
MEIS1 Is a Common Transcription Repressor of the miR-23a and NORHA Axis in Granulosa Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
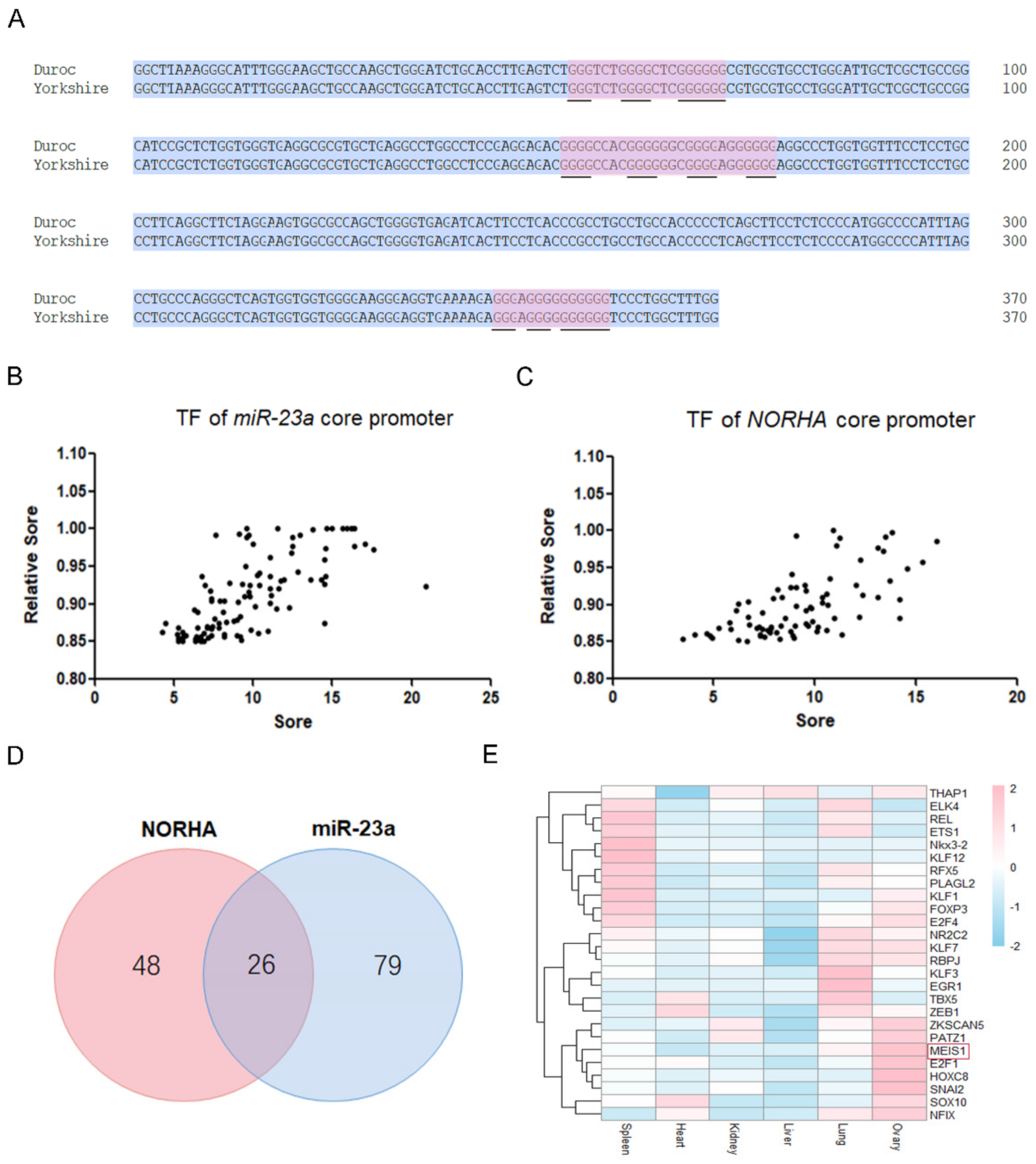
2.1. The miR-23a and NORHA Axis Share Potential Common Transcription Factors
2.2. MEIS1 Is a Transcription Factor Related to Sow Follicular Atresia
2.3. MEIS1 Is a Novel Suppressor of Apoptosis in GCs
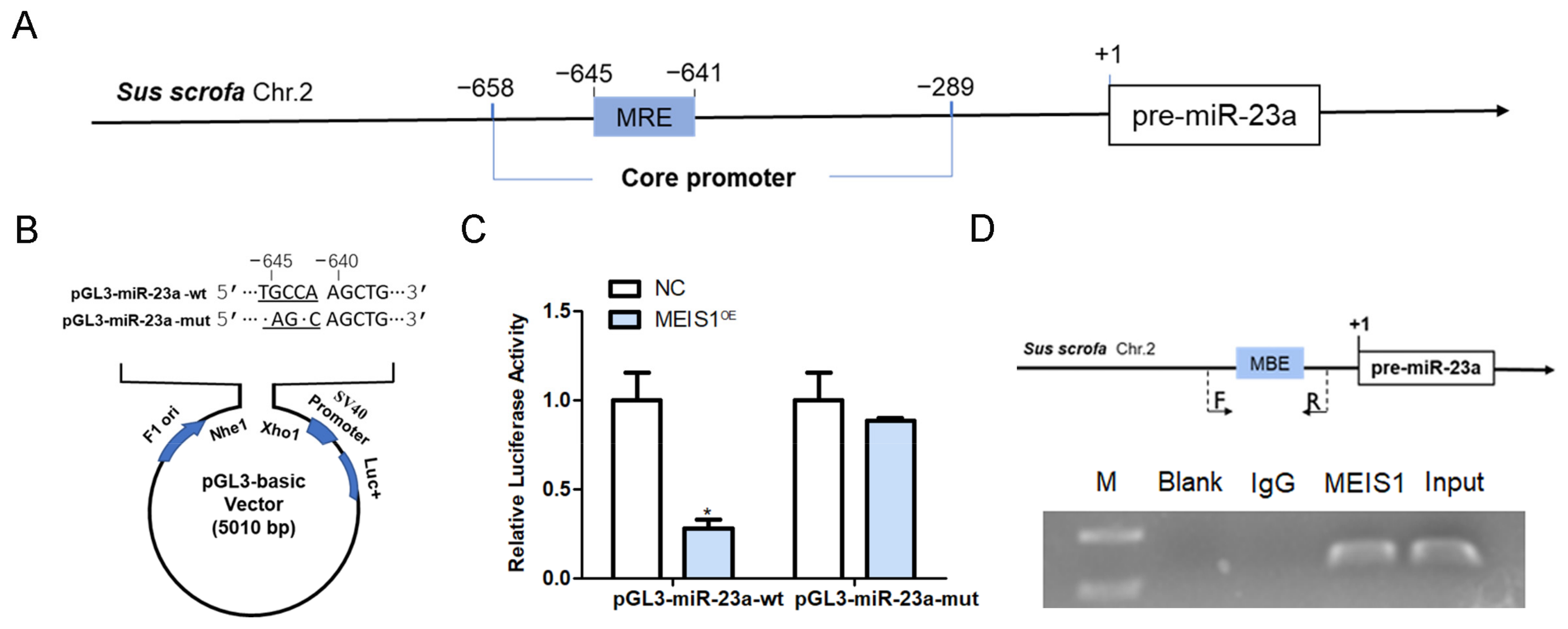
2.4. MEIS1 Inhibits miR-23a Transcription Activity by Binding to Its Core Promoter
2.5. MEIS1 Inhibits NORHA Transcription Activity by Binding to Its Core Promoter
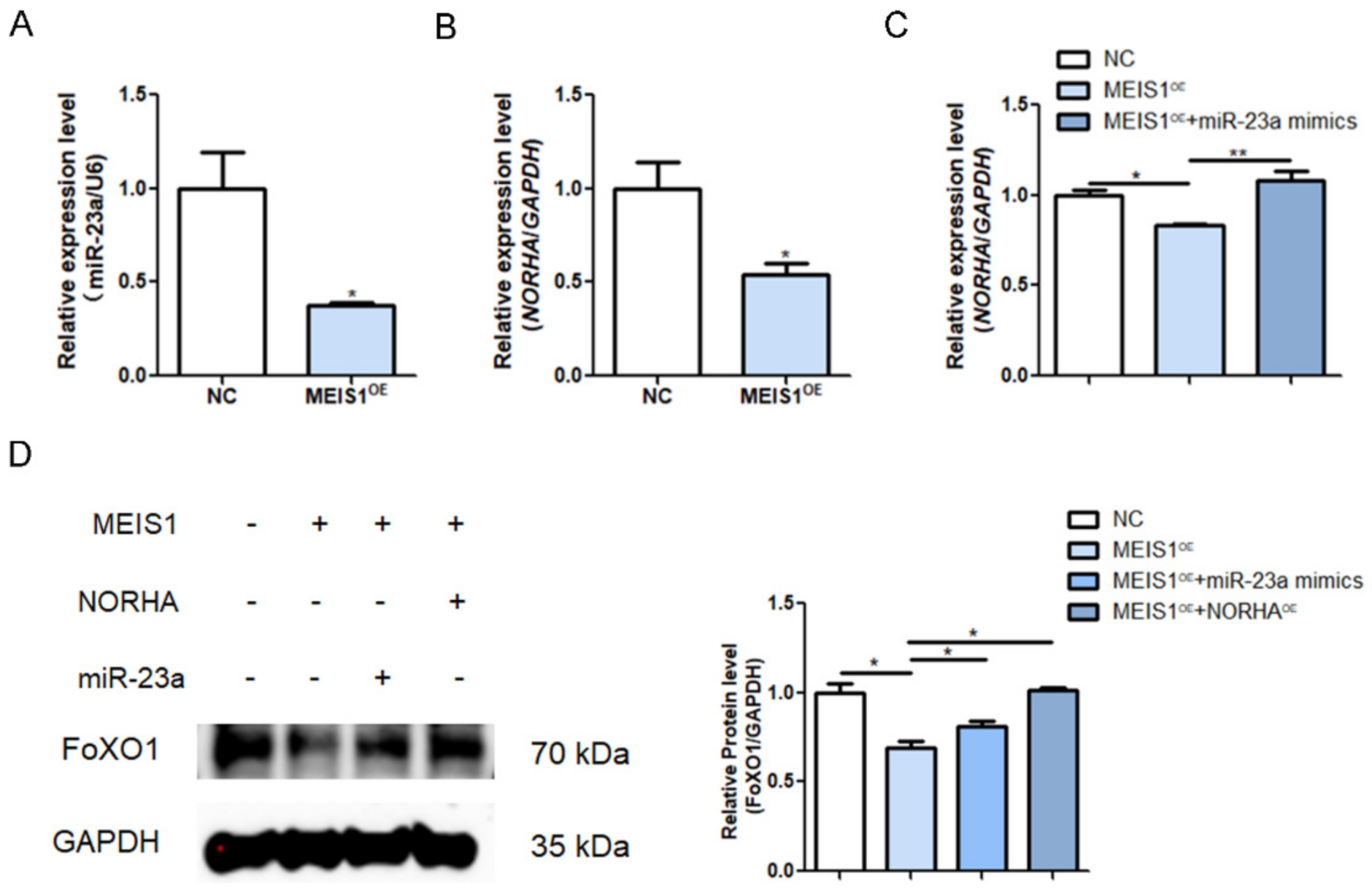
2.6. MEIS1 Commonly Transcriptional Represses the miR-23a/NORHA Axis in GCs
2.7. MEIS Inhibits GC Apoptosis through Inhibiting the miR-23a/NORHA Axis
3. Discussion
4. Materials and Methods
4.1. Bioinformatic Analysis
4.2. Immunohistochemistry (IHC)
4.3. Plasmid Construction and Oligonucleotide Synthesis
4.4. Cell Culture, Transfection, and Luciferase Reporter Assays
4.5. Chromatin Immunoprecipitation (ChIP)
4.6. RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.7. Western Blotting
4.8. Flow Cytometry
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Negrete, J.J.; Oates, A.C. Towards a physical understanding of developmental patterning. Nat. Rev. Genet. 2021, 22, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Vaddavalli, P.L.; Schumacher, B. The p53 network: Cellular and systemic DNA damage responses in cancer and aging. Trends Genet. 2022, 38, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.H.; Liu, S.Z.; Wang, G.Z.; Zhou, G.B. CXCL13 in cancer and other diseases: Biological functions, clinical significance, and therapeutic opportunities. Life 2021, 11, 1282. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Q.; Yang, L.; Li, Q.; Du, X. SMAD4 Feedback activates the canonical TGF-beta family signaling pathways. Int. J. Mol. Sci. 2021, 22, 10024. