
Emerging Role of SMILE in Liver Metabolism




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
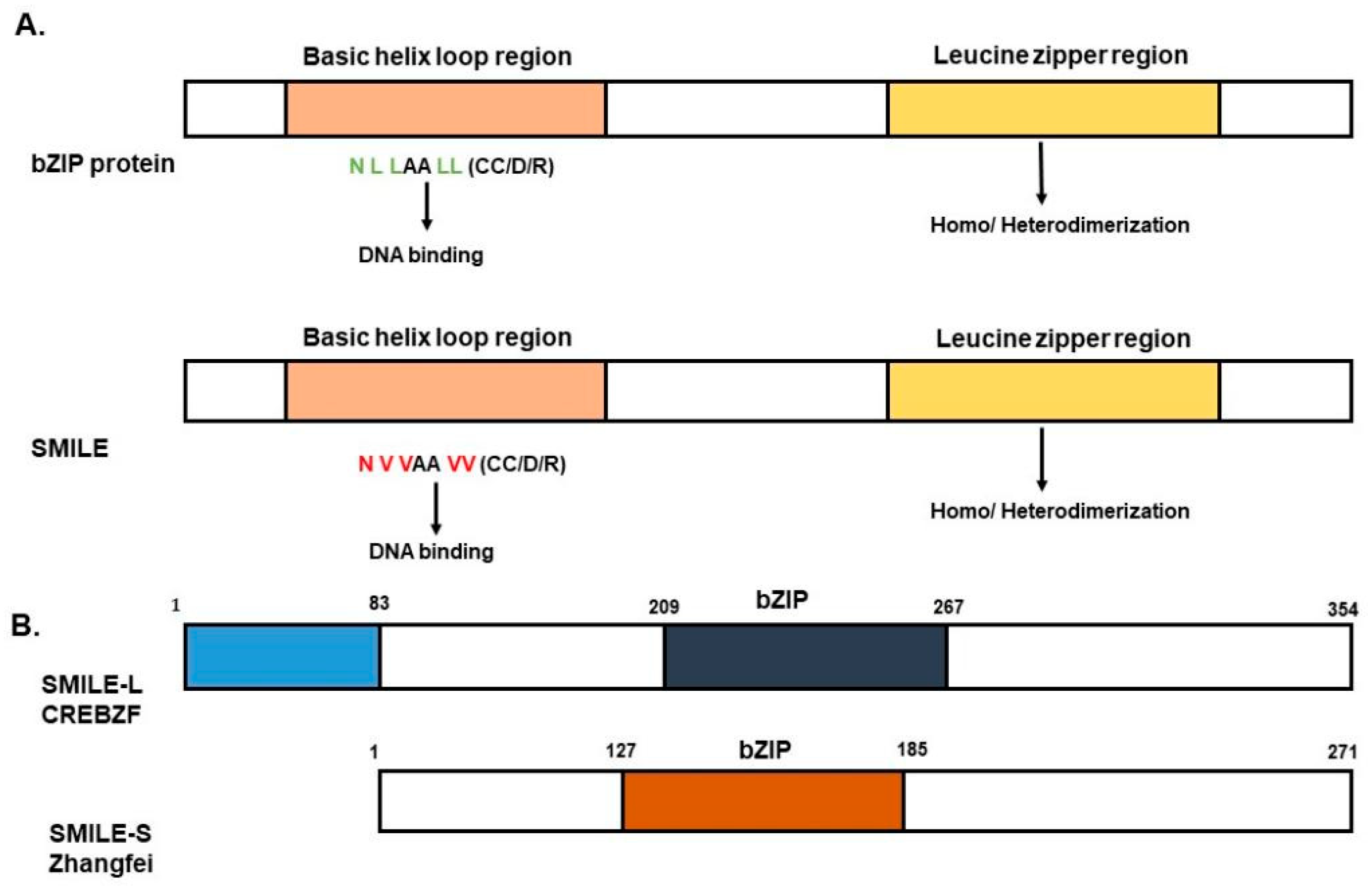
2. SMILE Background
3. Mode of Action of SMILE in Target Gene Regulation
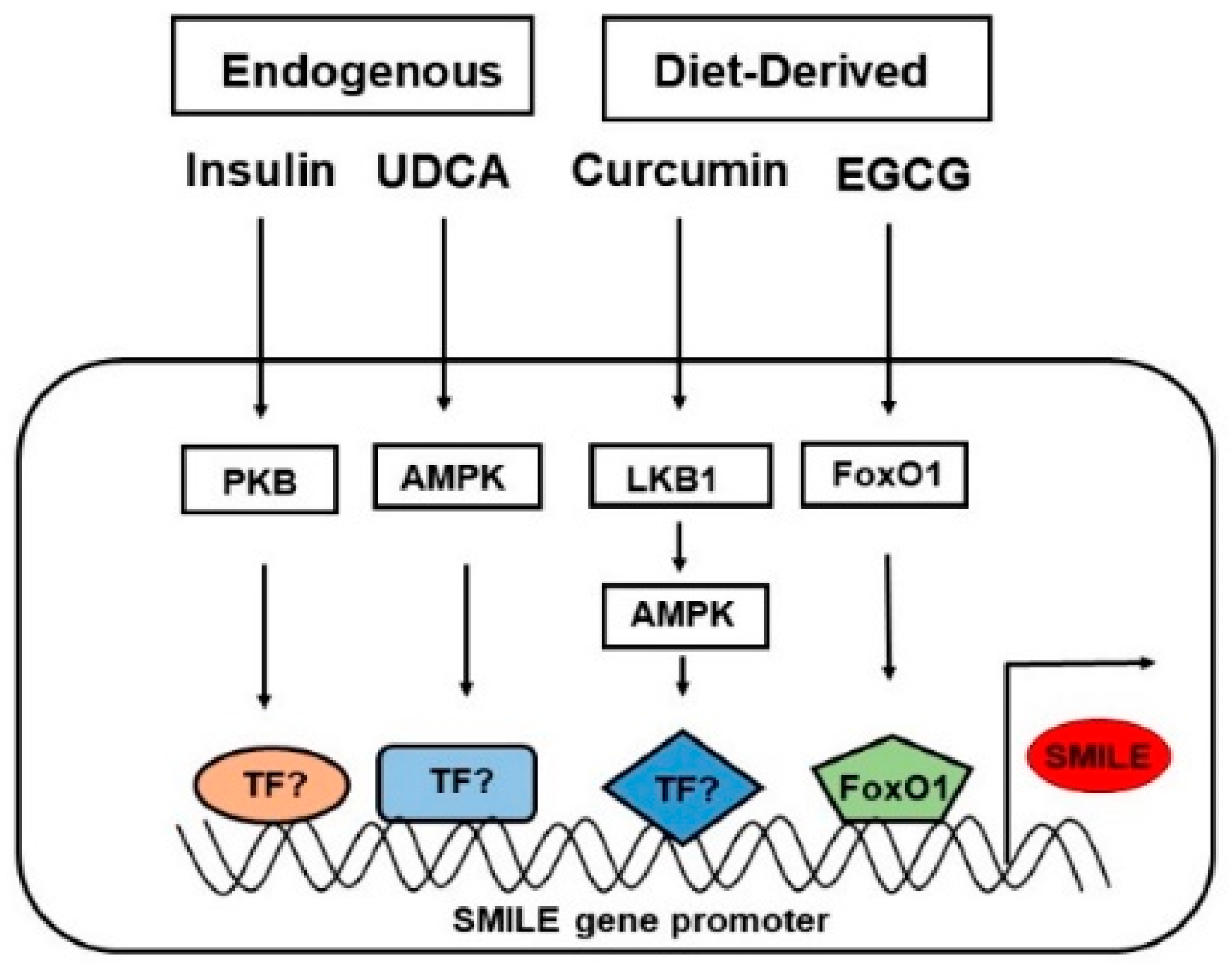
4. The Role of SMILE in Regulating Liver Metabolism via Various Signaling Pathways and Transcription Factors
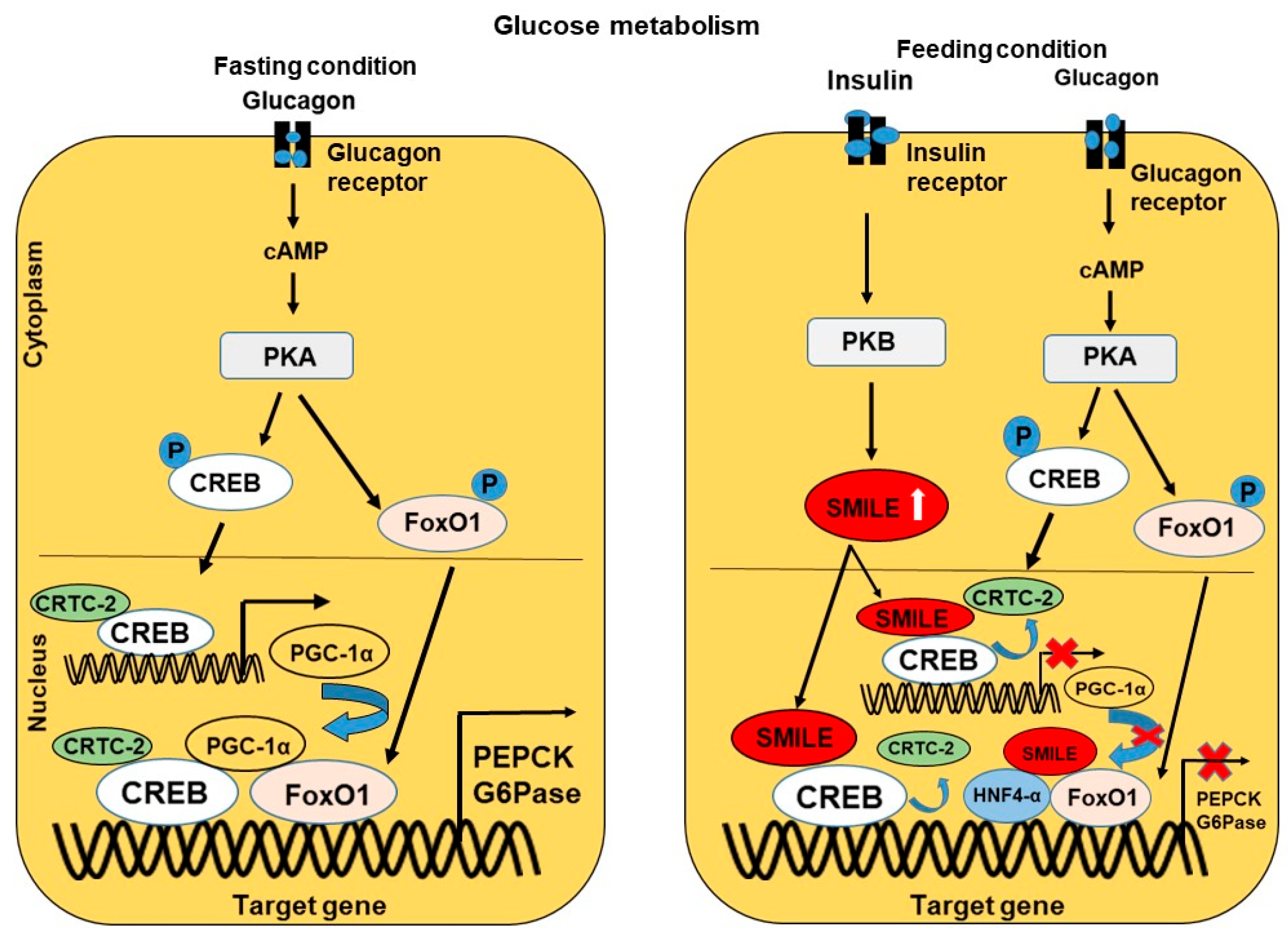
4.1. Glucose Metabolism and Insulin Resistance
4.2. Lipid Metabolism
4.3. Iron Metabolism and the Inflammatory Response
5. Non-Hepatic Functions of SMILE
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, R.; Misra, V. Zhangfei: A second cellular protein interacts with herpes simplex virus accessory factor HCF in a manner similar to Luman and VP16. Nucleic Acids Res. 2000, 28, 2446–2454. [Google Scholar] [CrossRef] [PubMed]
- Cockram, G.P.; Hogan, M.R.; Burnett, H.F.; Lu, R. Identification and characterization of the DNA-binding properties of a Zhangfei homologue in Japanese pufferfish, Takifugu rubripes. Biochem. Biophys. Res. Commun. 2006, 339, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Misra, V.; Rapin, N.; Akhova, O.; Bainbridge, M.; Korchinski, P. Zhangfei is a potent and specific inhibitor of the host cell factor-binding transcription factor Luman. J. Biol. Chem. 2005, 280, 15257–15266. [Google Scholar] [CrossRef] [PubMed]
- Hakoshima, T.; Fujii, Y.; Shimizu, T.; Toda, T.; Yanagida, M. Structural basis for the diversity of DNA recognition by bZIP transcription factors. Nat. Struct. Biol. 2000, 7, 889–893. [Google Scholar] [CrossRef]
- Ellenberger, T.E.; Brandl, C.J.; Struhl, K.; Harrison, S.C. The GCN4 basic region leucine zipper binds DNA as a dimer of uninterrupted alpha helices: Crystal structure of the protein-DNA complex. Cell 1992, 71, 1223–1237. [Google Scholar] [CrossRef]
- Vinson, C.R.; Sigler, P.; McKnight, S. Scissors-grip model for DNA recognition by a family of leucine zipper proteins. Science 1989, 246, 911–916. [Google Scholar] [CrossRef]
- Hurst, H.C. Transcription factors 1: bZIP proteins. Protein Profile 1995, 2, 101–168. [Google Scholar]
- Vinson, C.; Myakishev, M.; Acharya, A.; Mir, A.A.; Moll, J.R.; Bonovich, M. Classification of human B-ZIP proteins based on dimerization properties. Mol. Cell Biol. 2002, 22, 6321–6335. [Google Scholar] [CrossRef]
- Landschulz, W.H.; Johnson, P.; McKnight, S. The leucine zipper: A hypothetical structure common to a new class of DNA binding proteins. Science 1988, 240, 1759–1764. [Google Scholar] [CrossRef]
- Ron, D.; Habener, J. CHOP, a novel developmentally regulated nuclear protein that dimerizes with transcription factors C/EBP and LAP and functions as a dominant-negative inhibitor of gene transcription. Genes Dev. 1992, 6, 439–453. [Google Scholar] [CrossRef]
