

Long-Term Soil Drought Limits Starch Accumulation by Altering Sucrose Transport and Starch Synthesis in Sweet Potato Tuberous Root

 and
and
Abstract
:1. Introduction
2. Results
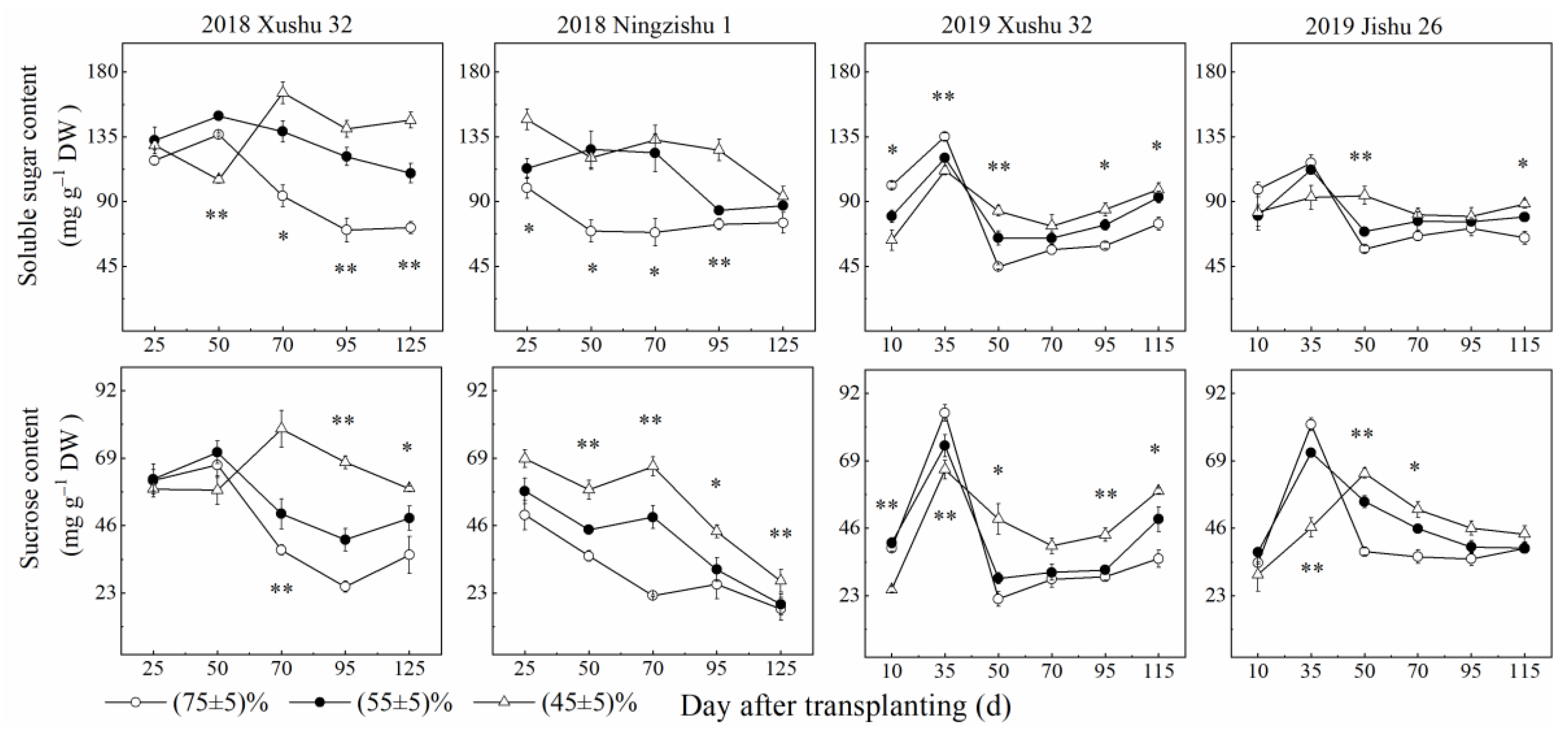
2.1. Effects of Long-Term Soil Drought Stress on Carbohydrate Content in Sweet Potato TRs
2.1.1. Soluble Sugar and Sucrose Content in TRs
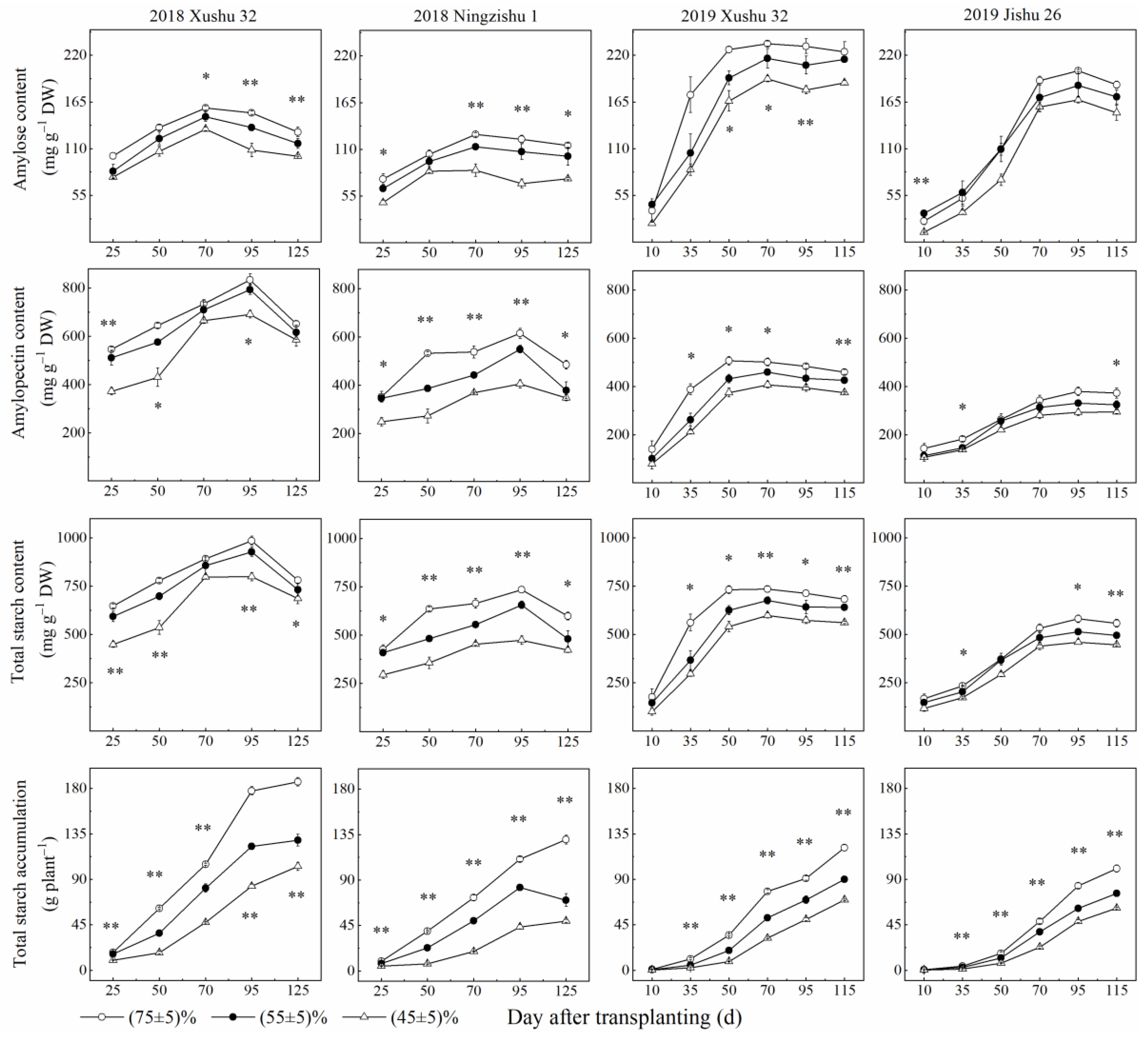
2.1.2. Starch Content and Starch Accumulation in TRs
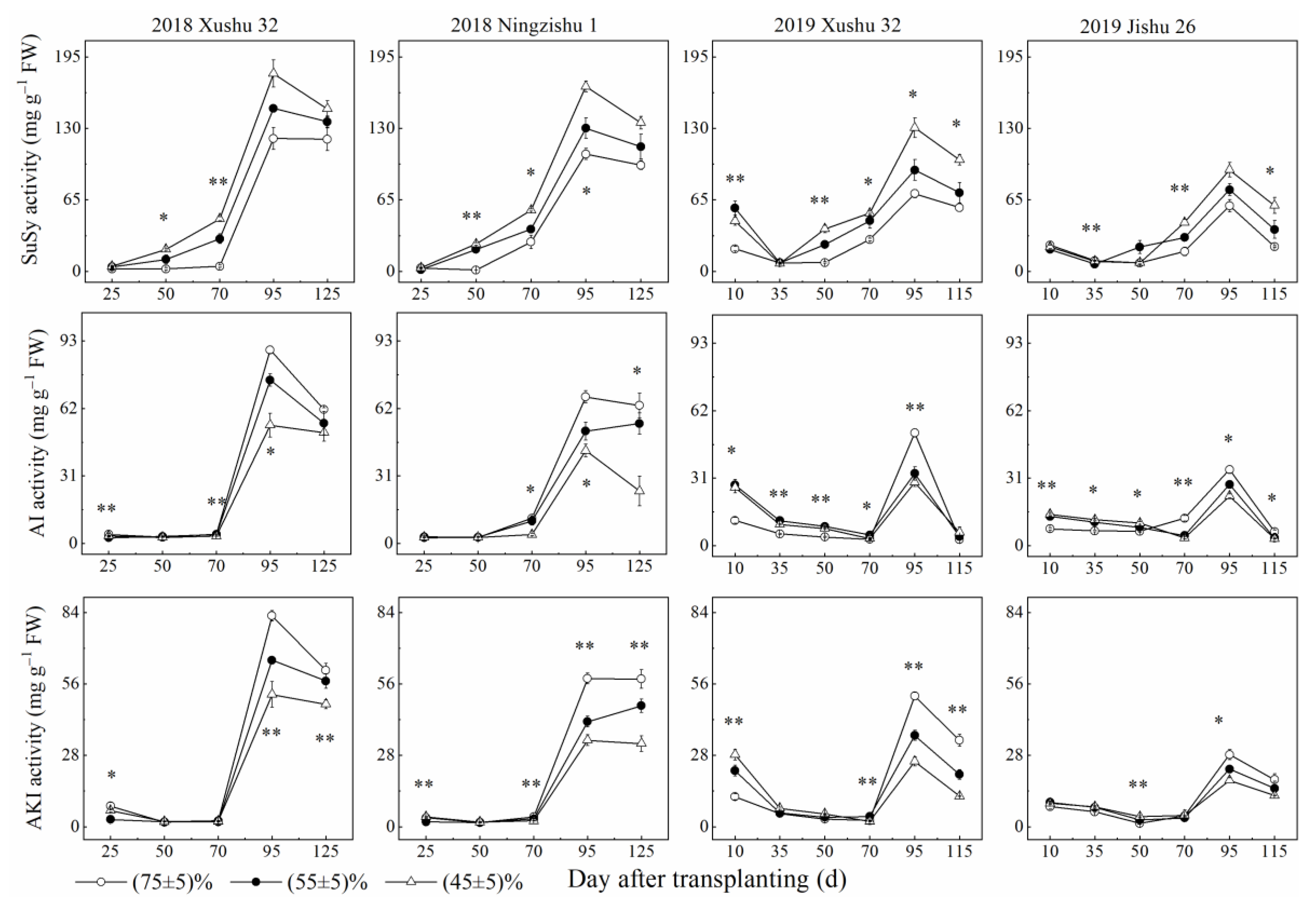
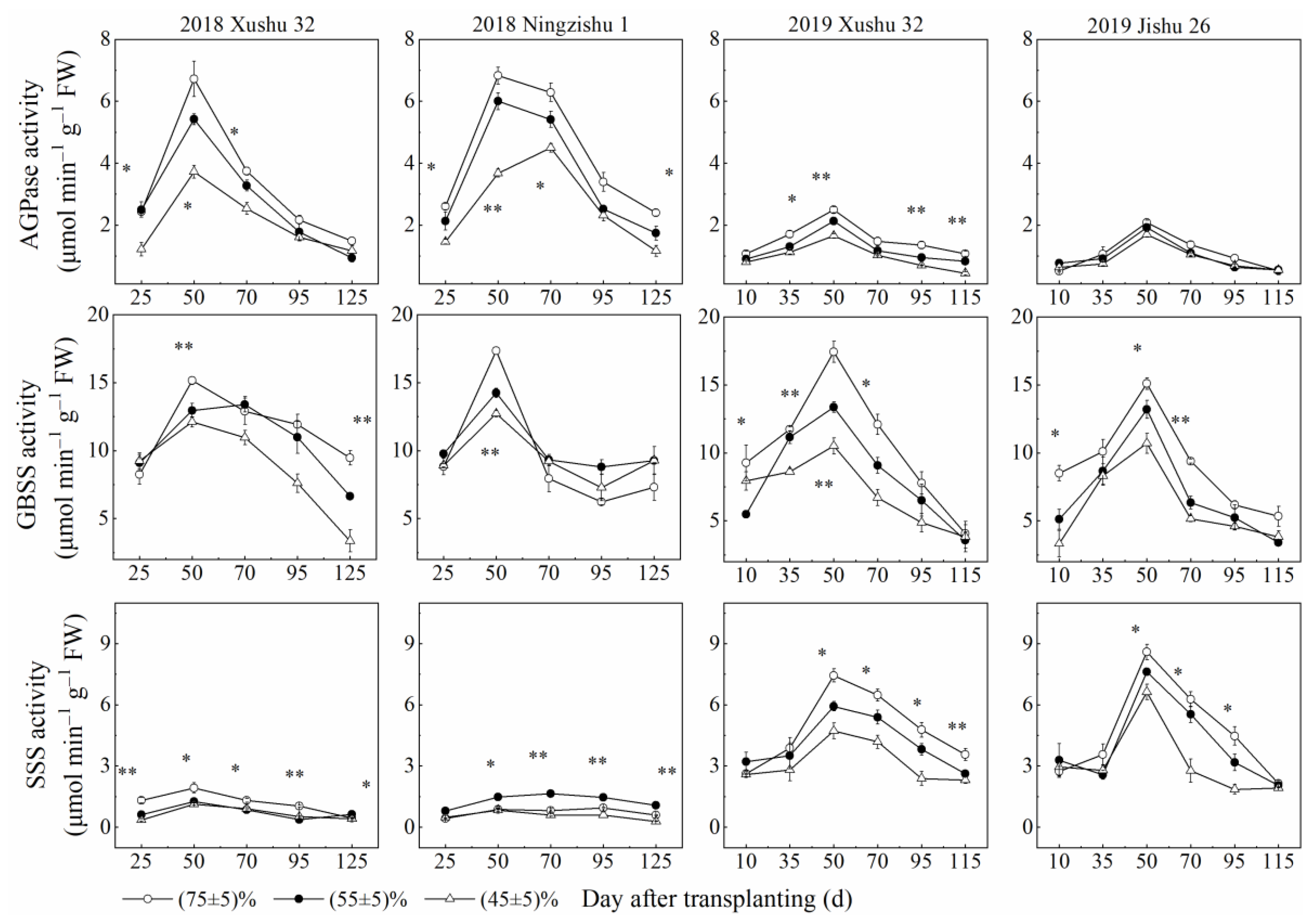
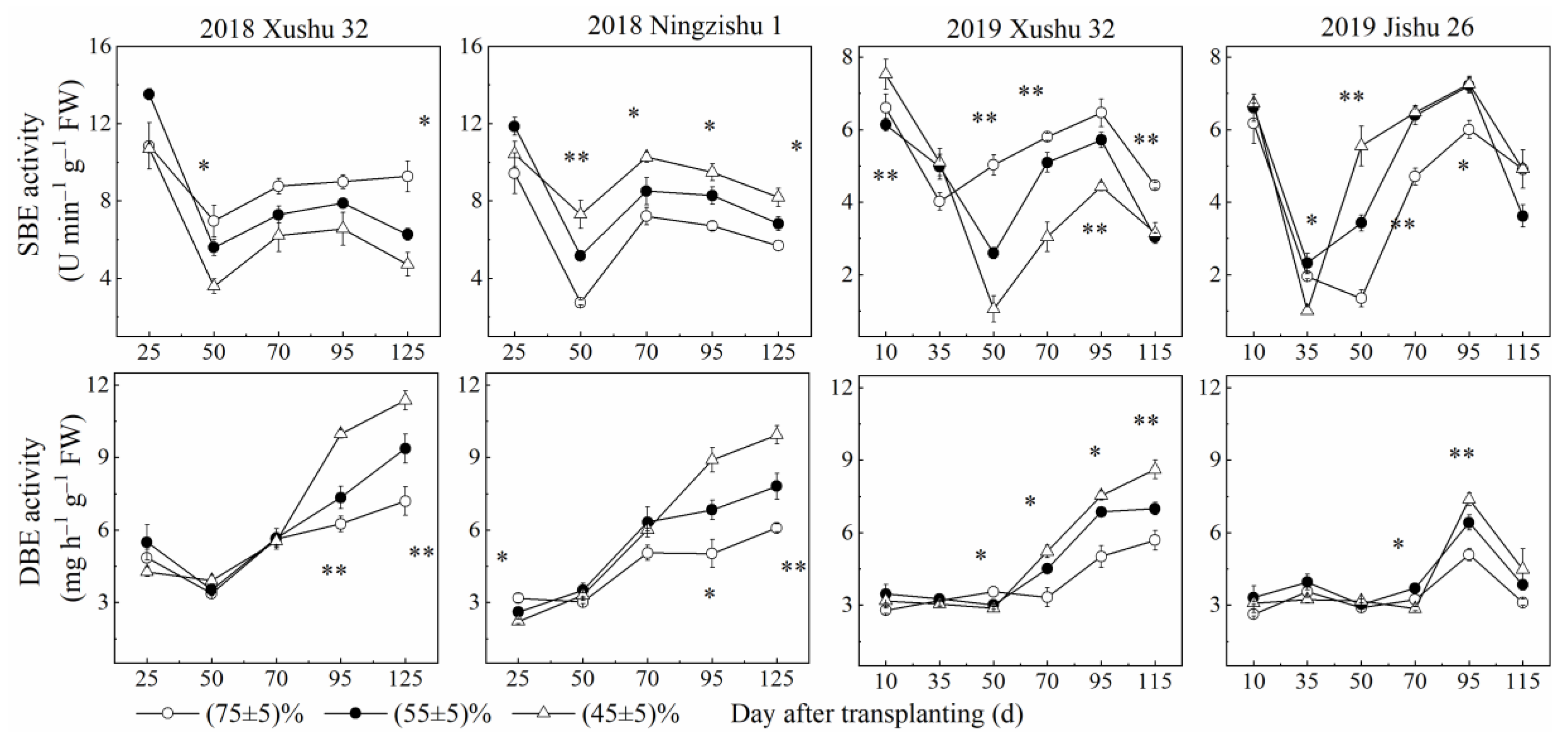
2.2. Effects of Long-Term Soil Drought Stress on Activities of Enzymes in Sucrose-Starch Conversion in Sweet Potato TRs
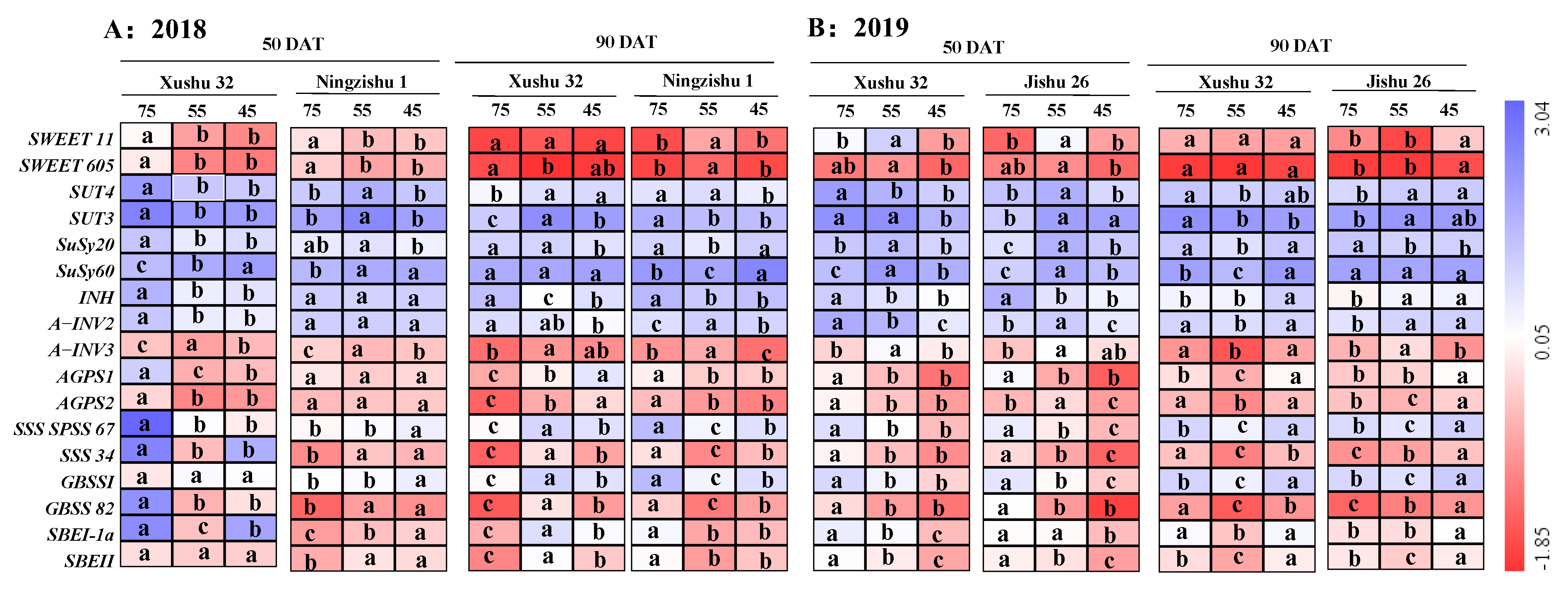
2.3. Effects of Long-Term Soil Drought Stress on the Genes Expression Levels in Sucrose-Starch Conversion of TRs
2.4. Correlation Analysis of between Starch Accumulation with Its Related Enzymes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
4.2. Sweet Potato Sampling
4.3. Extraction and Activity Assays for Sugars and Sucrose-Starch Metabolizing Enzymes
4.4. QRT-PCR Analysis in TRs
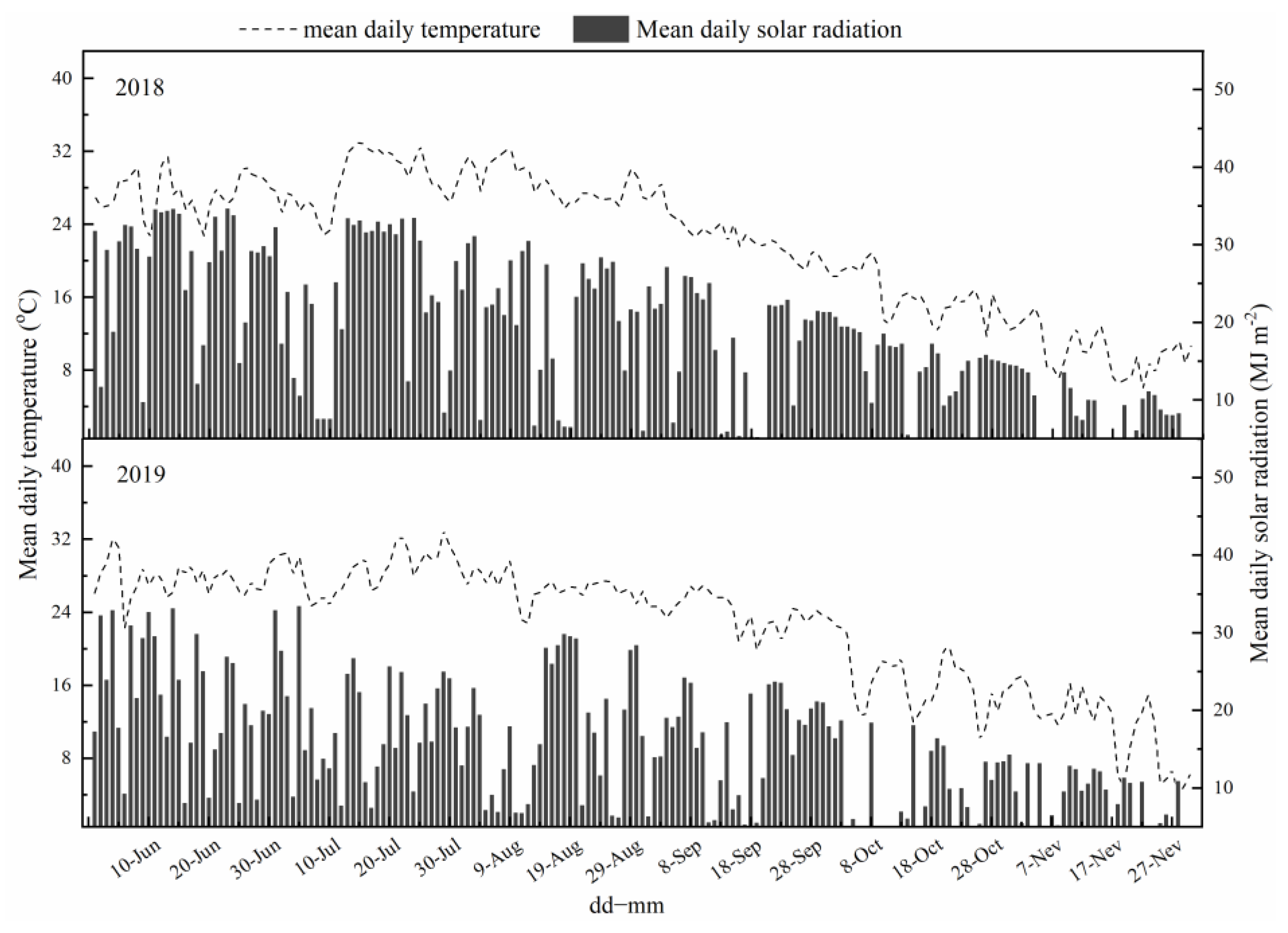
4.5. Statistical Analysis and Weather Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhai, H.; Wang, F.; Si, Z.; Huo, J.; Xing, L.; An, Y.; He, S.; Liu, Q. A myo-inositol-1-phosphate synthase gene, IbMIPS1, enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potato. Plant Biotechnol. J. 2016, 14, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Xu, T.; Zhang, J.; Shen, K.; Li, Z.; Liu, J. Drought-induced responses of nitrogen metabolism in Ipomoea batatas. Plants 2020, 9, 1341. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.; Li, J.; Wang, P.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Li, Q.; Cao, Q.; Niu, F.; Xie, Y.; Tang, J.; Li, H. Development and prospect of sweetpotato industry and its technologies in China. Jiangsu J. Agric. Sci. 2012, 28, 969–973. (In Chinese) [Google Scholar]
