
How Mechanical and Physicochemical Material Characteristics Influence Adipose-Derived Stem Cell Fate


Abstract
:1. Introduction
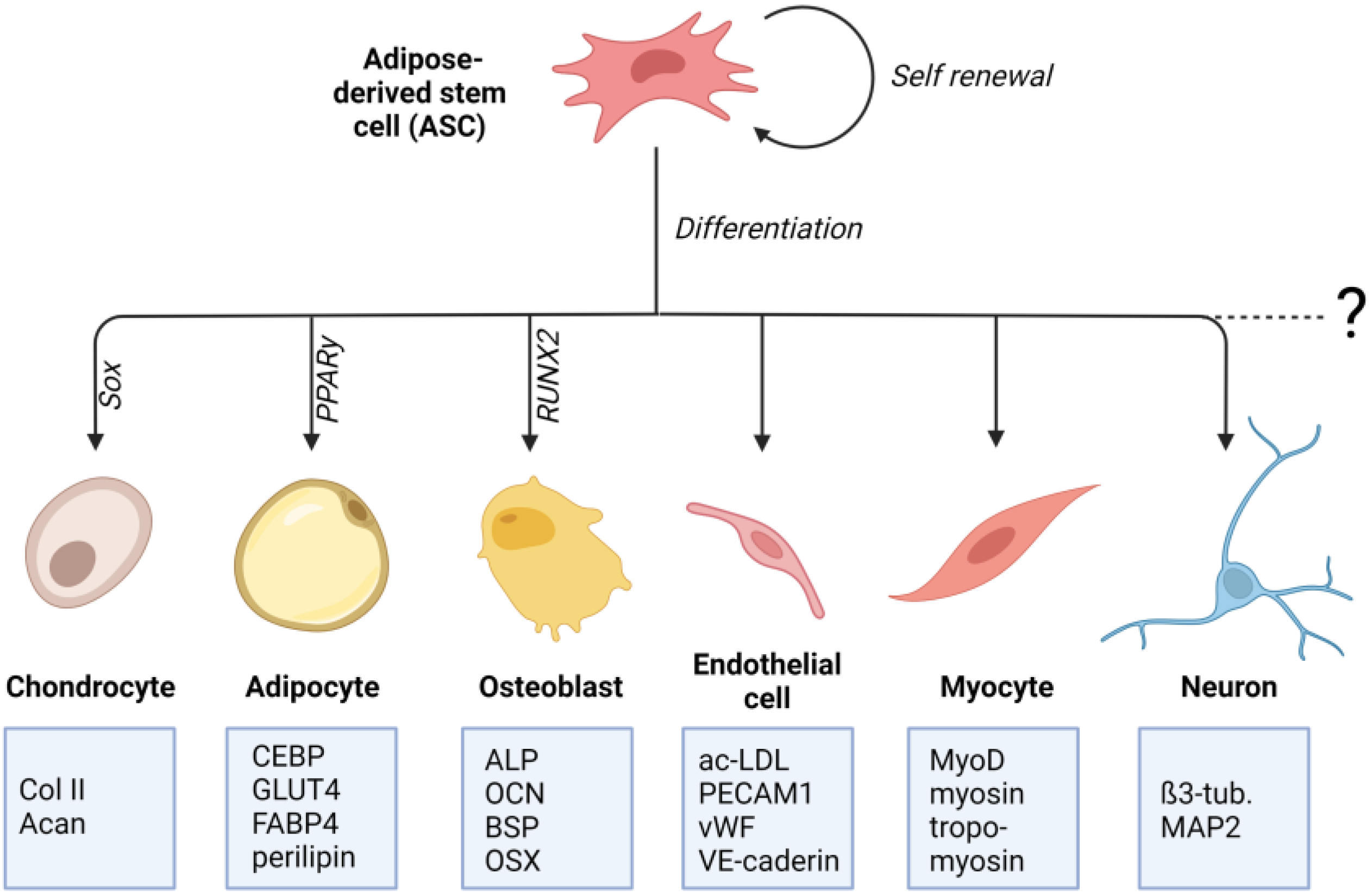
2. Adipose-Derived Stem Cells
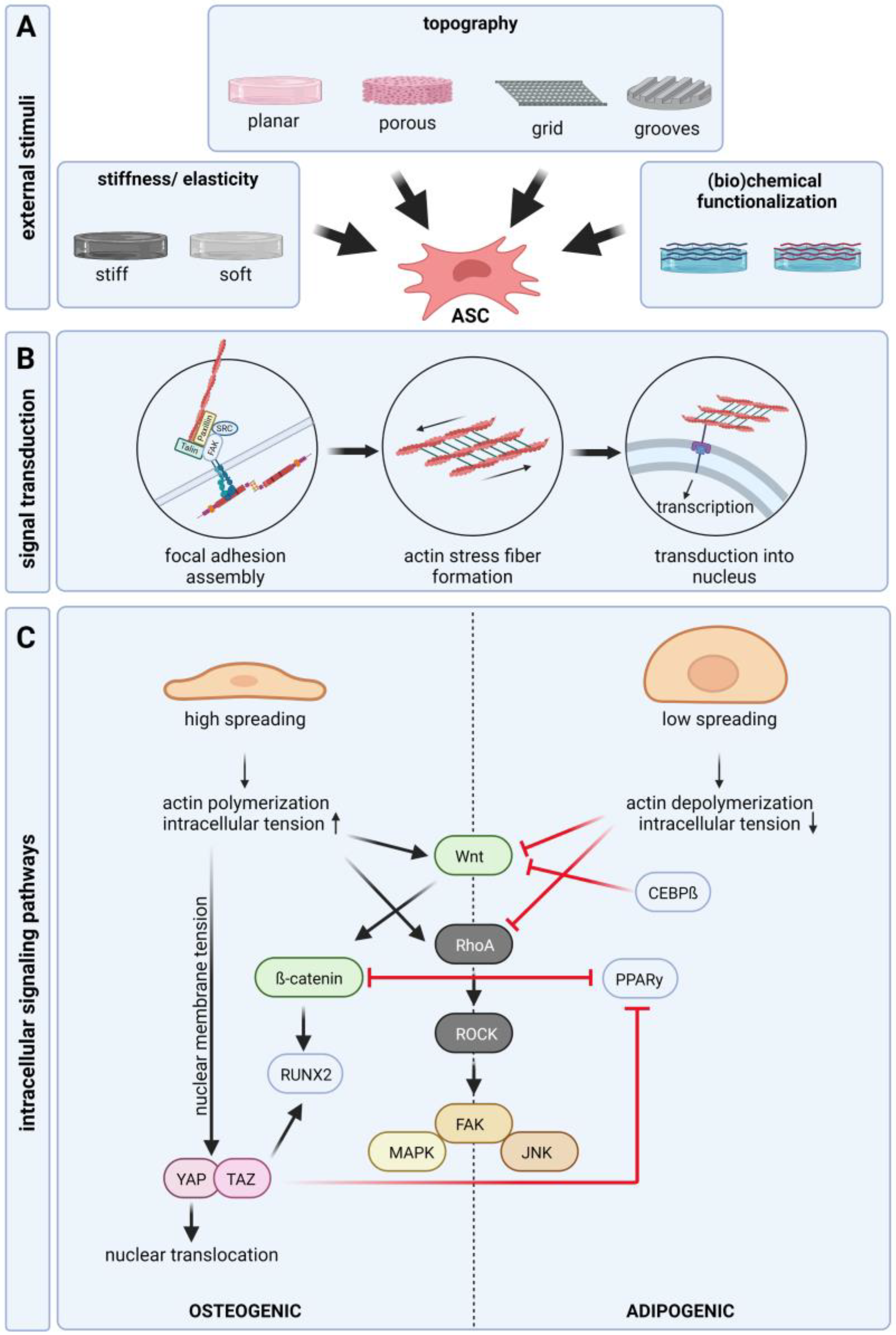
3. Signaling Pathways of Cell-Matrix/Cell-Material Interaction
4. Influence of Material Properties on Adipose-Derived Stem Cell Fate
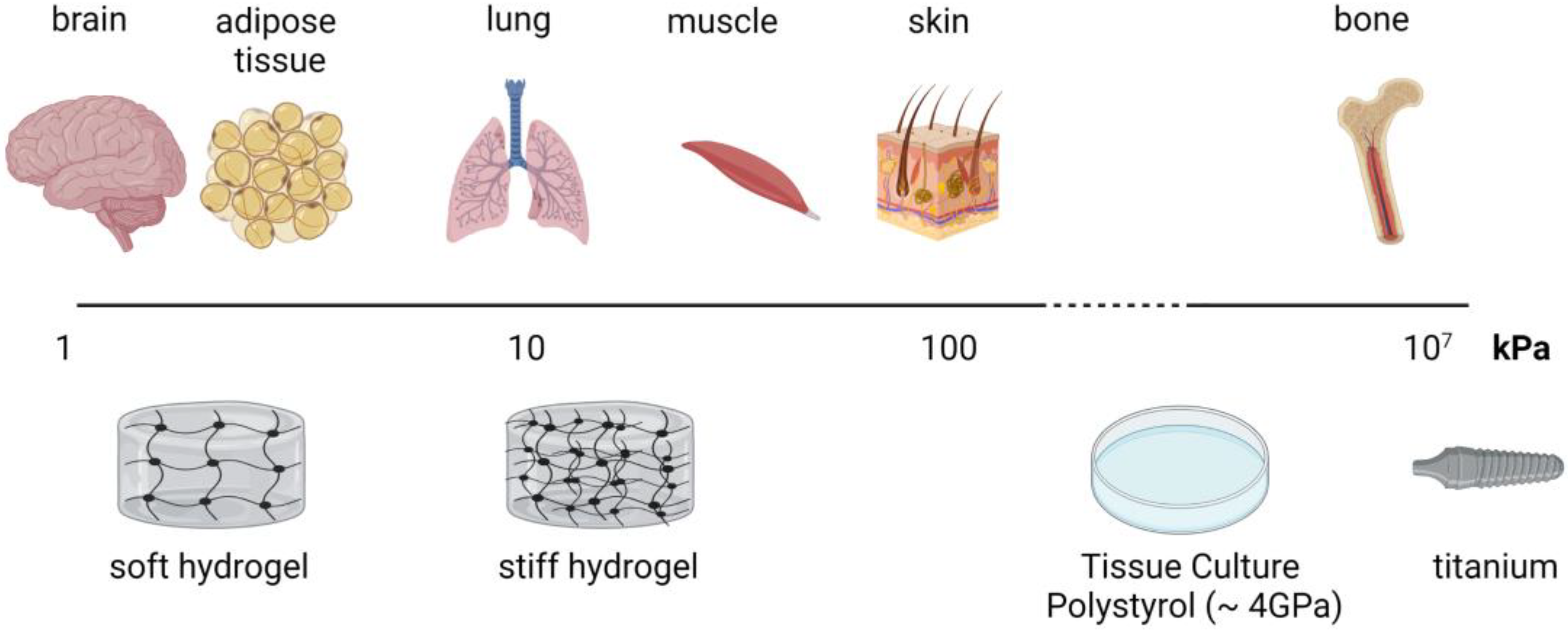
4.1. Stiffness

{kind=link}
{kind=link}
{kind=link}
| Differentiation | Stiffness (kPa) | Soluble Factors | Material | Results | Ref. | |
|---|---|---|---|---|---|---|
| Adipogenic | 1.4–6 | + | PDMS | 2D | Spreading ↓, disorganized actin filaments, Oil Red O ↑, Rho A ↓, ROCK 1/2 ↓, proteins of Wnt/β-catenin pathway ↓ | [93] |
| 0.5 | − | Polyacrylamide | 2D | Spreading ↓, PPARγ n.a., Nile Red ↑ Neuronal: β3 tubulin ↑, MAP2 ↑ | [92] | |
| 2 | − | Adipose ECM functionalized polyacrylamide | 2D | Rounded shape, spreading ↓, PPARγ ↑, CEBP ↑, ap2 ↑ | [98] | |
| 3 | + | Polyacrylamide | 2D | CEBPα ↑ | [81] | |
| ~4 | − | GelMA | 2D | PPARγ ↑ | [89] | |
| 5 | − | Polyacrylamide | 2D | Oil Red O ↑ | [90] | |
| 36 | + | Decellularized adipose tissue | 2D | Proliferation ↑, PPAR γ n.a., LPL ↑, adiponectin ↑; PLIN ↑, perilipin ↑ | [82] | |
| 46 | + | PDMS | 2D | Spreading ↓, β-catenin ↓, Oil Red O ↑, PPARγ ↑, CEBPα ↑ | [62] | |
| Myogenic | ~12 | − | GelMA | 2D | MyoD ↑ | [89] |
| Chondrogenic | 8 | + | Methacylated hyaluronan | 3D | Aggrecan ↑, collagen type II ↑, SOX9 ↓ | [83] |
| Osteogenic | 35 | + | Polyacrylamide | 2D | RUNX2 ↑, ALP ↑ | [81] |
| 37 | − | Polyacrylamide | 2D | ALP ↑, COL1A1 ↑, OCN ↑ | [90] | |
| 53.6–134 | + | PDMS | 2D | Spreading ↑, polygonal shape, bundled actin fibers, Alizarin Red ↑, Rho A ↑, Rock 1/2 ↑, proteins of Wnt/β-catenin pathway ↑ | [93] | |
| 61.8 | + | ELP-collagen | 3D | ALP ↑, osteocalcin ↑, Alizarin Red ↑ | [86] | |
| 660 | + | PTFE/PVA(/GO) | 2D | Alizarin Red ↑, ALP ↑, RUNX2 ↑, osteoclacin ↑, osteonectin ↑ | [99] | |
| 1000 | + | PDMS | 2D | Spreading ↑, β-catenin ↑, ALP ↑, RUNX ↑, OSX ↑ | [62] | |
| 4500 | − | Electrospun PEEU | 2D | Osteocalcin ↑, ALP ↑, hydroxyapatite ↑ | [100] |
4.2. Topography, Geometry, and Pore Size
4.2.1. Nanogrooves and Nanofibers
| Differentiation | Topography | Soluble Factors | Material | Results | Ref. | |
|---|---|---|---|---|---|---|
| Osteogenic | Aligned fibers | + | Tendon | 2D | Focal adhesion ↑, RUNX2 n.a., OPN ↑, COL I ↑, bone regeneration ↑ | [116] |
| Line patterns | + | Graphene oxid | 2D | Spreading ↑, Alizarin Red ↑, ALP ↑, OCN ↑ | [121] | |
| Fibers random | + | PLGA/PLC collagen I | 3D | Randomly orientated, wide-spreading, proliferation ↑, OCN ↑, RUNX2 ↑, OSX ↑, ALP n.a., Col I n.a. | [122] | |
| Nanofibers | + | PCL | 3D | ALP ↑, BMP-2 ↑, RUNX2 ↑, Col I ↑, β-catenin ↑, Smad3 ↑ | [118] | |
| Isotropic fibers | − | PCL | 3D | Proliferation ↑, ALP ↑, Alizarin Red ↑ | [114] | |
| Myogenic | Fibers aligned | + | PCL/PCU | 3D | Proliferation ↑, myosin ↑, tropomyosin ↑ | [123] |
| Endothelial | Grooves | + | Quartz | 2D | Spindle-shaped morphology, proliferation ↑, PECAM-1 ↑, vWF ↑, VE-cadherin ↑, tube formation ↑ | [119] |
| Dual-scale sinusoidal grooves | + | Polystyrene | 2D | Proliferation ↑, ac-LDL ↑ | [120] | |
| Neurogenic | Network pattern | + | Graphene oxid | 2D | Spreading ↓, Tuj-1 ↑, length↑ | [121] |
| Grooves | + | PDA coated polystyrene | 2D | Neurite length ↑, Tuj-1 ↑ | [117] | |
| Tendon | Nanofibers (663.5 nm) | + | PLGA | 3D | Proliferation ↑, SCX ↑, TNC ↑, COL I ↑, TNMD ↑ | [124] |
4.2.2. Tubes, Pores and Pillars
| Differentiation | Topography | Soluble Factors | Material | Results | Ref. | ||
|---|---|---|---|---|---|---|---|
| 15–65 nm | − | TiO2 | 2D | Proliferation ↑, ALP ↑ | [126] | ||
| Osteogenic | Tubes/pores | 70 nm | −/+ | TiO2 | 2D | Adhesion ↑, proliferation ↑, ALP ↑, Alizarin Red ↑, RUNX2 ↑, OC ↑ | [125] |
| 108 nm | + | TiO2 | 2D | Alizarin Red ↑, SP7 ↑, BGLAP ↑, RUNX2 n.a., SPARC n.a., BMP2 n.a., ALP n.a. | [132] | ||
| 112 µm | − | Silk-ECM-carbon nanotube hydrogels | 2D | ALP ↑, RUNX2 ↑, OPN ↑, COL I ↑ | [127] | ||
| Chondrogenic | 100 µm | + | Gelatin hydrogel | 3D | Proliferation ↑, GAG production ↑, aggrecan ↑, SOX9 ↑ | [128] | |
| Adipogenic | 200 nm | + | TCPS | 2D | Oil Red O ↑, FABP ↑, PPARγ ↑, GLUT4 ↑, integrin α6 ↑, integrin α3 ↓ | [102] | |
| Osteogenic | Pillars/nodules | 20 nm | − | Silicon | 2D | spreading ↓, Alizarin Red ↑, osteopontin ↑ | [133] |
| 200 nm | + | TCPS | 2D | Kossa staining ↑, BSP ↑, OCN ↑, RUNX2 ↑, integrin α5 ↑, integrin α2 ↑, integrin α3 ↓ | [102] | ||
| 200 nm | − | Polyetherether-ketone | 2D | Proliferation ↑, ALP ↑, RUNX2 ↑, OPN ↑, OCN ↑, Alizarin Red ↑, | [134] | ||
| 200–400 nm | − | Polystyrene | 2D | 200 nm + 500–750 nm: spreading ↓, viability ↓ 300 + 400 nm: spreading ↑, viability ↑ 200–400 nm: OCN ↑, OPN ↑, ALP ↑, RUNX2 ↑ | [135] | ||
| 530 nm | − | Ti | 2D | Alizarin Red ↑, ALP ↑ | [136] | ||
| 33.8 µm | − | Hydroxyapatite | 2D | Proliferation ↑, ALP ↑, RUNX2 ↑ | [131] | ||
| ? | − | Hydroxyapatite | 2D | Fibronectin absorbtion ↑, ALP ↑, BMP2 ↑, RUNX2 ↑, OCN ↑, OPN ↑, VEGF ↑ | [137] | ||
| osteo-chondrogenic | 92–267 nm | − | BCC | 2D | Spreading ↓, proliferation ↓, BSP ↑, RUNX2 n.a., OPN n.a., AGG ↑, SOX9 ↑, COL II ↑, PPARγ n.a., adiponectin n.a. | [130] | |
| adipogenic/chondrogenic/osteogenic | 15 µm | − | Honey silk fibroin | 2D | Spreading ↑, proliferation ↑, E-cadherin ↑, SOX9 ↑, RUNX2 ↑, PPARγ ↑, p53/p21 ↓, CDH1 ↑, CK-19 ↑ | [129] | |
4.2.3. Cell-Imprints
| Differentiation | Imprint | Soluble Factors | Material | Results | Ref. | |
|---|---|---|---|---|---|---|
| Chondrogenic | Chondrocyte-imprint | − | PDMS | 2D | Col I ↑, Col II ↑, aggrecan ↑, SOX9 ↑, | [138] |
| Keratinocytes | Keratinocyte-imprint | − | PDMS | 2D | Cytokeratin 14 ↑, involucrin ↑, p63 ↑ keratin 10 ↑ | [139,140] |
| Neurogenic | Neuronal-like cell-imprint | − | Chitosan/polyaniline | 2D | Spreading ↑, GFAP ↑, MAP2 ↑ | [143] |
| Osteogenic | Osteoblast-imprint | − | PDMS | 2D | Proliferation ↑, ALP ↑, Alizarin Red ↑, RUNX2 ↑, OCN ↑, Col I ↓ | [141] |
| Neurogenic | Schwann cell-imprint | − | PDMS | 2D | P75 ↑, S100 ↑ | [144] |
4.3. (Bio)Chemical Functionalization
| Differentiation | Functionalization | Soluble Factors | Material | Results | Ref. | |
|---|---|---|---|---|---|---|
| Osteogenic | NH2 | + | polystyrene | 2D | ALP ↑ spreading ↑, RUNX2 ↑ spreading ↑, focal adhesion ↑, ALP ↑, Col I ↑, OSC ↑ | [165,166,167,168,169] |
| Strontium | +/− | Ti HAP | 2D 3D | Focal adhesion ↑, Integrin α2 ↑, Integrin β1/β3 ↑, RhoA ↑, proliferation ↓, ALP ↑, RUNX2 ↑, BSP ↑, OC ↑, Wnt ↑, β-catenin ↑ ALP ↑, BMP2 ↑, RUNX2 ↑, OCN ↑, OX ↑, VEGF ↑ | [153,154] | |
| Zink/Silver and zink/Silver/Copper | − | HAP | 2D | ALP ↑ | [156] | |
| MgO | + | PCL | 3D | Viability ↑, ALP ↑, calcium ↑, RUNX2 ↑, Col I ↑, OPN ↑ | [170] | |
