
A Comprehensive Analysis of Cytokine Network in Centenarians

 ,
,  , , ,
, , , 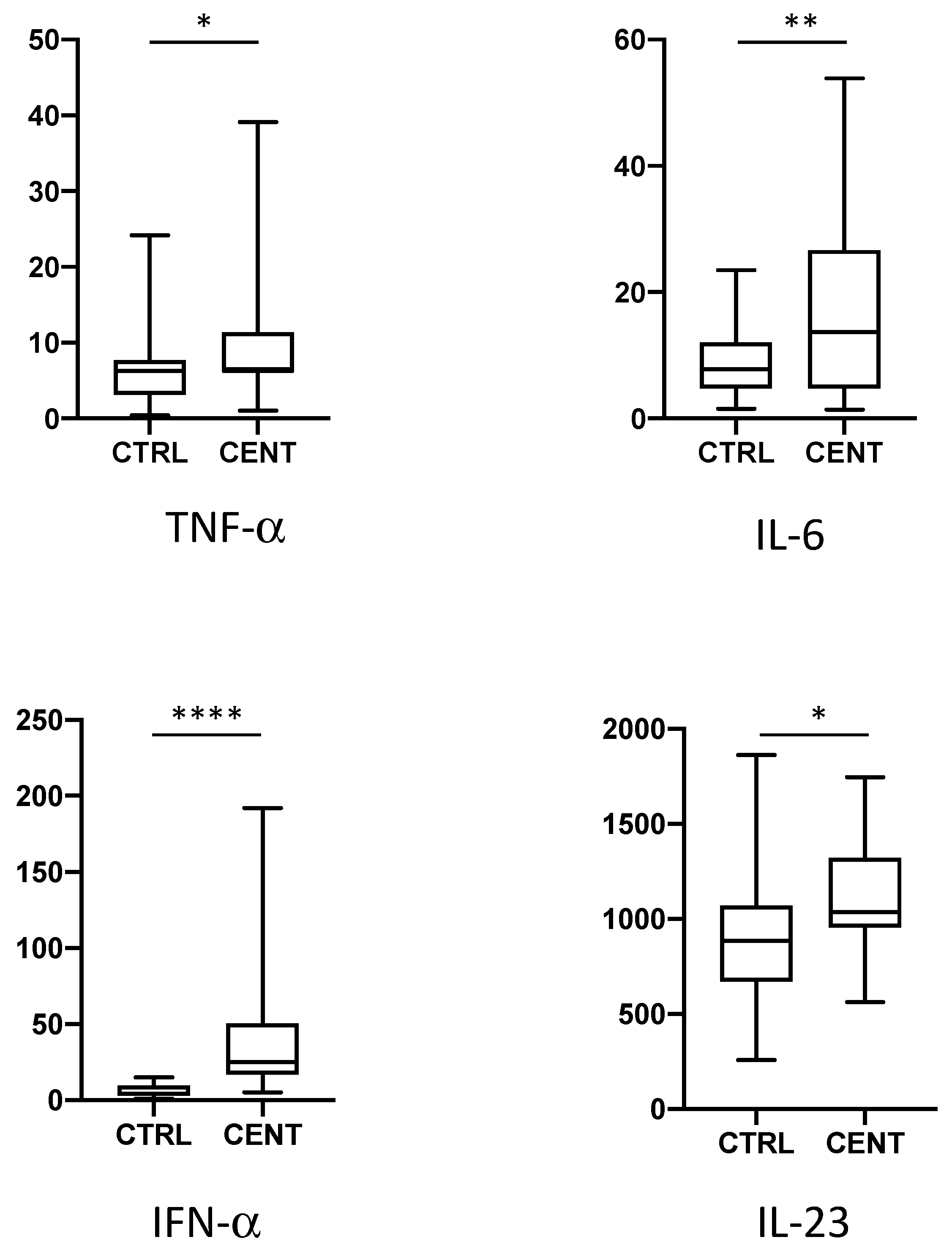{kind=link}
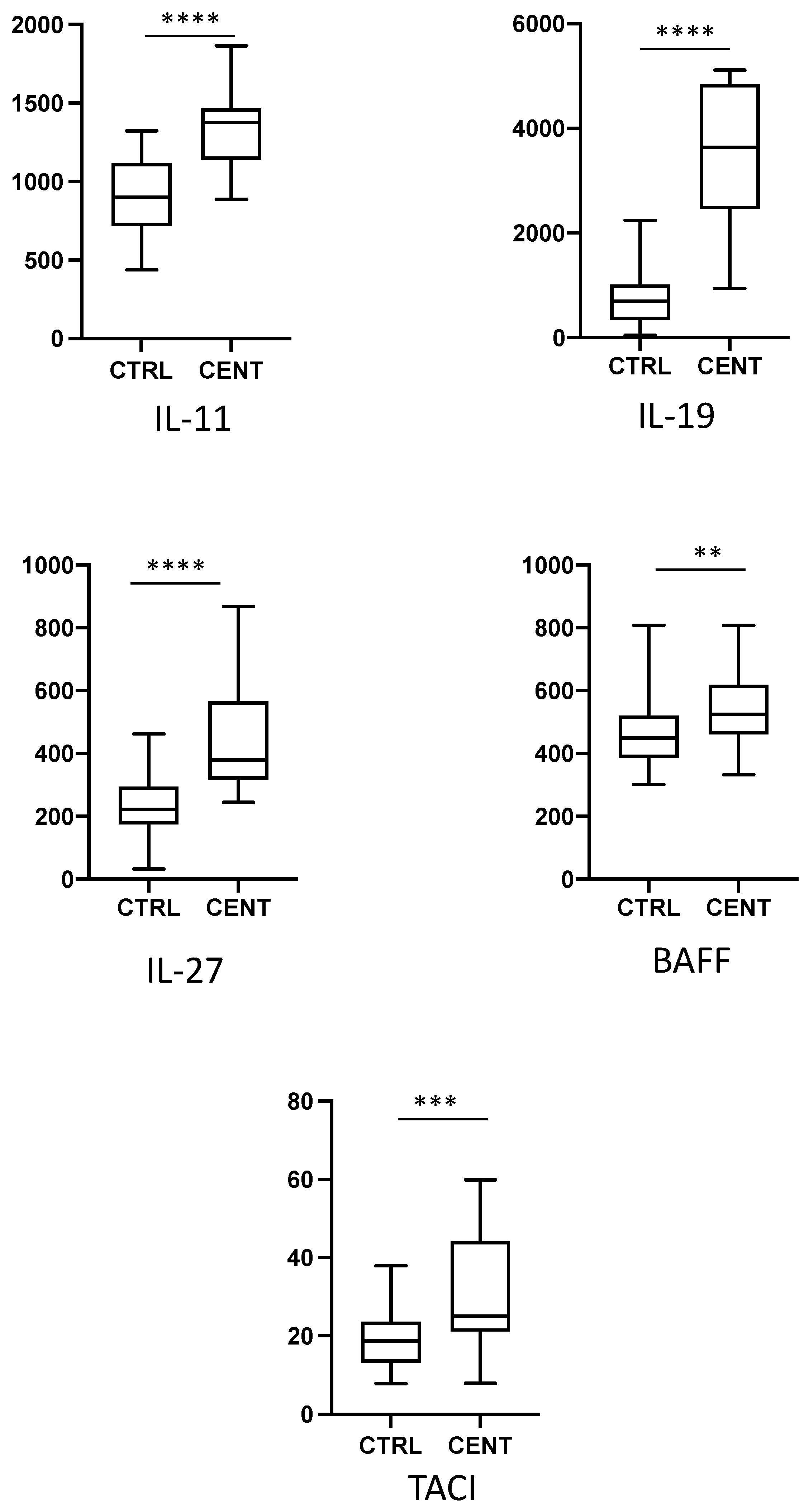{kind=link}
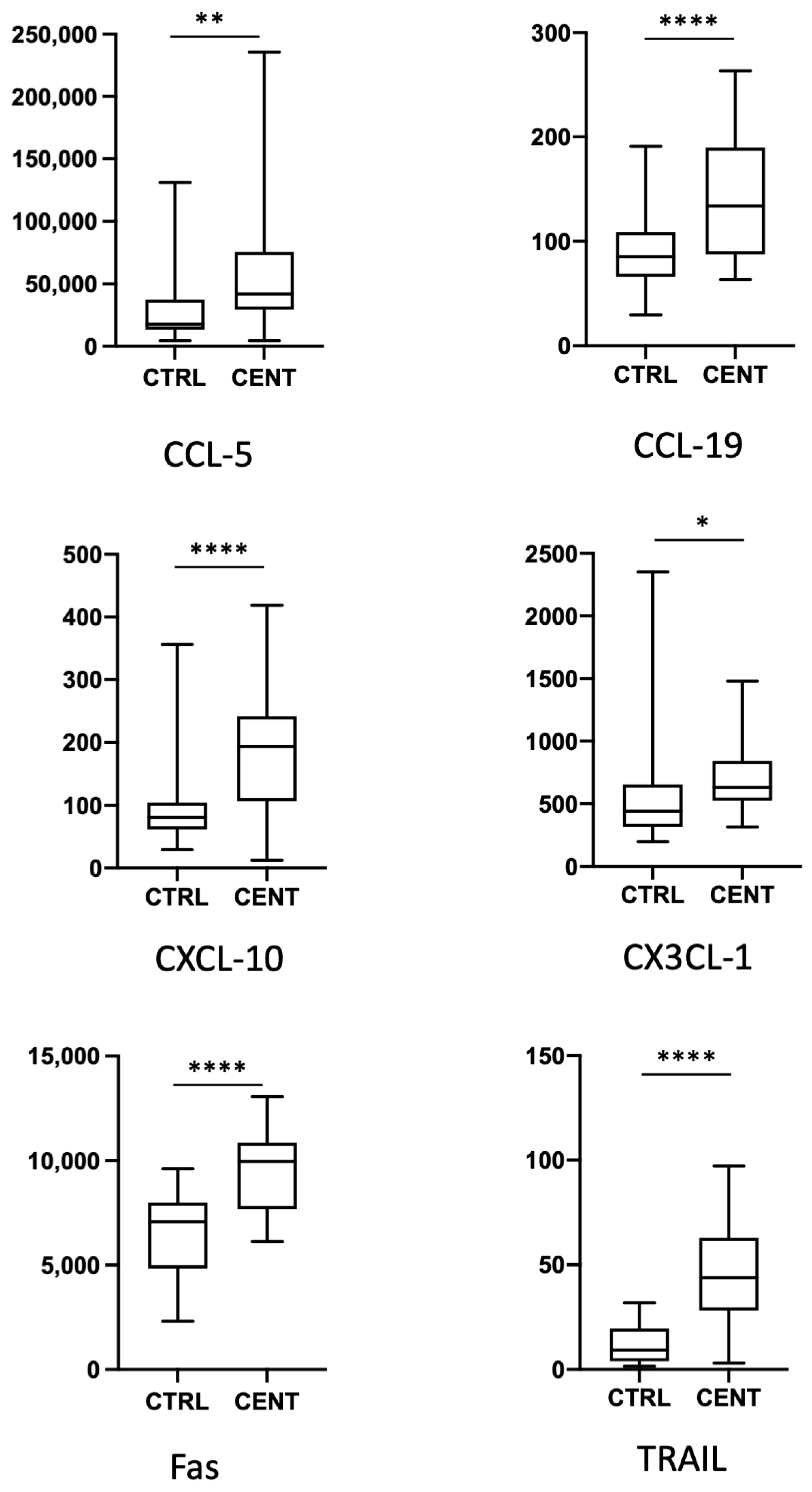{kind=link}
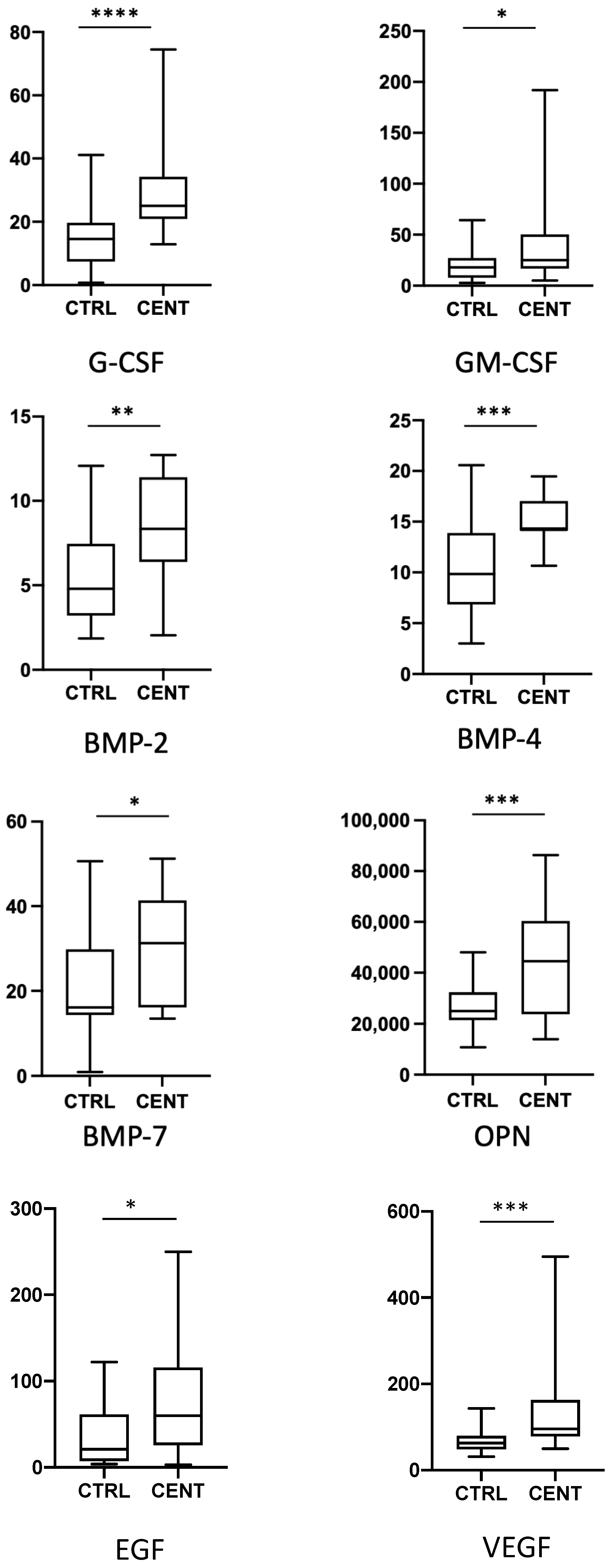{kind=link}
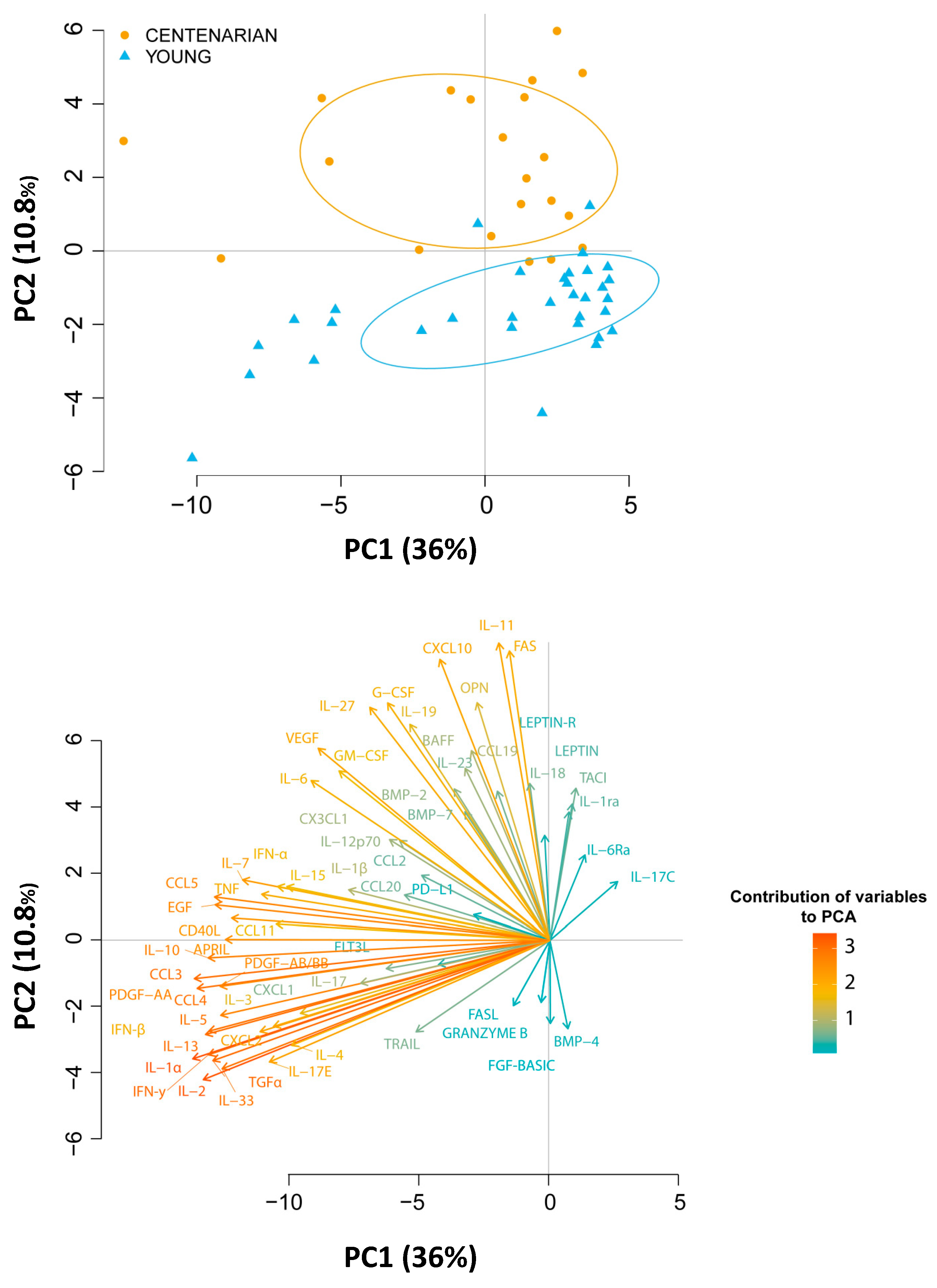{kind=link}
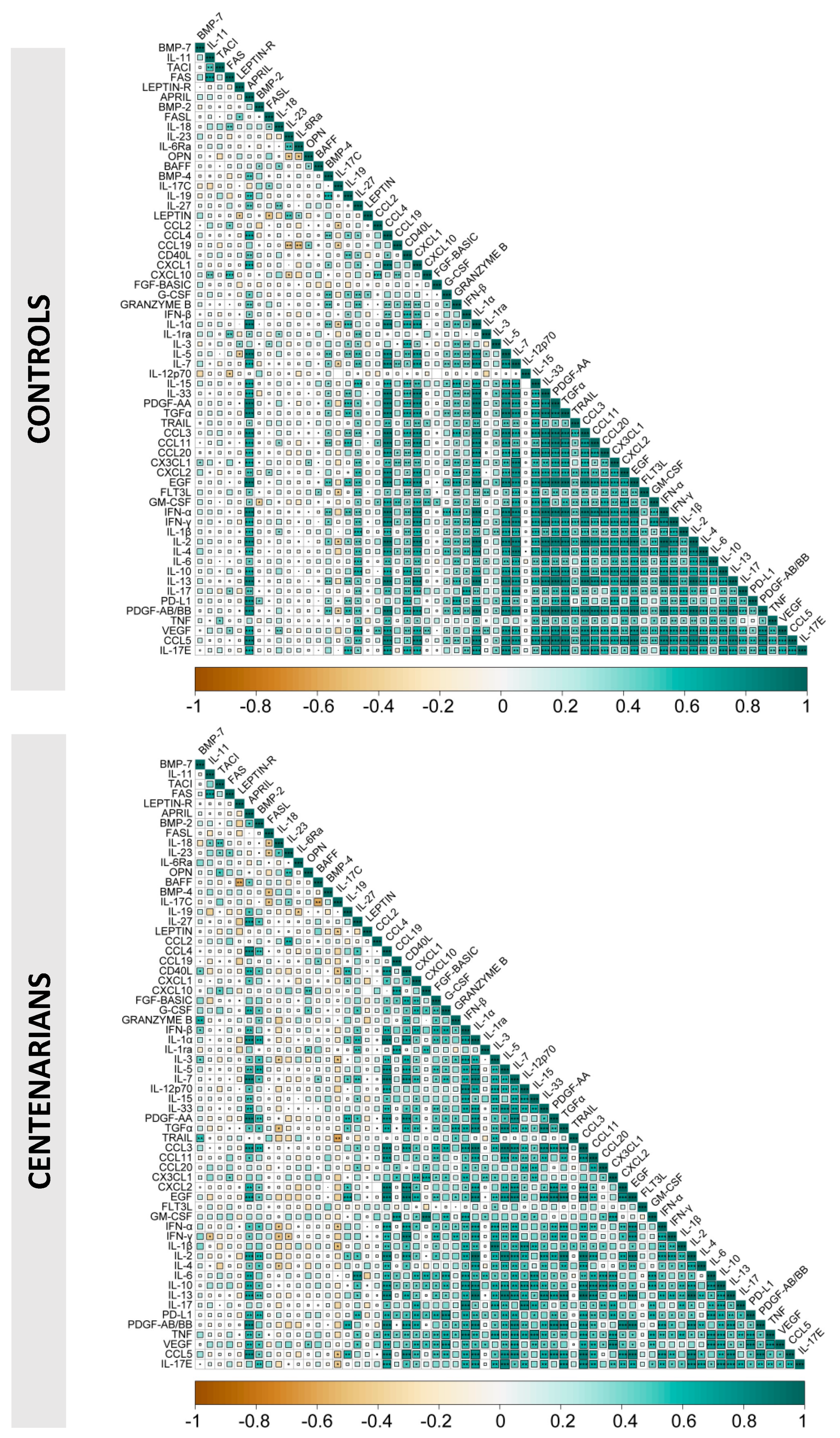{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Cytokine Quantification
4.3. Analysis of the Correlations among Plasma Cytokines in Young Donors and Centenarians
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Vasto, S.; Candore, G.; Balistreri, C.R.; Caruso, M.; Colonna-Romano, G.; Grimaldi, M.P.; Listi, F.; Nuzzo, D.; Lio, D.; Caruso, C. Inflammatory networks in ageing, age-related diseases and longevity. Mech. Ageing Dev. 2007, 128, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Valensin, S.; Bonafe, M.; Paolisso, G.; Yashin, A.I.; Monti, D.; De Benedictis, G. The network and the remodeling theories of aging: Historical background and new perspectives. Exp. Gerontol. 2000, 35, 879–896. [Google Scholar] [CrossRef]
- Caruso, C.; Passarino, G.; Puca, A.; Scapagnini, G. “Positive biology”: The centenarian lesson. Immun. Ageing I A 2012, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Monti, D.; Sansoni, P.; Cossarizza, A. The immunology of exceptional individuals: The lesson of centenarians. Immunol. Today 1995, 16, 12–16. [Google Scholar] [CrossRef]
- Caruso, C.; Puca, A.A. Special Issue “Centenarians-A Model to Study the Molecular Basis of Lifespan and Healthspan”. Int. J. Mol. Sci. 2021, 22, 2044. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2017, 8, 1960. [Google Scholar] [CrossRef]