- Ruan, L.; Chen, L.; Chen, Y.; He, J.; Zhang, W.; Gao, Z.; Zhang, Y. Expression of Arabidopsis HOMEODOMAIN GLABROUS 11 enhances tolerance to drought stress in transgenic sweet potato plants. J. Plant Biol. 2012, 55, 151–158. [Google Scholar] [CrossRef]
- Fan, W.; Zhang, Y.; Wu, Y.; Zhou, W.; Yang, J.; Yuan, L.; Zhang, P.; Wang, H. The H+-pyrophosphatase IbVP1 regulates carbon flux to influence the starch metabolism and yield of sweet potato. Hortic. Res. 2021, 8, 20. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, Z.; Tang, D.; Luo, K.; Lu, H.; Liu, Y.; Dong, J.; Wang, X.; Lv, C.; Wang, J.; et al. Comparative transcriptome analysis reveals critical function of sucrose metabolism related-enzymes in starch accumulation in the storage root of sweet potato. Front. Plant Sci. 2017, 8, 914. [Google Scholar] [CrossRef]
- Rudack, K.; Seddig, S.; Sprenger, H.; Köhl, K.; Uptmoor, R.; Ordon, F. Drought stress-induced changes in starch yield and physiological traits in potato. J. Agron. Crop Sci. 2017, 203, 494–505. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, W.Q.; Xia, H.Q.; Gao, Y.; Tang, Z.H.; Zhang, J.Y.; Li, Z.Y.; Liu, J.R. Effects of soil drought stress on photosynthesis and yield of sweet potato. J. Jiangsu Norm. Univ. Nat. Sci. Ed. 2019, 37, 21–25. (In Chinese) [Google Scholar]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Durand, M.; Porcheron, B.; Hennion, N.; Maurousset, L.; Lemoine, R.; Pourtau, N. Water deficit enhances c export to the roots in Arabidopsis thaliana plants with contribution of sucrose transporters in both shoot and roots. Plant Physiol. 