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Roy, C.S.; Mandal, G.; Purohit, S.; Gupta, A.; Bhattacharyya, A. RelA driven co-expression of CXCL13 and CXCR5 is governed by a multifaceted transcriptional program regulating breast cancer progression. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 502–511. [Google Scholar] [CrossRef]
- Du, X.; Wang, L.; Li, Q.; Wu, W.; Shang, P.; Chamba, Y.; Pan, Z.; Li, Q. miR-130a/TGF-beta1 axis is involved in sow fertility by controlling granulosa cell apoptosis. Theriogenology 2020, 157, 407–417. [Google Scholar] [CrossRef]
- Zhu, J.; Luo, Y.; Zhao, Y.; Kong, Y.; Zheng, H.; Li, Y.; Gao, B.; Ai, L.; Huang, H.; Huang, J.; et al. circEHBP1 promotes lymphangiogenesis and lymphatic metastasis of bladder cancer via miR-130a-3p/TGFbetaR1/VEGF-D signaling. Mol. Ther. 2021, 29, 1838–1852. [Google Scholar] [CrossRef]
- Ding, Y.; Hou, Y.; Liu, Y.; Yu, T.; Cui, Y.; Nie, H. MiR-130a-3p alleviates inflammatory and fibrotic phases of pulmonary fibrosis through proinflammatory factor TNF-alpha and profibrogenic receptor TGF-betaRII. Front. Pharmacol. 2022, 13, 863646. [Google Scholar] [CrossRef]
- Tian, X.; Fei, Q.; Du, M.; Zhu, H.; Ye, J.; Qian, L.; Lu, Z.; Zhang, W.; Wang, Y.; Peng, F.; et al. miR-130a-3p regulated TGF-beta1-induced epithelial-mesenchymal transition depends on SMAD4 in EC-1 cells. Cancer Med. 2019, 8, 1197–1208. [Google Scholar] [CrossRef]
- Ai, K.; Zhu, X.; Kang, Y.; Li, H.; Zhang, L. miR-130a-3p inhibition protects against renal fibrosis in vitro via the TGF-beta1/Smad pathway by targeting SnoN. Exp. Mol. Pathol. 2020, 112, 104358. [Google Scholar] [CrossRef]
