

Methyl Donor Micronutrients: A Potential Dietary Epigenetic Target in Systemic Lupus Erythematosus Patients
 , , , and
, , , and 
Abstract
:1. Introduction
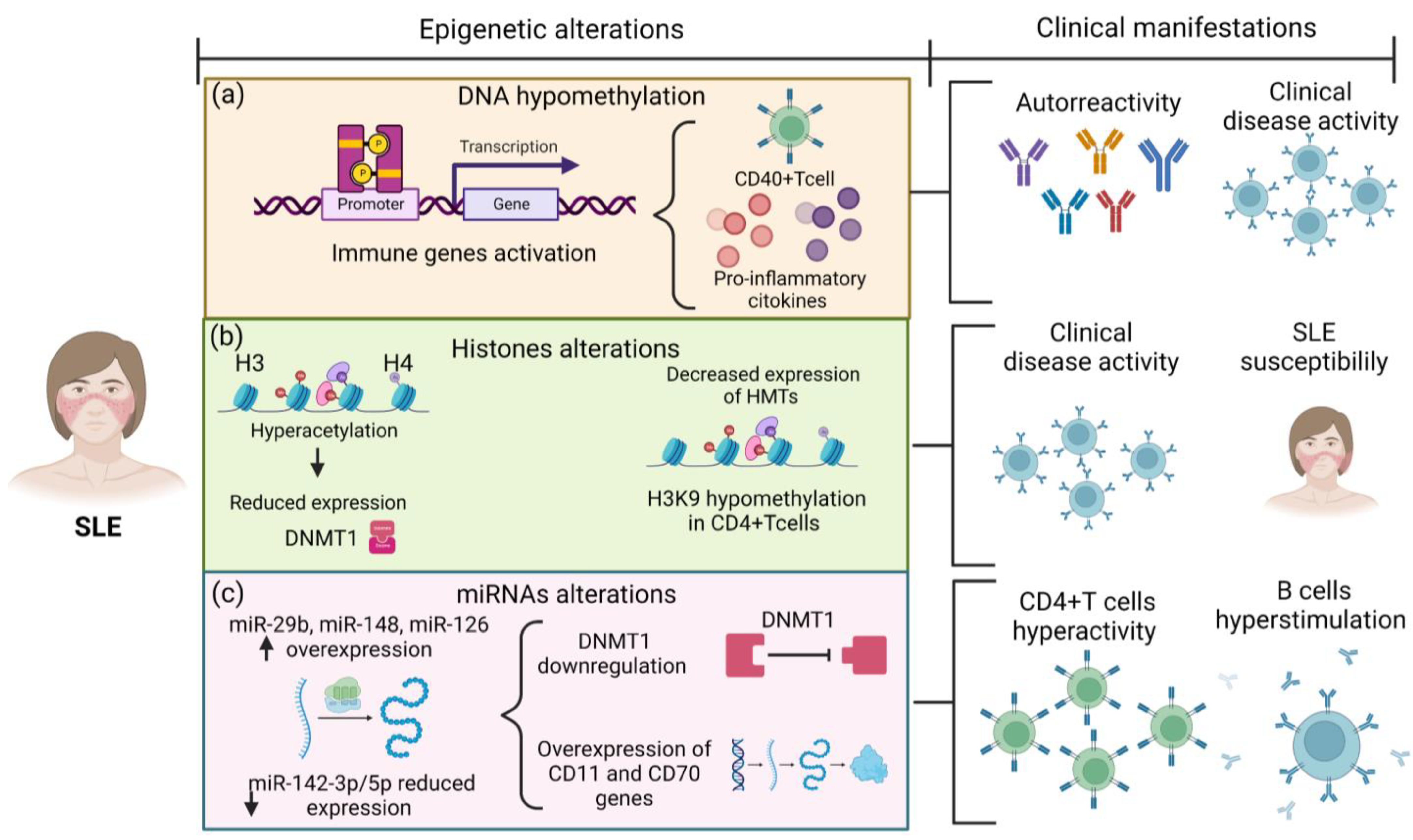
2. Epigenetic Mechanisms Related to SLE Pathophysiology
2.1. DNA Methylation and SLE
2.2. ncRNAs and SLE
2.3. Histones Post-Translational Modifications and SLE
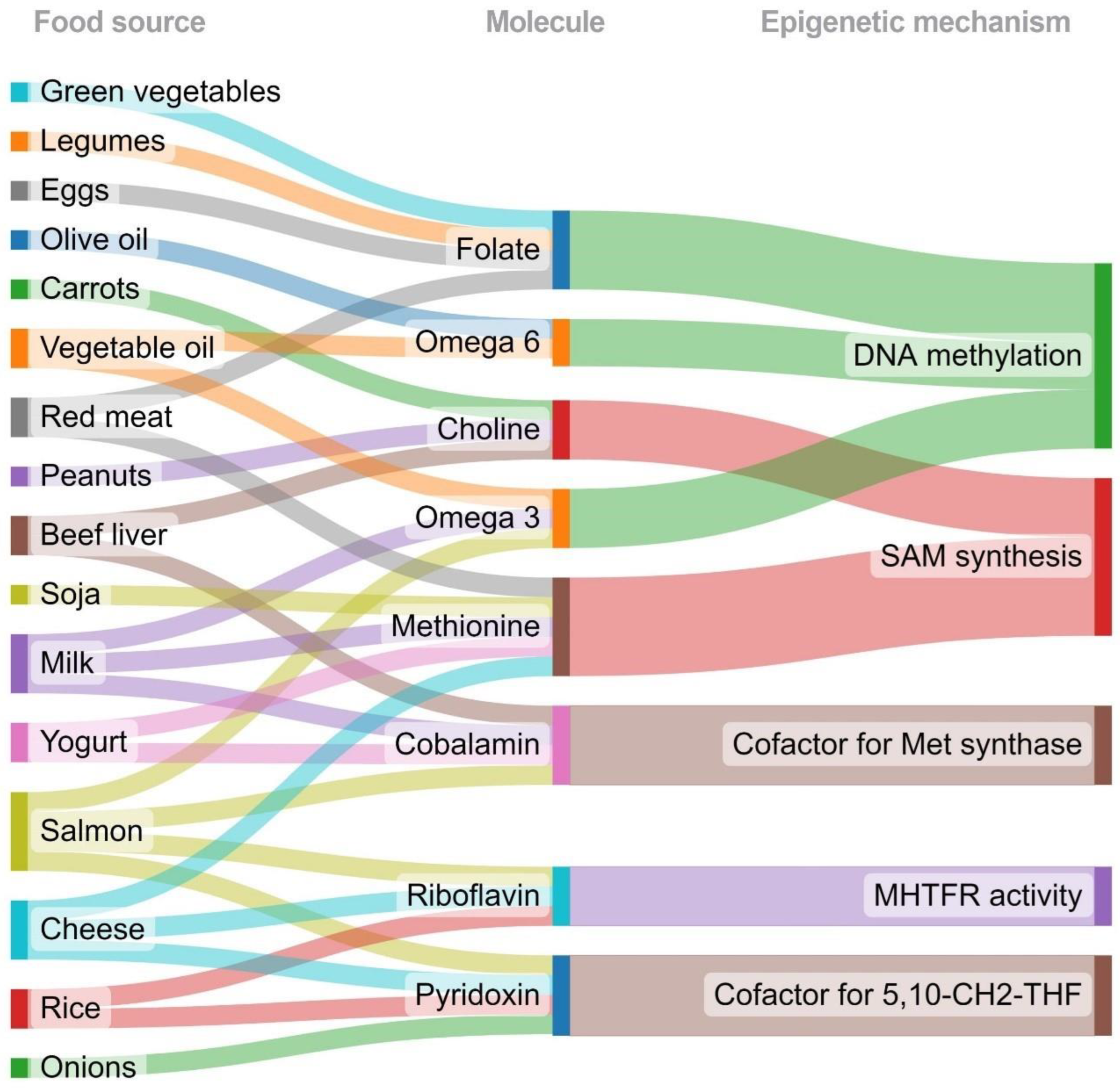
3. Nutrients’ Role in Epigenetic Modifications
3.1. Folate
3.2. Choline and Betaine
3.3. B Vitamins
3.4. Methionine
3.5. Fatty Acids
3.6. Other Environmental Factors That Impact DNA Methylation
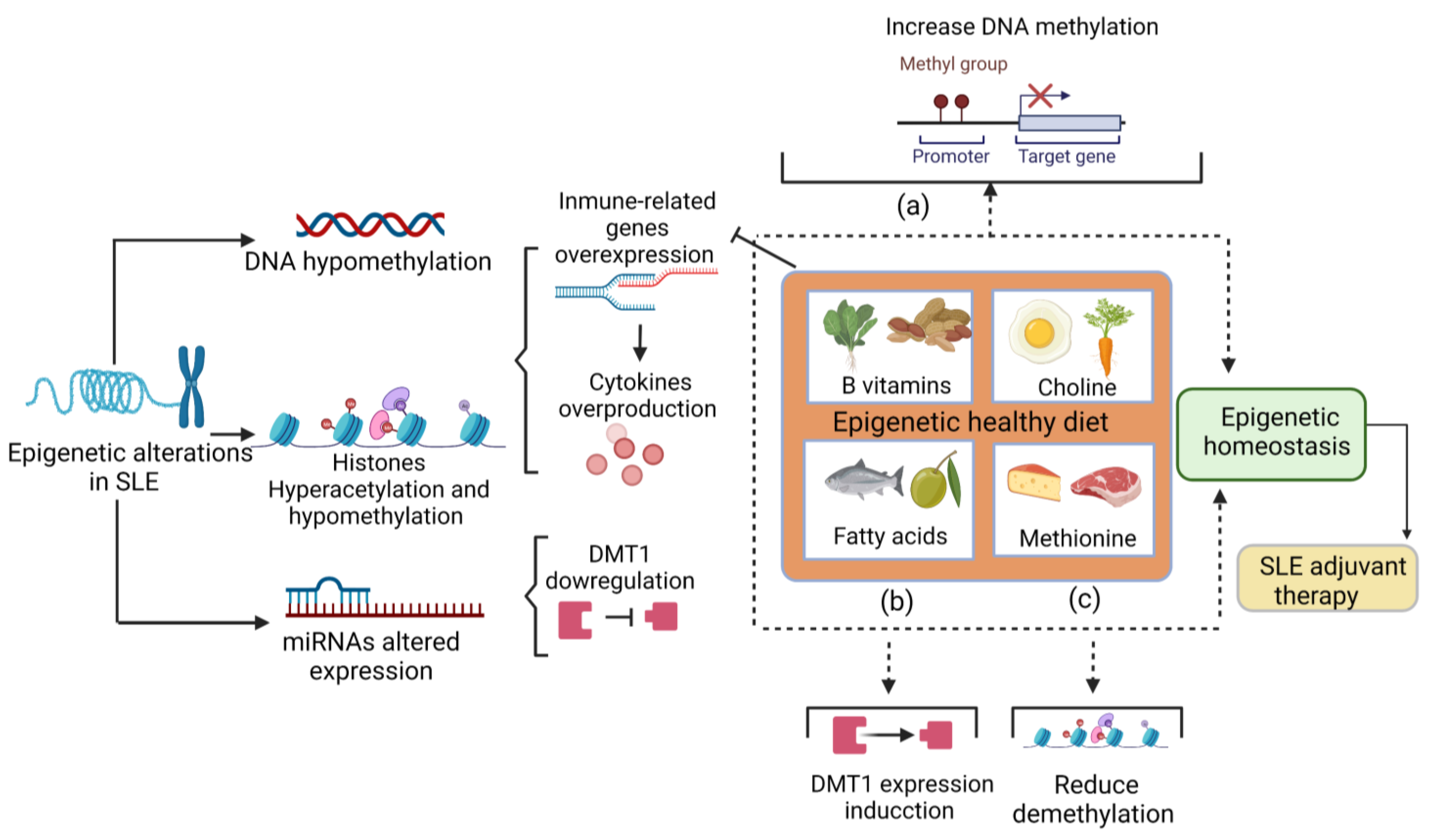
4. Diet as an Epigenetic Therapy in SLE: A New Paradigm?
A Diet to Increase Methylation in SLE
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SLE | systemic lupus erythematosus |
| miRNAs | micro RNAs |
| MTHFR | methylenetetrahydrofolate reductase |
| DNMTs | DNA methyltransferases |
| HMTs | histone methyltransferases |
| HDMTs | histone demethylases |
| 5-methylTHF | 5-methyltetrahydrofolate |
| SAM | S -adenosyl methionine |
| CDKN3 | cyclin-dependent kinase 3 |
| VEGF-C | vascular endothelial growth factor C |
| IGF-2 | insulin-like growth factor-2 |
| PUFAs | polyunsaturated fatty acids |
| DHA | docosahexaenoic acid |
| EPA | eicosapentaenoic acid |
| LA | linoleic acid |
References
- Montoya, T.; Castejón, M.L.; Muñoz-García, R.; Alarcón-de-la-Lastra, C. Epigenetic linkage of systemic lupus erythematosus and nutrition. Nutr. Res. Rev. 2021, 16, 1–21. [Google Scholar]
- Kaul, A.; Gordon, C.; Crow, M.K.; Touma, Z.; Urowitz, M.B.; Van Vollenhoven, R.; Ruiz-Irastorza, G.; Hughes, G. Systemic lupus erythematosus. Nat. Rev. Dis. Prim. 2016, 2, 16039. [Google Scholar] [CrossRef]
- Oku, K.; Atsumi, T. Systemic lupus erythematosus: Nothing stale her infinite variety. Mod. Rheumatol. 2018, 28, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Pons-Estel, G.J.; Alarcón, G.S.; Scofield, L.; Reinlib, L.; Cooper, G.S. Understanding the Epidemiology and Progression of Systemic Lupus Erythematosus. Semin. Arthritis Rheum. 2010, 39, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Renaudineau, Y.; Youinou, P. Epigenetics and Autoimmunity, with Special Emphasis on Methylation. Keio J. Med. 2011, 60, 10–16. [Google Scholar] [CrossRef] [PubMed]
- King, S.E.; Skinner, M.K. Epigenetic Transgenerational Inheritance of Obesity Susceptibility. Trends Endocrinol. Metab. 2020, 31, 478–494. [Google Scholar] [CrossRef]