- Pinti, M.; Nasi, M.; Lugli, E.; Gibellini, L.; Bertoncelli, L.; Roat, E.; De Biasi, S.; Mussini, C.; Cossarizza, A. T cell homeostasis in centenarians: From the thymus to the periphery. Curr. Pharm. Des. 2010, 16, 597–603. [Google Scholar] [CrossRef]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Pinti, M.; Appay, V.; Campisi, J.; Frasca, D.; Fulop, T.; Sauce, D.; Larbi, A.; Weinberger, B.; Cossarizza, A. Aging of the immune system: Focus on inflammation and vaccination. Eur. J. Immunol. 2016, 46, 2286–2301. [Google Scholar] [CrossRef]
- Palmeri, M.; Misiano, G.; Malaguarnera, M.; Forte, G.I.; Vaccarino, L.; Milano, S.; Scola, L.; Caruso, C.; Motta, M.; Maugeri, D.; et al. Cytokine serum profile in a group of Sicilian nonagenarians. J. Immunoass. Immunochem. 2012, 33, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Sansoni, P.; Vescovini, R.; Fagnoni, F.; Biasini, C.; Zanni, F.; Zanlari, L.; Telera, A.; Lucchini, G.; Passeri, G.; Monti, D.; et al. The immune system in extreme longevity. Exp. Gerontol. 2008, 43, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Michaud, M.; Balardy, L.; Moulis, G.; Gaudin, C.; Peyrot, C.; Vellas, B.; Cesari, M.; Nourhashemi, F. Proinflammatory cytokines, aging, and age-related diseases. J. Am. Med. Dir. Assoc. 2013, 14, 877–882. [Google Scholar] [CrossRef]
- Yende, S.; Tuomanen, E.I.; Wunderink, R.; Kanaya, A.; Newman, A.B.; Harris, T.; de Rekeneire, N.; Kritchevsky, S.B. Preinfection systemic inflammatory markers and risk of hospitalization due to pneumonia. Am. J. Respir. Crit. Care Med. 2005, 172, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Broman, N.; Rantasarkka, K.; Feuth, T.; Valtonen, M.; Waris, M.; Hohenthal, U.; Rintala, E.; Karlsson, A.; Marttila, H.; Peltola, V.; et al. IL-6 and other biomarkers as predictors of severity in COVID-19. Ann. Med. 2021, 53, 410–412. [Google Scholar] [CrossRef]