- Hu, W.; Zheng, X.; Liu, J.; Zhang, M.; Liang, Y.; Song, M. MicroRNA MiR-130a-3p promotes gastric cancer by targeting Glucosaminyl N-acetyl transferase 4 (GCNT4) to regulate the TGF-beta1/SMAD3 pathway. Bioengineered 2021, 12, 11634–11647. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; Zeng, Q.; Yang, L.; Du, X.; Li, Q. A mutation in endogenous saRNA miR-23a influences granulosa cells response to oxidative stress. Antioxidants 2022, 11, 1174. [Google Scholar] [CrossRef]
- Cheng, M.; Zeng, Y.; Zhang, T.; Xu, M.; Li, Z.; Wu, Y. Transcription factor ELF1 activates MEIS1 transcription and then regulates the GFI1/FBW7 axis to promote the development of glioma. Mol. Ther. Nucleic Acids 2021, 23, 418–430. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, J.; Tang, H.; Zhai, D.; Huang, D.; Ling, L.; Wang, X.; Liu, T.; Zhang, Q.; Zhang, Z.; et al. LncRNA SNORD3A specifically sensitizes breast cancer cells to 5-FU by sponging miR-185-5p to enhance UMPS expression. Cell Death Dis. 2020, 11, 329. [Google Scholar] [CrossRef]
- Schneider, E.; Pochert, N.; Ruess, C.; MacPhee, L.; Escano, L.; Miller, C.; Krowiorz, K.; Delsing, M.E.; Heravi-Moussavi, A.; Lorzadeh, A.; et al. MicroRNA-708 is a novel regulator of the Hoxa9 program in myeloid cells. Leukemia 2020, 34, 1253–1265. [Google Scholar] [CrossRef]
- Xiang, P.; Yang, X.; Escano, L.; Dhillon, I.; Schneider, E.; Clemans-Gibbon, J.; Wei, W.; Wong, J.; Wang, S.X.; Tam, D.; et al. Elucidating the importance and regulation of key enhancers for human MEIS1 expression. Leukemia 2022, 36, 1980–1989. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, X.; Mei, S. Autophagy in ovarian follicular development and atresia. Int. J. Biol. Sci. 2019, 15, 726–737. [Google Scholar] [CrossRef]
- Meng, L.; Zhao, K.; Wang, C.C.; Tao, J.; Wu, Z.; Teerds, K.; Zhang, S. characterization of long non-coding RNA profiles in porcine granulosa cells of healthy and atretic antral follicles: Implications for a potential role in apoptosis. Int. J. Mol. Sci. 2021, 22, 2677. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Zhang, J.; Du, X.; Li, Q.; Pan, Z. circSLC41A1 resists porcine granulosa cell apoptosis and follicular atresia by promoting srsf1 through miR-9820-5p sponging. Int. J. Mol. Sci. 2022, 23, 1509. [Google Scholar] [CrossRef]