- Daniel, M.; Tollefsbol, T.O. Epigenetic linkage of aging, cancer and nutrition. J. Exp. Biol. 2015, 218, 59–70. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef]
- Anderson, O.S.; Sant, K.E.; Dolinoy, D.C. Nutrition and epigenetics: An interplay of dietary methyl donors, one-carbon metabolism and DNA methylation. J. Nutr. Biochem. 2012, 23, 853–859. [Google Scholar] [CrossRef]
- Hughes, A.L.; Kelley, J.R.; Klose, R.J. Understanding the interplay between CpG island-associated gene promoters and H3K4 methylation. Biochim. Et Biophys. Acta (BBA) Gene Regul. Mech. 2020, 1863, 194567. [Google Scholar] [CrossRef]
- Surace, A.E.A.; Hedrich, C.M. The Role of Epigenetics in Autoimmune/Inflammatory Disease. Front. Immunol. 2019, 10, 1525. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ou, T.; Wu, C.; Li, R.; Lin, Y.; Lin, C.; Tsai, W.; Liu, H.; Yen, J. Global DNA methylation, DNMT1, and MBD2 in patients with systemic lupus erythematosus. Lupus 2011, 20, 131–136. [Google Scholar] [CrossRef]
- Catalina, M.D.; Owen, K.A.; Labonte, A.C.; Grammer, A.C.; Lipsky, P.E. The pathogenesis of systemic lupus erythematosus: Harnessing big data to understand the molecular basis of lupus. J. Autoimmun. 2020, 110, 102359. [Google Scholar] [CrossRef]
- Lu, Q.; Kaplan, M.; Ray, D.; Ray, D.; Zacharek, S.; Gutsch, D.; Richardson, B. Demethylation ofITGAL (CD11a) regulatory sequences in systemic lupus erythematosus. Arthritis Rheum. 2002, 46, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Balada, E.; Ordi-Ros, J.; Vilardell-Tarres, M. DNA Methylation and Systemic Lupus Erythematosus. Ann. N. Y. Acad. Sci. 2007, 1108, 127–136. [Google Scholar] [CrossRef]
- Ballestar, E.; Esteller, M.; Richardson, B.C. The Epigenetic Face of Systemic Lupus Erythematosus. J. Immunol. 2006, 176, 7143–7147. [Google Scholar] [CrossRef] [PubMed]
- Vordenbäumen, S.; Rosenbaum, A.; Gebhard, C.; Raithel, J.; Sokolowski, A.; Düsing, C.; Chehab, G.; Richter, J.G.; Brinks, R.; Rehli, M.; et al. Associations of site-specific CD4+-T-cell hypomethylation within CD40-ligand promotor and enhancer regions with disease activity of women with systemic lupus erythematosus. Lupus 2021, 30, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Tang, J.; Gao, F.; Wu, X.; Liang, Y.; Yin, H.; Lu, Q. Hypomethylation of IL10 and IL13 Promoters in CD4+ T Cells of Patients with Systemic Lupus Erythematosus. J. Biomed. Biotechnol. 2010, 2010, 931018. [Google Scholar] [CrossRef]