- Xie, Y.B.; Nedumaran, B.; Choi, H. Molecular characterization of SMILE as a novel corepressor of nuclear receptors. Nucleic Acids Res. 2009, 37, 4100–4115. [Google Scholar] [CrossRef] [Green Version]
- Sever, R.; Glass, C. Signaling by nuclear receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef]
- Frigo, D.E.; Bondesson, M.; Williams, C. Nuclear receptors: From molecular mechanisms to therapeutics. Essays Biochem. 2021, 65, 847–856. [Google Scholar] [CrossRef]
- Jenster, G. Coactivators and corepressors as mediators of nuclear receptor function: An update. Mol. Cell Endocrinol. 1998, 143, 1–7. [Google Scholar] [CrossRef]
- Xie, Y.B.; Park, J.H.; Kim, D.K.; Hwang, J.H.; Oh, S.; Park, S.B.; Shong, M.; Lee, I.K.; Choi, H.S. Transcriptional corepressor SMILE recruits SIRT1 to inhibit nuclear receptor estrogen receptor-related receptor gamma transactivation. J. Biol. Chem. 2009, 284, 28762–28774. [Google Scholar] [CrossRef]
- Yang, S.; Park, J.-S.; Hwang, S.-H.; Cho, K.-H.; Na, H.S.; Choi, J.; Jhun, J.; Kim, S.-J.; Lee, B.-I.; Park, S.-H.; et al. Metformin-Inducible Small Heterodimer Partner Interacting Leucine Zipper Protein Ameliorates Intestinal Inflammation. Front. Immunol. 2021, 12, 652709. [Google Scholar] [CrossRef]
- Lee, J.-M.; Seo, W.-Y.; Han, H.-S.; Oh, K.-J.; Lee, Y.-S.; Kim, D.-K.; Choi, S.; Choi, B.H.; Harris, R.A.; Lee, C.-H.; et al. Insulin-Inducible SMILE Inhibits Hepatic Gluconeogenesis. Diabetes 2016, 65, 62–73. [Google Scholar] [CrossRef]
- Lee, E.-J.; Kwon, J.-E.; Park, M.-J.; Jung, K.-A.; Kim, D.-S.; Kim, E.-K.; Lee, S.H.; Choi, J.Y.; Park, S.-H.; Cho, M.-L. Ursodeoxycholic acid attenuates experimental autoimmune arthritis by targeting Th17 and inducing pAMPK and transcriptional corepressor SMILE. Immunol. Lett. 2017, 188, 1–8. [Google Scholar] [CrossRef]
- Sandoval, V.; Sanz-Lamora, H.; Marrero, P.; Relat, J.; Haro, D. Lyophilized Maqui (Aristotelia chilensis) Berry Administration Suppresses High-Fat Diet-Induced Liver Lipogenesis through the Induction of the Nuclear Corepressor SMILE. Antioxidants 2021, 10, 637. [Google Scholar] [CrossRef]
- Lin, P.; Chen, F.; Wang, N.; Wang, X.; Li, X.; Zhou, J.; Jin, Y.; Wang, A. CREBZF expression and hormonal regulation in the mouse uterus. Reprod. Biol. Endocrinol. 2013, 11, 110. [Google Scholar] [CrossRef]
