association with Obesity and Metabolic Syndrome Characteristics and Monounsaturated Fat Intake. Chronobiol. Int. 2012, 29, 1180–1194. [Google Scholar] [CrossRef]
- Samblas, M.; Milagro, F.I.; Gómez-Abellán, P.; Martínez, J.A.; Garaulet, M. Methylation on the Circadian Gene BMAL1 Is Associated with the Effects of a Weight Loss Intervention on Serum Lipid Levels. J. Biol. Rhythm. 2016, 31, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Samblas, M.; Milagro, F.I.; Martínez, A. DNA methylation markers in obesity, metabolic syndrome, and weight loss. Epigenetics 2019, 14, 421–444. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, O.; Samblas, M.; Milagro, F.I.; Riezu-Boj, J.I.; Crujeiras, A.B.; Martinez, J.A.; Project, M. Circadian gene meth-ylation profiles are associated with obesity, metabolic disturbances and carbohydrate intake. Chronobiol. Int. 2018, 35, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Etchegaray, J.-P.; Lee, C.; Wade, P.A.; Reppert, S.M. Rhythmic histone acetylation underlies transcription in the mammalian circadian clock. Nature 2002, 421, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Curtis, A.M.; Seo, S.-B.; Westgate, E.J.; Rudic, R.D.; Smyth, E.M.; Chakravarti, D.; FitzGerald, G.A.; McNamara, P. Histone Acetyltransferase-dependent Chromatin Remodeling and the Vascular Clock. J. Biol. Chem. 2004, 279, 7091–7097. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Witmer, P.D.; Lumayag, S.; Kovacs, B.; Valle, D. MicroRNA (miRNA) Transcriptome of Mouse Retina and Identification of a Sensory Organ-specific miRNA Cluster. J. Biol. Chem. 2007, 282, 25053–25066. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, C.; Aoyama, K.; Matsumura, N.; Kikuchi-Utsumi, K.; Watabe, M.; Nakaki, T. Rhythmic oscillations of the microRNA miR-96-5p play a neuroprotective role by indirectly regulating glutathione levels. Nat. Commun. 2014, 5, 3823. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Impli-cations for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [Green Version]
- Figueredo, D.S.; Gitai, D.L.; Andrade, T.G. Daily variations in the expression of miR-16 and miR-181a in human leukocytes. Blood Cells Mol. Dis. 2015, 54, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Shende, V.R.; Goldrick, M.M.; Ramani, S.; Earnest, D.J. Expression and rhythmic modulation of circulating microRNAs tar-geting the clock gene Bmal1 in mice. PLoS ONE 2011, 6, 22586. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Zhang, P.; Zhou, L.; Yin, B.; Pan, H.; Peng, X. Clock-controlled mir-142-3p can target its activator, Bmal1. BMC Mol. Biol. 2012, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Daimiel-Ruiz, L.; Klett-Mingo, M.; Konstantinidou, V.; Mico, V.; Aranda, J.F.; Garcia, B.; Martinez-Botas, J.; Davalos, A.; Fernandez-Hernando, C.; Ordovas, J.M. Dietary lipids modulate the expression of miR-107 a miRNA that regulates the cir-cadian system. Mol. Nutr. Food Res. 2015, 59, 552–565. [Google Scholar] [CrossRef] [Green Version]
- Zampetaki, A.; Willeit, P.; Drozdov, I.; Kiechl, S.; Mayr, M. Profiling of circulating microRNAs: From single biomarkers to re-wired networks. Cardiovasc. Res. 2011, 93, 555–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintanilha, B.J.; Reis, B.Z.; Duarte, G.B.S.; Cozzolino, S.M.F.; Rogero, M.M. Nutrimiromics: Role of microRNAs and Nutrition in Modulating Inflammation and Chronic Diseases. Nutrients 2017, 9, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenburg, J.L.; Sonnenburg, E.D. Vulnerability of the industrialized microbiota. Science 2019, 366, eaaw9255. [Google Scholar] [CrossRef] [Green Version]
- Bishehsari, F.; Keshavarzian, A. Microbes help to track time. Science 2019, 365, 1379–1380. [Google Scholar] [CrossRef]