| Graphene | − | Ti | 3D | ALP ↑, bone regeneration ↑ | [171] | |
| Poly-dopamine | + | PLA | 3D | Proliferation ↑, ALP ↑, OC ↑, Alizarin Red ↑, vWF ↑, Ang1 ↑ | [162,163] | |
| CS (+Col I) | + | Glass | 2D | Proliferation ↑, ALP ↑, RUNX2 ↑, Col I ↑ | [158] | |
| Chondrogenic | COOH | + | Polystyrene | 2D | RUNX2 ↑, Collagen II ↑ spreading ↑, focal adhesion ↑, aggrecan ↑, Col II ↑ | [165,169] |
| Ca/Co alginate beads | − | Alginate | 3D | Sox9 ↑, VCAN ↑, Coll II ↑ | [157] | |
| Adipogenic | CH3 | + | Polystyrene | 2D | Spreading ↓, migration ↑, PPARγ ↑, | [169] |
| Myogenic | PLA | − | Glass | 2D | MyoD ↑, Myf5 ↑ | [164] |
| Neurogenic | Laminin | − | Poly-acrylamide | 2D | Β3-tubulin ↑ | [92] |
Extracellular Matrix
5. Conclusions and Further Perspectives
| Differentiation | Material Characteristics |
|---|---|
| Adipogenic | Softer materials (comparable to native tissue), lager pores that allow rounded shape and lipid storage, surface functionalization with methyl groups adipose tissue-derived and pre-adipocyte-derived ECM. |
| Chondrogenic | Material stiffness in the medium range, topography that allows the spheroid formation and chondrocyte imprint, surface functionalization with carboxy groups chondrocyte-derived ECM. |
| Osteogenic | Stiff materials, smaller pores, aligned fiber/grooves and nodular or pillar structures and osteoblast imprint, surface functionalization with amine groups or strontium bone tissue-derived and pre-osteoblast-derived ECM. |
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kshitiz; Park, J.; Kim, P.; Helen, W.; Engler, A.J.; Levchenko, A.; Kim, D.H. Control of stem cell fate and function by engineering physical microenvironments. Integr. Biol. 2012, 4, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-B.; Kim, J.-K.; Lee, G.; Kim, D.-H.; Han, S.-B.; Kim, J.-K.; Lee, G.D.; Kim, D.-H. Mechanical Properties of Materials for Stem Cell Differentiation. Adv. Biosyst. 2020, 4, 2000247. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.U.; Qu, R.; Fan, T.; Ouyang, J.; Dai, J. A glance on the role of actin in osteogenic and adipogenic differentiation of mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-Derived Stem Cells for Regenerative Medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, T.; Song, K.; Fan, X.; Ma, X.; Cui, Z. Adipose-derived stem cell: A better stem cell than BMSC. Cell Biochem. Funct. 2008, 26, 664–675. [Google Scholar] [CrossRef]
- Jung, S.; Panchalingam, K.M.; Rosenberg, L.; Behie, L.A. Ex vivo expansion of human mesenchymal stem cells in defined serum-free media. Stem Cells Int. 2012, 2012, 123030. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Trojahn Kølle, S.F.; Oliveri, R.S.; Glovinski, P.V.; Kirchhoff, M.; Mathiasen, A.B.; Elberg, J.J.; Andersen, P.S.; Drzewiecki, K.T.; Fischer-Nielsen, A. Pooled human platelet lysate versus fetal bovine serum-investigating the proliferation rate, chromosome stability and angiogenic potential of human adipose tissue-derived stem cells intended for clinical use. Cytotherapy 2013, 15, 1086–1097. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Scioli, M.G.; Bielli, A.; Gentile, P.; Mazzaglia, D.; Cervelli, V.; Orlandi, A. The biomolecular basis of adipogenic differentiation of adipose-derived stem cells. Int. J. Mol. Sci. 2014, 15, 6517–6526. [Google Scholar] [CrossRef] [Green Version]
- Mildmay-White, A.; Khan, W. Cell Surface Markers on Adipose-Derived Stem Cells: A Systematic Review. Curr. Stem Cell Res. Ther. 2017, 12, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jia, Z.; Yin, X.; Zhang, X.; Liu, Y.; Chen, P.; Ma, K.; Zhou, C. Comparative Analysis of Mesenchymal Stem Cells from Bone Marrow, Cartilage, and Adipose Tissue. Stem Cells Dev. 2008, 17, 761–773. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and Limits of Adipose-Derived Stem Cells (ADSCs) and Mesenchymal Stem Cells (MSCs) in Wound Healing. Int. J. Mol. Sci. 2020, 21, 1306. [Google Scholar] [CrossRef] [PubMed]
- Trzyna, A.; Banaś-Ząbczyk, A. Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”. Biomolecules 2021, 11, 878. [Google Scholar] [CrossRef]
- Kapur, S.K.; Katz, A.J. Review of the adipose derived stem cell secretome. Biochimie 2013, 95, 2222–2228. [Google Scholar] [CrossRef]
- Li, P.; Guo, X. A review: Therapeutic potential of adipose-derived stem cells in cutaneous wound healing and regeneration 11 Medical and Health Sciences 1103 Clinical Sciences 10 Technology 1004 Medical Biotechnology. Stem Cell Res. Ther. 2018, 9, 302. [Google Scholar] [CrossRef]
- Frese, L.; Dijkman, P.E.; Hoerstrup, S.P. Adipose Tissue-Derived Stem Cells in Regenerative Medicine. Transfus. Med. Hemotherapy 2016, 43, 268–274. [Google Scholar] [CrossRef]
- Alió Del Barrio, J.L.; El Zarif, M.; De Miguel, M.P.; Azaar, A.; Makdissy, N.; Harb, W.; El Achkar, I.; Arnalich-Montiel, F.; Alió, J.L. Cellular Therapy with Human Autologous Adipose-Derived Adult Stem Cells for Advanced Keratoconus. Cornea 2017, 36, 952–960. [Google Scholar] [CrossRef]
- Jurado, M.; De La Mata, C.; Ruiz-García, A.; López-Fernández, E.; Espinosa, O.; Remigia, M.J.; Moratalla, L.; Goterris, R.; García-Martín, P.; Ruiz-Cabello, F.; et al. Adipose tissue-derived mesenchymal stromal cells as part of therapy for chronic graft-versus-host disease: A phase I/II study. Cytotherapy 2017, 19, 927–936. [Google Scholar] [CrossRef]
- Pourmand, G.; Arjmand, B.; Safavi, M.; Heidari, R.; Aghayan, H.; Bazargani, S.T.; Dehghani, S.; Goodarzi, P.; Mohammadi-Jahani, F.; Heidari, F.; et al. Concomitant Transurethral and Transvaginal-Periurethral Injection of Autologous Adipose Derived Stem Cells for Treatment of Female Stress Urinary Incontinence: A Phase One Clinical Trial. Acta Med. Iran. 2017, 19, 368–374. [Google Scholar] [CrossRef]
- Sarveazad, A.; Newstead, G.L.; Mirzaei, R.; Joghataei, M.T.; Bakhtiari, M.; Babahajian, A.; Mahjoubi, B. A new method for treating fecal incontinence by implanting stem cells derived from human adipose tissue: Preliminary findings of a randomized double-blind clinical trial. Stem Cell Res. Ther. 2017, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.A.; Liu, R.S.; Lirng, J.F.; Yang, B.H.; Chang, C.H.; Wang, Y.C.; Wu, Y.S.; Ho, J.H.C.; Lee, O.K.; Soong, B.W. Treatment of Spinocerebellar Ataxia With Mesenchymal Stem Cells: A Phase I/IIa Clinical Study. Cell Transplant. 2017, 26, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.M.; Kapur, S.; Katz, A.J. Characterization of Adipose-Derived Stem Cells: An Update. Curr. Stem Cell Res. Ther. 2010, 5, 95–102. [Google Scholar] [CrossRef]
- Farmer, S.R. Regulation of PPARγ activity during adipogenesis. Int. J. Obes. 2005, 29 (Suppl. 1), S13–S16. [Google Scholar] [CrossRef]
- Bruderer, M.; Richards, R.G.; Alini, M.; Stoddart, M.J. Role and regulation of RUNX2 in osteogenesis. Eur. Cells Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef]