- López-Mateo, I.; Villaronga, M.A.; Llanos, S.; Belandia, B. The transcription factor CREBZF is a novel positive regulator of p53. Cell Cycle 2012, 11, 3887–3895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Qi, Z.; Yin, H.; Yang, G. Interaction between p53 and Ras signaling controls cisplatin resistance via HDAC4- and HIF-1α-mediated regulation of apoptosis and autophagy. Theranostics 2019, 9, 1096–1114. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wen, X.; Lin, P.; Chen, H.; Wang, A.; Jin, Y. Activation of CREBZF Increases Cell Apoptosis in Mouse Ovarian Granulosa Cells by Regulating the ERK1/2 and mTOR Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 3517. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-N.; Chang, H.-Y.; Wang, C.C.N.; Chu, F.-Y.; Shen, H.-Y.; Chen, C.-J.; Lim, Y.-P. Oleanolic Acid Inhibits Liver X Receptor Alpha and Pregnane X Receptor to Attenuate Ligand-Induced Lipogenesis. J. Agric. Food Chem. 2018, 66, 10964–10976. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M.; Gang, G.-T.; Kim, D.-K.; Kim, Y.D.; Koo, S.-H.; Lee, C.-H.; Choi, H.-S. Ursodeoxycholic acid inhibits liver X receptor α-mediated hepatic lipogenesis via induction of the nuclear corepressor SMILE. J. Biol. Chem. 2014, 289, 1079–1091. [Google Scholar] [CrossRef]
- Lee, K.-M.; Seo, Y.J.; Kim, M.-K.; Seo, H.-A.; Jeong, J.-Y.; Choi, H.-S.; Lee, I.-K.; Park, K.-G. Mediation of glucolipotoxicity in INS-1 rat insulinoma cells by small heterodimer partner interacting leucine zipper protein (SMILE). Biochem. Biophys. Res. Commun. 2012, 419, 768–773. [Google Scholar] [CrossRef]
- Chen, F.; Lin, P.F.; Li, X.; Sun, J.; Zhang, Z.; Du, E.; Wang, A.; Jin, Y.P. Construction and expression of lentiviral vectors encoding recombinant mouse CREBZF in NIH 3T3 cells. Plasmid 2014, 76, 24–31. [Google Scholar] [CrossRef]
- Xie, Y.-B.; Lee, O.-H.; Nedumaran, B.; Seong, H.-A.; Lee, K.-M.; Ha, H.; Lee, I.-K.; Yun, Y.; Choi, H.-S. SMILE, a new orphan nuclear receptor SHP-interacting protein, regulates SHP-repressed estrogen receptor transactivation. Biochem. J. 2008, 416, 463–473. [Google Scholar] [CrossRef]
- Hogan, M.R.; Cockram, G.; Lu, R. Cooperative interaction of Zhangfei and ATF4 in transactivation of the cyclic AMP response element. FEBS Lett. 2006, 580, 58–62. [Google Scholar] [CrossRef]
- Kim, M.Y.; Ahn, Y. SMILE Is an Insulin-Inducible Transcriptional Corepressor of Hepatic Gluconeogenic Gene Programs. Diabetes 2016, 65, 14–15. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Song, C.-H.; Xie, Y.-B.; Jung, C.; Choi, H.-S.; Lee, K. SMILE upregulated by metformin inhibits the function of androgen receptor in prostate cancer cells. Cancer Lett. 2014, 354, 390–397. [Google Scholar] [CrossRef]
- Mirza, A.Z.; Althagafi; Shamshad, H. Role of PPAR receptor in different diseases and their ligands: Physiological importance and clinical implications. Eur. J. Med. Chem. 2019, 166, 502–513. [Google Scholar] [CrossRef]
- Mansour, M. The roles of peroxisome proliferator-activated receptors in the metabolic syndrome. Prog. Mol. Biol. Transl. Sci. 2014, 121, 217–266. [Google Scholar]