- Di Iorio, A.; Ferrucci, L.; Sparvieri, E.; Cherubini, A.; Volpato, S.; Corsi, A.; Bonafe, M.; Franceschi, C.; Abate, G.; Paganelli, R. Serum IL-1beta levels in health and disease: A population-based study. ‘The InCHIANTI study’. Cytokine 2003, 22, 198–205. [Google Scholar] [CrossRef]
- Pinti, M.; Cevenini, E.; Nasi, M.; De Biasi, S.; Salvioli, S.; Monti, D.; Benatti, S.; Gibellini, L.; Cotichini, R.; Stazi, M.A.; et al. Circulating mitochondrial DNA increases with age and is a familiar trait: Implications for “inflamm-aging”. Eur. J. Immunol. 2014, 44, 1552–1562. [Google Scholar] [CrossRef]
- Zanni, F.; Vescovini, R.; Biasini, C.; Fagnoni, F.; Zanlari, L.; Telera, A.; Di Pede, P.; Passeri, G.; Pedrazzoni, M.; Passeri, M.; et al. Marked increase with age of type 1 cytokines within memory and effector/cytotoxic CD8+ T cells in humans: A contribution to understand the relationship between inflammation and immunosenescence. Exp. Gerontol. 2003, 38, 981–987. [Google Scholar] [CrossRef]
- Bruunsgaard, H.; Andersen-Ranberg, K.; Hjelmborg, J.; Pedersen, B.K.; Jeune, B. Elevated levels of tumor necrosis factor alpha and mortality in centenarians. Am. J. Med. 2003, 115, 278–283. [Google Scholar] [CrossRef]
- Giovannini, S.; Onder, G.; Liperoti, R.; Russo, A.; Carter, C.; Capoluongo, E.; Pahor, M.; Bernabei, R.; Landi, F. Interleukin-6, C-Reactive Protein, and Tumor Necrosis Factor-Alpha as Predictors of Mortality in Frail, Community-Living Elderly Individuals. J. Am. Geriatr. Soc. 2011, 59, 1679–1685. [Google Scholar] [CrossRef]
- Van Den Biggelaar, A.H.; De Craen, A.J.; Gussekloo, J.; Huizinga, T.W.; Heijmans, B.T.; Frolich, M.; Kirkwood, T.B.; Westendorp, R.G. Inflammation underlying cardiovascular mortality is a late consequence of evolutionary programming. FASEB J. 2004, 18, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Salvioli, S.; Capri, M.; Bucci, L.; Lanni, C.; Racchi, M.; Uberti, D.; Memo, M.; Mari, D.; Govoni, S.; Franceschi, C. Why do centenarians escape or postpone cancer? The role of IGF-1, inflammation and p53. Cancer Immunol. Immunother. 