- Du, X.; Li, Q.; Yang, L.; Liu, L.; Cao, Q.; Li, Q. SMAD4 activates Wnt signaling pathway to inhibit granulosa cell apoptosis. Cell Death Dis. 2020, 11, 373. [Google Scholar] [CrossRef]
- Yang, L.; Wang, S.; Pan, Z.; Du, X.; Li, Q. TGFBR2 is a novel substrate and indirect transcription target of deubiquitylase USP9X in granulosa cells. J. Cell. Physiol. 2022, 237, 2969–2979. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Pan, Z.; Du, X.; Zhang, J.; Liu, H.; Li, Q. NORHA, a novel follicular atresia-related lncRNA, promotes porcine granulosa cell apoptosis via the miR-183-96-182 cluster and FoxO1 axis. J. Anim. Sci. Biotechnol. 2021, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.; Yao, W.; Li, Q.; Liu, H.; Pan, Z. Initiation of follicular atresia: Gene networks during early atresia in pig ovaries. Reproduction 2018, 156, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Pan, Z.; Li, Q.; Liu, H.; Li, Q. SMAD4 feedback regulates the canonical TGF-beta signaling pathway to control granulosa cell apoptosis. Cell Death Dis. 2018, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Du, X.; Zhang, J.; Wang, Y.; Wang, M.; Pan, Z.; Li, Q. SMAD4-induced knockdown of the antisense long noncoding RNA BRE-AS contributes to granulosa cell apoptosis. Mol. Ther. Nucleic Acids 2021, 25, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, H.; Huang, C.L.; Chen, H.; Zhu, G.; Qian, K. Regulation of myeloid ecotropic viral integration site 1 and its expression in normal and abnormal endometrium. Fertil. Steril. 2014, 102, 856–863. [Google Scholar] [CrossRef]
- Karapetsas, A.; Tokamani, M.; Evangelou, C.; Sandaltzopoulos, R. The homeodomain transcription factor MEIS1 triggers chemokine expression and is involved in CD8+ T-lymphocyte infiltration in early stage ovarian cancer. Mol. Carcinog. 2018, 57, 1251–1263. [Google Scholar] [CrossRef]
