


Flavonoids from Hippophae rhamnoides Linn. Revert Doxorubicin-Induced Cardiotoxicity through Inhibition of Mitochondrial Dysfunction in H9c2 Cardiomyoblasts In Vitro


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
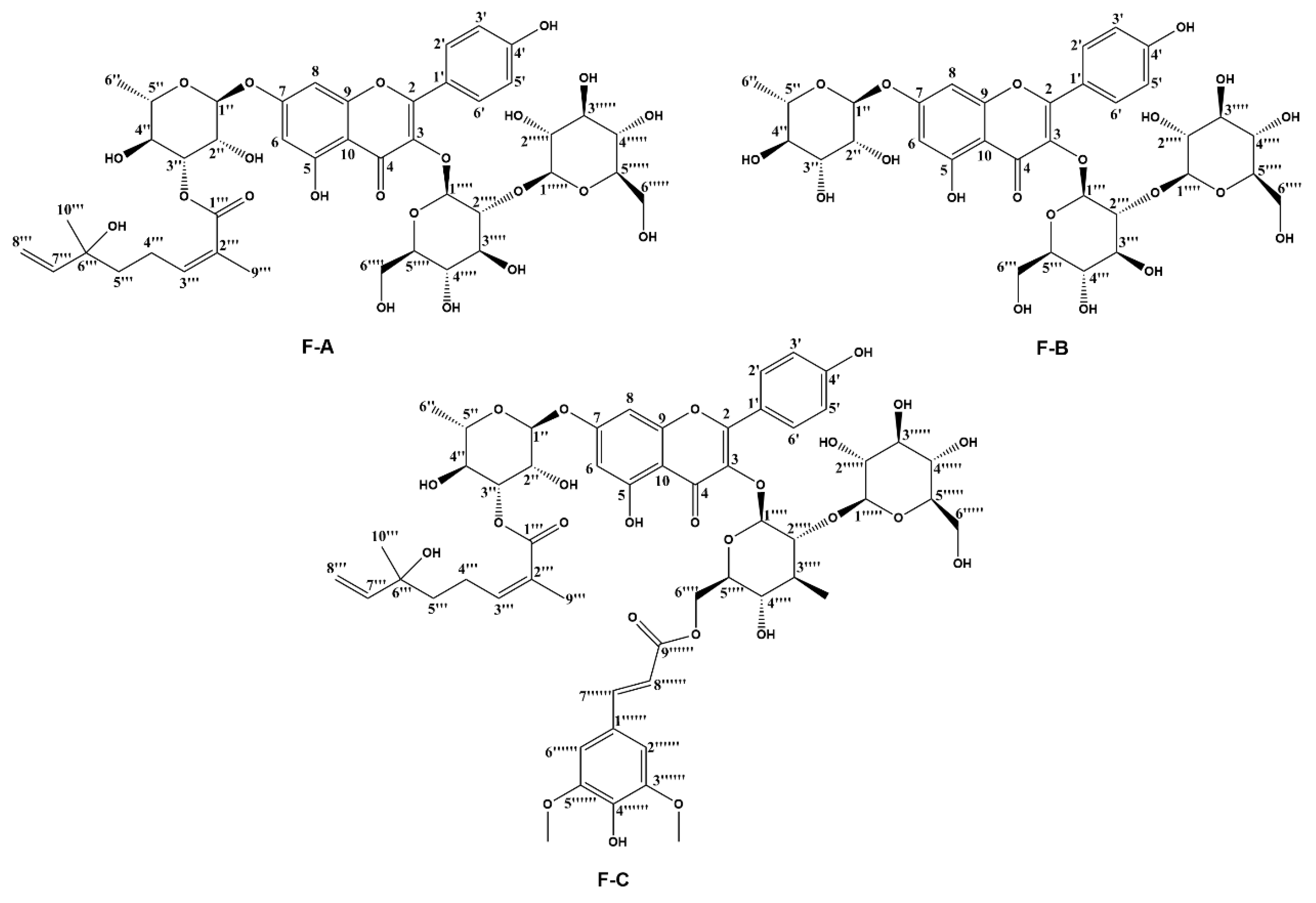
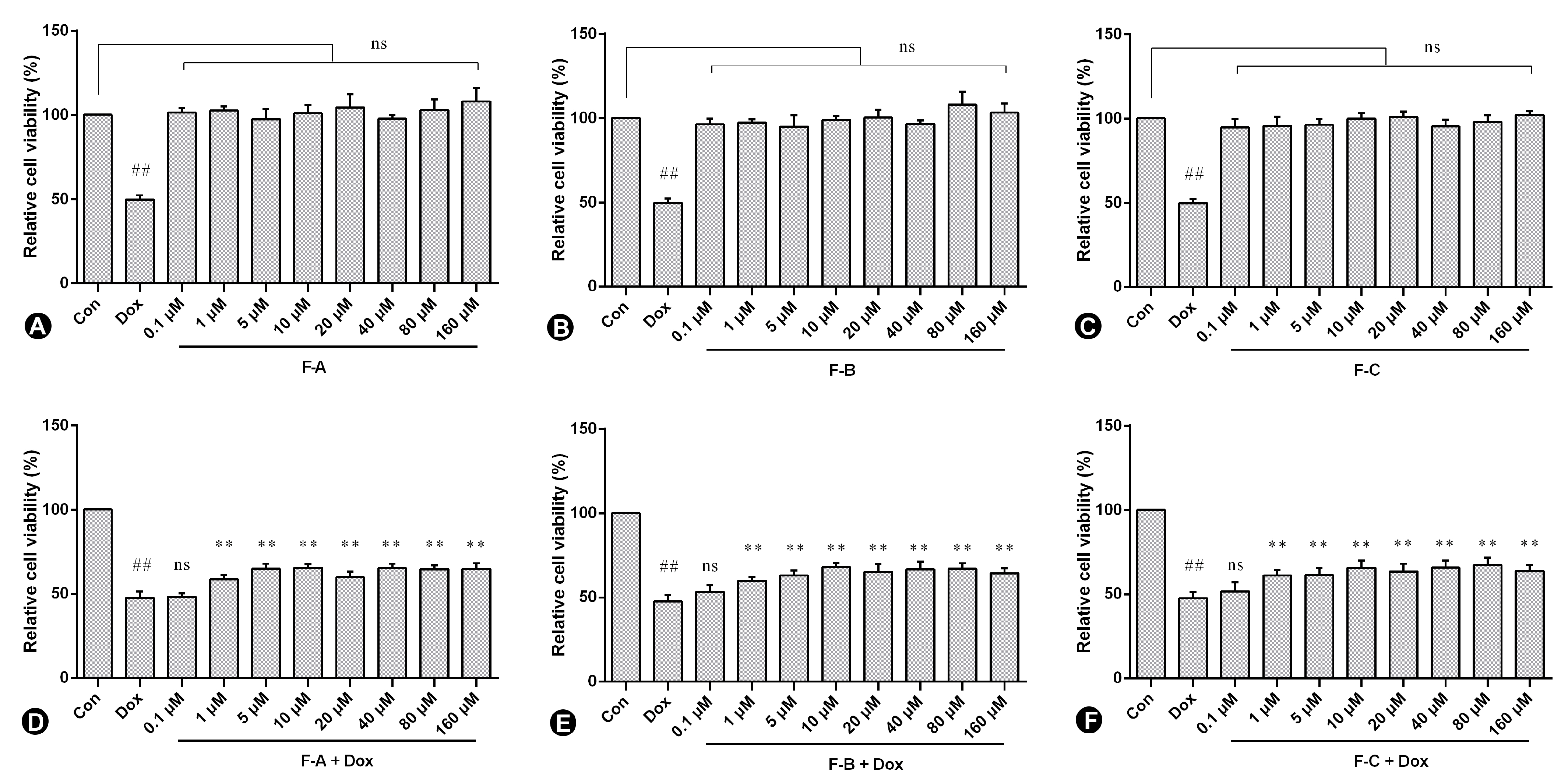
2.1. Flavonoids from Hippophae rhamnoides Linn. Inhibits Dox-Induced Cytotoxicity in H9c2 Cells