2009, 58, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- McElhaney, J.E.; Zhou, X.; Talbot, H.K.; Soethout, E.; Bleackley, R.C.; Granville, D.J.; Pawelec, G. The unmet need in the elderly: How immunosenescence, CMV infection, co-morbidities and frailty are a challenge for the development of more effective influenza vaccines. Vaccine 2012, 30, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Nakachi, K.; Ito, Y.; Kikuchi, S.; Tamakoshi, A.; Yagyu, K.; Watanabe, Y.; Inaba, Y.; Kazuo, T.; Jacc Study, G. Variations in serum transforming growth factor-beta1 levels with gender, age and lifestyle factors of healthy Japanese adults. Dis. Mrk. 2009, 27, 23–28. [Google Scholar] [CrossRef]
- Forsey, R.J.; Thompson, J.M.; Ernerudh, J.; Hurst, T.L.; Strindhall, J.; Johansson, B.; Nilsson, B.O.; Wikby, A. Plasma cytokine profiles in elderly humans. Mech. Ageing Dev. 2003, 124, 487–493. [Google Scholar] [CrossRef]
- Pinti, M.; Troiano, L.; Nasi, M.; Bellodi, C.; Ferraresi, R.; Mussi, C.; Salvioli, G.; Cossarizza, A. Balanced regulation of mRNA production for Fas and Fas ligand in lymphocytes from centenarians: How the immune system starts its second century. Circulation 2004, 110, 3108–3114. [Google Scholar] [CrossRef]
- Pinti, M.; Troiano, L.; Nasi, M.; Moretti, L.; Monterastelli, E.; Mazzacani, A.; Mussi, C.; Ventura, P.; Olivieri, F.; Franceschi, C.; et al. Genetic polymorphisms of Fas (CD95) and FasL (CD178) in human longevity: Studies on centenarians. Cell Death Differ. 2002, 9, 431–438. [Google Scholar] [CrossRef]
- Aggarwal, S.; Gupta, S. Increased apoptosis of T cell subsets in aging humans: Altered expression of Fas (CD95), Fas ligand, Bcl-2, and Bax. J. Immunol. 1998, 160, 1627–1637. [Google Scholar] [CrossRef]