- Ota, T.; Asahina, H.; Park, S.H.; Huang, Q.; Minegishi, T.; Auersperg, N.; Leung, P.C. HOX cofactors expression and regulation in the human ovary. Reprod. Biol. Endocrinol. 2008, 6, 49. [Google Scholar] [CrossRef]
- Huang, S.; Li, X.; Zheng, H.; Si, X.; Li, B.; Wei, G.; Li, C.; Chen, Y.; Chen, Y.; Liao, W.; et al. Loss of super-enhancer-regulated circRNA NFIX induces cardiac regeneration after myocardial infarction in adult mice. Circulation 2019, 139, 2857–2876. [Google Scholar] [CrossRef]
- Du, X.; Liu, L.; Li, Q.; Zhang, L.; Pan, Z.; Li, Q. NORFA, long intergenic noncoding RNA, maintains sow fertility by inhibiting granulosa cell death. Commun. Biol. 2020, 3, 131. [Google Scholar] [CrossRef] [Green Version]
- Piergentili, R.; Basile, G.; Nocella, C.; Carnevale, R.; Marinelli, E.; Patrone, R.; Zaami, S. Using ncRNAs as tools in cancer diagnosis and treatment-the way towards personalized medicine to improve patients’ health. Int. J. Mol. Sci. 2022, 23, 9353. [Google Scholar] [CrossRef]
- Cavaliere, A.F.; Perelli, F.; Zaami, S.; D’Indinosante, M.; Turrini, I.; Giusti, M.; Gullo, G.; Vizzielli, G.; Mattei, A.; Scambia, G.; et al. Fertility sparing treatments in endometrial cancer patients: The potential role of the new molecular classification. Int. J. Mol. Sci. 2021, 22, 12248. [Google Scholar] [CrossRef]
- Schneider, J.F.; Nonneman, D.J.; Wiedmann, R.T.; Vallet, J.L.; Rohrer, G.A. Genomewide association and identification of candidate genes for ovulation rate in swine. J. Anim. Sci. 2014, 92, 3792–3803. [Google Scholar] [CrossRef]
- Bidanel, J.P.; Rosendo, A.; Iannuccelli, N.; Riquet, J.; Gilbert, H.; Caritez, J.C.; Billon, Y.; Amigues, Y.; Prunier, A.; Milan, D. Detection of quantitative trait loci for teat number and female reproductive traits in Meishan x Large White F2 pigs. Animal 2008, 2, 813–820. [Google Scholar] [CrossRef]