- Yi, S.W.; Kim, H.J.; Oh, H.J.; Shin, H.; Lee, J.S.; Park, J.S.; Park, K.H. Gene expression profiling of chondrogenic differentiation by dexamethasone-conjugated polyethyleneimine with SOX trio genes in stem cells. Stem Cell Res. Ther. 2018, 9, 341. [Google Scholar] [CrossRef]
- Akiyama, H.; Chaboissier, M.C.; Martin, J.F.; Schedl, A.; De Crombrugghe, B. The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef]
- Nishimura, R.; Hata, K.; Ikeda, F.; Ichida, F.; Shimoyama, A.; Matsubara, T.; Wada, M.; Amano, K.; Yoneda, T. Signal transduction and transcriptional regulation during mesenchymal cell differentiation. J. Bone Miner. Metab. 2008, 26, 203–212. [Google Scholar] [CrossRef]
- Frith, J.; Genever, P. Transcriptional control of mesenchymal stem cell differentiation. Transfus. Med. Hemother. 2008, 35, 216–227. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J.; Liu, X.S.; et al. PPARγ and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, R.; Pedersen, T.Å.; Hagenbeek, D.; Moulos, P.; Siersbæk, R.; Megens, E.; Denissov, S.; Børgesen, M.; Francoijs, K.J.; Mandrup, S.; et al. Genome-wide profiling of PPARγ:RXR and RNA polymerase II occupancy reveals temporal activation of distinct metabolic pathways and changes in RXR dimer composition during adipogenesis. Genes Dev. 2008, 22, 2953–2967. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBPα and PPARγ controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ge, C.; Long, J.P.; Begun, D.L.; Rodriguez, J.A.; Goldstein, S.A.; Franceschi, R.T. Biomechanical Stimulation of Osteoblast Gene Expression Requires Phosphorylation of the RUNX2 Transcription Factor. J. Bone Miner. Res. 2012, 27, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; De Crombrugghe, B. The novel zinc finger-containing transcription factor Osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef]
- Olivares-Navarrete, R.; Raz, P.; Zhao, G.; Chen, J.; Wieland, M.; Cochran, D.L.; Chaudhri, R.A.; Ornoy, A.; Boyan, B.D.; Schwartz, Z. Integrin α2β1 plays a critical role in osteoblast response to micron-scale surface structure and surface energy of titanium substrates. Proc. Natl. Acad. Sci. USA 2008, 105, 15767–15772. [Google Scholar] [CrossRef]
- Gronthos, S.; Simmons, P.J.; Graves, S.E.; Robey, P.G. Integrin-mediated interactions between human bone marrow stromal precursor cells and the extracellular matrix. Bone 2001, 28, 174–181. [Google Scholar] [CrossRef]
- Sun, M.; Chi, G.; Li, P.; Lv, S.; Xu, J.; Xu, Z.; Xia, Y.; Tan, Y.; Xu, J.; Li, L.; et al. Effects of Matrix Stiffness on the Morphology, Adhesion, Proliferation and Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Med. Sci. 2018, 15, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; DeYoung, S.M.; Zhang, M.; Zhang, M.; Cheng, A.; Saltiel, A.R. Changes in integrin expression during adipocyte differentiation. Cell Metab. 2005, 2, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Uetaki, M.; Onishi, N.; Oki, Y.; Shimizu, T.; Sugihara, E.; Sampetrean, O.; Watanabe, T.; Yanagi, H.; Suda, K.; Fujii, H.; et al. Regulatory roles of fibronectin and integrin α5 in reorganization of the actin cytoskeleton and completion of adipogenesis. Mol. Biol. Cell 2022, 33, ar78. [Google Scholar] [CrossRef] [PubMed]
- Hamidouche, Z.; Fromigué, O.; Ringe, J.; Häupl, T.; Vaudin, P.; Pagès, J.C.; Srouji, S.; Livne, E.; Marie, P.J. Priming integrin α5 promotes human mesenchymal stromal cell osteoblast differentiation and osteogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 18587. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.; Zhang, X.; Ning, T.; Ma, D.; Ge, Y.; Xu, S.; Hao, Y.; Wu, B. Priming integrin alpha 5 promotes the osteogenic differentiation of human periodontal ligament stem cells due to cytoskeleton and cell cycle changes. J. Proteom. 2018, 179, 122–130. [Google Scholar] [CrossRef]
- Changede, R.; Sheetz, M. Integrin and cadherin clusters: A robust way to organize adhesions for cell mechanics. BioEssays 2017, 39, e201600123. [Google Scholar] [CrossRef]
- Parsons, J.T.; Horwitz, A.R.; Schwartz, M.A. Cell adhesion: Integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. Cell Biol. 2010, 11, 633–643. [Google Scholar] [CrossRef]
- Sneider, A.; Hah, J.; Wirtz, D.; Kim, D.H. Recapitulation of molecular regulators of nuclear motion during cell migration. Cell Adhes. Migr. 2019, 13, 50–62. [Google Scholar] [CrossRef]
- Cho, S.; Irianto, J.; Discher, D.E. Mechanosensing by the nucleus: From pathways to scaling relationships. J. Cell Biol. 2017, 216, 305–315. [Google Scholar] [CrossRef]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Yadav, V.; Sun, L.; Panilaitis, B.; Kaplan, D.L. In vitro chondrogenesis with lysozyme susceptible bacterial cellulose as a scaffold. J. Tissue Eng. Regen. Med. 2015, 9, E276–E288. [Google Scholar] [CrossRef]
- Yao, X.; Peng, R.; Ding, J. Effects of aspect ratios of stem cells on lineage commitments with and without induction media. Biomaterials 2013, 34, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, S.; Yan, C.; Liu, P.; Ding, J. Fabrication of RGD micro/nanopattern and corresponding study of stem cell differentiation. Nano Lett. 2015, 15, 1457–1467. [Google Scholar] [CrossRef]
- Clark, E.A.; Brugge, J.S. Integrins and Signal Transduction Pathways: The Road Taken. Science 1995, 268, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Aiyelabegan, H.T.; Sadroddiny, E. Fundamentals of protein and cell interactions in biomaterials. Biomed. Pharmacother. 2017, 88, 956–970. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.A. Integrins and extracellular matrix in mechanotransduction. Cold Spring Harb. Perspect. Biol. 2010, 2, a005066. [Google Scholar] [CrossRef]
- DeMali, K.A.; Sun, X.; Bui, G.A. Force transmission at cell-cell and cell-matrix adhesions. Biochemistry 2014, 53, 7706–7717. [Google Scholar] [CrossRef]
- Prestwich, T.C.; MacDougald, O.A. Wnt/beta-catenin signaling in adipogenesis and metabolism. Curr. Opin. Cell Biol. 2007, 19, 612–617. [Google Scholar] [CrossRef]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal-Puig, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. 2009, 20, 16–24. [Google Scholar] [CrossRef]
- Pospisilik, J.A.; Schramek, D.; Schnidar, H.; Cronin, S.J.F.; Nehme, N.T.; Zhang, X.; Knauf, C.; Cani, P.D.; Aumayr, K.; Todoric, J.; et al. Drosophila Genome-wide Obesity Screen Reveals Hedgehog as a Determinant of Brown versus White Adipose Cell Fate. Cell 2010, 140, 148–160. [Google Scholar] [CrossRef]
- Cousin, W.; Fontaine, C.; Dani, C.; Peraldi, P. Hedgehog and adipogenesis: Fat and fiction. Biochimie 2007, 89, 1447–1453. [Google Scholar] [CrossRef]