- Duvallet, E.; Semerano, L.; Assier, E.; Falgarone, G.; Boissier, M.C. Interleukin-23: A key cytokine in inflammatory diseases. Ann. Med. 2011, 43, 503–511. [Google Scholar] [CrossRef]
- Hamilton, J.A. GM-CSF in inflammation. J. Exp. Med. 2020, 217, e20190945. [Google Scholar] [CrossRef] [PubMed]
- Leigh, T.; Scalia, R.G.; Autieri, M.V. Resolution of inflammation in immune and nonimmune cells by interleukin-19. Am. J. Physiol. Cell Physiol. 2020, 319, C457–C464. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.M.; Vukmanovic-Stejic, M.; Dunne, P.J.; Birch, K.E.; Cook, J.E.; Jackson, S.E.; Salmon, M.; Rustin, M.H.; Akbar, A.N. Cytomegalovirus-specific CD4+ T cells in healthy carriers are continuously driven to replicative exhaustion. J. Immunol. 2005, 175, 8218–8225. [Google Scholar] [CrossRef] [PubMed]
- Karrer, U.; Mekker, A.; Wanke, K.; Tchang, V.; Haeberli, L. Cytomegalovirus and immune senescence: Culprit or innocent bystander? Exp. Gerontol. 2009, 44, 689–694. [Google Scholar] [CrossRef]
- Lied, G.A.; Berstad, A. Functional and clinical aspects of the B-cell-activating factor (BAFF): A narrative review. Scand. J. Immunol. 2011, 73, 1–7. [Google Scholar] [CrossRef]
- Hoffmann, F.S.; Kuhn, P.H.; Laurent, S.A.; Hauck, S.M.; Berer, K.; Wendlinger, S.A.; Krumbholz, M.; Khademi, M.; Olsson, T.; Dreyling, M.; et al. The immunoregulator soluble TACI is released by ADAM10 and reflects B cell activation in autoimmunity. J. Immunol. 2015, 194, 542–552. [Google Scholar] [CrossRef]
- Samy, E.; Wax, S.; Huard, B.; Hess, H.; Schneider, P. Targeting BAFF and APRIL in systemic lupus erythematosus and other antibody-associated diseases. Int. Rev. Immunol. 2017, 36, 3–19. [Google Scholar] [CrossRef]
- Ng, L.G.; Mackay, C.R.; Mackay, F. The BAFF/APRIL system: Life beyond B lymphocytes. Mol. Immunol. 2005, 42, 763–772. [Google Scholar] [CrossRef]