- Qin, H.; Zhu, X.; Liang, J.; Wu, J.; Yang, Y.; Wang, S.; Shi, W.; Xu, J. MicroRNA-29b contributes to DNA hypomethylation of CD4+ T cells in systemic lupus erythematosus by indirectly targeting DNA methyltransferase 1. J. Dermatol. Sci. 2013, 69, 61–67. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, Y.; Liang, Y.; Zhao, M.; Long, H.; Ding, S.; Yin, H.; Lu, Q. MicroRNA-126 regulates DNA methylation in CD4+ T cells and contributes to systemic lupus erythematosus by targeting DNA methyltransferase 1. Arthritis Rheum. 2011, 63, 1376–1386. [Google Scholar] [CrossRef]
- Ding, S.; Zhang, Q.; Luo, S.; Gao, L.; Huang, J.; Lu, J.; Chen, J.; Zeng, Q.; Guo, A.; Zeng, J.; et al. BCL-6 suppresses miR-142-3p/5p expression in SLE CD4+ T cells by modulating histone methylation and acetylation of the miR-142 promoter. Cell. Mol. Immunol. 2020, 17, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Qiu, X.; Luo, Y.; Yuan, J.; Li, Y.; Zhong, Q.; Zhao, M.; Lu, Q. Histone modifications and methyl-CpG-binding domain protein levels at the TNFSF7 (CD70) promoter in SLE CD4+ T cells. Lupus 2011, 20, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.J.; Lin, Y.Z.; Liu, C.C.; Lin, C.H.; Li, R.N.; Wu, C.C.; Ou, T.T.; Tsai, W.C.; Yen, J.H. Methylation and gene expression of histone deacetylases 6 in systemic lupus erythematosus. Int. J. Rheum. Dis. 2016, 19, 968–973. [Google Scholar] [CrossRef]
- Dai, R.; Wang, Z.; Ahmed, S.A. Epigenetic Contribution and Genomic Imprinting Dlk1-Dio3 miRNAs in Systemic Lupus Erythematosus. Genes 2021, 12, 680. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Daskalaki, M.G.; Tsatsanis, C.; Kampranis, S.C. Histone methylation and acetylation in macrophages as a mechanism for regulation of inflammatory responses. J. Cell. Physiol. 2018, 233, 6495–6507. [Google Scholar] [CrossRef] [PubMed]
- Hedrich, C.M. Epigenetics in SLE. Curr. Rheumatol. Rep. 2017, 19, 58. [Google Scholar] [CrossRef]
- Shen, Y.; Wei, W.; Zhou, D.X. Histone Acetylation Enzymes Coordinate Metabolism and Gene Expression. Trends Plant Sci. 2015, 20, 614–621. [Google Scholar] [CrossRef]
- Gautam, P.; Sharma, A.; Bhatnagar, A. Global histone modification analysis reveals hypoacetylated H3 and H4 histones in B Cells from systemic lupus erythematosus patients. Immunol. Lett. 2021, 240, 41–45. [Google Scholar] [CrossRef]
- Zhao, S.; Long, H.; Lu, Q. Epigenetic Perspectives in Systemic Lupus Erythematosus: Pathogenesis, Biomarkers, and Therapeutic Potentials. Clin. Rev. Allergy Immunol. 2010, 39, 3–9. [Google Scholar] [CrossRef]