2016, 170, 1460–1479. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Naskar, S.; Makeshkumar, T.; Babu, B.; Krishnan, B.P. Molecular physiology of storage root formation and development in sweet potato (Ipomoea batatas (L.) Lam.). J. Root Crop. 2009, 35, 1–27. [Google Scholar]
- Yang, D.; Xie, Y.; Sun, H.; Bian, X.; Ke, Q.; Kim, H.S.; Ji, C.y.; Jin, R.; Wang, W.; Zhang, C.; et al. IbINH positively regulates drought stress tolerance in sweetpotato. Plant Physiol. Biochem. 2020, 146, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Lin, L.; Deng, Q.; Luo, X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Aliche, E.B.; Theeuwen, T.P.J.M.; Oortwijn, M.; Visser, R.G.F.; van der Linden, C.G. Carbon partitioning mechanisms in potato under drought stress. Plant Physiol. Biochem. 2020, 146, 211–219. [Google Scholar] [CrossRef]
- Hu, W.; Huang, Y.; Loka, D.A.; Bai, H.; Liu, Y.; Wang, S.; Zhou, Z. Drought-induced disturbance of carbohydrate metabolism in anthers and male abortion of two Gossypium hirsutum cultivars differing in drought tolerance. Plant Cell Rep. 2020, 39, 195–206. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, P.; Sharma, M.; Tuli, R.; Dhaliwal, H.S.; Chaudhury, A.; Pal, D.; Roy, J. Expression patterns of genes involved in starch biosynthesis during seed development in bread wheat (Triticum aestivum). Mol. Breed. 2015, 35, 184. [Google Scholar] [CrossRef]
- Li, X.-Q.; Zhang, D. Gene expression activity and pathway selection for sucrose metabolism in developing storage root of sweet potato. Plant Cell Physiol. 2003, 44, 630–636. [Google Scholar] [CrossRef]
- Bahaji, A.; Li, J.; Sánchez-López, Á.M.; Baroja-Fernández, E.; Muñoz, F.J.; Ovecka, M.; Almagro, G.; Montero, M.; Ezquer, I.; Etxeberria, E.; et al. Starch biosynthesis, its regulation and biotechnological approaches to improve crop yields. Biotechnol. Adv. 2014, 32, 87–106. [Google Scholar] [CrossRef]