- Jin, R.; Kaneko, H.; Suzuki, H.; Arai, T.; Teramoto, T.; Fukao, T.; Kondo, N. Age-related changes in BAFF and APRIL profiles and upregulation of BAFF and APRIL expression in patients with primary antibody deficiency. Int. J. Mol. Med. 2008, 21, 233–238. [Google Scholar] [CrossRef]
- Paul, S.R.; Bennett, F.; Calvetti, J.A.; Kelleher, K.; Wood, C.R.; O’Hara, R.M., Jr.; Leary, A.C.; Sibley, B.; Clark, S.C.; Williams, D.A.; et al. Molecular cloning of a cDNA encoding interleukin 11, a stromal cell-derived lymphopoietic and hematopoietic cytokine. Proc. Natl. Acad. Sci. USA 1990, 87, 7512–7516. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, R.D.; Putoczki, T.L.; Griffin, M.D.W. Structural Understanding of Interleukin 6 Family Cytokine Signaling and Targeted Therapies: Focus on Interleukin 11. Front. Immunol. 2020, 11, 1424. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctot, K.L. A meta-analysis of cytokines in major depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.A.; Hargadon, B.; Shelley, M.; Parker, D.; Shaw, D.E.; Green, R.H.; Bradding, P.; Brightling, C.E.; Wardlaw, A.J.; Pavord, I.D. Evidence of a role of tumor necrosis factor alpha in refractory asthma. N. Engl. J. Med. 2006, 354, 697–708. [Google Scholar] [CrossRef]
- Ohta, H.; Wada, H.; Niwa, T.; Kirii, H.; Iwamoto, N.; Fujii, H.; Saito, K.; Sekikawa, K.; Seishima, M. Disruption of tumor necrosis factor-alpha gene diminishes the development of atherosclerosis in ApoE-deficient mice. Atherosclerosis 2005, 180, 11–17. [Google Scholar] [CrossRef]
- Reuter, M.A.; Pombo, C.; Betts, M.R. Cytokine production and dysregulation in HIV pathogenesis: Lessons for development of therapeutics and vaccines. Cytokine Growth Factor Rev. 2012, 23, 181–191. [Google Scholar] [CrossRef]