- Chen, L.; Shi, K.; Frary, C.E.; Ditzel, N.; Hu, H.; Qiu, W.; Kassem, M. Inhibiting actin depolymerization enhances osteoblast differentiation and bone formation in human stromal stem cells. Stem Cell Res. 2015, 15, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhang, D.; Zhou, C.; Yuan, Q.; Ye, L.; Zhou, X. Substrate elasticity regulates adipose-derived stromal cell differentiation towards osteogenesis and adipogenesis through β-catenin transduction. Acta Biomater. 2018, 79, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.L.; Rajamannan, N. Diseases of Wnt signaling. Rev. Endocr. Metab. Disord. 2006, 7, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Day, T.F.; Guo, X.; Garrett-Beal, L.; Yang, Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev. Cell 2005, 8, 739–750. [Google Scholar] [CrossRef]
- Takada, I.; Kouzmenko, A.P.; Kato, S. Wnt and PPARγ signaling in osteoblastogenesis and adipogenesis. Nat. Rev. Rheumatol. 2009, 5, 442–447. [Google Scholar] [CrossRef]
- Lecarpentier, Y.; Claes, V.; Duthoit, G.; Hébert, J.L. Circadian rhythms, Wnt/beta-catenin pathway and PPAR alpha/gamma profiles in diseases with primary or secondary cardiac dysfunction. Front. Physiol. 2014, 5, 429. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.S.; Lee, J.S.; Kim, M.; Ahn, B.Y.; Jung, H.S.; Lee, H.M.; Kim, J.W.; Park, K.S. Regulation of Wnt/beta-catenin signaling by CCAAT/enhancer binding protein β during adipogenesis. Obesity 2012, 20, 482–487. [Google Scholar] [CrossRef]
- Krishnan, V.; Bryant, H.U.; MacDougald, O.A. Regulation of bone mass by Wnt signaling. J. Clin. Investig. 2006, 116, 1202–1209. [Google Scholar] [CrossRef]
- Boland, G.M.; Perkins, G.; Hall, D.J.; Tuan, R.S. Wnt 3a promotes proliferation and suppresses osteogenic differentiation of adult human mesenchymal stem cells. J. Cell. Biochem. 2004, 93, 1210–1230. [Google Scholar] [CrossRef]
- Salzig, D.; Leber, J.; Merkewitz, K.; Lange, M.C.; Köster, N.; Czermak, P. Attachment, Growth, and Detachment of Human Mesenchymal Stem Cells in a Chemically Defined Medium. Stem Cells Int. 2016, 2016, 5246584. [Google Scholar] [CrossRef] [Green Version]
- Berrier, A.L.; Yamada, K.M. Cell–matrix adhesion. J. Cell. Physiol. 2007, 213, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.H.; Hwang, E.S.; McManus, M.T.; Amsterdam, A.; Tian, Y.; Kalmukova, R.; Mueller, E.; Benjamin, T.; Spiegelman, B.M.; Sharp, P.A.; et al. TAZ, a transcriptional modulator of mesenchymal stem cell differentiation. Science 2005, 309, 1074–1078. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.J.; Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nat. Cell Biol. 2018, 20, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, P.S.; Loboa, E.G. Cytoskeletal and Focal Adhesion Influences on Mesenchymal Stem Cell Shape, Mechanical Properties, and Differentiation Down Osteogenic, Adipogenic, and Chondrogenic Pathways. Tissue Eng. Part B Rev. 2012, 18, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Wang, L.; Wang, S.; Han, Y.; Wu, J.; Zhang, Q.; Xu, F.; Lu, T.J. Engineering three-dimensional cell mechanical microenvironment with hydrogels. Biofabrication 2012, 4, 042001. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, C.F.; Gasperini, L.; Marques, A.P.; Reis, R.L. The stiffness of living tissues and its implications for tissue engineering. Nat. Rev. Mater. 2020, 5, 351–370. [Google Scholar] [CrossRef]
- Stevens, L.R.; Gilmore, K.J.; Wallace, G.G.; in het Panhuis, M. Tissue engineering with gellan gum. Biomater. Sci. 2016, 4, 1276–1290. [Google Scholar] [CrossRef]
- Pacelli, S.; Paolicelli, P.; Petralito, S.; Subham, S.; Gilmore, D.; Varani, G.; Yang, G.; Lin, D.; Casadei, M.A.; Paul, A. Investigating the Role of Polydopamine to Modulate Stem Cell Adhesion and Proliferation on Gellan Gum-Based Hydrogels. ACS Appl. Bio Mater. 2020, 3, 945–951. [Google Scholar] [CrossRef]
- Albrecht, F.B.; Dolderer, V.; Nellinger, S.; Schmidt, F.F.; Kluger, P.J. Gellan Gum Is a Suitable Biomaterial for Manual and Bioprinted Setup of Long-Term Stable, Functional 3D-Adipose Tissue Models. Gels 2022, 8, 420. [Google Scholar] [CrossRef]
- Kumar, S.; Weaver, V.M. Mechanics, malignancy, and metastasis: The force journey of a tumor cell. Cancer Metastasis Rev. 2009, 28, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Major, L.G.; Choi, Y.S. Developing a high-throughput platform to direct adipogenic and osteogenic differentiation in adipose-derived stem cells. J. Tissue Eng. Regen. Med. 2018, 12, 2021–2028. [Google Scholar] [CrossRef] [PubMed]
- Shridhar, A.; Lam, A.Y.L.; Sun, Y.; Simmons, C.A.; Gillies, E.R.; Flynn, L.E. Culture on Tissue-Specific Coatings Derived from α-Amylase-Digested Decellularized Adipose Tissue Enhances the Proliferation and Adipogenic Differentiation of Human Adipose-Derived Stromal Cells. Biotechnol. J. 2020, 15, 1900118. [Google Scholar] [CrossRef] [PubMed]
- Teong, B.; Wu, S.C.; Chang, C.M.; Chen, J.W.; Chen, H.T.; Chen, C.H.; Chang, J.K.; Ho, M.L. The stiffness of a crosslinked hyaluronan hydrogel affects its chondro-induction activity on hADSCs. J. Biomed. Mater. Res. Part B Appl. Biomater. 2018, 106, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Zigon-Branc, S.; Markovic, M.; Van Hoorick, J.; Van Vlierberghe, S.; Dubruel, P.; Zerobin, E.; Baudis, S.; Ovsianikov, A. Impact of Hydrogel Stiffness on Differentiation of Human Adipose-Derived Stem Cell Microspheroids. Tissue Eng. Part A 2019, 25, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Sarangthem, V.; Singh, T.D.; Dinda, A.K. Emerging Role of Elastin-Like Polypeptides in Regenerative Medicine. Adv. Wound Care 2021, 10, 257–269. [Google Scholar] [CrossRef]
- Gurumurthy, B.; Bierdeman, P.C.; Janorkar, A.V. Composition of elastin like polypeptide-collagen composite scaffold influences in vitro osteogenic activity of human adipose derived stem cells. Dent. Mater. 2016, 32, 1270–1280. [Google Scholar] [CrossRef]
- Newman, K.; Clark, K.; Gurumurthy, B.; Pal, P.; Janorkar, A.V. Elastin-Collagen Based Hydrogels as Model Scaffolds to Induce Three-Dimensional Adipocyte Culture from Adipose Derived Stem Cells. Bioeng. 2020, 7, 110. [Google Scholar] [CrossRef]
- Betre, H.; Ong, S.R.; Guilak, F.; Chilkoti, A.; Fermor, B.; Setton, L.A. Chondrocytic differentiation of human adipose-derived adult stem cells in elastin-like polypeptide. Biomaterials 2006, 27, 91–99. [Google Scholar] [CrossRef]
- Kim, C.; Young, J.L.; Holle, A.W.; Jeong, K.; Major, L.G.; Jeong, J.H.; Aman, Z.M.; Han, D.W.; Hwang, Y.; Spatz, J.P.; et al. Stem Cell Mechanosensation on Gelatin Methacryloyl (GelMA) Stiffness Gradient Hydrogels. Ann. Biomed. Eng. 2020, 48, 893–902. [Google Scholar] [CrossRef]
- Banks, J.M.; Harley, B.A.C.; Bailey, R.C. Tunable, Photoreactive Hydrogel System to Probe Synergies between Mechanical and Biomolecular Cues on Adipose-Derived Mesenchymal Stem Cell Differentiation. ACS Biomater. Sci. Eng. 2015, 1, 718–725. [Google Scholar] [CrossRef]