- Zhang, W.; Gu, J.; Wang, Z.; Wei, C.; Yang, J.; Zhang, J. Comparison of structural and functional properties of wheat starch under different soil drought conditions. Sci. Rep. 2017, 7, 12312. [Google Scholar] [CrossRef]
- Wang, S.J.; Liu, L.F.; Chen, C.K.; Chen, L.W. Regulations of granule-bound starch synthase I gene expression in rice leaves by temperature and drought stress. Biol. Plant. 2006, 50, 537–541. [Google Scholar] [CrossRef]
- Kim, S.-H.; Mizuno, K.; Fujimura, T. Regulated expression of ADPglucose pyrophosphorylase and chalcone synthase during root development in sweet potato. Plant Growth Regul. 2002, 38, 173–179. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, H.; Liu, M.; Wang, W. Effect of drought on the contents of soluble sugars, starch and enzyme activities in cassava stem. Plant Physiol. J. 2017, 53, 795–806. (In Chinese) [Google Scholar]
- Zhou, Y.-X.; Chen, Y.-X.; Tao, X.; Cheng, X.-J.; Wang, H.-Y. Isolation and characterization of cDNAs and genomic DNAs encoding ADP-glucose pyrophosphorylase large and small subunits from sweet potato. Mol. Genet. Genom. 2016, 291, 609–620. [Google Scholar] [CrossRef]
- Gámez-Arjona, F.M.; Li, J.; Raynaud, S.; Baroja-Fernández, E.; Muñoz, F.J.; Ovecka, M.; Ragel, P.; Bahaji, A.; Pozueta-Romero, J.; Mérida, Á. Enhancing the expression of starch synthase class IV results in increased levels of both transitory and long-term storage starch. Plant Biotechnol. J. 2011, 9, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Saito, A. Heterogeneity of poly(A) Sites in the granule-bound starch synthase I gene in sweet potato (Ipomoea batatas (L.) Lam.). Biosci. Biotechnol. Biochem. 