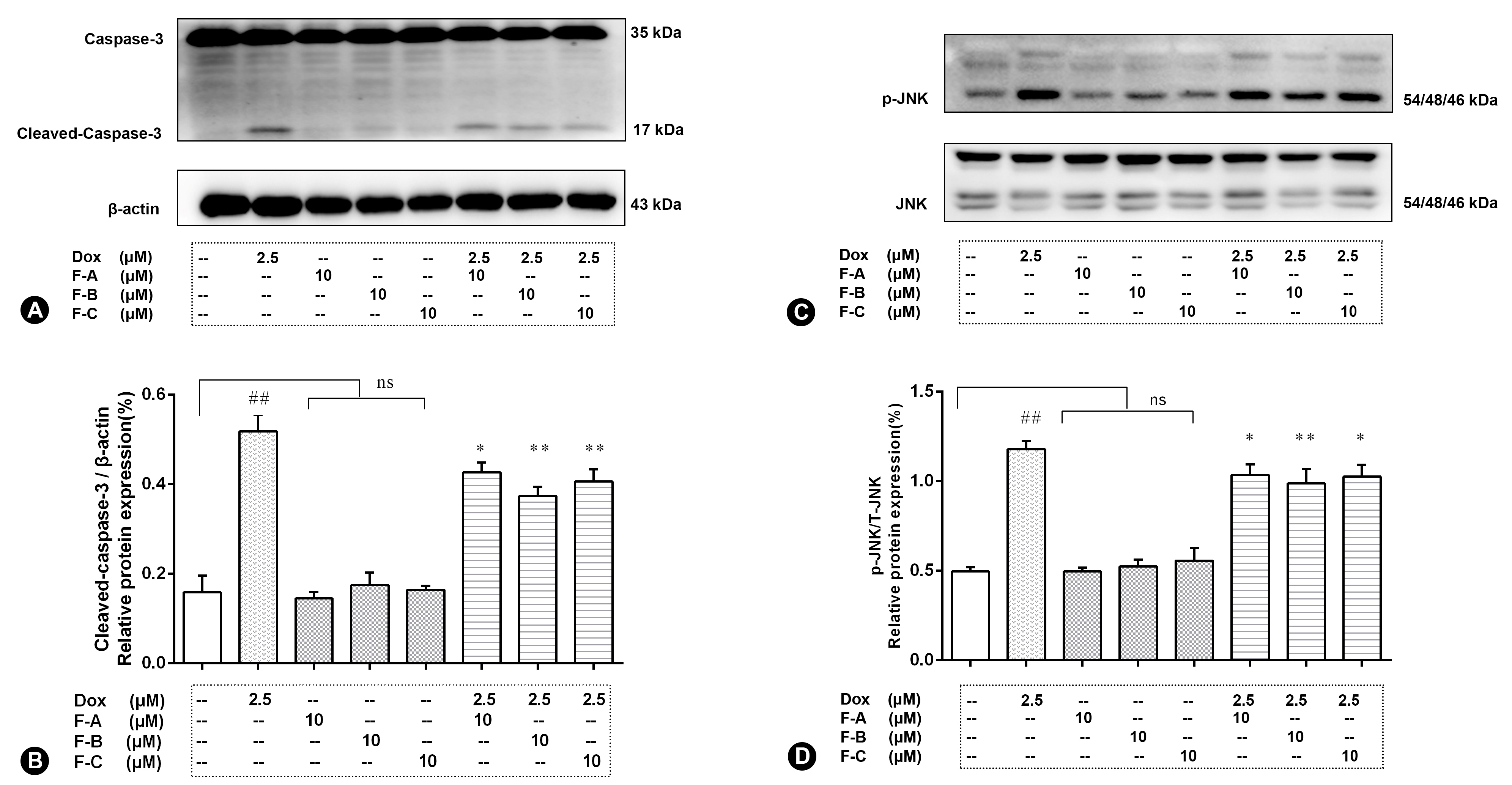
2.2. Flavonoids Significantly Decrease Dox-Induced Cell Apoptosis
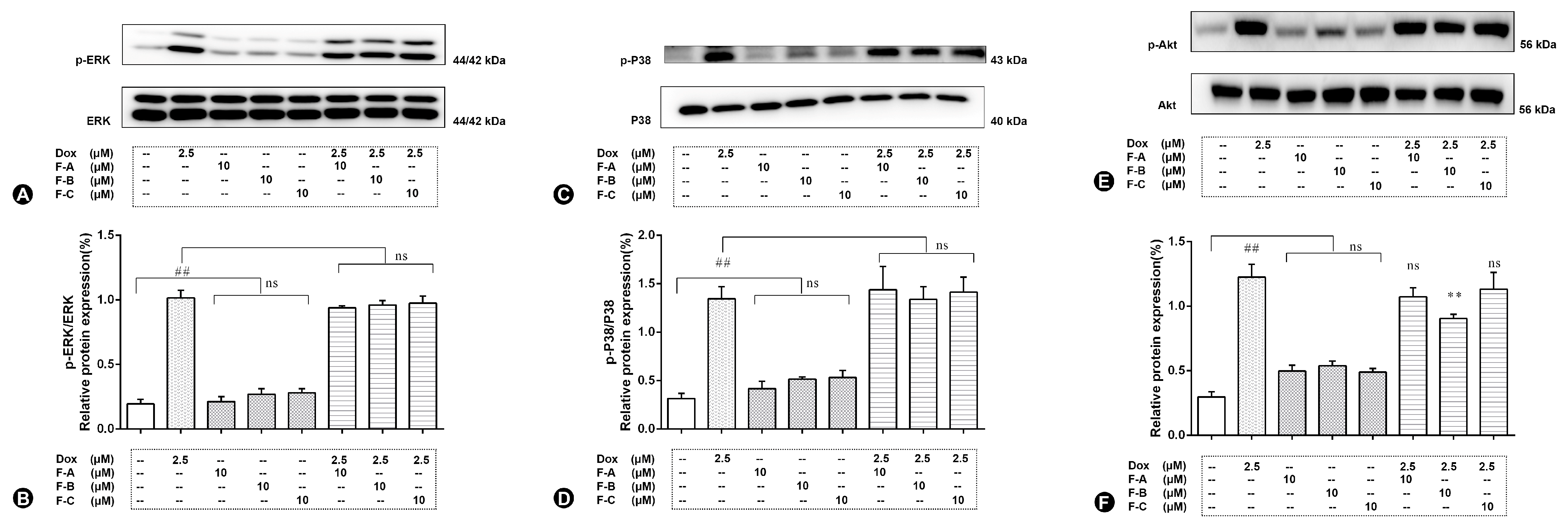
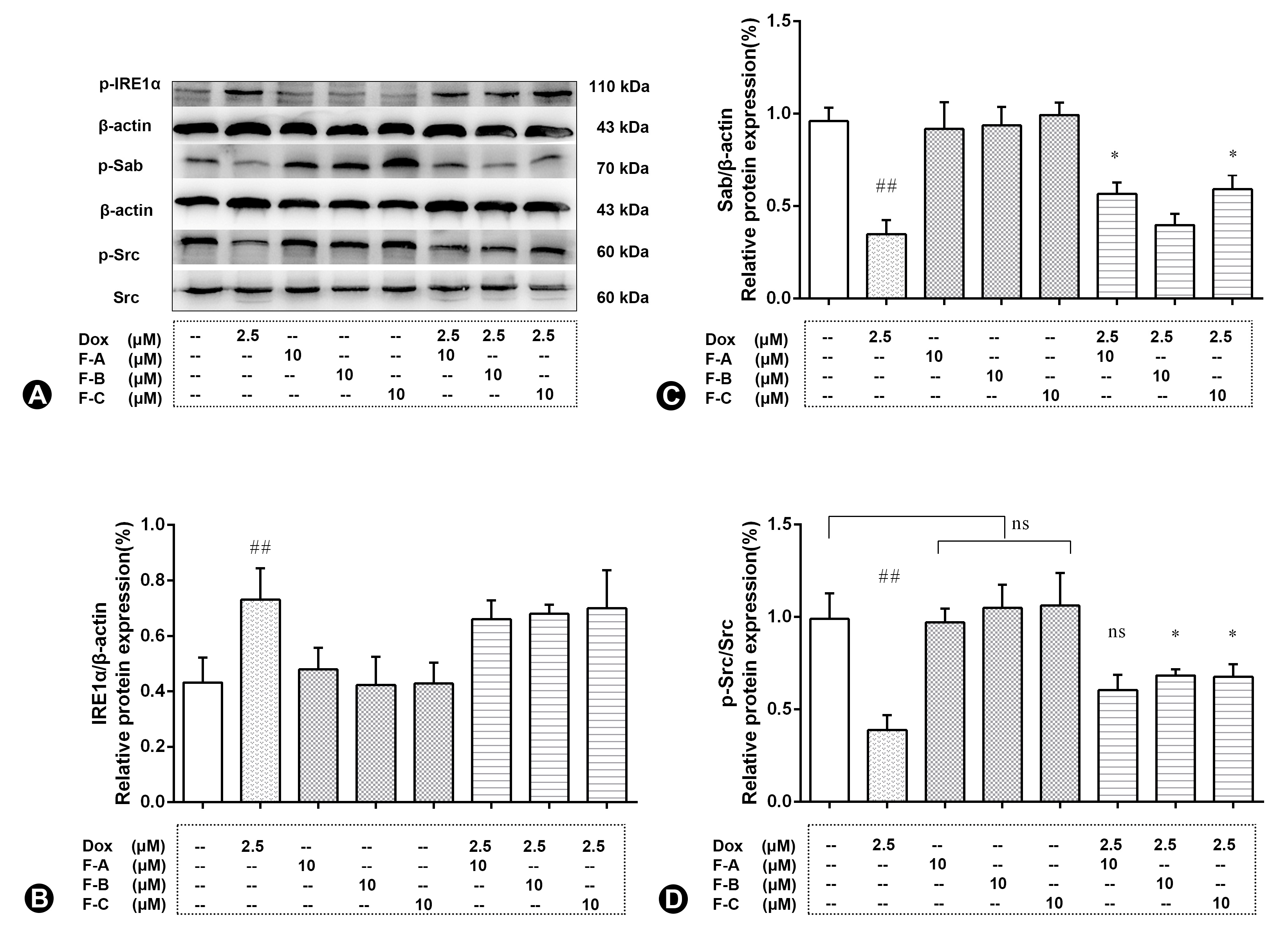
2.3. Flavonoids Inhibit JNK Activation in Dox-Treated H9c2 Cells
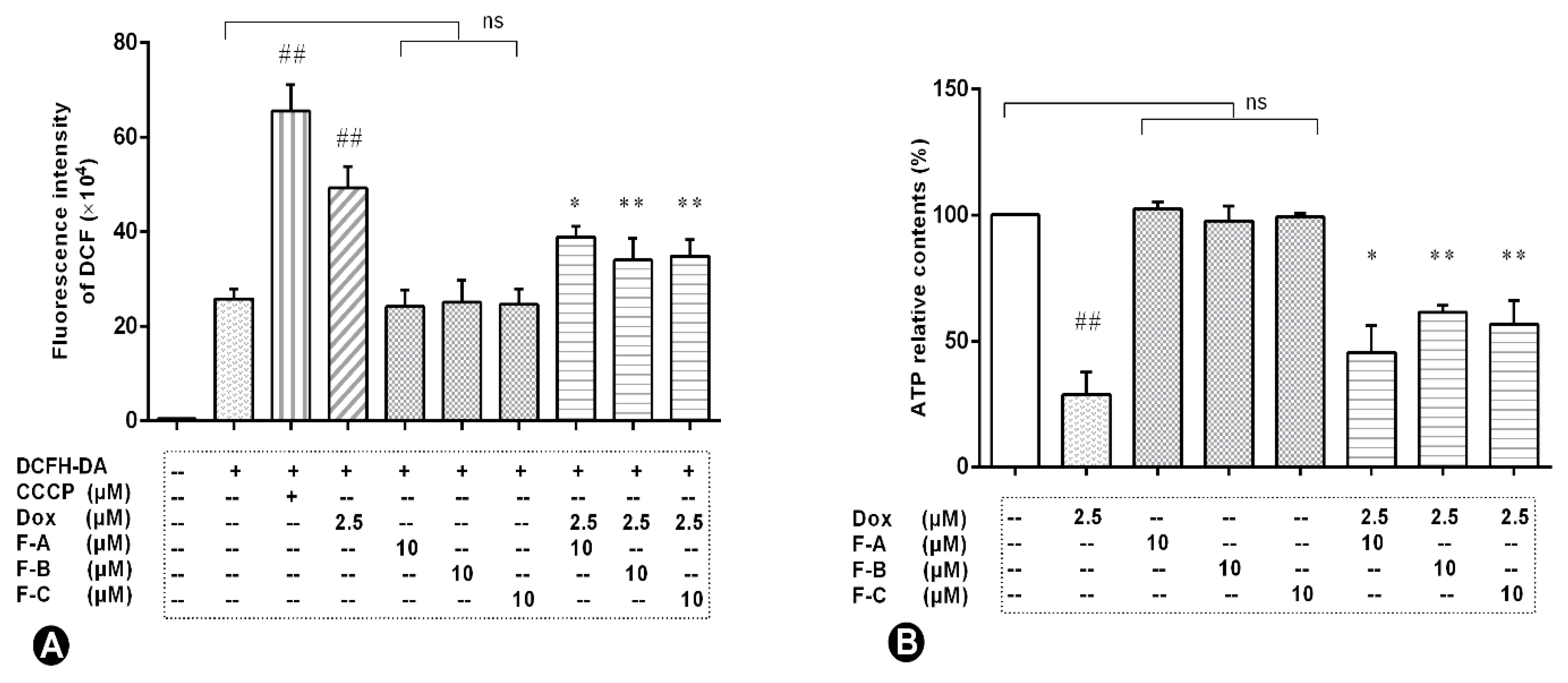
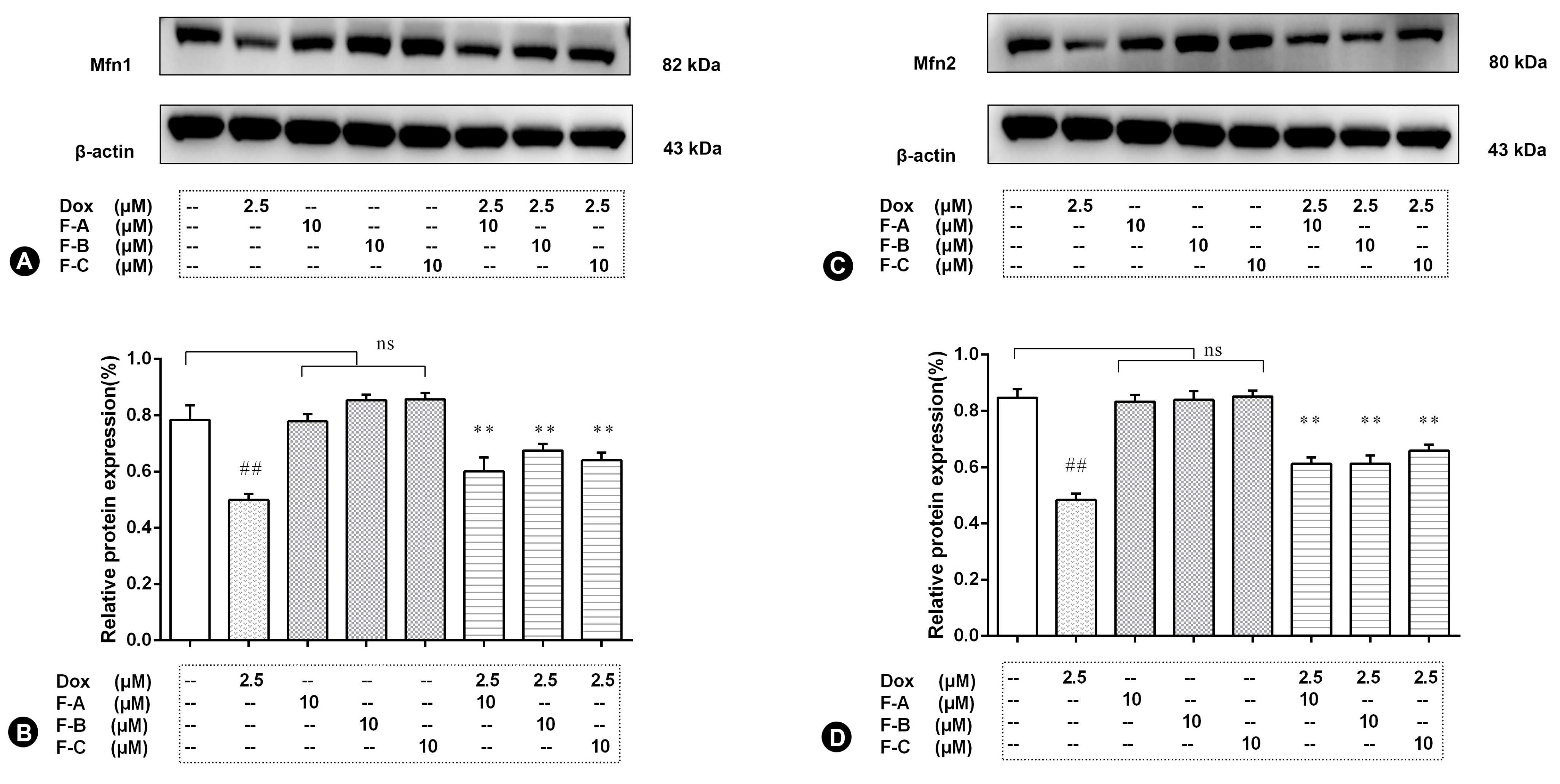
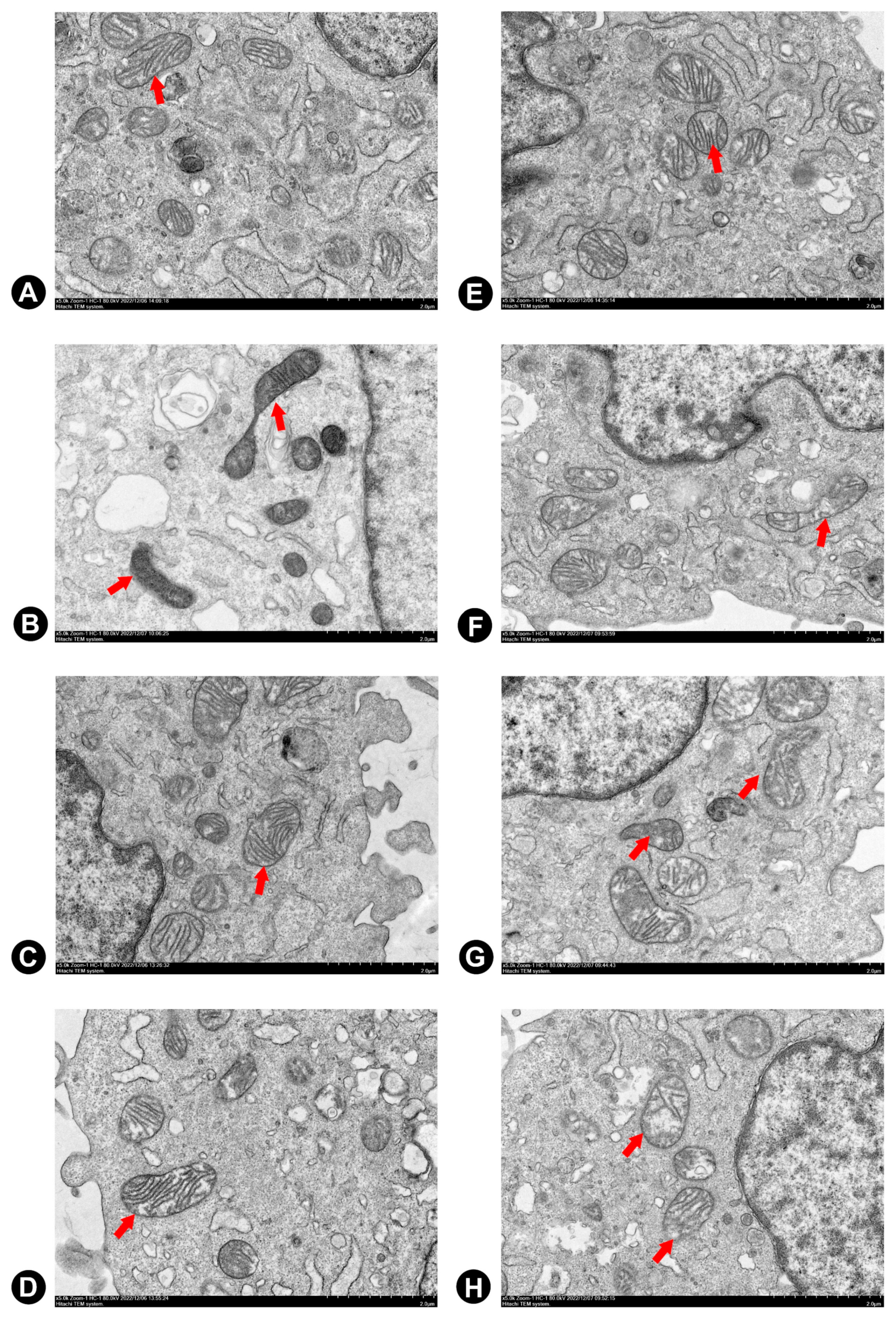
2.4. Flavonoids Effectively Improved Dox-Induced Mitochondrial Dysfunction in H9c2 Cells