- Guneta, V.; Loh, Q.L.; Choong, C. Cell-secreted extracellular matrix formation and differentiation of adipose-derived stem cells in 3D alginate scaffolds with tunable properties. J. Biomed. Mater. Res. Part A 2016, 104, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Abdeen, A.A.; Tang, X.; Saif, T.A.; Kilian, K.A. Matrix directed adipogenesis and neurogenesis of mesenchymal stem cells derived from adipose tissue and bone marrow. Acta Biomater. 2016, 42, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lin, S.; Shao, X.; Shi, S.; Zhang, Q.; Xue, C.; Lin, Y.; Zhu, B.; Cai, X. Regulating osteogenesis and adipogenesis in adipose-derived stem cells by controlling underlying substrate stiffness. J. Cell. Physiol. 2018, 233, 3418–3428. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.L.; Cooke, M.E.; Alliston, T. ECM stiffness primes the TGFβ pathway to promote chondrocyte differentiation. Mol. Biol. Cell 2012, 23, 3731–3742. [Google Scholar] [CrossRef]
- Gao, L.; McBeath, R.; Chen, C.S. Stem Cell Shape Regulates a Chondrogenic versus Myogenic Fate through Rac1 and N-cadherin. Stem Cells 2010, 28, 564–572. [Google Scholar] [CrossRef]
- Leight, J.L.; Wozniak, M.A.; Chen, S.; Lynch, M.L.; Chen, C.S. Matrix rigidity regulates a switch between TGF-β1-induced apoptosis and epithelial-mesenchymal transition. Mol. Biol. Cell 2012, 23, 781–791. [Google Scholar] [CrossRef]
- Park, J.S.; Chu, J.S.; Tsou, A.D.; Diop, R.; Tang, Z.; Wang, A.; Li, S. The effect of matrix stiffness on the differentiation of mesenchymal stem cells in response to TGF-β. Biomaterials 2011, 32, 3921–3930. [Google Scholar] [CrossRef]
- Young, D.A.; Choi, Y.S.; Engler, A.J.; Christman, K.L. Stimulation of adipogenesis of adult adipose-derived stem cells using substrates that mimic the stiffness of adipose tissue. Biomaterials 2013, 34, 8581–8588. [Google Scholar] [CrossRef]
- Khoramgah, M.S.; Ranjbari, J.; Abbaszadeh, H.A.; Mirakabad, F.S.T.; Hatami, S.; Hosseinzadeh, S.; Ghanbarian, H. Freeze-dried multiscale porous nanofibrous three dimensional scaffolds for bone regenerations. BioImpacts 2020, 10, 73–85. [Google Scholar] [CrossRef]
- Sun, X.; Tung, W.; Wang, W.; Xu, X.; Zou, J.; Gould, O.E.C.; Kratz, K.; Ma, N.; Lendlein, A. The effect of stiffness variation of electrospun fiber meshes of multiblock copolymers on the osteogenic differentiation of human mesenchymal stem cells. Clin. Hemorheol. Microcirc. 2019, 73, 219–228. [Google Scholar] [CrossRef]
- Ahn, H.H.; Lee, I.W.; Lee, H.B.; Kim, M.S. Cellular Behavior of Human Adipose-Derived Stem Cells on Wettable Gradient Polyethylene Surfaces. Int. J. Mol. Sci. 2014, 15, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Cha, K.J.; Han, I.B.; Shin, D.A.; Cho, D.W.; Lee, S.H.; Kim, D.S. Mass-producible Nano-featured Polystyrene Surfaces for Regulating the Differentiation of Human Adipose-derived Stem Cells. Macromol. Biosci. 2012, 12, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.W.; Tay, L.; Chua, K.H.; Ahmad, R.; Akbar, S.A.; Pingguan-Murphy, B. Proliferation and stemness preservation of human adipose-derived stem cells by surface-modified in situ TiO2 nanofibrous surfaces. Int. J. Nanomed. 2014, 9, 5389–5401. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.S.; Kang, E.H.; Ji, S.; Lee, S.B.; Kim, Y.O.; Yun, I.S.; Yeo, J.S. Quantitative Correlation of Nanotopography with Cell Spreading via Focal Adhesions Using Adipose-Derived Stem Cells. Adv. Biosyst. 2020, 4, 2000092. [Google Scholar] [CrossRef] [PubMed]
- Yim, E.K.F.; Darling, E.M.; Kulangara, K.; Guilak, F.; Leong, K.W. Nanotopography-induced changes in focal adhesions, cytoskeletal organization, and mechanical properties of human mesenchymal stem cells. Biomaterials 2010, 31, 1299–1306. [Google Scholar] [CrossRef]
- Mobasseri, A.; Faroni, A.; Minogue, B.M.; Downes, S.; Terenghi, G.; Reid, A.J. Polymer Scaffolds with Preferential Parallel Grooves Enhance Nerve Regeneration. Tissue Eng. Part A 2015, 21, 1152–1162. [Google Scholar] [CrossRef]
- Howe, A.; Aplin, A.E.; Alahari, S.K.; Juliano, R. Integrin signaling and cell growth control. Curr. Opin. Cell Biol. 1998, 10, 220–231. [Google Scholar] [CrossRef]
- Chrzanowska-Wodnicka, M.; Burridge, K. Rho-stimulated contractility drives the formation of stress fibers and focal adhesions. J. Cell Biol. 1996, 133, 1403–1415. [Google Scholar] [CrossRef]
- Nobes, C.D.; Hall, A. Rho, rac, and cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 1995, 81, 53–62. [Google Scholar] [CrossRef]
- Khatiwala, C.B.; Kim, P.D.; Peyton, S.R.; Putnam, A.J. ECM Compliance Regulates Osteogenesis by Influencing MAPK Signaling Downstream of RhoA and ROCK. J. Bone Miner. Res. 2009, 24, 886–898. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, R.K.; Jaiswal, N.; Bruder, S.P.; Mbalaviele, G.; Marshak, D.R.; Pittenger, M.F. Adult Human Mesenchymal Stem Cell Differentiation to the Osteogenic or Adipogenic Lineage Is Regulated by Mitogen-activated Protein Kinase. J. Biol. Chem. 2000, 275, 9645–9652. [Google Scholar] [CrossRef] [PubMed]
- Klees, R.F.; Salasznyk, R.M.; Kingsley, K.; Williams, W.A.; Boskey, A.; Plopper, G.E. Laminin-5 induces osteogenic gene expression in human mesenchymal stem cells through an ERK-dependent pathway. Mol. Biol. Cell 2005, 16, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Salasznyk, R.M.; Klees, R.F.; Hughlock, M.K.; Plopper, G.E. ERK Signaling Pathways Regulate the Osteogenic Differentiation of Human Mesenchymal Stem Cells on Collagen I and Vitronectin. Cell Commun. Adhes. 2009, 11, 137–153. [Google Scholar] [CrossRef]
- Calejo, I.; Reis, R.L.; Domingues, R.M.A.; Gomes, M.E. Texturing Hierarchical Tissues by Gradient Assembling of Microengineered Platelet-Lysates Activated Fibers. Adv. Healthc. Mater. 2022, 11, 2102076. [Google Scholar] [CrossRef]
- Calejo, I.; Costa-Almeida, R.; Reis, R.L.; Gomes, M.E. A Textile Platform Using Continuous Aligned and Textured Composite Microfibers to Engineer Tendon-to-Bone Interface Gradient Scaffolds. Adv. Healthc. Mater. 2019, 8, 1900200. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.; Alberti, K.; Lee, J.S.; Yang, K.; Jin, Y.; Shin, J.; Yang, H.S.; Xu, Q.; Cho, S.W. Nanostructured tendon-derived scaffolds for enhanced bone regeneration by human adipose-derived stem cells. ACS Appl. Mater. Interfaces 2016, 8, 22819–22829. [Google Scholar] [CrossRef]
- Chen, C.H.; Tsai, C.C.; Wu, P.T.; Wang, I.K.; Yu, J.; Tsai, W.B. Modulation of Neural Differentiation through Submicron-Grooved Topography Surface with Modified Polydopamine. ACS Appl. Bio Mater. 2019, 2, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Qian, Y.; Li, L.; Yao, G.; Yang, L.; Sun, Y. Polycaprolactone nanofiber scaffold enhances the osteogenic differentiation potency of various human tissue-derived mesenchymal stem cells. Stem Cell Res. Ther. 2017, 8, 148. [Google Scholar] [CrossRef]