2010, 74, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Yan, H.; Ma, M.; Kou, M.; Li, C.; Tang, W.; Yu, Y.; Hao, Q.; Nguyen, T.; Wang, X.; et al. Comparative transcriptome profiling reveals the genes involved in storage root expansion in sweetpotato (Ipomoea batatas (L.) Lam.). Genes 2022, 13, 1156. [Google Scholar] [CrossRef]
- Lai, Y.C.; Wang, S.Y.; Gao, H.Y.; Nguyen, K.M.; Nguyen, C.H.; Shih, M.C.; Lin, K.H. Physicochemical properties of starches and expression and activity of starch biosynthesis-related genes in sweet potatoes. Food Chem. 2016, 199, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, Y.; Chen, T.; Zhang, H.; Yang, J.; Zhang, J. Abscisic acid and the key enzymes and genes in sucrose-to-starch conversion in rice spikelets in response to soil drying during grain filling. Planta 2015, 241, 1091–1107. [Google Scholar] [CrossRef]
- Li, S.; Zhao, L.; Sun, N.; Liu, Q.; Li, H. Photosynthesis product allocation and yield in sweetpotato with different irrigation levels at mid-season. Agric. Water Manag. 2021, 246, 106708. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.; Liu, Q. Photosynthesis product allocation and yield in sweet potato with spraying exogenous hormones under drought stress. J. Plant Physiol. 2020, 253, 153265. [Google Scholar]
- Wang, J.; Li, S.; Liu, Q.; Li, H. Mechanism of spraying growth regulators to alleviate drought stress of sweet potato. Sci. Agric. Sin. 2020, 53, 500–512. [Google Scholar]
- Wang, J.; Li, H.; Liu, Q.; Zeng, L. Effects of exogenous plant hormones on physiological characteristics and yield of sweet potato under drought stress. Chin. J. Appl. Ecol. 2020, 31, 189–198. (In Chinese) [Google Scholar]