- Kota, B.P.; Huang, T.; Roufogalis, B. An overview on biological mechanisms of PPARs. Pharmacol. Res. 2005, 51, 85–94. [Google Scholar] [CrossRef]
- Kersten, S.; Stienstra, R. The role and regulation of the peroxisome proliferator activated receptor alpha in human liver. Biochimie 2017, 136, 75–84. [Google Scholar] [CrossRef]
- Jang, H.; Kim, H.-J.; Kim, D.-H.; Park, J.-K.; Sun, W.-S.; Hwang, S.; Oh, K.-B.; Jang, W.-G.; Lee, J.-W. Small heterodimer partner-interacting leucine zipper protein inhibits adipogenesis by regulating peroxisome proliferator-activated receptor γ activity. Life Sci. 2015, 132, 49–54. [Google Scholar] [CrossRef]
- Wang, Y.; Nakajima, T.; Gonzalez, F.J.; Tanaka, N. PPARs as Metabolic Regulators in the Liver: Lessons from Liver-Specific PPAR-Null Mice. Int. J. Mol. Sci. 2020, 21, 2061. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, K.-S.; Lim, D.; Yang, D.J.; Park, J.-I.; Kim, K.W.; Jeong, J.-H.; Choi, H.-S.; Kim, D.-K. Epigallocatechin-3-Gallate (EGCG)-Inducible SMILE Inhibits STAT3-Mediated Hepcidin Gene Expression. Antioxidants 2020, 9, 514. [Google Scholar] [CrossRef]
- Kim, Y.J.; Park, W.R.; Choi, B.; Choi, H.S.; Kim, D.K. Epigallocatechin-3-Gallate Suppresses BMP-6-Mediated SMAD1/5/8 Transactivation of Hepcidin Gene by Inducing SMILE in Hepatocytes. Antioxidants 2021, 10, 1590. [Google Scholar] [CrossRef]
- Son, H.E.; Kim, E.; Jang, W. Curcumin induces osteoblast differentiation through mild-endoplasmic reticulum stress-mediated such as BMP2 on osteoblast cells. Life Sci. 2018, 193, 34–39. [Google Scholar] [CrossRef]
- Bogan, A.A.; Dallas-Yang, Q.; Ruse, M.D.; Maeda, Y.; Jiang, G.; Nepomuceno, L.; Scanlan, T.S.; Cohen, F.E.; Sladek, F.M. Analysis of protein dimerization and ligand binding of orphan receptor HNF4alpha. J. Mol. Biol. 2000, 302, 831–851. [Google Scholar] [CrossRef] [PubMed]
- Sharabi, K.; Lin, H.; Tavares, C.D.; Dominy, J.E.; Camporez, J.P.; Perry, R.J.; Schilling, R.; Rines, A.K.; Lee, J.; Hickey, M.; et al. Selective Chemical Inhibition of PGC-1α Gluconeogenic Activity Ameliorates Type 2 Diabetes. Cell 2017, 169, 148–160.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, T.; Fairall, L.; Muskett, F.W.; Milano, C.P.; Watson, P.J.; Arnaudo, N.; Saleh, A.; Millard, C.J.; El-Mezgueldi, M.; Martino, F.; et al. Structural and functional characterization of a cell cycle associated HDAC1/2 complex reveals the structural basis for complex assembly and nucleosome targeting. Nucleic Acids Res. 2015, 43, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Han, H.-S.; Kang, G.; Kim, J.S.; Choi, B.H.; Koo, S.-H. Regulation of glucose metabolism from a liver-centric perspective. Exp. Mol. Med. 2016, 48, e218. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Pérez-Felpete, N.; Fernández-Fernández, C.; Donapetry-García, C.; Pazos-García, C. Liver glucose metabolism in humans. Biosci. Rep. 2016, 36, e00416. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Han, H.S.; Kwon, Y.; Koo, S. Role of CRTC2 in Metabolic Homeostasis: Key Regulator of Whole-Body Energy Metabolism? Diabetes Metab. J. 2020, 44, 498–508. [Google Scholar] [CrossRef]
- Wu, H.; Deng, X.; Shi, Y.; Su, Y.; Wei, J.; Duan, H. PGC-1α, glucose metabolism and type 2 diabetes mellitus. J. Endocrinol. 2016, 229, R99–R115. [Google Scholar] [CrossRef]
- Lee, J.-M.; Seo, W.-Y.; Song, K.-H.; Chanda, D.; Kim, Y.D.; Kim, D.-K.; Lee, M.-W.; Ryu, D.; Noh, J.-R.; Lee, C.-H.; et al. AMPK-dependent repression of hepatic gluconeogenesis via disruption of CREB.CRTC2 complex by orphan nuclear receptor small heterodimer partner. J. Biol. Chem. 2010, 285, 32182–32191. [Google Scholar] [CrossRef]
- Wang, Y.; Inoue, H.; Ravnskjaer, K.; Viste, K.; Miller, N.; Liu, Y.; Hedrick, S.; Vera, L.; Montminy, M. Targeted disruption of the CREB coactivator Crtc2 increases insulin sensitivity. Proc. Natl. Acad. Sci. USA 2010, 107, 3087–3092. [Google Scholar] [CrossRef]
- Herzig, S.; Long, F.; Jhala, U.S.; Hedrick, S.; Quinn, R.; Bauer, A.; Rudolph, D.; Schutz, G.; Yoon, C.; Puigserver, P.; et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature 2001, 413, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, I.; Hager, G. The Three Ds of Transcription Activation by Glucagon: Direct, Delayed, and Dynamic. Endocrinology 2018, 159, 206–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puigserver, P.; Rhee, J.; Donovan, J.; Walkey, C.J.; Yoon, J.C.; Oriente, F.; Kitamura, Y.; Altomonte, J.; Dong, H.; Accili, D.; et al. Insulin-regulated hepatic gluconeogenesis through FOXO1-PGC-1alpha interaction. Nature 2003, 423, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Soyal, S.; Krempler, F.; Oberkofler, H.; Patsch, W. PGC-1alpha: A potent transcriptional cofactor involved in the pathogenesis of type 2 diabetes. Diabetologia 2006, 49, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Hatting, M.; Tavares, C.D.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. N. Y. Acad. Sci. 2018, 1411, 21–35. [Google Scholar] [CrossRef]
- Dentin, R.; Liu, Y.; Koo, S.-H.; Hedrick, S.; Vargas, T.; Heredia, J.; Yates, J.; Montminy, M. Insulin modulates gluconeogenesis by inhibition of the coactivator TORC2. Nature 2007, 449, 366–369. [Google Scholar] [CrossRef]
- He, L.; Sabet, A.; Djedjos, S.; Miller, R.; Sun, X.; Hussain, M.A.; Radovick, S.; Wondisford, F.E. Metformin and insulin suppress hepatic gluconeogenesis through phosphorylation of CREB binding protein. Cell 2009, 137, 635–646. [Google Scholar] [CrossRef]