- Shi, Z.; Neoh, K.G.; Kang, E.T.; Poh, C.K.; Wang, W. Enhanced endothelial differentiation of adipose-derived stem cells by substrate nanotopography. J. Tissue Eng. Regen. Med. 2014, 8, 50–58. [Google Scholar] [CrossRef]
- Kim, H.W.; Lee, J.S.; Park, S.J.; Rhie, J.W.; Kim, D.S. Micro/Nano Dual-Scale Crossed Sinusoidal Wavy Patterns for Synergistic Promotion of Proliferation and Endothelial Differentiation of Human Adipose-Derived Stem Cells. Adv. Mater. Interfaces 2020, 7, 1901983. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Shah, S.; Yang, L.; Yin, P.T.; Hossain, M.K.; Conley, B.; Choi, J.W.; Lee, K.B. Controlling differentiation of adipose-derived stem cells using combinatorial graphene hybrid-pattern arrays. ACS Nano 2015, 9, 3780–3790. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qian, Y.; Xia, Y.; Chen, G.; Dai, Y.; Li, N.; Zhang, F.; Gu, N. Enhanced Osteogenesis of ADSCs by the Synergistic Effect of Aligned Fibers Containing Collagen I. ACS Appl. Mater. Interfaces 2016, 8, 29289–29297. [Google Scholar] [CrossRef] [PubMed]
- Bayati, V.; Altomare, L.; Tanzi, M.C.; Farè, S. Adipose-derived stem cells could sense the nano-scale cues as myogenic-differentiating factors. J. Mater. Sci. Mater. Med. 2013, 24, 2439–2447. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, R.; Zhou, F.; Streubel, P.N.; Chen, S.; Duan, B. Electrospun thymosin β-4 loaded PLGA/PLA nanofiber/microfiber hybrid yarns for tendon tissue engineering application. Mater. Sci. Eng. C 2020, 106, 110268. [Google Scholar] [CrossRef]
- Lv, L.; Liu, Y.; Zhang, P.; Zhang, X.; Liu, J.; Chen, T.; Su, P.; Li, H.; Zhou, Y. The nanoscale geometry of TiO2 nanotubes influences the osteogenic differentiation of human adipose-derived stem cells by modulating H3K4 trimethylation. Biomaterials 2015, 39, 193–205. [Google Scholar] [CrossRef]
- Ehlert, M.; Radtke, A.; Jedrzejewski, T.; Roszek, K.; Bartmanski, M.; Piszczek, P. In vitro studies on nanoporous, nanotubular and nanosponge-like titania coatings, with the use of adipose-derived stem cells. Materials 2020, 13, 1574. [Google Scholar] [CrossRef] [PubMed]
- Lemos, R.; Maia, F.R.; Ribeiro, V.P.; Costa, J.B.; Coutinho, P.J.G.; Reis, R.L.; Oliveira, J.M. Carbon nanotube-reinforced cell-derived matrix-silk fibroin hierarchical scaffolds for bone tissue engineering applications. J. Mater. Chem. B 2021, 9, 9561–9574. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.C.; Chen, I.H.; Yang, Y.T.; Hsiao, J.K.; Wang, C.C. Effects of scaffold geometry on chondrogenic differentiation of adipose-derived stem cells. Mater. Sci. Eng. C 2020, 110, 110733. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Das, A.; Mukherjee, S.; Rajput, M.; Gope, A.; Chaudhary, A.; Choudhury, K.; Barui, A.; Chatterjee, J.; Mukherjee, R. Improved Mesenchymal Stem Cell Proliferation, Differentiation, Epithelial Transition, and Restrained Senescence on Hierarchically Patterned Porous Honey Silk Fibroin Scaffolds. ACS Appl. Bio Mater. 2021, 4, 4328–4344. [Google Scholar] [CrossRef]
- Wang, P.Y.; Thissen, H.; Kingshott, P. Stimulation of Early Osteochondral Differentiation of Human Mesenchymal Stem Cells Using Binary Colloidal Crystals (BCCs). ACS Appl. Mater. Interfaces 2016, 8, 4477–4488. [Google Scholar] [CrossRef]
- Ramaswamy, Y.; Roohani, I.; No, Y.J.; Madafiglio, G.; Chang, F.; Zhang, F.; Lu, Z.; Zreiqat, H. Nature-inspired topographies on hydroxyapatite surfaces regulate stem cells behaviour. Bioact. Mater. 2021, 6, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Malec, K.; Góralska, J.; Hubalewska-Mazgaj, M.; Głowacz, P.; Jarosz, M.; Brzewski, P.; Sulka, G.D.; Jaskuła, M.; Wybrańska, I. Effects of nanoporous anodic titanium oxide on human adipose derived stem cells. Int. J. Nanomed. 2016, 11, 5349–5360. [Google Scholar] [CrossRef] [PubMed]
- Brammer, K.S.; Choi, C.; Frandsen, C.J.; Oh, S.; Jin, S. Hydrophobic nanopillars initiate mesenchymal stem cell aggregation and osteo-differentiation. Acta Biomater. 2011, 7, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Feng, Z.; Hu, Y.; Zhao, D.; Guo, X.; Du, F.; Wang, N.; Sun, C.; Liu, C.; Liu, H. Endowing Polyetheretherketone Implants with Osseointegration Properties: In Situ Construction of Patterned Nanorod Arrays. Small 2022, 18, 2105589. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Song, X.; Lu, X. Directional osteo-differentiation effect of hadscs on nanotopographical self-assembled polystyrene nanopit surfaces. Int. J. Nanomed. 2020, 15, 3281–3290. [Google Scholar] [CrossRef]
- Martel-Frachet, V.; Ivanova, E.P.; Le Clainche, T.; Linklater, D.; Wong, S.; Le, P.; Juodkazis, S.; Le Guevel, X.; Coll, J.L. Mechano-bactericidal titanium surfaces for bone tissue engineering. ACS Appl. Mater. Interfaces 2020, 12, 48272–48283. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, L.; Gao, H.; Shi, X.; Wang, Y. In Situ Formation of Hexagon-like Column Array Hydroxyapatite on 3D-Plotted Hydroxyapatite Scaffolds by Hydrothermal Method and Its Effect on Osteogenic Differentiation. ACS Appl. Bio Mater. 2020, 3, 1753–1760. [Google Scholar] [CrossRef]
- Bonakdar, S.; Mahmoudi, M.; Montazeri, L.; Taghipoor, M.; Bertsch, A.; Shokrgozar, M.A.; Sharifi, S.; Majidi, M.; Mashinchian, O.; Sekachaei, M.H.; et al. Cell-Imprinted Substrates Modulate Differentiation, Redifferentiation, and Transdifferentiation. ACS Appl. Mater. Interfaces 2016, 8, 13777–13784. [Google Scholar] [CrossRef]
- Keyhanvar, N.; Zarghami, N.; Seifalian, A.; Keyhanvar, P.; Sarvari, R.; Salehi, R.; Rahbarghazi, R.; Ranjkesh, M.; Akbarzadeh, M.; Mahdipour, M.; et al. The Combined Thermoresponsive Cell-Imprinted Substrate, Induced Differentiation, and “KLC Sheet” Formation. Adv. Pharm. Bull. 2022, 12, 356–365. [Google Scholar] [CrossRef]
- Mashinchian, O.; Bonakdar, S.; Taghinejad, H.; Satarifard, V.; Heidari, M.; Majidi, M.; Sharifi, S.; Peirovi, A.; Saffar, S.; Taghinejad, M.; et al. Cell-imprinted substrates act as an artificial niche for skin regeneration. ACS Appl. Mater. Interfaces 2014, 6, 13280–13292. [Google Scholar] [CrossRef]
- Babaei, M.; Nasernejad, B.; Sharifikolouei, E.; Shokrgozar, M.A.; Bonakdar, S. Bioactivation of 3D Cell-Imprinted Polydimethylsiloxane Surfaces by Bone Protein Nanocoating for Bone Tissue Engineering. ACS Omega 2022, 7, 26353–26367. [Google Scholar] [CrossRef] [PubMed]
- Nazbar, A.; Samani, S.; Yazdian Kashani, S.; Amanzadeh, A.; Shoeibi, S.; Bonakdar, S. Molecular imprinting as a simple way for the long-term maintenance of the stemness and proliferation potential of adipose-derived stem cells: An in vitro study. J. Mater. Chem. B 2022, 10, 6816–6830. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, B.S.; Eskandari, M.; Janmey, P.A.; Samadikuchaksaraei, A.; Gholipourmalekabadi, M. Conductive chitosan/polyaniline hydrogel with cell-imprinted topography as a potential substrate for neural priming of adipose derived stem cells. RSC Adv. 2021, 11, 15795–15807. [Google Scholar] [CrossRef] [PubMed]
