
Regulation of Inflammasome by microRNAs in Triple-Negative Breast Cancer: New Opportunities for Therapy



Abstract
:1. Introduction
2. Molecular Characteristics of TNBC
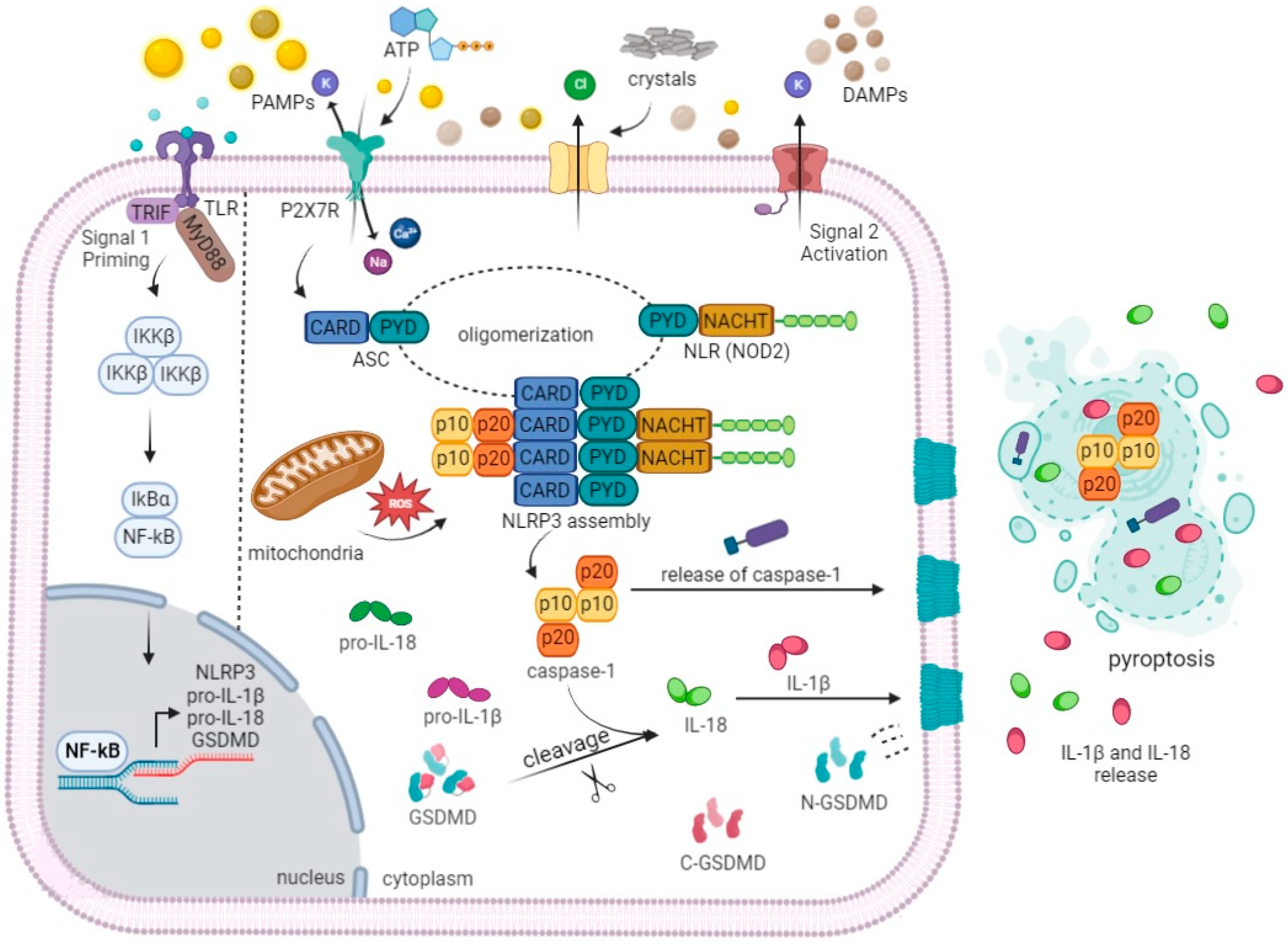
3. Molecular Mechanisms of Inflammasome Activation during TNBC
4. Non-Coding RNAs in Oncology
5. MicroRNAs Interplay with TNBC and NLRP3
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, N.A.J.; Tirona, M. An updated review of epidemiology, risk factors, and management of male breast cancer. Med. Oncol. 2021, 38, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Oskar, S.; Kehm, R.; Terry, M.B. Breast Tissue Composition—Why It Matters and How Can We Measure It More Accurately in Epidemiology Studies. Cancer Epidemiol. Biomarkers Prev. 2021, 30, 590–592. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Wang, L. Early Diagnosis of Breast Cancer. Sensors 2017, 17, 1572. [Google Scholar] [CrossRef] [PubMed]
- Teichgraeber, D.C.; Guirguis, M.S.; Whitman, G.J. Breast Cancer Staging: Updates in the AJCC Cancer Staging Manual, 8th Edition, and Current Challenges for Radiologists, From the AJR Special Series on Cancer Staging. Am. J. Roentgenol. 2021, 217, 278–290. [Google Scholar] [CrossRef]
- Kalli, S.; Semine, A.; Cohen, S.; Naber, S.P.; Makim, S.S.; Bahl, M. American Joint Committee on Cancer’s Staging System for Breast Cancer, Eighth Edition: What the Radiologist Needs to Know. Radiographics 2018, 38, 1921–1933. [Google Scholar] [CrossRef]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2019, 27, 27–35. [Google Scholar] [CrossRef]
- Kumar, P.; Aggarwal, R. An overview of triple-negative breast cancer. Arch. Gynecol. Obstet. 2016, 293, 247–269. [Google Scholar] [CrossRef]
- Jackson, H.W.; Fischer, J.R.; Zanotelli, V.R.T.; Ali, H.R.; Mechera, R.; Soysal, S.D.; Moch, H.; Muenst, S.; Varga, Z.; Weber, W.P.; et al. The single-cell pathology landscape of breast cancer. Nature 2020, 578, 615–620. [Google Scholar] [CrossRef]
- Yeo, S.K.; Guan, J.-L. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef]
- Sukumar, J.; Gast, K.; Quiroga, D.; Lustberg, M.; Williams, N. Triple-negative breast cancer: Promising prognostic biomarkers currently in development. Expert Rev. Anticancer. Ther. 2020, 21, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, C.; Scarpi, E.; Sonnessa, M.; Cioffi, A.; Buccino, F.; Giotta, F.; Pastena, M.I.; Zito, F.A.; Mangia, A. Prognostic Value of NLRP3 Inflammasome and TLR4 Expression in Breast Cancer Patients. Front. Oncol. 2021, 11, 705331. [Google Scholar] [CrossRef] [PubMed]
- Faria, S.S.; Costantini, S.; de Lima, V.C.C.; de Andrade, V.P.; Rialland, M.; Cedric, R.; Budillon, A.; Magalhaes, K.G. NLRP3 inflammasome-mediated cytokine production and pyroptosis cell death in breast cancer. J. Biomed. Sci. 2021, 28, 26. [Google Scholar] [CrossRef] [PubMed]
- Tezcan, G.; Martynova, E.V.; Gilazieva, Z.E.; McIntyre, A.; Rizvanov, A.A.; Khaiboullina, S.F. MicroRNA Post-transcriptional Regulation of the NLRP3 Inflammasome in Immunopathologies. Front. Pharmacol. 2019, 10, 451. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, G.; Liu, X.-L.; Zhang, G.; Zhao, S.-Q.; Zhang, S.-L.; Luo, L.-H.; Yin, D.-C.; Zhang, C.-Y. Progress of non-coding RNAs in triple-negative breast cancer. Life Sci. 2021, 272, 119238. [Google Scholar] [CrossRef]
- Ding, X.-F. Roles of miRNA and IncRNA in triple-negative breast cancer. J. Zhejiang Univ. Sci. B 2020, 21, 673–689. [Google Scholar] [CrossRef]
- Zamani, P.; Oskuee, R.K.; Atkin, S.L.; Navashenaq, J.G.; Sahebkar, A. MicroRNAs as important regulators of the NLRP3 inflammasome. Prog. Biophys. Mol. Biol. 2019, 150, 50–64. [Google Scholar] [CrossRef]
- Zhang, L.; Li, H.; Zang, Y.; Wang, F. NLRP3 inflammasome inactivation driven by miR-223-3p reduces tumor growth and increases anticancer immunity in breast cancer. Mol. Med. Rep. 2019, 19, 2180–2188. [Google Scholar] [CrossRef]
- Lyons, T.G. Targeted Therapies for Triple-Negative Breast Cancer. Curr. Treat. Options Oncol. 2019, 20, 82. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [Green Version]
- Burstein, M.D.; Tsimelzon, A.; Poage, G.M.; Covington, K.R.; Contreras, A.; Fuqua, S.A.; Savage, M.I.; Osborne, C.K.; Hilsenbeck, S.G.; Chang, J.C.; et al. Comprehensive genomic analysis identifies novel subtypes and targets of triple-negative breast cancer. Clin. Cancer Res. 2015, 21, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Jézéquel, P.; Loussouarn, D.; Guérin-Charbonnel, C.; Campion, L.; Vanier, A.; Gouraud, W.; Lasla, H.; Guette, C.; Valo, I.; Verrièle, V.; et al. Gene-expression molecular subtyping of triple-negative breast cancer tumours: Importance of immune response. Breast Cancer Res. 2015, 17, 43. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, M.; Czech, T.; Müller, H. Biological Subtypes of Triple-Negative Breast Cancer. Breast Care 2017, 12, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Abramson, V.G.; Lehmann, B.D.; Ballinger, T.J.; Pietenpol, J.A. Subtyping of triple-negative breast cancer: Implications for therapy. Cancer 2015, 121, 8–16. [Google Scholar] [CrossRef]
- Bareche, Y.; Buisseret, L.; Gruosso, T.; Girard, E.; Venet, D.; Dupont, F.; Desmedt, C.; Larsimont, D.; Park, M.; Rothé, F.; et al. Unraveling Triple-Negative Breast Cancer Tumor Microenvironment Heterogeneity: Towards an Optimized Treatment Approach. J. Natl. Cancer Inst. 2020, 112, 708–719. [Google Scholar] [CrossRef]
- Uscanga-Peralesa, G.I.; Santuario-Facio, S.K.; Ortiz-López, R. Triple negative breast cancer: Deciphering the biology and heterogeneity. Med. Univ. 2016, 18, 105–114. [Google Scholar] [CrossRef]
- Gibson, G.R.; Qian, D.; Ku, J.K.; Lai, L.L. Metaplastic Breast Cancer: Clinical Features and Outcomes. Am. Surg. 2005, 71, 725–730. [Google Scholar] [CrossRef]
- Hayes, M.J.; Thomas, D.; Emmons, A.; Giordano, T.J.; Kleer, C.G. Genetic Changes of Wnt Pathway Genes Are Common Events in Metaplastic Carcinomas of the Breast. Clin. Cancer Res. 2008, 14, 4038–4044. [Google Scholar] [CrossRef]
- Gerratana, L.; Basile, D.; Buono, G.; De Placido, S.; Giuliano, M.; Minichillo, S.; Coinu, A.; Martorana, F.; De Santo, I.; Del Mastro, L.; et al. Androgen receptor in triple negative breast cancer: A potential target for the targetless subtype. Cancer Treat. Rev. 2018, 68, 102–110. [Google Scholar] [CrossRef]
- Ahn, S.G.; Kim, S.J.; Kim, C.; Jeong, J. Molecular Classification of Triple-Negative Breast Cancer. J. Breast Cancer 2016, 19, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-Z.; Ma, D.; Suo, C.; Shi, J.; Xue, M.; Hu, X.; Xiao, Y.; Yu, K.-D.; Liu, Y.-R.; Yu, Y.; et al. Genomic and Transcriptomic Landscape of Triple-Negative Breast Cancers: Subtypes and Treatment Strategies. Cancer Cell 2019, 35, 428–440.e5. [Google Scholar] [CrossRef]
- Mahmoud, R.; Ordonez-Moran, P.; Allegrucci, C. Challenges for Triple Negative Breast Cancer Treatment: Defeating Heterogeneity and Cancer Stemness. Cancers 2022, 14, 4280. [Google Scholar] [CrossRef]
