


Carcinoid Syndrome: Preclinical Models and Future Therapeutic Strategies

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
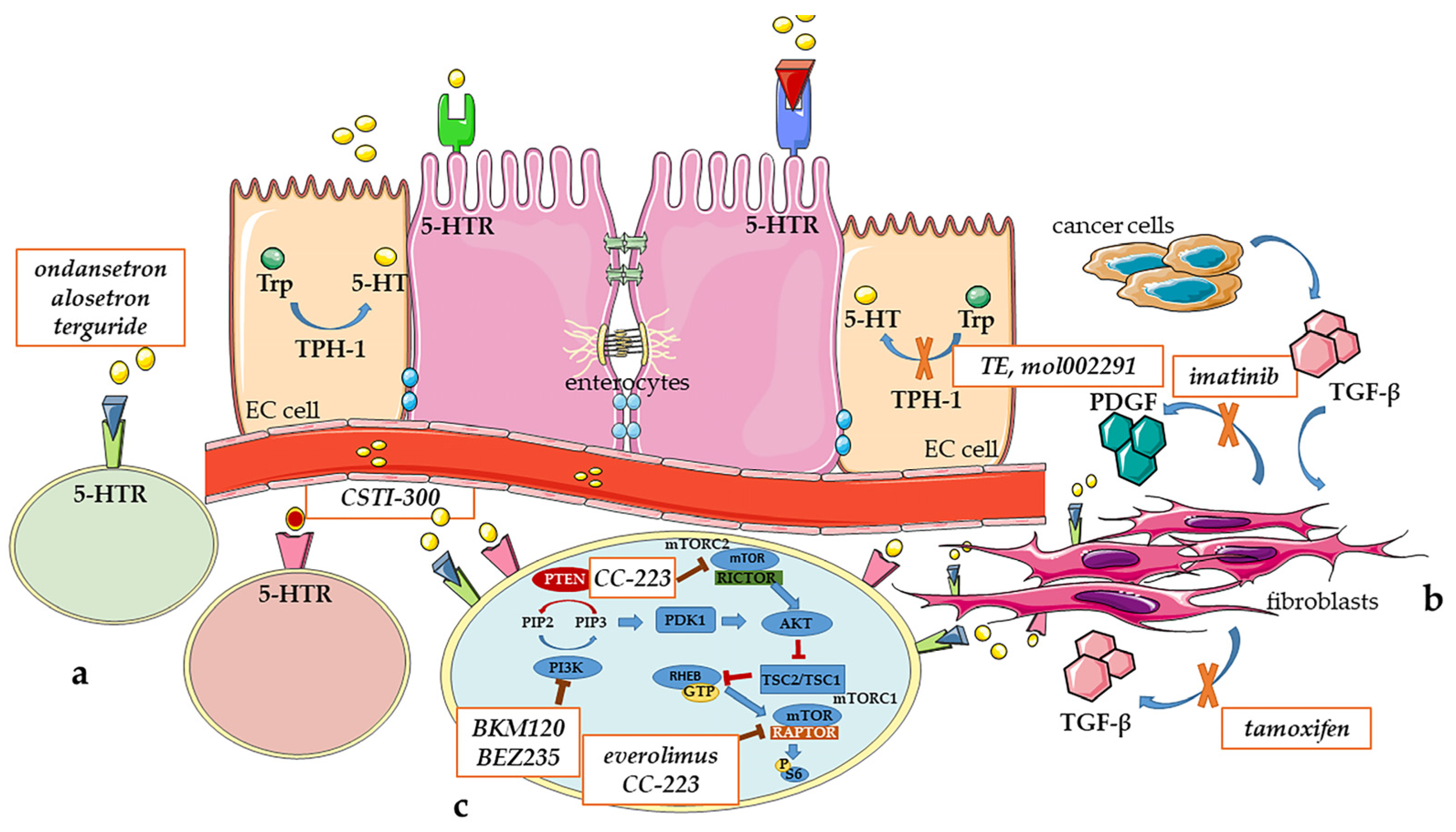
2. Preclinical Models of Carcinoid Syndrome
2.1. In Vitro Models
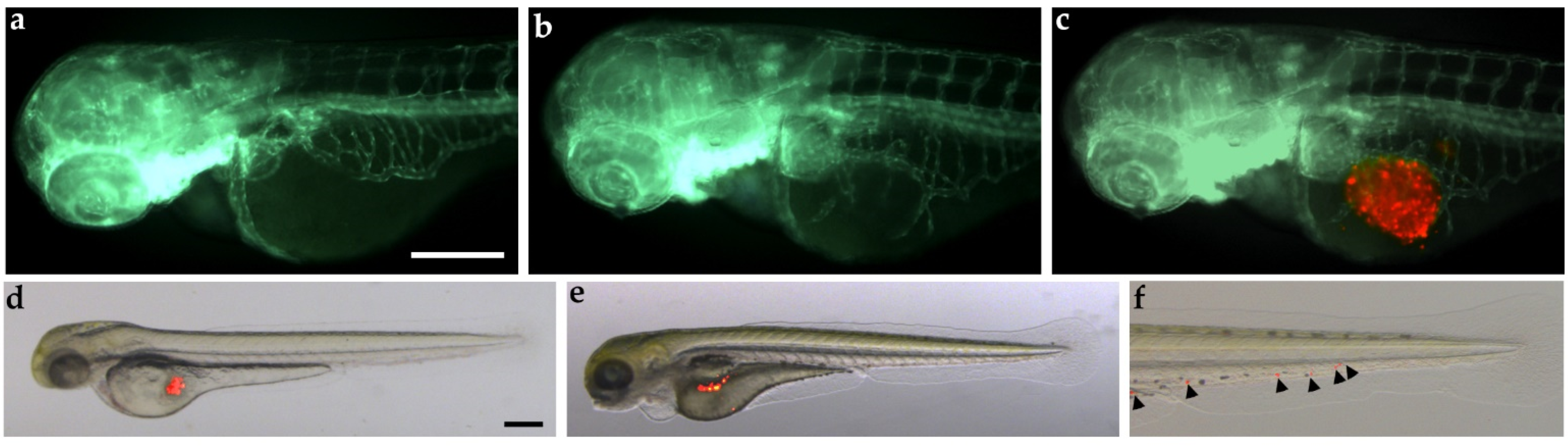
2.2. In Vivo Models
3. Future Therapies for Carcinoid Syndrome
4. Future Development of Preclinical Models in Carcinoid Syndrome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dasari, A.; Shen, C.; Halperin, D.; Zhao, B.; Zhou, S.; Xu, Y.; Shih, T.; Yao, J.C. Trends in the Incidence, Prevalence, and Survival Outcomes in Patients with Neuroendocrine Tumors in the United States. JAMA Oncol. 2017, 3, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Grozinsky-Glasberg, S.; Grossman, A.B.; Gross, D.J. Carcinoid Heart Disease: From Pathophysiology to Treatment–‘Something in the Way It Moves’. Neuroendocrinology 2015, 101, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Kimbrough, C.W.; Beal, E.W.; Dillhoff, M.E.; Schmidt, C.R.; Pawlik, T.M.; Lopez-Aguiar, A.G.; Poultsides, G.; Makris, E.; Rocha, F.G.; Crown, A.; et al. Influence of carcinoid syndrome on the clinical characteristics and outcomes of patients with gastroenteropancreatic neuroendocrine tumors undergoing operative resection. Surgery 2019, 165, 657–663. [Google Scholar] [CrossRef]
- Mota, J.M.; Sousa, L.G.; Riechelmann, R.P. Complications from carcinoid syndrome: Review of the current evidence. Ecancermedicalscience 2016, 10, 662. [Google Scholar] [CrossRef] [PubMed]