- Dadashkhan, S.; Irani, S.; Bonakdar, S.; Ghalandari, B. P75 and S100 gene expression induced by cell-imprinted substrate and beta-carotene to nerve tissue engineering. J. Appl. Polym. Sci. 2021, 138, 50624. [Google Scholar] [CrossRef]
- Webb, K.; Hlady, V.; Tresco, P.A. Relationships among cell attachment, spreading, cytoskeletal organization, and migration rate for anchorage-dependent cells on model surfaces. J. Biomed. Mater. Res. 2000, 49, 362–368. [Google Scholar] [CrossRef]
- Dalton, B.A.; Mc Farland, C.D.; Gengenbach, T.R.; Griesser, H.J.; Steele, J.G. Polymer surface chemistry and bone cell migration. J. Biomater. Sci. Polym. Ed. 2012, 9, 781–799. [Google Scholar] [CrossRef]
- Yang, J.; Shi, G.; Bei, J.; Wang, S.; Cao, Y.; Shang, Q.; Yang, G.; Wang, W. Fabrication and surface modification of macroporous poly(L-lactic acid) and poly(L-lactic-co-glycolic acid) (70/30) cell scaffolds for human skin fibroblast cell culture. J. Biomed. Mater. Res. 2002, 62, 438–446. [Google Scholar] [CrossRef]
- Kleinhans, C.; Schmohl, L.; Barz, J.; Kluger, P.J. Low-pressure plasma activation enables enhanced adipose-derived stem cell adhesion. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 1527–1535. [Google Scholar] [CrossRef]
- Firkowska-Boden, I.; Zhang, X.; Jandt, K.D. Controlling Protein Adsorption through Nanostructured Polymeric Surfaces. Adv. Healthc. Mater. 2018, 7, 1700995. [Google Scholar] [CrossRef]
- Wilson, C.J.; Clegg, R.E.; Leavesley, D.I.; Pearcy, M.J. Mediation of biomaterial-cell interactions by adsorbed proteins: A review. Tissue Eng. 2005, 11, 1–18. [Google Scholar] [CrossRef]
- Daum, R.; Mrsic, I.; Hutterer, J.; Junginger, A.; Hinderer, S.; Meixner, A.J.; Gauglitz, G.; Chassé, T.; Schenke-Layland, K. Fibronectin adsorption on oxygen plasma-treated polyurethane surfaces modulates endothelial cell response. J. Mater. Chem. B 2021, 9, 1647–1660. [Google Scholar] [CrossRef] [PubMed]
- Aimaiti, A.; Maimaitiyiming, A.; Boyong, X.; Aji, K.; Li, C.; Cui, L. Low-dose strontium stimulates osteogenesis but high-dose doses cause apoptosis in human adipose-derived stem cells via regulation of the ERK1/2 signaling pathway. Stem Cell Res. Ther. 2017, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, Y.J.; Jang, J.H.; Park, J.W. Surface Engineering of Nanostructured Titanium Implants with Bioactive Ions. J. Dent. Res. 2016, 95, 558–565. [Google Scholar] [CrossRef]
- Wei, Y.; Gao, H.; Hao, L.; Shi, X.; Wang, Y. Constructing a Sr2+-substituted surface hydroxyapatite hexagon-like microarray on 3d-plotted hydroxyapatite scaffold to regulate osteogenic differentiation. Nanomaterials 2020, 10, 1672. [Google Scholar] [CrossRef] [PubMed]
- El-Rashidy, A.A.; El Moshy, S.; Radwan, I.A.; Rady, D.; Abbass, M.M.S.; Dörfer, C.E.; El-Sayed, K.M.F. Effect of Polymeric Matrix Stiffness on Osteogenic Differentiation of Mesenchymal Stem/Progenitor Cells: Concise Review. Polymers 2021, 13, 2950. [Google Scholar] [CrossRef] [PubMed]
- Bostancioglu, R.B.; Gurbuz, M.; Akyurekli, A.G.; Dogan, A.; Koparal, A.S.; Koparal, A.T. Adhesion profile and differentiation capacity of human adipose tissue derived mesenchymal stem cells grown on metal ion (Zn, Ag and Cu) doped hydroxyapatite nano-coated surfaces. Colloids Surf. B Biointerfaces 2017, 155, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Focaroli, S.; Teti, G.; Salvatore, V.; Orienti, I.; Falconi, M. Calcium/Cobalt Alginate Beads as Functional Scaffolds for Cartilage Tissue Engineering. Stem Cells Int. 2016, 2016, 2030478. [Google Scholar] [CrossRef]
- Zhao, M.; Altankov, G.; Grabiec, U.; Bennett, M.; Salmeron-Sanchez, M.; Dehghani, F.; Groth, T. Molecular composition of GAG-collagen I multilayers affects remodeling of terminal layers and osteogenic differentiation of adipose-derived stem cells. Acta Biomater. 2016, 41, 86–99. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, T.; Liu, J. Recent Development of Polydopamine Anti-Bacterial Nanomaterials. Int. J. Mol. Sci. 2022, 23, 7278. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, L.; Zhang, J.; Hu, J.; Duan, G.; Liu, X.; Li, Y.; Gu, Z. Polydopamine antibacterial materials. Mater. Horiz. 2021, 8, 1618–1633. [Google Scholar] [CrossRef]
- Li, Y.; Li, C.; Yu, R.; Ding, Y. Application of polydopamine on the implant surface modification. Polym. Bull. 2022, 79, 5613–5633. [Google Scholar] [CrossRef]
- Kao, C.T.; Lin, C.C.; Chen, Y.W.; Yeh, C.H.; Fang, H.Y.; Shie, M.Y. Poly(dopamine) coating of 3D printed poly(lactic acid) scaffolds for bone tissue engineering. Mater. Sci. Eng. C 2015, 56, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Fu, S.J. Osteogenesis of human adipose-derived stem cells on poly(dopamine)-coated electrospun poly(lactic acid) fiber mats. Mater. Sci. Eng. C 2016, 58, 254–263. [Google Scholar] [CrossRef]
- Foldberg, S.; Petersen, M.; Fojan, P.; Gurevich, L.; Fink, T.; Pennisi, C.P.; Zachar, V. Patterned poly(lactic acid) films support growth and spontaneous multilineage gene expression of adipose-derived stem cells. Colloids Surf. B Biointerfaces 2012, 93, 92–99. [Google Scholar] [CrossRef]
- Griffin, M.F.; Ibrahim, A.; Seifalian, A.M.; Butler, P.E.M.; Kalaskar, D.M.; Ferretti, P. Chemical group-dependent plasma polymerisation preferentially directs adipose stem cell differentiation towards osteogenic or chondrogenic lineages. Acta Biomater. 2017, 50, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, Q.; Bachhuka, A.; Vasilev, K. Surface modification by allylamine plasma polymerization promotes osteogenic differentiation of human adipose-derived stem cells. ACS Appl. Mater. Interfaces 2014, 6, 9733–9741. [Google Scholar] [CrossRef]
- Chaves, C.; Alshomer, F.; Palgrave, R.G.; Kalaskar, D.M. Plasma Surface Modification of Polyhedral Oligomeric Silsequioxane-Poly(carbonate-urea) Urethane with Allylamine Enhances the Response and Osteogenic Differentiation of Adipose-Derived Stem Cells. ACS Appl. Mater. Interfaces 2016, 8, 18701–18709. [Google Scholar] [CrossRef]
- Liu, X.; Shi, S.; Feng, Q.; Bachhuka, A.; He, W.; Huang, Q.; Zhang, R.; Yang, X.; Vasilev, K. Surface Chemical Gradient Affects the Differentiation of Human Adipose-Derived Stem Cells via ERK1/2 Signaling Pathway. ACS Appl. Mater. Interfaces 2015, 7, 18473–18482. [Google Scholar] [CrossRef]
- Chieh, H.F.; Su, F.C.; Lin, S.C.; Shen, M.R.; Liao, J. Der Migration patterns and cell functions of adipose-derived stromal cells on self-assembled monolayers with different functional groups. J. Biomater. Sci. Polym. Ed. 2013, 24, 94–117. [Google Scholar] [CrossRef]
- Niknam, Z.; Golchin, A.; Rezaei-Tavirani, M.; Ranjbarvan, P.; Zali, H.; Omidi, M.; Mansouri, V. Osteogenic Differentiation Potential of Adipose-Derived Mesenchymal Stem Cells Cultured on Magnesium Oxide/Polycaprolactone Nanofibrous Scaffolds for Improving Bone Tissue Reconstruction. Adv. Pharm. Bull. 2022, 12, 142–154. [Google Scholar] [CrossRef]