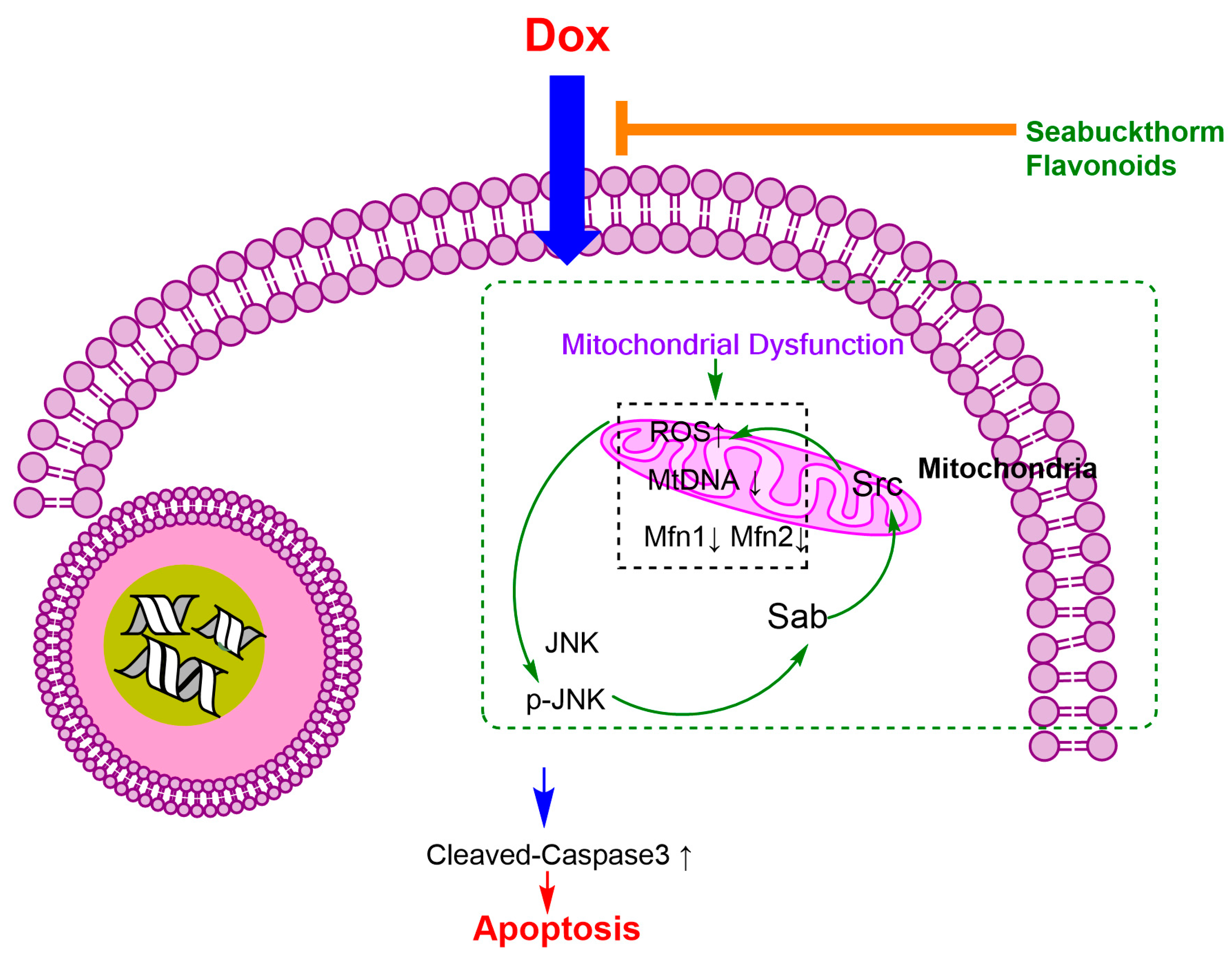
2.5. Flavonoids Based on the “JNK-Sab-ROS” Pathway Effectively Alleviate Dox-Induced Cardiotoxicity
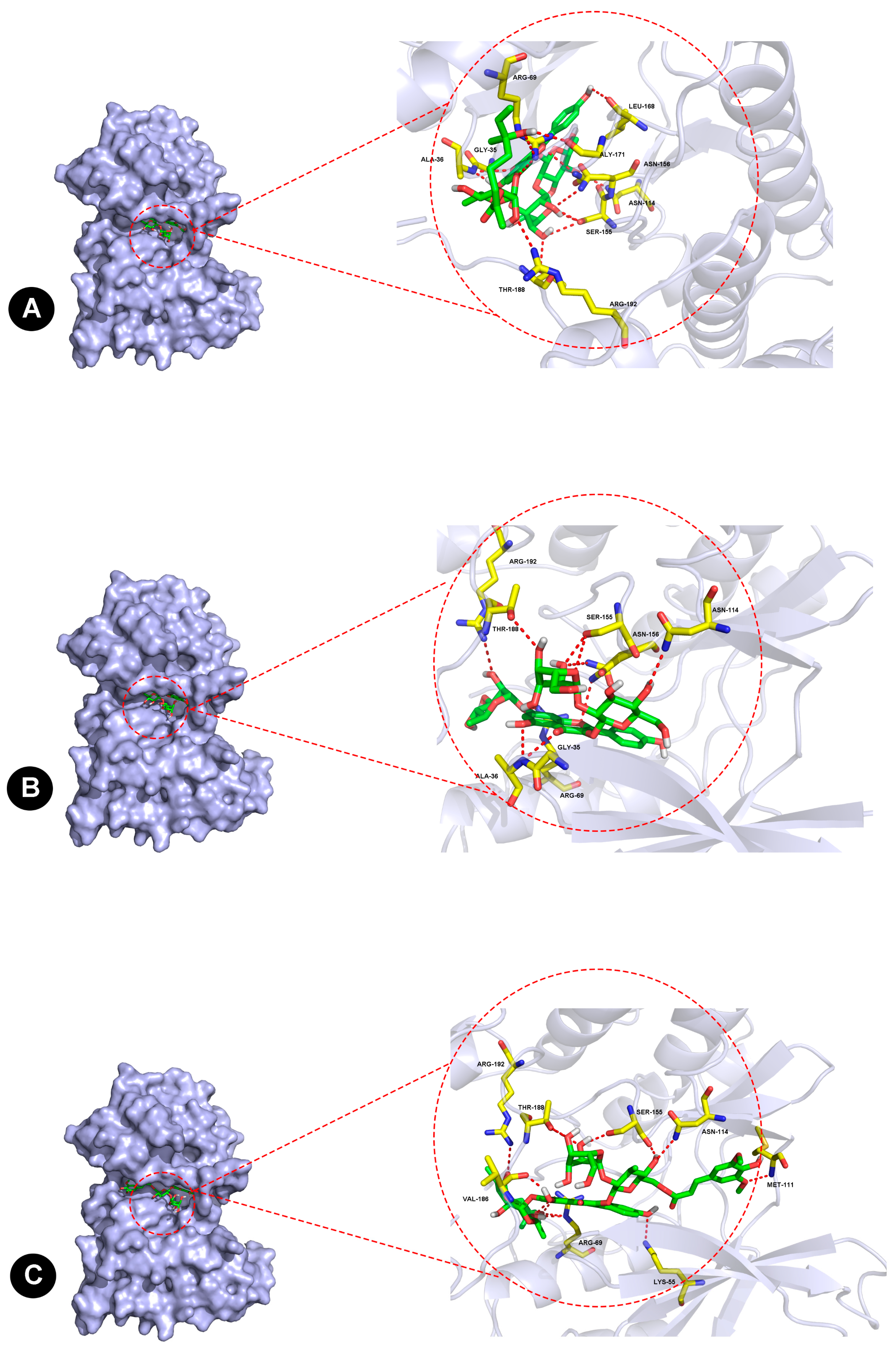
2.6. Molecular Docking of JNK1 and Three Flavonoids
3. Discussion
4. Materials and Methods
4.1. Preparation of Purified Compounds
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Determination of Intracellular ROS Generation