- Lu, Q.; Qiu, X.; Hu, N.; Wen, H.; Su, Y.; Richardson, B.C. Epigenetics, disease, and therapeutic interventions. Ageing Res. Rev. 2006, 5, 449–467. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.A.; Gretz, D.M.; Taylor, P.C.; James, S.J.; Pogribny, I.P.; Miller, B.J.; Henning, S.M.; Swendseid, M.E. Moderate Folate Depletion Increases Plasma Homocysteine and Decreases Lymphocyte DNA Methylation in Postmenopausal Women. J. Nutr. 1998, 128, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.M.; Mellott, T.J.; Kovacheva, V.P.; Blusztajn, J.K. Gestational Choline Supply Regulates Methylation of Histone H3, Expression of Histone Methyltransferases G9a (Kmt1c) and Suv39h1 (Kmt1a), and DNA Methylation of Their Genes in Rat Fetal Liver and Brain. J. Biol. Chem. 2009, 284, 1982–1989. [Google Scholar] [CrossRef]
- Kulkarni, A.; Dangat, K.; Kale, A.; Sable, P.; Chavan-Gautam, P.; Joshi, S. Effects of Altered Maternal Folic Acid, Vitamin B12 and Docosahexaenoic Acid on Placental Global DNA Methylation Patterns in Wistar Rats. PLoS ONE 2011, 6, e17706. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, S.; Duca, R.; Devlieger, R.; Freson, K.; Straetmans, D.; Van Herck, E.; Huybrechts, I.; Koppen, G.; Godderis, L. Maternal Methyl-Group Donor Intake and Global DNA (Hydroxy)Methylation before and during Pregnancy. Nutrients 2016, 8, 474. [Google Scholar] [CrossRef]
- Sarabi, M.M.; Naghibalhossaini, F. The impact of polyunsaturated fatty acids on DNA methylation and expression of DNMTs in human colorectal cancer cells. Biomed. Pharmacother. 2018, 101, 94–99. [Google Scholar] [CrossRef]
- Scaglione, F.; Panzavolta, G. Folate, folic acid and 5-methyltetrahydrofolate are not the same thing. Xenobiotica 2014, 44, 480–488. [Google Scholar] [CrossRef]
- Courtemanche, C.; Huang, A.C.; Elson-Schwab, I.; Kerry, N.; Ng, B.Y.; Ames, B.N. Folate deficiency and ionizing radiation cause DNA breaks in primary human lymphocytes: A comparison. FASEB J. 2004, 18, 209–211. [Google Scholar] [CrossRef]
- Wu, C.H.; Huang, T.C.; Lin, B.F. Folate deficiency affects dendritic cell function and subsequent T helper cell differentiation. J. Nutr. Biochem. 2017, 41, 65–72. [Google Scholar] [CrossRef]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA Methylation: A Review of Molecular Mechanisms and the Evidence for Folate’s Role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Ross, S.A.; Wise, C.; Pogribna, M.; Jones, E.A.; Tryndyak, V.P.; James, S.J.; Dragan, Y.P.; Poirier, L.A. Irreversible global DNA hypomethylation as a key step in hepatocarcinogenesis induced by dietary methyl deficiency. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2006, 593, 80–87. [Google Scholar] [CrossRef]