- Lee, J.-M.; Han, H.-S.; Jung, Y.S.; Harris, R.A.; Koo, S.-H.; Choi, H.-S. The SMILE transcriptional corepressor inhibits cAMP response element-binding protein (CREB)-mediated transactivation of gluconeogenic genes. J. Biol. Chem. 2018, 293, 13125–13133. [Google Scholar] [CrossRef]
- Schultz, J.R.; Tu, H.; Luk, A.; Repa, J.J.; Medina, J.C.; Li, L.; Schwendner, S.; Wang, S.; Thoolen, M.; Mangelsdorf, D.J.; et al. Role of LXRs in control of lipogenesis. Genes Dev. 2000, 14, 2831–2838. [Google Scholar] [CrossRef]
- Bertolio, R.; Napoletano, F.; Mano, M.; Maurer-Stroh, S.; Fantuz, M.; Zannini, A.; Bicciato, S.; Sorrentino, G.; Del Sal, G. Sterol regulatory element binding protein 1 couples mechanical cues and lipid metabolism. Nat. Commun. 2019, 10, 1326. [Google Scholar] [CrossRef]
- Deng, Q.; Li, X.; Fu, S.; Yin, L.; Zhang, Y.; Wang, T.; Wang, J.; Liu, L.; Yuan, X.; Sun, G.; et al. SREBP-1c gene silencing can decrease lipid deposits in bovine hepatocytes cultured in vitro. Cell Physiol. Biochem. 2014, 33, 1568–1578. [Google Scholar] [CrossRef]
- Huss, J.M.; Torra, I.P.; Staels, B.; Giguère, V.; Kelly, D.P. Estrogen-related receptor alpha directs peroxisome proliferator-activated receptor alpha signaling in the transcriptional control of energy metabolism in cardiac and skeletal muscle. Mol. Cell Biol. 2004, 24, 9079–9091. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.K.; Kim, J.R.; Koh, M.; Kim, Y.D.; Lee, J.M.; Chanda, D.; Park, S.B.; Min, J.J.; Lee, C.H.; Park, T.S.; et al. Estrogen-related receptor γ (ERRγ) is a novel transcriptional regulator of phosphatidic acid phosphatase, LIPIN1, and inhibits hepatic insulin signaling. J. Biol. Chem. 2011, 286, 38035–38042. [Google Scholar] [CrossRef]
- Chen, W.; Chen, G.; Head, D.L.; Mangelsdorf, D.J.; Russell, D.W. Enzymatic reduction of oxysterols impairs LXR signaling in cultured cells and the livers of mice. Cell Metab. 2007, 5, 73–79. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Kliewer, S.A.; Moore, L.B.; Smith-Oliver, T.A.; Oliver, B.B.; Su, J.-L.; Sundseth, S.S.; Winegar, D.A.; Blanchard, D.E.; Spencer, T.A.; et al. Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway. J. Biol. Chem. 1997, 272, 3137–3140. [Google Scholar] [CrossRef]
- Lin, Y.N.; Wang, C.C.; Chang, H.Y.; Chu, F.Y.; Hsu, Y.A.; Cheng, W.K.; Ma, W.C.; Chen, C.J.; Wan, L.; Lim, Y.P. Ursolic Acid, a Novel Liver X Receptor α (LXRα) Antagonist Inhibiting Ligand-Induced Nonalcoholic Fatty Liver and Drug-Induced Lipogenesis. J. Agric. Food Chem. 2018, 66, 11647–11662. [Google Scholar] [CrossRef]