4.5. ATP Content Measurement
4.6. Transmission Electron Microscopy (TEM)
4.7. Western Blot Analysis
4.8. Molecular Docking
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sritharan, S.; Sivalingam, N.A. Comprehensive review on time-tested anticancer drug doxorubicin. Life Sci. 2021, 278, 119527. [Google Scholar] [CrossRef] [PubMed]
- Sheibani, M.; Azizi, Y.; Shayan, M.; Nezamoleslami, S.; Eslami, F.; Farjoo, M.H.; Dehpour, A.R. Doxorubicin-Induced Cardiotoxicity: An Overview on Pre-clinical Therapeutic Approaches. Cardiovasc. Toxicol. 2022, 22, 292–310. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.; Santos, R.X.; Cardoso, S.; Correia, S.; Oliveira, P.J.; Santos, M.S.; Moreira, P.I. Doxorubicin: The good, the bad and the ugly effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Eikenberry, S. A tumor cord model for doxorubicin delivery and dose optimization in solid tumors. Theor. Biol. Med. Model. 2009, 6, 16. [Google Scholar] [CrossRef]
- Govender, Y.J. Mitochondrial catastrophe during doxorubicin-induced cardiotoxicity: An evaluation of the protective role of melatonin. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2017. Available online: http://hdl.handle.net/10019.1/101150 (accessed on 31 March 2017).