- Wiedeman, A.; Barr, S.; Green, T.; Xu, Z.; Innis, S.; Kitts, D. Dietary Choline Intake: Current State of Knowledge Across the Life Cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.; Ali, M. Methyl Donor Micronutrients that Modify DNA Methylation and Cancer Outcome. Nutrients 2019, 11, 608. [Google Scholar] [CrossRef]
- Zhao, G.; He, F.; Wu, C.; Li, P.; Li, N.; Deng, J.; Zhu, G.; Ren, W.; Peng, Y. Betaine in Inflammation: Mechanistic Aspects and Applications. Front. Immunol. 2018, 9, 1070. [Google Scholar] [CrossRef]
- Kennedy, D. B Vitamins and the Brain: Mechanisms, Dose and Efficacy—A Review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Cappuccilli, M.; Bergamini, C.; Giacomelli, F.A.; Cianciolo, G.; Donati, G.; Conte, D.; Natali, T.; La Manna, G.; Capelli, I. Vitamin B Supplementation and Nutritional Intake of Methyl Donors in Patients with Chronic Kidney Disease: A Critical Review of the Impact on Epigenetic Machinery. Nutrients 2020, 12, 1234. [Google Scholar] [CrossRef]
- Peracchu, M.; Bamonti, C.; Pomati, M.; De Franceschi, M.; Scalabrino, G. Human cobalamin deficiency: Alterations in serum tumor necrosis factor- α and epidermal growth factor. Eur. J. Haematol. 2001, 67, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Depeint, F.; Bruce, W.R.; Shangari, N.; Mehta, R.; O’Brien, P.J. Mitochondrial function and toxicity: Role of B vitamins on the one-carbon transfer pathways. Chem. Biol. Interact. 2006, 163, 113–132. [Google Scholar] [CrossRef]
- Lourdudoss, C.; Elkan, A.C.; Hafström, I.; Jogestrand, T.; Gustafsson, T.; van Vollenhoven, R.; Frostegård, J. Dietary micronutrient intake and atherosclerosis in systemic lupus erythematosus. Lupus 2016, 25, 1602–1609. [Google Scholar] [CrossRef]
- Suwannasom, N.; Kao, I.; Pruß, A.; Georgieva, R.; Bäumler, H. Riboflavin: The Health Benefits of a Forgotten Natural Vitamin. Int. J. Mol. Sci. 2020, 21, 950. [Google Scholar] [CrossRef] [PubMed]
- Wanders, D.; Hobson, K.; Ji, X. Methionine Restriction and Cancer Biology. Nutrients 2020, 12, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, M.; Ogura, Y.; Monno, I.; Xu, J.; Koya, D. Effect of Methionine Restriction on Aging: Its Relationship to Oxidative Stress. Biomedicines 2021, 9, 130. [Google Scholar] [CrossRef]