- Wu, H.; Guo, Q.; Wang, J.; Li, H.; Liu, Q. Effects of water supply on photosynthesis and fluorescence characteristics of sweet potato [Ipomoea batatas (L.) Lam.] leaves and comparison of light response models. Chin. J. Eco-Agric. 2019, 27, 908–918. [Google Scholar]
- Hu, W.; Huang, Y.; Bai, H.; Liu, Y.; Wang, S.; Zhou, Z. Influence of drought stress on pistil physiology and reproductive success of two Gossypium hirsutum cultivars differing in drought tolerance. Physiol. Plant 2019, 168, 909–920. [Google Scholar] [CrossRef]
- Kuai, J.; Liu, Z.; Wang, Y.; Meng, Y.; Chen, B.; Zhao, W.; Zhou, Z.; Oosterhuis, D.M. Waterlogging during flowering and boll forming stages affects sucrose metabolism in the leaves subtending the cotton boll and its relationship with boll weight. Plant Sci. 2014, 223, 79–98. [Google Scholar] [CrossRef]
- Du, X.; Zhang, X.; Xi, M.; Kong, L. Split application enhances sweetpotato starch production by regulating the conversion of sucrose to starch under reduced nitrogen supply. Plant Physiol. Biochem. 2020, 151, 743–750. [Google Scholar] [CrossRef]
- Liu, H.J.; Yao, H.L.; Shi, C.Y.; Zhang, L.M. Effect of potassium application time on starch accumulation and related enzyme activities of sweet potato variety Jixu 23. Sci. Agric. Sin. 2014, 47, 43–52. (In Chinese) [Google Scholar]
- Liu, J.R.; Ma, Y.N.; Lv, F.J.; Chen, J.; Zhou, Z.G.; Wang, Y.H.; Abudurezike, A.; Oosterhuis, D.M. Changes of sucrose metabolism in leaf subtending to cotton boll under cool temperature due to late planting. Field Crop. Res. 2013, 144, 200–211. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, Z.; Xia, H.; Sheng, M.; Liu, M.; Pan, S.; Li, Z.; Liu, J. Potassium fertilization stimulates sucrose-to-starch conversion and root formation in sweet potato (Ipomoea batatas (L.) Lam.). Int. J. Mol. Sci. 2021, 22, 4826. [Google Scholar] [CrossRef]