- Kabir, S.; Lingappa, N.; Mayrovitz, H. Potential Therapeutic Treatments for Doxorubicin-Induced Cardiomyopathy. Cureus 2022, 14, e21154. [Google Scholar] [CrossRef]
- Octavia, Y.; Tocchetti, C.G.; Gabrielson, K.L.; Janssens, S.; Crijns, H.J.; Moens, A.L. Doxorubicin-induced cardiomyopathy: From molecular mechanisms to therapeutic strategies. J. Mol. Cell. Cardiol. 2012, 52, 1213–1225. [Google Scholar] [CrossRef]
- Li, Z.; Chinnathambi, A.; Ali Alharbi, S.; Yin, F. Plumbagin protects the myocardial damage by modulating the cardiac biomarkers, antioxidants, and apoptosis signaling in the doxorubicin-induced cardiotoxicity in rats. Environ. Toxicol. 2020, 35, 1374–1385. [Google Scholar] [CrossRef]
- Archana, V.; Indumathy, R. In-vitro Antioxidant and Cardio-protective effect of Delonix elata (L.) Leaf extract against Doxorubicin induced toxicity in H9c2 Cardio-myocyte cell line. Res. J. Pharm. Technol. 2021, 14, 5635–5641. [Google Scholar] [CrossRef]
- Monahan, D.S.; Flaherty, E.; Hameed, A.; Duffy, G.P. Resveratrol significantly improves cell survival in comparison to dexrazoxane and carvedilol in a h9c2 model of doxorubicin induced cardiotoxicity. Biomed. Pharmacother. 2021, 140, 111702. [Google Scholar] [CrossRef]
- Rossini, M.; Filadi, R. Sarcoplasmic reticulum-mitochondria kissing in cardiomyocytes: Ca2+, ATP, and undisclosed secrets. Front. Cell Dev. Biol. 2020, 8, 532. [Google Scholar] [CrossRef]
- Carvalho, F.S.; Burgeiro, A.; Garcia, R.; Moreno, A.J.; Carvalho, R.A.; Oliveira, P.J. Doxorubicin-induced cardiotoxicity: From bioenergetic failure and cell death to cardiomyopathy. Med. Res. Rev. 2014, 34, 106–135. [Google Scholar] [CrossRef] [PubMed]
- Kawalec, P.; Martens, M.D.; Field, J.T.; Mughal, W.; Caymo, A.M.; Chapman, D.; Xiang, B.; Ghavami, S.; Dolinsky, V.W.; Gordon, J. Differential Impact of Doxorubicin Dose on Cell Death and Autophagy Pathways during Acute Cardiotoxicity. Toxicol. Appl. Pharmacol. 2022, 453, 116210. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.B.; Leung, K.T.; Poon, E.N.-Y. Mitochondrial-Targeted Therapy for Doxorubicin-Induced Cardiotoxicity. Int. J. Mol. Sci. 2022, 23, 1912. [Google Scholar] [CrossRef]
- Shuaifeng, C.; Xuming, Y. Clinical effect of Tibetan medicine Tongxinshu Capsule combined with Nicodil on angina pectoris of coronary heart disease. J. Med. Phar. Chin. Minor. 2021, 27, 17–18. [Google Scholar] [CrossRef]
- Gulimire, A.; Ybadaiti, T.; Rena, K.; Ting, F. Protective effect of total flavonoids of H. rhamnoides L. sunsp. turkestanica Rousi against adriamycin-induced cardiotoxicity in rats. J. Xinjiang Med. Univ. 2010, 33, 383–385. [Google Scholar] [CrossRef]
- Sun, B.; Sun, G.B.; Xiao, J.; Chen, R.C.; Wang, X.; Wu, Y.; Cao, L.; Yang, Z.H.; Sun, X.B. Isorhamnetin inhibits H2O2-induced activation of the intrinsic apoptotic pathway in H9c2 cardiomyocytes through scavenging reactive oxygen species and ERK inactivation. J. Cell. Biochem. 2012, 113, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Sun, G.; Meng, X.; Wang, H.; Luo, Y.; Qin, M.; Ma, B.; Wang, M.; Cai, D.; Guo, P. Isorhamnetin protects against doxorubicin-induced cardiotoxicity in vivo and in vitro. PLoS ONE 2013, 8, e64526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Yuan, Z.; Li, G.; Ouyang, J.; Suo, Y.; Wang, H. Isolation and structure determination of a new flavone glycoside from seed residues of seabuckthorn (Hippophae rhamnoides L.). Nat. Prod. Res. 2018, 32, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ouyang, J.; Hu, N.; Li, G.; Wang, H. Protective effect of two alkaloids from Hippophae rhamnoides Linn. Against doxorubicin-induced toxicity in H9c2 Cardiomyoblasts. Molecules 2021, 26, 1946. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.Y.; Xu, D.P. Extraction, Isolation and Protective Effect of Alkaloid from Seabuckthorn Seeds on Injured Cardiomyocytes in Rats. Food Sci. 2010, 4, 234–237. [Google Scholar] [CrossRef]
- Li, G.; Chu, M.; Tong, Y.; Liang, Y.; Wang, S.; Ma, C.J.; Wang, Z.H.; Zhou, W.N. Protective Effects of Hippophae rhamnoides L. Phenylpropanoids on Doxorubicin-Induced Cardiotoxicity in Zebrafish. Molecules 2022, 27, 8858. [Google Scholar] [CrossRef] [PubMed]
- Takemura, G.; Fujiwara, H. Doxorubicin-induced cardiomyopathy: From the cardiotoxic mechanisms to management. Prog. Cardiovasc. Dis. 2007, 49, 330–352. [Google Scholar] [CrossRef] [PubMed]