- Burdge, G.C.; Lillycrop, K.A. Fatty acids and epigenetics. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of Omega-3 Fatty Acids on Immune Cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef]
- Lu, S.C.; Huang, Z.Z.; Yang, H.; Mato, J.M.; Avila, M.A.; Tsukamoto, H. Changes in methionine adenosyltransferase and S -adenosylmethionine homeostasis in alcoholic rat liver. Am. J. Physiol.-Gastrointest. Liver Physiol. 2000, 279, G178–G185. [Google Scholar] [CrossRef]
- Vordenbäumen, S.; Sokolowski, A.; Rosenbaum, A.; Gebhard, C.; Raithel, J.; Düsing, C.; Chehab, G.; Richter, J.G.; Brinks, R.; Rehli, M.; et al. Methyl donor micronutrients, CD40-ligand methylation and disease activity in systemic lupus erythematosus: A cross-sectional association study. Lupus 2021, 30, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, B.; Kapoor, D.; Gautam, S.; Singh, R.; Bhardwaj, S. Dietary Polyunsaturated Fatty Acids (PUFAs): Uses and Potential Health Benefits. Curr. Nutr. Rep. 2021, 10, 232–242. [Google Scholar] [CrossRef]
- Islam, M.A.; Khandker, S.S.; Kotyla, P.J.; Hassan, R. Immunomodulatory Effects of Diet and Nutrients in Systemic Lupus Erythematosus (SLE): A Systematic Review. Front. Immunol. 2020, 11, 1477. [Google Scholar] [CrossRef]
- Stanger, O.; Wonisch, W. Enzymatic and Non-enzymatic Antioxidative Effects of Folic Acid and Its Reduced Derivates. In Water Soluble Vitamins; Stanger, O., Ed.; Part of the Subcellular Biochemistry Book Series (SCBI, Volume 56); Springer: Dordrecht, The Netherlands, 2012; pp. 131–161. [Google Scholar] [CrossRef]
- Moat, S.J.; Lang, D.; McDowell, I.F.W.; Clarke, Z.L.; Madhavan, A.K.; Lewis, M.J.; Goodfellow, J. Folate, homocysteine, endothelial function and cardiovascular disease. J. Nutr. Biochem. 2004, 15, 64–79. [Google Scholar] [CrossRef]
- Urowitz, M.B. The Bimodal Mortality Pattern of Systemic Lupus Erythematosus. Am. J. Med. 1976, 60, 221–225. [Google Scholar] [CrossRef]
- Salomão, R.G.; de Carvalho, L.M.; Izumi, C.; Czernisz, É.S.; Rosa, J.C.; Antonini, S.R.R.; Bueno, A.C.; Almada, M.O.R.D.V.; de Coelho-Landell, C.A.; Jordão, A.A.; et al. Homocysteine, folate, hs-C-reactive protein, tumor necrosis factor-alpha and inflammatory proteins: Are these biomarkers related to nutritional status and cardiovascular risk in childhood-onset systemic lupus erythematosus? Pediatr. Rheumatol. 