- Molchanov, O.; Maistrenko, D.; Granov, D.; Vasina, L.; Popova, A.; Vasilevskaya, I.; Mikolaichuk, O.; Shemchuk, O.; Protas, A.; Sharoyko, V.; et al. Biomarkers and potential targets for immune and cellular therapy in triple negative breast cancer. Cell. Ther. Transplant. 2022, 11, 16–30. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Pietenpol, J.A. Identification and use of biomarkers in treatment strategies for triple-negative breast cancer subtypes. J. Pathol. 2014, 232, 142–150. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Merkher, Y.; Chen, L.; Liu, N.; Leonov, S.; Chen, Y. Recent advances in therapeutic strategies for triple-negative breast cancer. J. Hematol. Oncol. 2022, 29, 121. [Google Scholar] [CrossRef]
- Ghafouri, S.R.; Guvvala, S.; Jones, C.; Philipovskiy, A. Recently approved treatment options for patients with metastatic triple-negative and HER2-neu-positive breast cancer. J. Investig. Med. 2022, 70, 1329–1341. [Google Scholar] [CrossRef]
- Xiao, Y.; Ma, D.; Zhao, S.; Suo, C.; Shi, J.; Xue, M.-Z.; Ruan, M.; Wang, H.; Zhao, J.; Li, Q.; et al. Multi-Omics Profiling Reveals Distinct Microenvironment Characterization and Suggests Immune Escape Mechanisms of Triple-Negative Breast Cancer. Clin. Cancer Res. 2019, 25, 5002–5014. [Google Scholar] [CrossRef] [PubMed]
- Heusinkveld, M.; van der Burg, S.H. Identification and manipulation of tumor associated macrophages in human cancers. J. Transl. Med. 2011, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhou, H.; Krueger, J.; Kaplan, C.; Lee, S.H.; Dolman, C.; Markowitz, D.; Wu, W.; Liu, C.; Reisfeld, R.A.; et al. Targeting tumor-associated macrophages as a novel strategy against breast cancer. J. Clin. Investig. 2006, 116, 2132–2141. [Google Scholar] [CrossRef] [Green Version]
- Prasmickaite, L.; Tenstad, E.M.; Pettersen, S.; Jabeen, S.; Egeland, E.V.; Nord, S.; Pandya, A.; Haugen, M.H.; Kristensen, V.N.; Børresen-Dale, A.; et al. Basal-like breast cancer engages tumor-supportive macrophages via secreted factors induced by extracellular S100A4. Mol. Oncol. 2018, 12, 1540–1558. [Google Scholar] [CrossRef] [PubMed]
- Pe, K.C.S.; Saetung, R.; Yodsurang, V.; Chaotham, C.; Suppipat, K.; Chanvorachote, P.; Tawinwung, S. Triple-negative breast cancer influences a mixed M1/M2 macrophage phenotype associated with tumor aggressiveness. PLoS ONE 2022, 17, e0273044. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.K.; Kadel, S.; Townsend, M.G.; Oliwa, M.; Guerriero, J.L. Macrophage Biology and Mechanisms of Immune Suppression in Breast Cancer. Front. Immunol. 2021, 12, 643771. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y. M1 and M2 polarization of macrophages: A mini-review. Med. Biol. Sci. Eng. 2019, 2, 1–5. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The Inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Ershaid, N.; Sharon, Y.; Doron, H.; Raz, Y.; Shani, O.; Cohen, N.; Monteran, L.; Leider-Trejo, L.; Ben-Shmuel, A.; Yassin, M.; et al. NLRP3 inflammasome in fibroblasts links tissue damage with inflammation in breast cancer progression and metastasis. Nat. Commun. 2019, 10, 4375. [Google Scholar] [CrossRef]
- George, M.A.; Lustberg, M.B.; Orchard, T.S. Psychoneurological symptom cluster in breast cancer: The role of inflammation and diet. Breast Cancer Res. Treat. 2020, 184, 1–9. [Google Scholar] [CrossRef]
- Sonnessa, M.; Cioffi, A.; Brunetti, O.; Silvestris, N.; Zito, F.A.; Saponaro, C.; Mangia, A. NLRP3 Inflammasome From Bench to Bedside: New Perspectives for Triple Negative Breast Cancer. Front. Oncol. 2020, 10, 1587. [Google Scholar] [CrossRef]
- Medina, M.A.; Oza, G.; Sharma, A.; Arriaga, L.G.; Hernandez, J.M.; Rotello, V.M.; Ramirez, J.T. Triple-Negative Breast Cancer: A Review of Conventional and Advanced Therapeutic Strategies. Int. J. Environ. Res. Public Health 2020, 17, 2078. [Google Scholar] [CrossRef]
- Pauls, M.; Chia, S.; LeVasseur, N. Current and New Novel Combination Treatments for Metastatic Triple-Negative Breast Cancer. Curr. Oncol. 2022, 29, 4748–4767. [Google Scholar] [CrossRef]
- Carey, L.A.; Dees, E.C.; Sawyer, L.; Gatti, L.; Moore, D.T.; Collichio, F.; Ollila, D.W.; Sartor, C.I.; Graham, M.L.; Perou, C.M. The Triple Negative Paradox: Primary Tumor Chemosensitivity of Breast Cancer Subtypes. Clin. Cancer Res. 2007, 13, 2329–2334. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.; Ruiz, A.; Ruiz Borrego, M.; Barnadas, A.; González, S.; Calvo, L.; Vila, M.M.; Antón, A.; Rodríguez-Lescure, A.; Seguí-Palmer, M.A.; et al. Fluorouracil, Doxorubicin, and Cyclophosphamide (FAC) Versus FAC Followed by Weekly Paclitaxel As Adjuvant Therapy for High-Risk, Node-Negative Breast Cancer: Results From the GEICAM/2003-02 Study. J. Clin. Oncol. 2013, 31, 2593–2599. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Ahn, J.-H.; Kim, S.-B. How shall we treat early triple-negative breast cancer (TNBC): From the current standard to upcoming immuno-molecular strategies. ESMO Open 2018, 3, e000357. [Google Scholar] [CrossRef] [PubMed]
- Bou Zerdan, M.; Ghorayeb, T.; Saliba, F.; Allam, S.; Bou Zerdan, M.; Yaghi, M.; Bilani, N.; Jaafar, R.; Nahleh, Z. Triple Negative Breast Cancer: Updates on Classification and Treatment in 2021. Cancers 2022, 14, 1253. [Google Scholar] [CrossRef]
- Emens, L.A. Breast Cancer Immunotherapy: Facts and Hopes. Clin. Cancer Res. 2018, 24, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Pandy, J.G.P.; Balolong-Garcia, J.C.; Cruz-Ordinario, M.V.B.; Que, F.V.F. Triple negative breast cancer and platinum-based systemic treatment: A meta-analysis and systematic review. BMC Cancer 2019, 19, 1253. [Google Scholar] [CrossRef]
- Liu, M.; Mo, Q.-G.; Wei, C.-Y.; Qin, Q.-H.; Huang, Z.; He, J. Platinum-based chemotherapy in triple-negative breast cancer: A meta-analysis. Oncol. Lett. 2012, 5, 983–991. [Google Scholar] [CrossRef]
- Bian, L.; Yu, P.; Wen, J.; Na Li, N.; Huang, W.; Xie, X.; Ye, F. Survival benefit of platinum-based regimen in early stage triple negative breast cancer: A meta-analysis of randomized controlled trials. npj Breast Cancer 2021, 7, 157. [Google Scholar] [CrossRef]
- Thu, K.L.; Soria-Bretones, I.; Mak, T.W.; Cescon, D.W. Targeting the cell cycle in breast cancer: Towards the next phase. Cell Cycle 2018, 17, 1871–1885. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, D.; Yao, Z.; Lin, X.; Liu, J.; Gu, Q.; Dong, X.; Liu, F.; Wang, Y.; Yao, N.; et al. Anti-angiogenic treatment promotes triple-negative breast cancer invasion via vasculogenic mimicry. Cancer Biol. Ther. 2017, 18, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Wu, Y.; He, P.; Fan, Y.; Zhong, X.; Zheng, H.; Luo, T. PI3K/AKT/mTOR-Targeted Therapy for Breast Cancer. Cells 2022, 11, 2508. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.L.B.; Han, H.S.; Gradishar, W.J. Targeting the PI3K/AKT/mTOR pathway in triple-negative breast cancer: A review. Breast Cancer Res. Treat. 2018, 169, 397–406. [Google Scholar] [CrossRef]