- DU, Y.-W. Protective effect and mechanism of total flavonoids of Dracocephalum moldevica on doxorubicin myocardial toxicity. Chin Tradit. Herbal Drugs 2019, 50, 6045–6051. [Google Scholar] [CrossRef]
- Ouyang, J. Preparation of Active Components from the Seed Residue of Hippophae rhamnoides Linn. and Studies of the Mechanism on Type II Diabetes; Northwest Institute of Plateau Biology, University of Chinese Academy of Sciences: Xining, China, 2015; pp. 48–49. [Google Scholar]
- Dikalov, S.I.; Ungvari, Z. Role of mitochondrial oxidative stress in hypertension. Am. J. Physiol.-Heart C 2013, 305, H1417–H1427. [Google Scholar] [CrossRef]
- Marin-Garcia, J.; Goldenthal, M.J.; Moe, G.W. Mitochondrial pathology in cardiac failure. Cardiovasc. Res. 2001, 49, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, M.P.; Weiner, A.; Kaminaris, A.; Li, C.; Cai, F.; Zhao, F.; Kobayashi, S.; Kobayashi, T.; Huang, Y.; Sesaki, H. Doxorubicin-induced cardiomyocyte death is mediated by unchecked mitochondrial fission and mitophagy. Faseb J. 2019, 33, 11096–11108. [Google Scholar] [CrossRef]
- Chen, M.-B.; Wu, X.-Y.; Gu, J.-H.; Guo, Q.-T.; Shen, W.-X.; Lu, P.-H. Activation of AMP-activated protein kinase contributes to doxorubicin-induced cell death and apoptosis in cultured myocardial H9c2 cells. Cell Biochem. Biophys. 2011, 60, 311–322. [Google Scholar] [CrossRef]
- Yong, S.; Yong-Kwan, C.; Nam-Ho, K.; Hun-Taeg, C.; Kang, D.G.; Hyun-Ock, P. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J. Sig. Transd. 2011, 2011, 792639. [Google Scholar] [CrossRef]
- Brantley-Finley, C.; Lyle, C.S.; Du, L.; Goodwin, M.E.; Hall, T.; Szwedo, D.; Kaushal, G.P.; Chambers, T.C. The JNK, ERK and p53 pathways play distinct roles in apoptosis mediated by the antitumor agents vinblastine, doxorubicin, and etoposide. Biochem. Pharmacol. 2003, 66, 459–469. [Google Scholar] [CrossRef]
- Hirata, Y.; Inoue, A.; Suzuki, S.; Takahashi, M.; Matsui, R.; Kono, N.; Noguchi, T.; Matsuzawa, A. Trans-Fatty acids facilitate DNA damage-induced apoptosis through the mitochondrial JNK-Sab-ROS positive feedback loop. Sci. Rep. 2020, 10, 2743. [Google Scholar] [CrossRef] [Green Version]
- Tie, F.F.; Luan, G.X.; Zhou, W.N.; Wang, Z.H.; Shi, X.B.; Li, G.; Wang, H.L. Effects of the oligostilbenes from Iris lactea Pall. var. chinensis (Fisch.) Koidz on the adipocytes differentiation of 3T3-L1 cells. Pharmazie 2018, 73, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Luan, G.; Wang, Y.; Wang, Z.; Zhou, W.; Hu, N.; Li, G.; Wang, H. Flavonoid Glycosides from Fenugreek Seeds Regulate Glycolipid Metabolism by Improving Mitochondria! Function in 3T3-L1 Adipocytes In Vitro. J. Agric. Food Chem. 2018, 66, 3169–3178. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Han, L.; Tie, F.; Wang, Z.; Ma, C.; Li, J.; Wang, H.; Li, G. (20S)-Protopanaxadiol Ginsenosides Induced Cytotoxicity via Blockade of Autophagic Flux in HGC-27 Cells. Chem. Biodivers. 2020, 17, e2000187. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, W.; Ouyang, J.; Hu, N.; Wang, H. Flavonoids from Hippophae rhamnoides Linn. Revert Doxorubicin-Induced Cardiotoxicity through Inhibition of Mitochondrial Dysfunction in H9c2 Cardiomyoblasts In Vitro. Int. J. Mol. Sci. 2023, 24, 3174. https://doi.org/10.3390/ijms24043174
Zhou W, Ouyang J, Hu N, Wang H. Flavonoids from Hippophae rhamnoides Linn. Revert Doxorubicin-Induced Cardiotoxicity through Inhibition of Mitochondrial Dysfunction in H9c2 Cardiomyoblasts In Vitro. International Journal of Molecular Sciences. 2023; 24(4):3174. https://doi.org/10.3390/ijms24043174
Chicago/Turabian StyleZhou, Wenna, Jian Ouyang, Na Hu, and Honglun Wang. 2023. "Flavonoids from Hippophae rhamnoides Linn. Revert Doxorubicin-Induced Cardiotoxicity through Inhibition of Mitochondrial Dysfunction in H9c2 Cardiomyoblasts In Vitro" International Journal of Molecular Sciences 24, no. 4: 3174. https://doi.org/10.3390/ijms24043174
APA StyleZhou, W., Ouyang, J., Hu, N., & Wang, H. (2023). Flavonoids from Hippophae rhamnoides Linn. Revert Doxorubicin-Induced Cardiotoxicity through Inhibition of Mitochondrial Dysfunction in H9c2 Cardiomyoblasts In Vitro. International Journal of Molecular Sciences, 24(4), 3174. https://doi.org/10.3390/ijms24043174