- Shu, H.M.; Zhou, Z.G.; Xu, N.Y.; Wang, Y.H.; Zheng, M. Sucrose metabolism in cotton (Gossypium hirsutum L.) fibre under low temperature during fibre development. Eur. J. Agron. 2009, 31, 61–68. [Google Scholar] [CrossRef]
- Liu, J.; Peng, J.; Xia, H.; Li, P.; Li, Z.; Sun, M.; Zheng, C.; Dong, H. High soil available phosphorus favors carbon metabolism in cotton leaves in pot trials. J. Plant Growth Regul. 2021, 40, 974–985. [Google Scholar] [CrossRef]
- Nakamura, Y.; Umemoto, T.; Ogata, N.; Kuboki, Y.; Yano, M.; Sasaki, T. Starch debranching enzyme (R-enzyme or pullulanase) from developing rice endosperm: Purification, cDNA and chromosomal localization of the gene. Planta 1996, 199, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.A.; Petreikov, M. Sucrose-to-starch metabolism in tomato fruit undergoing transient starch accumulation. Plant Physiol. 1997, 113, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, Y.H.; Ji, C.Y.; Park, S.; Jeong, J.C.; Lee, H.S.; Kwak, S.S. Stable internal reference genes for the normalization of Real-time PCR in different sweetpotato cultivars subjected to abiotic stress conditions. PLoS ONE 2012, 7, e51502. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Cultivar | SRWC (%) | R2 | Smax | t1 | T | VT | Vmax |
|---|---|---|---|---|---|---|---|---|
| 2018 | Xushu 32 | 75 ± 5 | 0.9901 ** | 193.0 | 42.1 | 31.8 | 2.4 | 2.7 |
| 55 ± 5 | 0.9659 ** | 155.7 | 45.2 | 37.6 | 1.7 | 1.8 | ||
| 45 ± 5 | 0.9761 ** | 124.1 | 51.9 | 39.9 | 1.3 | 1.4 | ||
| Ningzishu 1 | 75 ± 5 | 0.9949 ** | 125.7 | 42.2 | 27.4 | 1.8 | 2.0 | |
| 55 ± 5 | 0.9867 ** | 96.3 | 46.8 | 30.8 | 1.3 | 1.4 | ||
| 45 ± 5 | 0.9523 ** | 61.6 | 58.0 | 41.0 | 0.6 | 0.7 | ||
| 2019 | Xushu 32 | 75 ± 5 | 0.9847 ** | 102.5 | 43.5 | 18.6 | 2.2 | 2.5 |
| 55 ± 5 | 0.9936 ** | 88.5 | 50.2 | 20.4 | 1.7 | 1.9 | ||