- Shen, J.; Obin, M.S.; Zhao, L. The gut microbiota, obesity and insulin resistance. Mol. Asp. Med. 2013, 34, 39–58. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Cardona, F.; Tinahones, F.J.; Queipo-Ortuã±O, M.I. Impact of the gut microbiota on the development of obesity and type 2 diabetes mellitus. Front. Microbiol. 2014, 5, 190. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.L.; Davies, I.G.; Waraksa, W.; Khayyatzadeh, S.S.; Al-Asmakh, M.; Mazidi, M. The Impact of Microbial Com-position on Postprandial Glycaemia and Lipidaemia: A Systematic Review of Current Evidence. Nutrients 2021, 13, 3887. [Google Scholar] [CrossRef]
- Vitacolonna, E.; Masulli, M.; Palmisano, L.; Stuppia, L.; Franzago, M. Inositols, Probiotics, and Gestational Diabetes: Clinical and Epigenetic Aspects. Nutrients 2022, 14, 1543. [Google Scholar] [CrossRef]
- Nova, E.; Gómez-Martinez, S.; González-Soltero, R. The Influence of Dietary Factors on the Gut Microbiota. Microorganisms 2022, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.G.; Erbil, R.; Greve, J.W.; Buurman, W.A.; de Vos, W.M.; Rensen, S.S. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obesity 2013, 21, E607–E615. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [Green Version]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and Feeding Pattern Affect the Diurnal Dynamics of the Gut Microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom Control of Microbiota Diurnal Oscillations Promotes Metabolic Homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [Green Version]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510. [Google Scholar] [CrossRef] [Green Version]
- Paulose, J.K.; Wright, J.M.; Patel, A.G.; Cassone, V.M. Human gut bacteria are sensitive to melatonin and express en-dogenous circadian rhythmicity. PLoS ONE 2016, 11, e0146643. [Google Scholar] [CrossRef]
- Paulose, J.; Cassone, C.V.; Graniczkowska, K.B.; Cassone, V.M. Entrainment of the Circadian Clock of the Enteric Bacterium Klebsiella aerogenes by Temperature Cycles. Iscience 2019, 19, 1202–1213. [Google Scholar] [CrossRef]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [Green Version]
- Berry, S.E.; Valdes, A.M.; Drew, D.A.; Asnicar, F.; Mazidi, M.; Wolf, J.; Capdevila, J.; Hadjigeorgiou, G.; Davies, R.; Al Khatib, H.; et al. Human postprandial responses to food and potential for precision nutrition. Nat. Med. 2020, 26, 964–973. [Google Scholar] [CrossRef]
- Parkar, S.G.; Kalsbeek, A.; Cheeseman, J.F. Potential Role for the Gut Microbiota in Modulating Host Circadian Rhythms and Metabolic Health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef]
- Tuohy, K.M.; Conterno, L.; Gasperotti, M.; Viola, R. Up-regulating the Human Intestinal Microbiome Using Whole Plant Foods, Polyphenols, and/or Fiber. J. Agric. Food Chem. 2012, 60, 8776–8782. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, K.; Cheung, B.W.; Schröder, H. The possible roles of food-derived bioactive peptides in reducing the risk of car-diovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Potter, G.D.M.; Cade, J.; Grant, P.J.; Hardie, L.J. Nutrition and the circadian system. Br. J. Nutr. 2016, 116, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Sheward, W.J.; Maywood, E.S.; French, K.L.; Horn, J.M.; Hastings, M.H.; Seckl, J.R.; Holmes, M.C.; Harmar, A.J. Entrainment to Feeding but Not to Light: Circadian Phenotype of VPAC2 Receptor-Null Mice. J. Neurosci. 2007, 27, 4351–4358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwer, A.; van Raalte, D.H.; Diamant, M.; Rutters, F.; van Someren, E.J.; Snoek, F.J.; Beekman, A.T.; Bremmer, M.A. Light therapy for better mood and insulin sensitivity in patients with major depression and type 2 diabetes: A randomised, double-blind, parallel-arm trial. BMC Psychiatry 2015, 15, 169. [Google Scholar] [CrossRef] [Green Version]