- Deng, Y.H.; Deng, Z.H.; Hao, H.; Wu, X.L.; Gao, H.; Tang, S.H.; Tang, H. MicroRNA-23a promotes colorectal cancer cell survival by targeting PDK4. Exp. Cell Res. 2018, 373, 171–179. [Google Scholar] [CrossRef]
- Li, Y.; Gan, Y.; Liu, J.; Li, J.; Zhou, Z.; Tian, R.; Sun, R.; Liu, J.; Xiao, Q.; Li, Y.; et al. Downregulation of MEIS1 mediated by ELFN1-AS1/EZH2/DNMT3a axis promotes tumorigenesis and oxaliplatin resistance in colorectal cancer. Signal Transduct. Target Ther. 2022, 7, 87. [Google Scholar] [CrossRef]
- Choi, J.H.; Jang, T.Y.; Jeon, S.E.; Kim, J.H.; Lee, C.J.; Yun, H.J.; Jung, J.Y.; Park, S.Y.; Nam, J.S. The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and metastasis by suppressing meis1 expression. Int. J. Mol. Sci. 2021, 22, 13413. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, S.J.; Choo, J.; Heo, G.; Yoo, J.W.; Jung, Y.; Rhee, S.H.; Im, E. miR-23a-3p is a key regulator of IL-17c-induced tumor angiogenesis in colorectal cancer. Cells 2020, 9, 1363. [Google Scholar] [CrossRef]
- Yachi, K.; Tsuda, M.; Kohsaka, S.; Wang, L.; Oda, Y.; Tanikawa, S.; Ohba, Y.; Tanaka, S. miR-23a promotes invasion of glioblastoma via HOXD10-regulated glial-mesenchymal transition. Signal Transduct. Target Ther. 2018, 3, 33. [Google Scholar] [CrossRef]
- Xiao, L.; Li, X.; Mu, Z.; Zhou, J.; Zhou, P.; Xie, C.; Jiang, S. FTO inhibition enhances the antitumor effect of temozolomide by targeting MYC-miR-155/23a cluster-MXI1 feedback circuit in Glioma. Cancer Res. 2020, 80, 3945–3958. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Hung, J.Y.; Chang, W.A.; Lin, Y.S.; Pan, Y.C.; Tsai, P.H.; Wu, C.Y.; Kuo, P.L. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Bandara, G.; Cho, Y.E.; Komarow, H.D.; Donahue, D.R.; Karim, B.; Baek, M.C.; Kim, H.M.; Metcalfe, D.D.; Olivera, A. Mastocytosis-derived extracellular vesicles deliver miR-23a and miR-30a into pre-osteoblasts and prevent osteoblastogenesis and bone formation. Nat. Commun. 2021, 12, 2527. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, C.; Mostile, G.; Baglieri, G.; Giunta, F.; Luca, A.; Raciti, L.; Zappia, M.; Purrello, M.; Ragusa, M.; Nicoletti, A. Specific signatures of serum miRNAs as potential biomarkers to discriminate clinically similar neurodegenerative and Vascular-Related diseases. Cell. Mol. Neurobiol. 2020, 40, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Salemi, M.; Marchese, G.; Lanza, G.; Cosentino, F.; Salluzzo, M.G.; Schillaci, F.A.; Ventola, G.M.; Cordella, A.; Ravo, M.; Ferri, R. Role and dysregulation of miRNA in patients with parkinson’s disease. Int. J. Mol. Sci. 2022, 24, 712. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wang, Y.; Chen, Y.; Li, Y.; Du, X.; Li, Y.; Li, Q. MEIS1 Is a Common Transcription Repressor of the miR-23a and NORHA Axis in Granulosa Cells. Int. J. Mol. Sci. 2023, 24, 3589. https://doi.org/10.3390/ijms24043589
Wang S, Wang Y, Chen Y, Li Y, Du X, Li Y, Li Q. MEIS1 Is a Common Transcription Repressor of the miR-23a and NORHA Axis in Granulosa Cells. International Journal of Molecular Sciences. 2023; 24(4):3589. https://doi.org/10.3390/ijms24043589
Chicago/Turabian StyleWang, Siqi, Yang Wang, Yibo Chen, Yuqi Li, Xing Du, Yinxia Li, and Qifa Li. 2023. "MEIS1 Is a Common Transcription Repressor of the miR-23a and NORHA Axis in Granulosa Cells" International Journal of Molecular Sciences 24, no. 4: 3589. https://doi.org/10.3390/ijms24043589
APA StyleWang, S., Wang, Y., Chen, Y., Li, Y., Du, X., Li, Y., & Li, Q. (2023). MEIS1 Is a Common Transcription Repressor of the miR-23a and NORHA Axis in Granulosa Cells. International Journal of Molecular Sciences, 24(4), 3589. https://doi.org/10.3390/ijms24043589