- Kondo, N.; Kuroda, T.; Kobayashi, D. Cytokine Networks in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 10922. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Liu, M.; Fu, Y.J.; Jiang, Y.J.; Zhang, Z.N. Alterations in levels of cytokine following treatment to predict outcome of sepsis: A meta-analysis. Cytokine 2023, 161, 156056. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, K.; Sakurai, M.; Yamamoto, Y.; Suzuki, E.; Tsuda, M.; Kataoka, T.R.; Hirata, M.; Nishie, M.; Nojiri, T.; Kumazoe, M.; et al. Alteration of specific cytokine expression patterns in patients with breast cancer. Sci. Rep. 2019, 9, 2924. [Google Scholar] [CrossRef] [PubMed]
- Lippitz, B.E.; Harris, R.A. Cytokine patterns in cancer patients: A review of the correlation between interleukin 6 and prognosis. Oncoimmunology 2016, 5, e1093722. [Google Scholar] [CrossRef]
- Wang, M.; Zhai, X.; Li, J.; Guan, J.; Xu, S.; Li, Y.; Zhu, H. The Role of Cytokines in Predicting the Response and Adverse Events Related to Immune Checkpoint Inhibitors. Front. Immunol. 2021, 12, 670391. [Google Scholar] [CrossRef]
- Ellsworth, S.G.; Rabatic, B.M.; Chen, J.; Zhao, J.; Campbell, J.; Wang, W.; Pi, W.; Stanton, P.; Matuszak, M.; Jolly, S.; et al. Principal component analysis identifies patterns of cytokine expression in non-small cell lung cancer patients undergoing definitive radiation therapy. PLoS ONE 2017, 12, e0183239. [Google Scholar] [CrossRef] [PubMed]
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Gozzi, L.; Iannone, A.; Lo Tartaro, D.; Mattioli, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef] [PubMed]
- De Biasi, S.; Tartaro, D.L.; Gibellini, L.; Paolini, A.; Quong, A.; Petes, C.; Awong, G.; Douglas, S.; Lin, D.; Nieto, J.; et al. Endogenous control of inflammation characterizes pregnant women with asymptomatic or paucisymptomatic SARS-CoV-2 infection. Nat. Commun. 2021, 12, 4677. [Google Scholar] [CrossRef] [PubMed]