- Khan, M.A.; Jain, V.K.; Rizwanullah, M.; Ahmad, J.; Jain, K. PI3K/AKT/mTOR pathway inhibitors in triple-negative breast cancer: A review on drug discovery and future challenges. Drug Discov. Today 2019, 24, 2181–2191. [Google Scholar] [CrossRef]
- Singh, D.D.; Parveen, A.; Yadav, D.K. Role of PARP in TNBC: Mechanism of Inhibition, Clinical Applications, and Resistance. Biomedicines 2021, 9, 1512. [Google Scholar] [CrossRef]
- Maennling, A.E.; Tur, M.K.; Niebert, M.; Klockenbring, T.; Zeppernick, F.; Gattenlöhner, S.; Meinhold-Heerlein, I.; Hussain, A.F. Molecular Targeting Therapy against EGFR Family in Breast Cancer: Progress and Future Potentials. Cancers 2019, 11, 1826. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Lau, K.-Y.; Hsu, C.-C.; Chen, J.-L.; Lee, C.-H.; Huang, T.-T.; Chen, Y.-T.; Huang, C.-T.; Lin, P.-H.; Tseng, L.-M. Combination of palbociclib with enzalutamide shows in vitro activity in RB proficient and androgen receptor positive triple negative breast cancer cells. PLoS ONE 2017, 12, e0189007. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Jain, S. Immune signalling by supramolecular assemblies. Immunology 2018, 155, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Kanneganti, T.-D. Inflammasome activation and assembly at a glance. J. Cell Sci. 2017, 130, 3955–3963. [Google Scholar] [CrossRef]
- Komada, T.; Muruve, D.A. The role of inflammasomes in kidney disease. Nat. Rev. Nephrol. 2019, 15, 501–520. [Google Scholar] [CrossRef]
- Yi, Y.-S. Regulatory Roles of Caspase-11 Non-Canonical Inflammasome in Inflammatory Liver Diseases. Int. J. Mol. Sci. 2022, 23, 4986. [Google Scholar] [CrossRef]
- Kantono, M.; Guo, B. Inflammasomes and Cancer: The Dynamic Role of the Inflammasome in Tumor Development. Front. Immunol. 2017, 8, 1132. [Google Scholar] [CrossRef] [PubMed]
- Montico, B.; Nigro, A.; Casolaro, V.; Col, J.D. Immunogenic Apoptosis as a Novel Tool for Anticancer Vaccine Development. Int. J. Mol. Sci. 2018, 19, 594. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal Transduct. Target. Ther. 2021, 6, 128. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Ding, J.; Wang, C.; Zhou, X.; Gao, W.; Huang, H.; Shao, F.; Liu, Z. A bioorthogonal system reveals antitumour immune function of pyroptosis. Nature 2020, 579, 421–426. [Google Scholar] [CrossRef]
- Poli, G.; Fabi, C.; Bellet, M.M.; Costantini, C.; Nunziangeli, L.; Romani, L.; Brancorsini, S. Epigenetic Mechanisms of Inflammasome Regulation. Int. J. Mol. Sci. 2020, 21, 5758. [Google Scholar] [CrossRef]
- Rodríguez-Bautista, R.; Caro-Sánchez, C.H.; Cabrera-Galeana, P.; Alanis-Funes, G.J.; Gutierrez-Millán, E.; Ávila-Ríos, S.; Matías-Florentino, M.; Reyes-Terán, G.; Díaz-Chávez, J.; Villarreal-Garza, C.; et al. Immune Milieu and Genomic Alterations Set the Triple-Negative Breast Cancer Immunomodulatory Subtype Tumor Behavior. Cancers 2021, 13, 6256. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wei, Q.; Xu, C.; Dong, M.; Zhao, W. Immune landscape and risk prediction based on pyroptosis-related molecular subtypes in triple-negative breast cancer. Front. Immunol. 2022, 13, 933703. [Google Scholar] [CrossRef]
- Deets, K.A.; Vance, R.E. Inflammasomes and adaptive immune responses. Nat. Immunol. 2021, 22, 412–422. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Xu, Y.; Peng, T.; Meng, X.; Zou, F. NLRP3 induces the autocrine secretion of IL-1β to promote epithelial–mesenchymal transition and metastasis in breast cancer. Biochem. Biophys. Res. Commun. 2021, 560, 72–79. [Google Scholar] [CrossRef]
- Hamarsheh, S.; Zeiser, R. NLRP3 Inflammasome Activation in Cancer: A Double-Edged Sword. Front. Immunol. 2020, 11, 1444. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, H.; Hao, Y.; Lin, H.; Dong, M.; Ye, J.; Song, L.; Wang, Y.; Li, Q.; Shan, B.; et al. Myeloid PTEN promotes chemotherapy-induced NLRP3-inflammasome activation and antitumour immunity. Nat. Cell Biol. 2020, 22, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef]
- Bakr, N.M.; Mahmoud, M.S.; Nabil, R.; Boushnak, H.; Swellam, M. Impact of circulating miRNA-373 on breast cancer diagnosis through targeting VEGF and cyclin D1 genes. J. Genet. Eng. Biotechnol. 2021, 19, 84. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; Anand, P.K.; Malireddi, R.K.; Vande Walle, L.; Van Opdenbosch, N.; Dillon, C.P.; Weinlich, R.; Green, D.; Lamkanfi, M.; Kanneganti, T.-D. FADD and Caspase-8 Mediate Priming and Activation of the Canonical and Noncanonical Nlrp3 Inflammasomes. J. Immunol. 2014, 192, 1835–1846. [Google Scholar] [CrossRef]
- Lv, Z.; Xin, H.; Yang, Z.; Wang, W.; Dong, J.; Jin, L.; Li, F. miR-135b promotes proliferation and metastasis by targeting APC in triple-negative breast cancer. J. Cell. Physiol. 2019, 234, 10819–10826. [Google Scholar] [CrossRef] [PubMed]
- Halappanavar, S.; Nikota, J.; Wu, D.; Williams, A.; Yauk, C.L.; Stampfli, M. IL-1 Receptor Regulates microRNA-135b Expression in a Negative Feedback Mechanism during Cigarette Smoke–Induced Inflammation. J. Immunol. 2013, 190, 3679–3686. [Google Scholar] [CrossRef]
- Marwarha, G.; Røsand, Ø.; Scrimgeour, N.; Slagsvold, K.H.; Høydal, M.A. miR-210 Regulates Apoptotic Cell Death during Cellular Hypoxia and Reoxygenation in a Diametrically Opposite Manner. Biomedicines 2021, 10, 42. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, X.; Kong, X.; Zhai, J.; Fang, Y.; Guan, X.; Wang, J. Expression signatures and roles of microRNAs in inflammatory breast cancer. Cancer Cell Int. 2019, 19, 23. [Google Scholar] [CrossRef]
- Michaille, J.-J.; Piurowski, V.; Rigot, B.; Kelani, H.; Fortman, E.C.; Tili, E. MiR-663, a MicroRNA Linked with Inflammation and Cancer That Is under the Influence of Resveratrol. Medicines 2018, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- Carden, T.; Singh, B.; Mooga, V.; Bajpai, P.; Singh, K.K. Epigenetic modification of miR-663 controls mitochondria-to-nucleus retrograde signaling and tumor progression. J. Biol. Chem. 2017, 292, 20694–20706. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, K.; Yu, J.; Hao, X.-M.; Liu, Y.-L.; Xing, A.-Y. miR-638 Serves as a Biomarker of 5-Fluorouracil Sensitivity to Neoadjuvant Chemotherapy in Breast Cancer. J. Breast Cancer 2022, 25, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Bandiera, S.; Rüberg, S.; Girard, M.; Cagnard, N.; Hanein, S.; Chrétien, D.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. Nuclear Outsourcing of RNA Interference Components to Human Mitochondria. PLoS ONE 2011, 6, e20746. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, C.; Li, J.; Li, X.; Zhou, P. MicroRNA-217 aggravates breast cancer through activation of NF1-mediated HSF1/ATG7 axis and c-Jun/ATF3/MMP13 axis. Hum. Cell 2023, 36, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, Y.; Qian, X.; Hu, Y.; Liu, J.; Zhang, S.; Zhang, J. MiR-340 Inhibits Triple-Negative Breast Cancer Progression by Reversing EZH2 Mediated miRNAs Dysregulated Expressions. J. Cancer 2017, 8, 3037–3048. [Google Scholar] [CrossRef] [PubMed]