- Zhang, F.; Hu, Z.; Li, G.; Huo, S.; Ma, F.; Cui, A.; Xue, Y.; Han, Y.; Gong, Q.; Gao, J.; et al. Hepatic CREBZF couples insulin to lipogenesis by inhibiting insig activity and contributes to hepatic steatosis in diet-induced insulin-resistant mice. Hepatology 2018, 68, 1361–1375. [Google Scholar] [CrossRef]
- Ma, F.; Liu, Y.; Hu, Z.; Xue, Y.; Liu, Z.; Cai, G.; Su, W.; Zheng, Z.; Fang, X.; Yan, X.; et al. Intrahepatic osteopontin signaling by CREBZF defines a checkpoint for steatosis-to-NASH progression. Hepatology 2023. [Google Scholar] [CrossRef]
- Vecchi, C.; Montosi, G.; Zhang, K.; Lamberti, I.; Duncan, S.A.; Kaufman, R.J.; Pietrangelo, A. ER stress controls iron metabolism through induction of hepcidin. Science 2009, 325, 877–880. [Google Scholar] [CrossRef]
- Pietrangelo, A.; Dierssen, U.; Valli, L.; Garuti, C.; Rump, A.; Corradini, E.; Ernst, M.; Klein, C.; Trautwein, C. STAT3 is required for IL-6-gp130-dependent activation of hepcidin in vivo. Gastroenterology 2007, 132, 294–300. [Google Scholar] [CrossRef]
- Saad, H.K.M.; Rahman, A.A.A.; Ab Ghani, A.S.; Taib, W.R.W.; Ismail, I.; Johan, M.F.; Al-Wajeeh, A.S.; Al-Jamal, H.A.N. Activation of STAT and SMAD Signaling Induces Hepcidin Re-Expression as a Therapeutic Target for β-Thalassemia Patients. Biomedicines 2022, 10, 189. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Kim, Y.H.; Jung, Y.S.; Kim, K.S.; Jeong, J.H.; Lee, Y.S.; Yuk, J.M.; Oh, B.C.; Choy, H.E.; Dooley, S.; et al. Orphan nuclear receptor SHP regulates iron metabolism through inhibition of BMP6-mediated hepcidin expression. Sci. Rep. 2016, 6, 34630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-H.; Lee, G.T.; Kwon, S.J.; Jeong, J.; Ha, Y.-S.; Kim, W.-J.; Kim, I.Y. CREBZF, a novel Smad8-binding protein. Mol. Cell. Biochem. 2012, 368, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.; Lee, J.; Lou, Z.; Lee, B.S.; Kim, C.H.; Lee, S.H. Identification of epithelial-specific ETS-1 (ESE-1) as a tumor suppressor and molecular target of green tea compound, EGCG. Mol. Carcinog. 2019, 58, 922–932. [Google Scholar] [CrossRef]
- Omori, Y. CREB-H: A novel mammalian transcription factor belonging to the CREB/ATF family and functioning via the box-B element with a liver-specific expression. Nucleic Acids Res. 2001, 29, 2154–2162. [Google Scholar] [CrossRef]
- Misra, J.; Chanda, D.; Kim, D.-K.; Li, T.; Koo, S.-H.; Back, S.-H.; Chiang, J.Y.; Choi, H.-S. Curcumin differentially regulates endoplasmic reticulum stress through transcriptional corepressor SMILE (small heterodimer partner-interacting leucine zipper protein)-mediated inhibition of CREBH (cAMP responsive element-binding protein H). J. Biol. Chem. 2011, 286, 41972–41984. [Google Scholar] [CrossRef]