- Brouwer, A.; van Raalte, D.H.; Nguyen, H.-T.; Rutters, F.; van de Ven, P.M.; Elders, P.J.; Moll, A.C.; Van Someren, E.J.; Snoek, F.J.; Beekman, A.T.; et al. Effects of Light Therapy on Mood and Insulin Sensitivity in Patients With Type 2 Diabetes and Depression: Results From a Randomized Placebo-Controlled Trial. Diabetes Care 2019, 42, 529–538. [Google Scholar] [CrossRef] [PubMed]


| Authors (Ref) | Sample Size | Study Design | Methylation Profiles in Target Genes | Evidence |
|---|---|---|---|---|
| Milagro et al., 2012 [101] | N = 20 Normal weight (BMI < 25 kg/m2), N = 20 overweight/obese (BMI = 29–35 kg/m2) N = 20 morbidly obese women (BMI > 40 kg/m2), aged 25 to 53 years | Cross-sectional study (28 weeks of treatment) | CLOCK BMAL1 PER2 | Association between methylation status of CpG sites located in CLOCK, BMAL1, and PER2 with obesity, MetS. The methylation status of different CpG sites in CLOCK and PER2 has been proposed as biomarkers of weight loss success. |
| Ramos-Lopez et al., 2018 [104] | N = 474 adults belonging to the MENA project | Cross-sectional study | RORA PRKAG2 PER3 BHLHE40 FBXL3 RORA CREB1 PRKAG2 PRKAG2 | Correlation between DNA methylation patterns at six circadian rhythm pathway genes with BMI. Correlation between methylation signatures at cg09578018 (RORA), cg24061580 (PRKAG2), cg01180628 (BHLHE40), and cg10059324 (PER3) with insulin resistance (p < 0.0001) and mean arterial blood pressure (p < 0.0001). Relevant correlations between methylation at cg09578018 (RORA) and cg01180628 (BHLHE40) with total energy and carbohydrate intakes (p < 0.05). |
| Samblas et al., 2016 [102] | N = 61 women (BMI = 28.6 ± 3.4 kg/m2; age: 42.2 ± 11.4 years) | Cross-sectional study Weight loss treatment (nutritional program based on a Mediterranean dietary pattern) | BMAL1 CLOCK NR1D1 | The energy-restricted intervention modified the methylation levels of different CpG sites in BMAL1 and NR1D1. The changes in BMAL1 methylation level with the intervention, positively correlated with the eveningness profile (p = 0.019). The baseline methylation at BMAL1 positively correlated with energy (p = 0.047) and carbohydrate (p = 0.017) intake and negatively correlated with the effect of the weight loss intervention on TC (p = 0.032) and low-density lipoprotein cholesterol (p = 0.005). Significant and positive correlations were found between changes in methylation levels in the CpG region of BMAL1 due to the intervention and changes in serum lipids (p < 0.05). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franzago, M.; Alessandrelli, E.; Notarangelo, S.; Stuppia, L.; Vitacolonna, E. Chrono-Nutrition: Circadian Rhythm and Personalized Nutrition. Int. J. Mol. Sci. 2023, 24, 2571. https://doi.org/10.3390/ijms24032571
Franzago M, Alessandrelli E, Notarangelo S, Stuppia L, Vitacolonna E. Chrono-Nutrition: Circadian Rhythm and Personalized Nutrition. International Journal of Molecular Sciences. 2023; 24(3):2571. https://doi.org/10.3390/ijms24032571
Chicago/Turabian StyleFranzago, Marica, Elisa Alessandrelli, Stefania Notarangelo, Liborio Stuppia, and Ester Vitacolonna. 2023. "Chrono-Nutrition: Circadian Rhythm and Personalized Nutrition" International Journal of Molecular Sciences 24, no. 3: 2571. https://doi.org/10.3390/ijms24032571
APA StyleFranzago, M., Alessandrelli, E., Notarangelo, S., Stuppia, L., & Vitacolonna, E. (2023). Chrono-Nutrition: Circadian Rhythm and Personalized Nutrition. International Journal of Molecular Sciences, 24(3), 2571. https://doi.org/10.3390/ijms24032571