- Zimta, A.-A.; Cenariu, D.; Irimie, A.; Magdo, L.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. The Role of Nrf2 Activity in Cancer Development and Progression. Cancers 2019, 11, 1755. [Google Scholar] [CrossRef]
- Yu, T.; Ma, P.; Wu, D.; Shu, Y.; Gao, W. Functions and mechanisms of microRNA-31 in human cancers. Biomed. Pharmacother. 2018, 108, 1162–1169. [Google Scholar] [CrossRef]
- Hashemi, Z.S.; Moghadam, M.F.; Farokhimanesh, S.; Rajabibazl, M.; Sadroddiny, E. Inhibition of breast cancer metastasis by co-transfection of miR-31/193b-mimics. Iran J Basic Med. Sci 2018, 21, 427–433. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, X.; Han, B.; Ji, L.; Yao, L.; Wang, Z. High Expression of microRNA-223 Indicates a Good Prognosis in Triple-Negative Breast Cancer. Front. Oncol. 2021, 11, 630432. [Google Scholar] [CrossRef]
- Favero, A.; Segatto, I.; Perin, T.; Belletti, B. The many facets of miR -223 in cancer: Oncosuppressor, oncogenic driver, therapeutic target, and biomarker of response. Wiley Interdiscip. Rev. RNA 2021, 12, e1659. [Google Scholar] [CrossRef]
- Socinski, M.A.; Jotte, R.M.; Cappuzzo, F.; Orlandi, F.; Stroyakovskiy, D.; Nogami, N.; Rodríguez-Abreu, D.; Moro-Sibilot, D.; Thomas, C.A.; Barlesi, F.; et al. Atezolizumab for First-Line Treatment of Metastatic Nonsquamous NSCLC. N. Engl. J. Med. 2018, 378, 2288–2301. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Xu, J.; Zhang, B.; Liu, J.; Liang, C.; Hua, J.; Meng, Q.; Yu, X.; Shi, S. Ferroptosis, necroptosis, and pyroptosis in anticancer immunity. J. Hematol. Oncol. 2020, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Alyami, N.M. MicroRNAs Role in Breast Cancer: Theranostic Application in Saudi Arabia. Front. Oncol. 2021, 11, 717759. [Google Scholar] [CrossRef]
- Otmani, K.; Lewalle, P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef]
- Mehrgou, A.; Akouchekian, M. Therapeutic impacts of microRNAs in breast cancer by their roles in regulating processes involved in this disease. J. Res. Med. Sci. 2017, 22, 130. [Google Scholar] [CrossRef]
- Wu, J.; Sun, Z.; Sun, H.; Li, Y. MicroRNA-27a promotes tumorigenesis via targeting AKT in triple negative breast cancer. Mol. Med. Rep. 2018, 17, 562–570. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Wang, J. MicroRNA-384 inhibits the progression of breast cancer by targeting ACVR1. Oncol. Rep. 2018, 39, 2563–2574. [Google Scholar] [CrossRef]
- Dong, G.; Liang, X.; Wang, D.; Gao, H.; Wang, L.; Wang, L.; Liu, J.; Du, Z. High expression of miR-21 in triple-negative breast cancers was correlated with a poor prognosis and promoted tumor cell in vitro proliferation. Med. Oncol. 2014, 31, 57. [Google Scholar] [CrossRef]
- Cheng, H.-Y.; Hsieh, C.-H.; Lin, P.-H.; Chen, Y.-T.; Hsu, D.S.-S.; Tai, S.-K.; Chu, P.-Y.; Yang, M.-H. Snail-regulated exosomal microRNA-21 suppresses NLRP3 inflammasome activity to enhance cisplatin resistance. J. Immunother. Cancer 2022, 10, e004832. [Google Scholar] [CrossRef]
- Song, H.; Li, D.; Wu, T.; Xie, D.; Hua, K.; Hu, J.; Deng, X.; Ji, C.; Deng, Y.; Fang, L. MicroRNA-301b promotes cell proliferation and apoptosis resistance in triple-negative breast cancer by targeting CYLD. BMB Rep. 2018, 51, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.-J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. miR-221/222 Targeting of Trichorhinophalangeal 1 (TRPS1) Promotes Epithelial-to-Mesenchymal Transition in Breast Cancer. Sci. Signal. 2011, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Niedzielski, M.; Broncel, M.; Gorzelak-Pabiś, P.; Woźniak, E. New possible pharmacological targets for statins and ezetimibe. Biomed. Pharmacother. 2020, 129, 110388. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cano, I.; Pattanayak, B.; Adam-Artigues, A.; Lameirinhas, A.; Torres-Ruiz, S.; Tormo, E.; Cervera, R.; Eroles, P. MicroRNAs as a clue to overcome breast cancer treatment resistance. Cancer Metastasis Rev. 2022, 41, 77–105. [Google Scholar] [CrossRef]
- Vlacil, A.-K.; Vollmeister, E.; Bertrams, W.; Schoesser, F.; Oberoi, R.; Schuett, J.; Schuett, H.; Huehn, S.; Bedenbender, K.; Schmeck, B.T.; et al. Identification of microRNAs involved in NOD-dependent induction of pro-inflammatory genes in pulmonary endothelial cells. PLoS ONE 2020, 15, e0228764. [Google Scholar] [CrossRef]
- Tormo, E.; Ballester, S.; Adam-Artigues, A.; Burgués, O.; Alonso, E.; Bermejo, B.; Menéndez, S.; Zazo, S.; Madoz-Gúrpide, J.; Rovira, A.; et al. The miRNA-449 family mediates doxorubicin resistance in triple-negative breast cancer by regulating cell cycle factors. Sci. Rep. 2019, 29, 5316. [Google Scholar] [CrossRef]
- Qu, Y.; Dou, B.; Tan, H.; Feng, Y.; Wang, N.; Wang, D. Tumor microenvironment-driven non-cell-autonomous resistance to antineoplastic treatment. Mol. Cancer 2019, 18, 69. [Google Scholar] [CrossRef]
- Cataldo, A.; Romero-Cordoba, S.; Plantamura, I.; Cosentino, G.; Hidalgo-Miranda, A.; Tagliabue, E.; Iorio, M.V. MiR-302b as a Combinatorial Therapeutic Approach to Improve Cisplatin Chemotherapy Efficacy in Human Triple-Negative Breast Cancer. Cancers 2020, 12, 2261. [Google Scholar] [CrossRef]
- Ma, T.; Liu, X.; Cen, Z.; Xin, C.; Guo, M.; Zou, C.; Song, W.; Xie, R.; Wang, K.; Zhou, H.; et al. MicroRNA-302b negatively regulates IL-1β production in response to MSU crystals by targeting IRAK4 and EphA2. Arthritis Res. Ther. 2018, 20, 34. [Google Scholar] [CrossRef]
- Lee, S.J.; Jeong, J.-H.; Kang, S.H.; Kang, J.; Kim, E.A.; Lee, J.; Jung, J.H.; Park, H.Y.; Chae, Y.S. MicroRNA-137 Inhibits Cancer Progression by Targeting Del-1 in Triple-Negative Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 6162. [Google Scholar] [CrossRef]
- Ying, X.; Sun, Y.; He, P. MicroRNA-137 inhibits BMP7 to enhance the epithelial-mesenchymal transition of breast cancer cells. Oncotarget 2017, 8, 18348–18358. [Google Scholar] [CrossRef] [PubMed]
- Zimta, A.-A.; Tigu, A.B.; Muntean, M.; Cenariu, D.; Slaby, O.; Berindan-Neagoe, I. Molecular Links between Central Obesity and Breast Cancer. Int. J. Mol. Sci. 2019, 20, 5364. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Song, C.; Wang, X.; Li, Y.; Bai, X.; Liang, X.; Wu, J.; Liu, J. Downregulation of miR-155-5p enhances the anti-tumor effect of cetuximab on triple-negative breast cancer cells via inducing cell apoptosis and pyroptosis. Aging 2021, 13, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Bar, I.; Theate, I.; Haussy, S.; Beniuga, G.; Carrasco, J.; Canon, J.-L.; Delrée, P.; Merhi, A. MiR-210 Is Overexpressed in Tumor-infiltrating Plasma Cells in Triple-negative Breast Cancer. J. Histochem. Cytochem. 2020, 68, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Pottoo, F.H.; Iqubal, A.; Iqubal, M.K.; Salahuddin, M.; Rahman, J.U.; AlHajri, N.; Shehadeh, M. miRNAs in the Regulation of Cancer Immune Response: Effect of miRNAs on Cancer Immunotherapy. Cancers 2021, 13, 6145. [Google Scholar] [CrossRef]