2018, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Pocovi-Gerardino, G.; Correa-Rodríguez, M.; Callejas-Rubio, J.L.; Ríos-Fernández, R.; Ortego-Centeno, N.; Rueda-Medina, B. Dietary intake and nutritional status in patients with systemic lupus erythematosus. Endocrinol. Diabetes Y Nutr. 2018, 65, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Qiang, Y.; Li, Q.; Xin, Y.; Fang, X.; Tian, Y.; Ma, J.; Wang, J.; Wang, Q.; Zhang, R.; Wang, J.; et al. Intake of Dietary One-Carbon Metabolism-Related B Vitamins and the Risk of Esophageal Cancer: A Dose-Response Meta-Analysis. Nutrients 2018, 10, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Epigenetic Mechanisms | Change in SLE | Outcomes | References |
|---|---|---|---|
| DNA methylation | Hypomethylation of CD40L promotor in CD40+ T cells | Association with clinical disease activity | Vordenbäumen et al., 2021 [17] |
| Hypomethylation of IL-10 and IL-13 gene regulatory domains | Overexpression of IL-10 and IL-13 | Zhao et al., 2010 [18] | |
| ncRNAs | Overexpression of miR-29b | Reduction of DNMT1 levels, DNA hypomethylation, and upregulation of genes encoding CD11 and CD70 | Qin et al., 2013 [19] |
| Overexpression of miR-126 in CD4+ T cells | Contribution to T cell autoreactivity by targeting DNMT1 | Zhao et al., 2011 [20] | |
| Reduced expression of miR-142-3p/5p in CD4+ T cells | T cell hyperactivity and B cell hyperstimulation | Ding et al., 2020 [21] | |
| Histone modifications | Elevated histone H3 acetylation in CD4+ T cells | Correlation with clinical disease activity | Zhou et al., 2011 [22] |
| High methylation in the HDAC6 promoter | Lower HDAC6 levels and SLE susceptibility | Fang et al., 2016 [23] |
| Dietary Compound | Population/Tissue | Study Design | Outcomes | References |
|---|---|---|---|---|
| Folate | Colorectal adenoma patients | Randomized controlled trial 400 μg/day for 10 weeks | Increase DNA methylation of 31% in leukocytes and 25% in colonic mucosa | Lu et al., 2006 [31] |
| Postmenopausal women | Randomized controlled trial | DNA hypomethylation was reversed with high folate intake (286–516 μg/day) | Jacob et al.,1998 [32] | |
| Choline/betaine | Rat fetal liver and brain | Experimental study Rats fed with several choline doses | Maternal choline supply modifies fetal histone and DNA methylation. | Davison et al., 2009 [33] |
| Cobalamin (B12) | Wistar rats | Experimental study Rats with and without the absence of B12 | Cobalamin deprivation-induced global hypomethylation | Kulkami et al., 2011 [34] |
| Methionine | Women of reproductive age | Observational cohort study | High intake of methionine in pre-pregnancy reduced hydroxymethylation | Pauwels et al., 2016 [35] |