| 45 ± 5 | 0.9927 ** | 70.3 | 57.7 | 21.1 | 1.3 | 1.5 | ||
| Jishu 26 | 75 ± 5 | 0.9970 ** | 97.4 | 55.2 | 20.4 | 1.9 | 2.1 | |
| 55 ± 5 | 0.9898 ** | 76.8 | 56.8 | 21.7 | 1.4 | 1.6 | ||
| 45 ± 5 | 0.9865 ** | 61.8 | 61.0 | 19.2 | 1.3 | 1.4 |
| Correlation with | T | VT | Vmax | SuSy | AI | AKI | AGPase | GBSS | SSS | SBE | DBE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Starch accumulation | 0.391 | 0.662 * | 0.664 * | 0.128 | 0.828 ** | 0.894 ** | 0.578 * | 0.741 ** | −0.396 | 0.599 * | 0.419 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, M.; Xia, H.; Ding, H.; Pan, D.; He, J.; Li, Z.; Liu, J. Long-Term Soil Drought Limits Starch Accumulation by Altering Sucrose Transport and Starch Synthesis in Sweet Potato Tuberous Root. Int. J. Mol. Sci. 2023, 24, 3053. https://doi.org/10.3390/ijms24033053
Sheng M, Xia H, Ding H, Pan D, He J, Li Z, Liu J. Long-Term Soil Drought Limits Starch Accumulation by Altering Sucrose Transport and Starch Synthesis in Sweet Potato Tuberous Root. International Journal of Molecular Sciences. 2023; 24(3):3053. https://doi.org/10.3390/ijms24033053
Chicago/Turabian StyleSheng, Minfei, Houqiang Xia, Huizi Ding, Dongyu Pan, Jinping He, Zongyun Li, and Jingran Liu. 2023. "Long-Term Soil Drought Limits Starch Accumulation by Altering Sucrose Transport and Starch Synthesis in Sweet Potato Tuberous Root" International Journal of Molecular Sciences 24, no. 3: 3053. https://doi.org/10.3390/ijms24033053
APA StyleSheng, M., Xia, H., Ding, H., Pan, D., He, J., Li, Z., & Liu, J. (2023). Long-Term Soil Drought Limits Starch Accumulation by Altering Sucrose Transport and Starch Synthesis in Sweet Potato Tuberous Root. International Journal of Molecular Sciences, 24(3), 3053. https://doi.org/10.3390/ijms24033053