- Piasecka, D.; Braun, M.; Kordek, R.; Sadej, R.; Romanska, H. MicroRNAs in regulation of triple-negative breast cancer progression. J. Cancer Res. Clin. Oncol. 2018, 144, 1401–1411. [Google Scholar] [CrossRef]
- Rippo, M.R.; Olivieri, F.; Monsurrò, V.; Prattichizzo, F.; Albertini, M.C.; Procopio, A.D. MitomiRs in human inflamm-aging: A hypothesis involving miR-181a, miR-34a and miR-146a. Exp. Gerontol. 2014, 56, 154–163. [Google Scholar] [CrossRef]
- Li, D.; Hu, J.; Song, H.; Xu, H.; Wu, C.; Zhao, B.; Xie, D.; Wu, T.; Zhao, J.; Fang, L. miR-143-3p targeting LIM domain kinase 1 suppresses the progression of triple-negative breast cancer cells. Am. J. Transl. Res. 2017, 9, 2276–2285. [Google Scholar]
- Meng, L.; Li, L.; Lu, S.; Li, K.; Su, Z.; Wang, Y.; Fan, X.; Li, X.; Zhao, G. The protective effect of dexmedetomidine on LPS-induced acute lung injury through the HMGB1-mediated TLR4/NF-κB and PI3K/Akt/mTOR pathways. Mol. Immunol. 2018, 94, 7–17. [Google Scholar] [CrossRef]
- Li, J.; Lai, Y.; Ma, J.; Liu, Y.; Bi, J.; Zhang, L.; Chen, L.; Yao, C.; Lv, W.; Chang, G.; et al. miR-17-5p suppresses cell proliferation and invasion by targeting ETV1 in triple-negative breast cancer. BMC Cancer 2017, 17, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, W.; Wang, Y.; Xu, X.; Lv, S.; Dong, X. miR-17-5p promotes migration and invasion in breast cancer cells by repressing netrin 4. Int. J. Clin. Exp. Pathol. 2019, 12, 1649–1657. [Google Scholar]
- Zhou, W.; Song, F.; Wu, Q.; Liu, R.; Wang, L.; Liu, C.; Peng, Y.; Mao, S.; Feng, J.; Chen, C. miR-217 inhibits triple-negative breast cancer cell growth, migration, and invasion through targeting KLF5. PLoS ONE 2017, 12, e0176395. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L.; Zhang, Z.-J.; Yi, Z.-B.; Li, J.-J. MicroRNA-211-5p suppresses tumour cell proliferation, invasion, migration and metastasis in triple-negative breast cancer by directly targeting SETBP1. Br. J. Cancer 2017, 117, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Yarahmadi, S.; Abdolvahabi, Z.; Hesari, Z.; Tavakoli-Yaraki, M.; Yousefi, Z.; Seiri, P.; Hosseinkhani, S.; Nourbakhsh, M. Inhibition of sirtuin 1 deacetylase by miR-211-5p provides a mechanism for the induction of cell death in breast cancer cells. Gene 2019, 711, 143939. [Google Scholar] [CrossRef]
- Tang, H.; Liu, P.; Yang, L.; Xie, X.; Ye, F.; Wu, M.; Liu, X.; Chen, B.; Zhang, L.; Xie, X. miR-185 Suppresses Tumor Proliferation by Directly Targeting E2F6 and DNMT1 and Indirectly Upregulating BRCA1 in Triple-Negative Breast Cancer. Mol. Cancer Ther. 2014, 13, 3185–3197. [Google Scholar] [CrossRef]
- Toda, H.; Kurozumi, S.; Kijima, Y.; Idichi, T.; Shinden, Y.; Yamada, Y.; Arai, T.; Maemura, K.; Fujii, T.; Horiguchi, J.; et al. Molecular pathogenesis of triple-negative breast cancer based on microRNA expression signatures: Antitumor miR-204-5p targets AP1S3. J. Hum. Genet. 2018, 63, 1197–1210. [Google Scholar] [CrossRef]
- Xiao, M.; Lou, C.; Xiao, H.; Yang, Y.; Cai, X.; Li, C.; Jia, S.; Huang, Y. MiR-128 regulation of glucose metabolism and cell proliferation in triple-negative breast cancer. Br. J. Cancer 2018, 105, 75–85. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, D.; Shen, P.; Yu, Y.; Yan, Y.; You, W. Hsa_circ_0092276 promotes doxorubicin resistance in breast cancer cells by regulating autophagy via miR-348/ATG7 axis. Transl. Oncol. 2021, 14, 101045. [Google Scholar] [CrossRef]
- Rogers, T.J.; Christenson, J.L.; Greene, L.I.; O’Neill, K.I.; Williams, M.M.; Gordon, M.A.; Nemkov, T.; D’Alessandro, A.; Degala, G.D.; Shin, J.; et al. Reversal of Triple-Negative Breast Cancer EMT by miR-200c Decreases Tryptophan Catabolism and a Program of Immunosuppression. Mol. Cancer Res. 2019, 17, 30–41. [Google Scholar] [CrossRef]
- Carlomosti, F.; D’Agostino, M.; Beji, S.; Torcinaro, A.; Rizzi, R.; Zaccagnini, G.; Maimone, B.; Di Stefano, V.; De Santa, F.; Cordisco, S.; et al. Oxidative Stress-Induced miR-200c Disrupts the Regulatory Loop Among SIRT1, FOXO1, and eNOS. Antioxid. Redox Signal. 2017, 27, 328–344. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Barbano, R.; Dama, E.; Pasculli, B.; Rendina, M.; Morritti, M.G.; Melocchi, V.; Castelvetere, M.; Valori, V.M.; Ravaioli, S.; et al. Combined analysis of miR-200 family and its significance for breast cancer. Sci. Rep. 2021, 11, 2980. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-K.; Moon, Y.-A.; Kil Song, C.; Baskaran, R.; Bae, S.; Yang, S.-G. Tumor-suppressing miR-141 gene complex-loaded tissue-adhesive glue for the locoregional treatment of hepatocellular carcinoma. Theranostics 2018, 8, 3891–3901. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.-J.; Song, Y.; Ren, X.-Y.; Xu, Y.-L.; Zhou, Y.-D.; Liang, Z.-Y.; Sun, Q. MicroRNA-27b-3p Promotes Tumor Progression and Metastasis by Inhibiting Peroxisome Proliferator-Activated Receptor Gamma in Triple-Negative Breast Cancer. Front. Oncol. 2020, 10, 1371. [Google Scholar] [CrossRef]
- Lamberti, M.J.; Nigro, A.; Casolaro, V.; Rumie Vittar, N.B.R.; Dal Col, J.D. Damage-Associated Molecular Patterns Modulation by microRNA: Relevance on Immunogenic Cell Death and Cancer Treatment Outcome. Cancers 2021, 13, 2566. [Google Scholar] [CrossRef]
- Chen, M.-Y.; Ye, X.-J.; He, X.-H.; Ouyang, D.-Y. The Signaling Pathways Regulating NLRP3 Inflammasome Activation. Inflammation 2021, 44, 1229–1245. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Inflammasome activation and regulation: Toward a better understanding of complex mechanisms. Cell Discov. 2020, 6, 36. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Y.; Chang, W.; Zhang, L.; Syrigos, K.N.; Li, P. Noncoding RNA-mediated regulation of pyroptotic cell death in cancer. Front. Oncol. 2022, 12, 1015587. [Google Scholar] [CrossRef]
- Wang, J.G.; Jian, W.J.; Li, Y.; Zhang, J. Nobiletin promotes the pyroptosis of breast cancer via regulation of miR -200b/JAZF1 axis. Kaohsiung J. Med. Sci. 2021, 37, 572–582. [Google Scholar] [CrossRef]
- Glinsky, G.V. SNP-guided microRNA maps (MirMaps) of 16 common human disorders identify a clinically accessible therapy reversing transcriptional aberrations of nuclear import and inflammasome pathways. Cell Cycle 2008, 7, 3564–3576. [Google Scholar] [CrossRef]