| Fatty acids | Human colorectal cancer cells | In vitro study Cells treated with 100 μM of DHA, EPA, and LA for 6 days | Increase the expression of DNMTs in human colorectal cancer cell line HT29/219 | Sarabi et al., 2018 [36] |
| Dietary Compound | DRI | Food Source |
|---|---|---|
| Folate | 400 µg DFE/day | Green vegetables, sweet peppers, legumes, oranges, eggs, red meat |
| Choline | 400 mg/day | Beef liver, egg, soybeans, potatoes, wheat germ, quinoa, peanuts, carrots, apples, broccoli |
| Riboflavin (B2) | 1.1 mg/day | Oats, quinoa, apple, spinach, tomatoes, rice, salmon |
| Pyridoxine (B6) | 1.3 mg/day | Chickpeas, potatoes, salmon, tuna, cottage cheese, onions, rice, nuts, watermelon |
| Cobalamin (B12) | 2.4 µg/day | Milk, beef liver, beans, strawberry, banana, spinach, salmon, tuna, yogurt, cheese |
| Methionine | - | Eggs, yogurt, cheese, red meat, soja, milk |
| Omega 3 fatty acids | 1.1 g/day | Chia seeds, sardines, whole bread, milk, beans, salmon, soybean, canola oils, flaxseed |
| Omega 6 fatty acids | - | Olive oil, safflower, sunflowers oils, peanut oil |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pesqueda-Cendejas, K.; Campos-López, B.; Mora-García, P.E.; Moreno-Ortiz, J.M.; De la Cruz-Mosso, U. Methyl Donor Micronutrients: A Potential Dietary Epigenetic Target in Systemic Lupus Erythematosus Patients. Int. J. Mol. Sci. 2023, 24, 3171. https://doi.org/10.3390/ijms24043171
Pesqueda-Cendejas K, Campos-López B, Mora-García PE, Moreno-Ortiz JM, De la Cruz-Mosso U. Methyl Donor Micronutrients: A Potential Dietary Epigenetic Target in Systemic Lupus Erythematosus Patients. International Journal of Molecular Sciences. 2023; 24(4):3171. https://doi.org/10.3390/ijms24043171
Chicago/Turabian StylePesqueda-Cendejas, Karen, Bertha Campos-López, Paulina E. Mora-García, José M. Moreno-Ortiz, and Ulises De la Cruz-Mosso. 2023. "Methyl Donor Micronutrients: A Potential Dietary Epigenetic Target in Systemic Lupus Erythematosus Patients" International Journal of Molecular Sciences 24, no. 4: 3171. https://doi.org/10.3390/ijms24043171
APA StylePesqueda-Cendejas, K., Campos-López, B., Mora-García, P. E., Moreno-Ortiz, J. M., & De la Cruz-Mosso, U. (2023). Methyl Donor Micronutrients: A Potential Dietary Epigenetic Target in Systemic Lupus Erythematosus Patients. International Journal of Molecular Sciences, 24(4), 3171. https://doi.org/10.3390/ijms24043171