- Jang, H. Regulation of Cyclic AMP-Response Element Binding Protein Zhangfei (CREBZF) Expression by Estrogen in Mouse Uterus. Dev. Reprod. 2018, 22, 95–104. [Google Scholar] [CrossRef]
- Zhang, R.; Rapin, N.; Ying, Z.; Shklanka, E.; Bodnarchuk, T.W.; Verge, V.M.K.; Misra, V. Zhangfei/CREB-ZF—A potential regulator of the unfolded protein response. PLoS ONE 2013, 8, e77256. [Google Scholar] [CrossRef]
- Jang, H. Differential Expression of Cyclic AMP-Response Element Binding Protein Zhangfei (CREBZF) in the Mouse Testis during Postnatal Development. Dev. Reprod. 2018, 22, 65–72. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, R.; Yu, T.; Wang, L.; Liu, S.; Cai, R.; Guo, X.; Jia, Y.; Wang, A.; Jin, Y.; et al. CREBZF regulates testosterone production in mouse Leydig cells. J. Cell Physiol. 2019, 234, 22819–22832. [Google Scholar] [CrossRef]
- Truong, X.T.; Lee, Y.-S.; Nguyen, T.T.P.; Kim, H.-J.; Kim, S.-H.; Moon, C.; Kim, D.-K.; Choi, H.-S.; Jeon, T.-I. SMILE Downregulation during Melanogenesis Induces MITF Transcription in B16F10 Cells. Int. J. Mol. Sci. 2022, 23, 15094. [Google Scholar] [CrossRef]
- Jang, W.-G.; Kim, E.-J.; Kim, D.-K.; Ryoo, H.-M.; Lee, K.-B.; Kim, S.-H.; Choi, H.-S.; Koh, J.-T. BMP2 protein regulates osteocalcin expression via Runx2-mediated Atf6 gene transcription. J. Biol. Chem. 2012, 287, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.; Kim, E.-J.; Park, J.-K.; Kim, D.-E.; Kim, H.-J.; Sun, W.-S.; Hwang, S.; Oh, K.-B.; Koh, J.-T.; Jang, W.-G.; et al. SMILE inhibits BMP-2-induced expression of osteocalcin by suppressing the activity of the RUNX2 transcription factor in MC3T3E1 cells. Bone 2014, 61, 10–18. [Google Scholar] [CrossRef]
- Lv, X.; Gao, F.; Li, T.P.; Xue, P.; Wang, X.; Wan, M.; Hu, B.; Chen, H.; Jain, A.; Shao, Z.; et al. Skeleton interoception regulates bone and fat metabolism through hypothalamic neuroendocrine NPY. eLife 2021, 10, e70324. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadasivam, N.; Radhakrishnan, K.; Choi, H.-S.; Kim, D.-K. Emerging Role of SMILE in Liver Metabolism. Int. J. Mol. Sci. 2023, 24, 2907. https://doi.org/10.3390/ijms24032907
Sadasivam N, Radhakrishnan K, Choi H-S, Kim D-K. Emerging Role of SMILE in Liver Metabolism. International Journal of Molecular Sciences. 2023; 24(3):2907. https://doi.org/10.3390/ijms24032907
Chicago/Turabian StyleSadasivam, Nanthini, Kamalakannan Radhakrishnan, Hueng-Sik Choi, and Don-Kyu Kim. 2023. "Emerging Role of SMILE in Liver Metabolism" International Journal of Molecular Sciences 24, no. 3: 2907. https://doi.org/10.3390/ijms24032907
APA StyleSadasivam, N., Radhakrishnan, K., Choi, H. -S., & Kim, D. -K. (2023). Emerging Role of SMILE in Liver Metabolism. International Journal of Molecular Sciences, 24(3), 2907. https://doi.org/10.3390/ijms24032907