- Tang, H.-C.; Lai, Y.-Y.; Zheng, J.; Jiang, H.-Y.; Xu, G. miR-223-3p Inhibits Antigen Endocytosis and Presentation and Promotes the Tolerogenic Potential of Dendritic Cells through Targeting Mannose Receptor Signaling and Rhob. J. Immunol. Res. 2020, 1379458. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, X.; Zhang, P.; Luo, J.; Mou, E.; Liu, S. miR-143-5p suppresses breast cancer progression by targeting the HIF-1α-related GLUT1 pathway. Oncol. Lett. 2022, 23, 147. [Google Scholar] [CrossRef] [PubMed]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The Pervasive Role of the miR-181 Family in Development, Neurodegeneration, and Cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef]
- Heneghan, H.M.; Miller, N.; Lowery, A.; Sweeney, K.J.; Kerin, M.J. MicroRNAs as Novel Biomarkers for Breast Cancer. J. Oncol. 2010, 1950201. [Google Scholar] [CrossRef]
- Keklikoglou, I.; Koerner, C.; Schmidt, C.; Zhang, J.D.; Heckmann, D.; Shavinskaya, A.; Allgayer, H.; Guckel, B.; Fehm, T.F.; Schneeweiss, A.; et al. MicroRNA-520/373 family functions as a tumor suppressor in estrogen receptor negative breast cancer by targeting NF-κB and TGF-β signaling pathways. Oncogene 2012, 31, 4150–4163. [Google Scholar] [CrossRef]
- Ding, Q.; Shen, L.; Nie, X.; Lu, B.; Pan, X.; Su, Z.; Yan, A.; Yan, R.; Zhou, Y.; Li, L.; et al. MiR-223-3p overexpression inhibits cell proliferation and migration by regulating inflammation-associated cytokines in glioblastomas. Pathol. Res. Pr. 2018, 214, 1330–1339. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Attar, R.; Tanriover, G.; Sabitaliyevich, U.Y.; Zhailganov, A.; Rabandiyarov, M. Regulation of NLRP3 by non-coding RNAs in different cancers: Interplay between non-coding RNAs and NLRP3 in carcinogenesis and metastasis. Cell. Mol. Biol. 2020, 66, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, Z.; Fan, Y.; Zhang, J.; Chen, K.; Gao, L.; Zeng, H.; Cao, J.; Wang, C. MicroRNA-9 Inhibits NLRP3 Inflammasome Activation in Human Atherosclerosis Inflammation Cell Models through the JAK1/STAT Signaling Pathway. Cell. Physiol. Biochem. 2017, 41, 1555–1571. [Google Scholar] [CrossRef] [PubMed]
- Kaboli, P.J.; Rahmat, A.; Ismail, P.; Ling, K.-H. MicroRNA-based therapy and breast cancer: A comprehensive review of novel therapeutic strategies from diagnosis to treatment. Pharmacol. Res. 2015, 97, 104–121. [Google Scholar] [CrossRef]
- Grimaldi, A.M.; Salvatore, M.; Incoronato, M. miRNA-Based Therapeutics in Breast Cancer: A Systematic Review. Front. Oncol. 2021, 11, 668464. [Google Scholar] [CrossRef]
- Hussen, B.M.; Abdullah, S.T.; Rasul, M.F.; Salihi, A.; Ghafouri-Fard, S.; Hidayat, H.J.; Taheri, M. MicroRNAs: Important Players in Breast Cancer Angiogenesis and Therapeutic Targets. Front. Mol. Biosci. 2021, 8, 764025. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Wang, R.; Chen, L. Micro RNA-145: A potent tumour suppressor that regulates multiple cellular pathways. J. Cell. Mol. Med. 2014, 18, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X. miR-1290 modulates the radioresistance of triple-negative breast cancer by targeting NLRP3-mediated pyroptosis. Clin. Transl. Oncol. 2022, 24, 1764–1775. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Xi, Q.; Liu, H.; Guo, X.; Zhang, J.; Zhang, Z.; Li, Y.; Yang, G.; Zhou, D.; Yang, H.; et al. miR-21 promotes NLRP3 inflammasome activation to mediate pyroptosis and endotoxic shock. Cell Death Dis. 2019, 10, 461. [Google Scholar] [CrossRef]
- Kotiyal, S.; Bhattacharya, S. Biochemical and biophysical research communications breast cancer stem cells, EMT and therapeutic targets. Biochem. Biophys. Res. Commun. 2014, 453, 112–116. [Google Scholar] [CrossRef]
- Chabot, S.; Teissié, J.; Golzio, M. Targeted electro-delivery of oligonucleotides for RNA interference: siRNA and antimiR. Adv. Drug Deliv. Rev. 2015, 81, 161–168. [Google Scholar] [CrossRef]
- Conde, J.; Oliva, N.; Atilano, M.; Song, H.S.; Artzi, N. Self-assembled RNA-triple-helix hydrogel scaffold for microRNA modulation in the tumour microenvironment. Nat. Mater. 2016, 15, 353–363. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, S.; Tang, Q.; Yan, Z. Knockdown of miR-205-5p alleviates the inflammatory response in allergic rhinitis by targeting B-cell lymphoma 6. Mol. Med. Rep. 2021, 24, 181. [Google Scholar] [CrossRef]
- Zhang, G.-F.; Wu, J.-C.; Wang, H.-Y.; Jiang, W.-D.; Qiu, L. Overexpression of microRNA-205-5p exerts suppressive effects on stem cell drug resistance in gallbladder cancer by down-regulating PRKCE. Biosci. Rep. 2020, 40, BSR20194509. [Google Scholar] [CrossRef]
- Lee, P.-S.; Chiou, Y.-S.; Ho, C.-T.; Pan, M.-H. Chemoprevention by resveratrol and pterostilbene: Targeting on epigenetic regulation. Biofactors 2017, 44, 26–35. [Google Scholar] [CrossRef]
- Song, J.; Ouyang, Y.; Che, J.; Li, X.; Zhao, Y.; Yang, K.; Zhao, X.; Chen, Y.; Fan, C.; Yuan, W. Potential Value of miR-221/222 as Diagnostic, Prognostic, and Therapeutic Biomarkers for Diseases. Front. Immunol. 2017, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Ebert, M.S.; Sharp, P.A. MicroRNA sponges: Progress and possibilities. RNA 2010, 16, 2043–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, F.C.; Lim, J.K.; Zhu, H.; Hin, L.C.; Wang, S. Using artificial microRNA sponges to achieve microRNA loss-of-function in cancer cells. Adv. Drug Deliv. Rev. 2014, 81, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, J.; Song, Q.; Yang, X.; Wang, D.; Liu, W.; Shi, J.; Zhang, Z. DNA Nanosponge for Adsorption and Clearance of Intracellular miR-21 and Enhanced Antitumor Chemotherapy. ACS Appl. Mater. Interfaces 2019, 11, 46604–46613. [Google Scholar] [CrossRef]
- Mandal, C.C.; Ghosh-Choudhury, T.; Dey, N.; Choudhury, G.G.; Ghosh-Choudhury, N. miR-21 is targeted by omega-3 polyunsaturated fatty acid to regulate breast tumor CSF-1 expression. Carcinogenesis 2012, 33, 1897–1908. [Google Scholar] [CrossRef]
- Singh, S.K.; Singh, S.; Lillard, J.W., Jr.; Singh, R. Drug delivery approaches for breast cancer. Int. J. Nanomed. 2017, 12, 6205–6218. [Google Scholar] [CrossRef]
- Adinew, G.; Taka, E.; Mendonca, P.; Messeha, S.; Soliman, K. The Anticancer Effects of Flavonoids through miRNAs Modulations in Triple-Negative Breast Cancer. Nutrients 2021, 13, 1212. [Google Scholar] [CrossRef]
- Cassidy, A.; Minihane, A.M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2016, 105, 10–22. [Google Scholar] [CrossRef]
- Wang, M.; Chen, X.; Yu, F.; Zhang, L.; Zhang, Y.; Chang, W. The Targeting of Noncoding RNAs by Quercetin in Cancer Prevention and Therapy. Oxidative Med. Cell. Longev. 2022, 2022, 4330681. [Google Scholar] [CrossRef]
- La Rosa, F.; Mancuso, R.; Agostini, S.; Piancone, F.; Marventano, I.; Saresella, M.; Hernis, A.; Fenoglio, C.; Galimberti, D.; Scarpini, E.; et al. Pharmacological and Epigenetic Regulators of NLRP3 Inflammasome Activation in Alzheimer’s Disease. Pharmaceuticals 2021, 14, 1187. [Google Scholar] [CrossRef]