- Sun, X.; Tong, S.; Yang, S.; Guo, S. The effects of graphene on the biocompatibility of a 3D-printed porous Titanium alloy. Coatings 2021, 11, 1509. [Google Scholar] [CrossRef]
- Nellinger, S.; Schmidt, I.; Heine, S.; Volz, A.C.; Kluger, P.J. Adipose stem cell-derived extracellular matrix represents a promising biomaterial by inducing spontaneous formation of prevascular-like structures by mvECs. Biotechnol. Bioeng. 2020, 117, 3160–3172. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Kim, G.H. A bioprinted complex tissue model for myotendinous junction with biochemical and biophysical cues. Bioeng. Transl. Med. 2022, 7, e10321. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Qi, J.; Wang, Q.; Qu, Y.; Fu, S.; Luan, J. Investigating the Adipogenic Effects of Different Tissue-Derived Decellularized Matrices. Front. Bioeng. Biotechnol. 2022, 10, 872897. [Google Scholar] [CrossRef]
- Zhao, Y.; Fan, J.; Bai, S. Biocompatibility of injectable hydrogel from decellularized human adipose tissue in vitro and in vivo. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 1684–1694. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; McCann, J.; Ravnic, D.J.; Gimble, J.M.; Hayes, D.J. Hybrid adipose graft materials synthesized from chemically modified adipose extracellular matrix. J. Biomed. Mater. Res. Part A 2022, 110, 156–163. [Google Scholar] [CrossRef]
- Ruff, S.M.; Keller, S.; Wieland, D.E.; Wittmann, V.; Tovar, G.E.M.; Bach, M.; Kluger, P.J. clickECM: Development of a cell-derived extracellular matrix with azide functionalities. Acta Biomater. 2017, 52, 159–170. [Google Scholar] [CrossRef]
- Nellinger, S.; Rapp, M.A.; Southan, A.; Wittmann, V.; Kluger, P.J. An Advanced “clickECM” That Can be Modified by the Inverse-Electron-Demand Diels-Alder Reaction. Chembiochem 2022, 23, e202100266. [Google Scholar] [CrossRef]
- Yeleswarapu, S.; Chameettachal, S.; Bera, A.K.; Pati, F. Smooth muscle matrix bioink promotes myogenic differentiation of encapsulated adipose-derived stem cells. J. Biomed. Mater. Res. Part A 2022, 110, 1761–1773. [Google Scholar] [CrossRef]
- Mohiuddin, O.A.; O’Donnell, B.T.; Poche, J.N.; Iftikhar, R.; Wise, R.M.; Motherwell, J.M.; Campbell, B.; Savkovic, S.D.; Bunnell, B.A.; Hayes, D.J.; et al. Human adipose-derived hydrogel characterization based on in vitro ASC biocompatibility and differentiation. Stem Cells Int. 2019, 2019, 9276398. [Google Scholar] [CrossRef] [Green Version]
- Shridhar, A.; Amsden, B.G.; Gillies, E.R.; Flynn, L.E. Investigating the Effects of Tissue-Specific Extracellular Matrix on the Adipogenic and Osteogenic Differentiation of Human Adipose-Derived Stromal Cells Within Composite Hydrogel Scaffolds. Front. Bioeng. Biotechnol. 2019, 7, 402. [Google Scholar] [CrossRef] [PubMed]
- Coronado, R.E.; Somaraki-Cormier, M.; Ong, J.L.; Halff, G.A. Hepatocyte-like cells derived from human amniotic epithelial, bone marrow, and adipose stromal cells display enhanced functionality when cultured on decellularized liver substrate. Stem Cell Res. 2019, 38, 101471. [Google Scholar] [CrossRef]
- Asadi, M.; Lotfi, H.; Salehi, R.; Mehdipour, A.; Zarghami, N.; Akbarzadeh, A.; Alizadeh, E. Hepatic cell-sheet fabrication of differentiated mesenchymal stem cells using decellularized extracellular matrix and thermoresponsive polymer. Biomed. Pharmacother. 2021, 134, 111096. [Google Scholar] [CrossRef] [PubMed]
- Guneta, V.; Zhou, Z.; Tan, N.S.; Sugii, S.; Wong, M.T.C.; Choong, C. Recellularization of decellularized adipose tissue-derived stem cells: Role of the cell-secreted extracellular matrix in cellular differentiation. Biomater. Sci. 2018, 6, 168–178. [Google Scholar] [CrossRef]
- Zhang, Z.; Qu, R.; Fan, T.; Ouyang, J.; Lu, F.; Dai, J. Stepwise Adipogenesis of Decellularized Cellular Extracellular Matrix Regulates Adipose Tissue-Derived Stem Cell Migration and Differentiation. Stem Cells Int. 2019, 2019, 1845926. [Google Scholar] [CrossRef]
- Blum, J.C.; Schenck, T.L.; Birt, A.; Giunta, R.E.; Wiggenhauser, P.S. Artificial decellularized extracellular matrix improves the regenerative capacity of adipose tissue derived stem cells on 3D printed polycaprolactone scaffolds. J. Tissue Eng. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Chen, H.; Pan, T.; Li, T.; Zhang, Z.; Lv, X.; Wang, J.; Ji, Z.; He, Y.; Li, L.; et al. Autologous decellularized extracellular matrix promotes adipogenic differentiation of adipose derived stem cells in low serum culture system by regulating the ERK1/2-PPARγ pathway. Adipocyte 2021, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]



| Tissue/Cell-Source | Soluble Factors | Material | Results | Ref. | |
|---|---|---|---|---|---|
| Native ECM | Muscle tissue | − | Bioink | Myogenic: α-smooth muscle actin ↑, myosin ↑ | [179] |
| − | Coating/bioink | Myogenic: MyoD ↑, Myh2 ↑ | [173] | ||
| Adipose tissue | − | Hydrogel | Adipogenic: Proliferation ↑, Oil Red O ↑ | [175] | |
| + | Hydrogel | Adipogenic: adiponectin ↑, FABP4 ↑, PPARγ ↑ | [82,176,180,181] | ||
| Liver tissue | + | Hepatocyte-like: AFP ↑, PCK ↑, CYP ↑, CK-18 ↑ | [182,183] | ||
| Dermis | − | Hydrogel | Adipogenic: PPARγ ↑ | [174] | |
| Tendon | − | Coating/bioink | Tendon: Scx ↑, Tnmd ↑ | [173] | |
| Bone | + | Hydrogel | Osteogenic: ALP ↑ | [181] | |
| Cell-derived ECM | Pre-adipocytes | − | Coating | Adipogenic: PPARγ ↑, adiponectin ↑ | [184] |
| + | Coating | Adipogenic: PPARγ ↑, FABP4 ↑ | [185] | ||
| Pre-chondrocytes | + | Coating | Chondrogenic: Col IX ↑ | [186] | |
| Pre-osteoblasts | − | Coating | Osteogenic: Acan ↑, Col I ↑, ALP ↑, SPP1 ↑, Col IX ↑ | [186] | |
| Adipose-derived stem cells | + | Coating | Adipogenic: PPARγ ↑, CEBPα ↑ | [187] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nellinger, S.; Kluger, P.J. How Mechanical and Physicochemical Material Characteristics Influence Adipose-Derived Stem Cell Fate. Int. J. Mol. Sci. 2023, 24, 3551. https://doi.org/10.3390/ijms24043551
Nellinger S, Kluger PJ. How Mechanical and Physicochemical Material Characteristics Influence Adipose-Derived Stem Cell Fate. International Journal of Molecular Sciences. 2023; 24(4):3551. https://doi.org/10.3390/ijms24043551
Chicago/Turabian StyleNellinger, Svenja, and Petra Juliane Kluger. 2023. "How Mechanical and Physicochemical Material Characteristics Influence Adipose-Derived Stem Cell Fate" International Journal of Molecular Sciences 24, no. 4: 3551. https://doi.org/10.3390/ijms24043551
APA StyleNellinger, S., & Kluger, P. J. (2023). How Mechanical and Physicochemical Material Characteristics Influence Adipose-Derived Stem Cell Fate. International Journal of Molecular Sciences, 24(4), 3551. https://doi.org/10.3390/ijms24043551