- Lo Tartaro, D.; Neroni, A.; Paolini, A.; Borella, R.; Mattioli, M.; Fidanza, L.; Quong, A.; Petes, C.; Awong, G.; Douglas, S.; et al. Molecular and cellular immune features of aged patients with severe COVID-19 pneumonia. Commun. Biol. 2022, 5, 590. [Google Scholar] [CrossRef]
- De Biasi, S.; Lo Tartaro, D.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Mattioli, M.; Paolini, A.; Gozzi, L.; et al. Expansion of plasmablasts and loss of memory B cells in peripheral blood from COVID-19 patients with pneumonia. Eur. J. Immunol. 2020, 50, 1283–1294. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinti, M.; Gibellini, L.; Lo Tartaro, D.; De Biasi, S.; Nasi, M.; Borella, R.; Fidanza, L.; Neroni, A.; Troiano, L.; Franceschi, C.; et al. A Comprehensive Analysis of Cytokine Network in Centenarians. Int. J. Mol. Sci. 2023, 24, 2719. https://doi.org/10.3390/ijms24032719
Pinti M, Gibellini L, Lo Tartaro D, De Biasi S, Nasi M, Borella R, Fidanza L, Neroni A, Troiano L, Franceschi C, et al. A Comprehensive Analysis of Cytokine Network in Centenarians. International Journal of Molecular Sciences. 2023; 24(3):2719. https://doi.org/10.3390/ijms24032719
Chicago/Turabian StylePinti, Marcello, Lara Gibellini, Domenico Lo Tartaro, Sara De Biasi, Milena Nasi, Rebecca Borella, Lucia Fidanza, Anita Neroni, Leonarda Troiano, Claudio Franceschi, and et al. 2023. "A Comprehensive Analysis of Cytokine Network in Centenarians" International Journal of Molecular Sciences 24, no. 3: 2719. https://doi.org/10.3390/ijms24032719
APA StylePinti, M., Gibellini, L., Lo Tartaro, D., De Biasi, S., Nasi, M., Borella, R., Fidanza, L., Neroni, A., Troiano, L., Franceschi, C., & Cossarizza, A. (2023). A Comprehensive Analysis of Cytokine Network in Centenarians. International Journal of Molecular Sciences, 24(3), 2719. https://doi.org/10.3390/ijms24032719