- Xie, Q.; Wei, M.; Zhang, B.; Kang, X.; Liu, D.; Zheng, W.; Pan, X.; Quan, Y.; Liao, D.; Shen, J. MicroRNA-33 regulates the NLRP3 inflammasome signaling pathway in macrophages. Mol. Med. Rep. 2017, 17, 3318–3327. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wu, D.; Feng, X.; Shi, Z. Research on the effect of interfering with miRNA-155 on triple-negative breast cancer cells. Genes Genom. 2022, 44, 1117–1124. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| TNBC Classification | Method of Analyses | Number of Patients | Subtypes | Abnormal Mechanisms | Relevant Markers | Therapeutic Strategies | Refs. |
|---|---|---|---|---|---|---|---|
| The Vanderbilt Subtype | K-means clustering | 586 | Basal-like 1 | Cell cycle Cell proliferation DNA damage response | MYC, PIK3CA, CDK6, AKT2, KRAS, FGFR1, IGF1R, CCNE1, CDKN2A/B, BRCA2, PTEN, MDM2, RB1, TP53, KI67 | PARP inhibitors HDAC/DNMT inhibitors Natural-killer therapy Cisplatin, | [20,24,25,26] |
| Basal-like 2 | EGFR, MET, NGF, Wnt/β-catenin, TP63, IGF1R signaling pathway Glycolysis Gluconeogenesis | TP53, TP63, EGFR, MET, BRCA1, RB1, PTEN, CDKN2A, UTX | mTOR inhibitors Growth factor inhibitors (lapatinib, gefitinib, cetuximab, etc.) | [20,24,27] | |||
| Immunomodulatory | Th1/2, IL-7, IL-12 signaling pathway | TP53, CTNNA1, DDX18, HUWE1, NFKBIA, APC, BRAF, MAP K4, RB1, CTLA4, PDL1 | PD1/PDL1/CTLA4 inhibitors Cisplatin PARP inhibitors | [20,24] | |||
| Mesenchymal-like | Cell motility Cell proliferation Cell differentiation Wnt, TGFβ, Notch signaling pathway Epithelial-mesenchymal transition | PTEN, RB1, TP53, PIK3CA, VEGFR2, PI3KCA | mTOR inhibitors Drugs targeting epithelial–mesenchymal transition Abl/Src inhibitor Dasatinib | [20,24,28] | |||
| Mesenchymal stem-like | Cell motility Cell differentiation Growth factor signaling Epithelial–mesenchymal transition Low proliferation | ABCA8, PROCR, ENG, ALDHA1, PER1, ABCB1, TERT2IP, BCL2, BMP2, THY, HOXA5, HOXA10, MEIS1, MEIS2, MEOX1, MEOX2, MSX1, BMP2, ENG, ITGAV, KDR, NGFR, NT5E, PDGFR, THY1, VCAM1, VEGFR2 | mTOR/MEK/PI3K inhibitors, Src antagonists Antiangiogenic drugs Abl/Src inhibitor Dasatinib | [20,24] | |||
| Luminal androgen receptor | Steroid synthesis, porphyrin metabolism, Androgen/estrogen metabolism | DHCR24, CD166, FASN, FKBP5, APOD, PIP, SPDEF, CLDN8 | Anti-AR therapy PI3K/CDK4/6 inhibitors | [20,24,26,29] | |||
| The Baylor Subtype | Non-negative matrix factorization | 198 | Luminal androgen receptor | Steroid hormone biosynthesis Porphyrin and chlorophyll metabolism PPAR signaling pathway Androgen and estrogen metabolism Hormonale-mediated signaling | TP53, PI3KCA, AKT1, ERBB2, ERBB4, CDK4/6, AR, MUC1, ER, CDH1, KRT7, KRT8, KRT18, KRT19, XBP1, FOXA1 | Anti-AR/MUC1 therapy | [21,30,31,32,33,34] |
| Mesenchymal | Cell motility Epithelial–mesenchymal transition Focal adhesion TGF-β signaling pathway Adipocytokine signaling pathway | PIK3CA, PTEN, STAT3, IGF1, prostaglandin, TGF-β, Wnt, β-catenin, PDGFRα, c-Kit, ABC transporter | TKI/RAS/mTOR inhibitor Growth factor inhibitors | [21,30,31,32,33] | |||
| Basal-like immunosuppressed | Mitotic cell cycle Mitotic prometaphase M phase of mitotic cell cycle DNA replication DNA repair Immune response Innate immune response | VTCN1, TP53, CENPF, BUB1, PRC1, VTCN1, MS4A6A, MTBP, FGFR2, BARD1, RNASE6 | VTCN1 inhibition | [21,31] | |||
| Basal-like immune-activated | Cytokine–cytokine receptor interaction T cell receptor signaling pathway B cell receptor signaling pathway Chemokine signaling pathway NF-kB signaling pathway | CCR2, CXCL13, CXCL11, CD1C, CXCL10, CCL5, STAT | Drugs targeting stat signal transduction molecules and cytokines | [21,31,35] | |||
| The French Subtype | Fuzzy clustering | 194 | Cluster 1 | Luminal androgen receptor enriched | AR, Hsp90, PI3K, FGFR4, TTN, TNR, PKHD1L1, SPTA1, NCKAP5, COL15A1, ANKRD11, MYLK | Anti-AR therapy | [36,37,38] |
| Cluster 2 | Basal-like with low immune response High M2-like macrophages High pro-tumorigenic Low anti-tumor immune response | CCL2, CCL5, CCL18, CCL10, CXCL22, IL4, IL8, IL10, IL13, TGFβ1, CD206, CD204, VEGF, Aginase1, PIK3CA, NF1, AKT1, FBN3, ABCC1, DNHD1 | M2 inhibition Repolarization of M2 into M1 macrophages | [20,38,39,40,41,42,43,44,45] | |||
| Cluster 3 | Basal-enriched High immune response Low M2-like macrophages Low pro-tumorigenic High anti-tumor immune response | IL-1β, IL-6, IL-12, IL-23,CXCL9, TNF-α, CCL2, IFNγ, GSF10, DNAH1, CDH23, AHNAK2, GTF3C1 | Repolarization of M2 into M1 macrophages | [38,41,45] |
| Approach | Class of Agents | Examples of Therapy | Mechanism of Action | Refs. |
|---|---|---|---|---|
| Neoadjuvant | Anthracycline + Taxane | Doxorubicin + Cyclophosphamide + Paclitaxel Epirubicin + Cyclophosphamide + Nab-paclitaxel | Inhibition of DNA and RNA synthesis Inhibition of topoisomerase II enzyme Generation of reactive oxygen species (ROS) Stabilization of microtubules | [49,51] |
| Fluoropyrimidine + Taxane | Capecitabine + Docetaxel | |||
| Fluoropyrimidine + Epothilone | Capecitabine + Ixabepilone | |||
| Adjuvant | Anthracycline + Taxane | Doxorubicin + Cyclophosphamide + Docetaxel | [49,52] |
| Class of Agents | Examples of Therapy | Mechanism of Action | Refs. |
|---|---|---|---|
| PD-1 and PD-L1 inhibitors | Pembrolizumab + Paclitaxel Doxorubicin + Cyclophosphamide Pembrolizumab + Paclitaxel + Carboplatin Durvalumab + Nab-paclitaxel Atezolizumab + Nab-paclitaxel Atezolizumab + Nab-paclitaxel + Carboplatin | Reactivation of the anti-tumor immune response PD-1/PD-L1 complex formation inhibition | [55] |
| Platinum-based therapy | Carboplatin + Eribulin Gemcitabine + Carboplatin + Iniparib Carboplatin + Bevacizumab Cisplatin + Paclitaxel + Everolimus Paclitaxel + Carboplatin | Double-strand DNA break Apoptosis initiation | [56,57,58] |
| Cell cycle inhibitors | Trilaciclib, etoposide, abemaciclib, prexasertib | Activate the spindle assembly/mitotic checkpoint Prolonged mitotic arrest Cell death initiation | [59] |
| Angiogenesis inhibitors | Cisplatin + Bevacizumab Anlotinib, apatinib, afatinib, lenvatinib, erlotinib, famitinib, pyrotinib | Blocking new blood vessel formation Tumor growth inhibition VEGF signaling pathway disruption | [60] |
| PI3K/AKT/mTOR inhibitors | Rapamycin, ipatasertip, buparlisib, pictilisib, alpelisib | Cancer cells migration and invasion inhibition Apoptosis initiation | [61,62] |
| PARP inhibitors | Olaparib + Carboplatin + Paclitaxel Veliparib + Carboplatin Cisplatin + Rucaparib Veliparib, niraparib, talazoparib | Double-strand DNA break Cell death initiation Base excision repair Relax/condense chromatin bind nucleosom PARylate H1/H2B | [63] |
| EGFR inhibitors | Bintrafusp Alfa, dasatinib, geftinib, sorafenib, nimotuzumab, panitumumab, erlotinib, osimertinib | Cell death initiation Inhibition of cancer cell proliferation Blocking dimerization of receptors, auto-phosphorylation and downstream signaling Inducing receptor internalization, degradation and stable downregulation | [64] |
| Androgen receptor (AR) antagonists | Bicalutamide, enzalutamide, abiraterone, palbociclib | Decrease in cancer cell viability G1 phase arrest Apoptosis induction | [30,65] |
| Antibody drug conjugates | Sacituzumab govitecan, Ladiratuzumab vedotin, Trastuzumab deruxtecan | Cell growth and migration inhibition Binding to the topoisomerase in DNA replication inhibition S-phase-specific cell death initiation DNA damage | [66] |
| miRNA | Regulation | Targets | Main Biological Mechanisms | Inflammasome Modulation | Refs. |
|---|---|---|---|---|---|
| Oncogenic miRNAs | |||||
| miR-135b | Up-regulated | APC, TGF-β | Promotes proliferation, invasion, migration and metastasis | Alters expression of inflammatory mediators (IL-1R1) Suppresses CASP1 expression following IL-1α stimulation | [86,87] |
| miR-21 | Up-regulated | PTEN | Promotes TNBC cell proliferation Inhibits apoptosis | Modulates NLRP3 phosphorylation. Inhibits the assembly of NLRP3 inflammasomes | [110,111] |
| miR-301b | Up-regulated | CYLD | Promotes cell proliferation Resistance to apoptosis | Targets the TLR4/NF-kBsignaling pathway | [89,112] |
| miR-221 | Up-regulated | FOSL1, MEK, ZEB2 | Decreases the expression of epithelial-specific genes Increases the expression of mesenchymal-specific genes Suppresses oxidative | Suppression of NLRP3/ASC/CASP1 signaling pathway | [113,114] |
| miR-298 | Up-regulated | P-gp | Drug efflux Promotes VEGF signaling Regulates NF-kBsignaling pathway | Regulates the secretion of pro-inflammatory cytokines | [115,116] |
| miR-449 | Up-regulated | E2F1, E2F3, CDK2 | Cell-cycle regulation | Regulation of NLRP3 inflammasome activation | [117,118] |
| miR-302b | Up-regulated | E2F1, ATM | Cell-cycle progression DNA damage repair | Inhibits IL-1β secretion and maturation Regulates the TLR/NF-kB signaling pathway | [119,120] |
| miR-663a | Up-regulated | HSPG2 | Anti-apoptotic activity | Regulates the pro-inflammatory IL-1β expression | [90,91] |
| miR-638 | Up-regulated | BRCA1 | DNA damage repair | Involved in ATP synthesis-coupled electron transport | [92,121] |
| miR-137 | Up-regulated | FSTL1 | Wnt/β-catenin signaling Cellular stemness | Reduces the oxidative stress and inflammation via MAPK signaling pathway Modulates TLR4 expression levels | [121,122] |
| miR-140 | Up-regulated | Wnt/β-catenin | Cellular stemness | Mediates inflammatory cytokines production Modulates TLR4 signaling pathway | [105,123] |
| miR-155 | Up-regulated | CD44, CD90, ABCG2 | Cellular stemness Drug efflux | Inhibition of TLR4/MyD88/NF-kB signaling | [14,124] |
| miR-210 | Up-regulated | HIF1α | Promotes hypoxia | Modulates the necroptosis and pyroptosis in hypoxic conditions | [88,125] |
| miR-9 | Up-regulated | ELAVL1, JAK1 | Inhibition of EMT | Suppression of the NLRP3 inflammasome Inhibition of CASP1 expression and secretion of pro-inflammatory mediators | [11,126,127] |
| miR-181a | Up-regulated | ATG5, ATG22B | Autophagy Cellular stemness | Down-regulates the Bcl-2 expression Modulates the expression of IL-1 | [11,128] |
| miR-373 | Up-regulated | CD44 | Invasion, intravasation, migration and metastasis | Modulates the activation of inflammasome via caspase-8 | [84,85] |
| Tumor-Suppressor miRNAs | |||||
| miR-143-3p | Down-regulated | LIMK1 | Inhibits cell growth, proliferation, migration and invasion | Regulates the MyD88/NF-kB signaling pathway Decreases the level of inflammatory mediators | [129,130] |
| miR-17-5p | Up or down regulated | ETV1, PDCD4, PTEN, DR4 | Inhibits cell proliferation, invasion and apoptosis | Inhibits TXNIP/NLRP3 inflammasome pathway Inhibits pyroptosis | [131,132] |
| miR-217 | Down-regulated | KLF5 | Inhibits cell growth and migration | Modulates oxidative stress and pro-inflammatory cytokines release | [94,133] |
| miR-211-5p | Down-regulated | SETBP1 | Cell proliferation, migration and metatasis | Inhibits pyroptosis Inhibits the expression of CASP1 and caspase-4 | [134,135] |
| miR-185 | Down-regulated | E2F6, DNMT1 | Inhibits cell proliferation | Modulates the expression of NLRP family genes | [76,136] |
| miR-204-5p | Down-regulated | APL1S3 | Promotes cancer cell aggressiveness | Decreases the CASP1 and ASC expression levels Inhibits the activation of NLRP3 inflammasome Suppresses the inflammatory responses | [137,138] |
| miR-128 | Down-regulated | INSR, IRS1 | Inhibits cell proliferation | Constrains the inflammasome assembly Modulates the pyroptotic cell death | [138,139] |
| miR-340 | Down-regulated | SOX2, p16, p27 | Cell cycle | Inhibits pyroptosis and inflammation Mediates the NEK7/NLRP3 signaling pathway | [95,96] |
| miR-384 | Down-regulated | ACVR1 | Inhibits cell proliferation and migration | Regulates inflammation | [109,140] |
| miR-200c | Down-regulated | XIAP | Inhibits cell proliferation promotes apoptosis | Decreases the NLRP3, CASP1, and GSDMD expression levels Inhibits NLRP3 inflammasome activation via NEK7 targeting Inhibits ATP and LPS-induced pyroptosis | [141,142,143] |
| miR-31 | Down-regulated | BCL2, PKCε | Induces apoptosis Inhibits NF-kB signaling Modulates inflammation and oxidative stress | Inhibits caspase-1 activity Decreases NLRP3 and ASC expression levels Inhibits IL-1β and IL-18 releases | [97,98] |
| miR-200a | Down-regulated | PKCα, UBASH3B, XIAP | Suppresses proliferation, migration, invasion, and metastasis Promotes apoptosis | Regulates NOD2 expression | [116,144] |
| miR-141 | Down-regulated | P27, CDK6, STAT5 | Maintenance of epithelial phenotype | Modulates the expression of NLRP family genes | [76,145] |
| miR-27b-3p | Down-regulated | CBLB, GRB2 | Regulation of PI3K/AKT and MAPK/ERK signaling | Attenuates NLRP3, CASP1, GSDMD, IL-1β and IL-18 and expression Anti-pyroptosic effects | [146,147] |
| miR-223 | Down-regulated | HAX-1 | Stimulates apoptosis Blocks the growth and the immunosuppressive ability of TNBC | Inhibits the activity of the NLRP3 inflammasome by binding the 3′-UTR of NLRP3 mRNA | [99,100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
, L.-R.B.; Dinescu, S.; Costache, M. Regulation of Inflammasome by microRNAs in Triple-Negative Breast Cancer: New Opportunities for Therapy. Int. J. Mol. Sci. 2023, 24, 3245. https://doi.org/10.3390/ijms24043245
L-RB, Dinescu S, Costache M. Regulation of Inflammasome by microRNAs in Triple-Negative Breast Cancer: New Opportunities for Therapy. International Journal of Molecular Sciences. 2023; 24(4):3245. https://doi.org/10.3390/ijms24043245
Chicago/Turabian Style(Stămat), Liliana-Roxana Balahura, Sorina Dinescu, and Marieta Costache. 2023. "Regulation of Inflammasome by microRNAs in Triple-Negative Breast Cancer: New Opportunities for Therapy" International Journal of Molecular Sciences 24, no. 4: 3245. https://doi.org/10.3390/ijms24043245
APA Style, L. -R. B., Dinescu, S., & Costache, M. (2023). Regulation of Inflammasome by microRNAs in Triple-Negative Breast Cancer: New Opportunities for Therapy. International Journal of Molecular Sciences, 24(4), 3245. https://doi.org/10.3390/ijms24043245