- Tamagno, G.; Bennett, A.; Ivanovski, I. Lights and darks of neuroendocrine tumors of the appendix. Minerva Endocrinol. 2020, 45, 381–392. [Google Scholar] [CrossRef]
- Halperin, D.M.; Shen, C.; Dasari, A.; Xu, Y.; Chu, Y.; Zhou, S.; Shih, Y.T.; Yao, J.C. Frequency of carcinoid syndrome at neuroendocrine tumour diagnosis: A population-based study. Lancet Oncol. 2017, 18, 525–534. [Google Scholar] [CrossRef]
- Connolly, H.M.; Schaff, H.V.; Mullany, C.J.; Rubin, J.; Abel, M.D.; Pellikka, P.A. Surgical management of left-sided carcinoid heart disease. Circulation 2001, 104, I36–I40. [Google Scholar] [CrossRef]
- Moller, J.E.; Pellikka, P.A.; Bernheim, A.M.; Schaff, H.V.; Rubin, J.; Connolly, H.M. Prognosis of carcinoid heart disease: Analysis of 200 cases over two decades. Circulation 2005, 112, 3320–3327. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Davar, J.; Dreyfus, G.; Caplin, M.E. Carcinoid heart disease. Circulation 2007, 116, 2860–2865. [Google Scholar] [CrossRef]
- Macfie, R.; McCully, B.H.; Ratzlaff, A.N.; Lim, J.; Song, H.K.; Dewey, E.N.; Pommier, S.J.; Bhamidipati, C.M.; Pommier, R.F. The prevalence, operations, and outcomes of carcinoid heart disease. Am. J. Surg. 2022, 224, 665–669. [Google Scholar] [CrossRef]
- Clement, D.; Ramage, J.; Srirajaskanthan, R. Update on Pathophysiology, Treatment, and Complications of Carcinoid Syndrome. J. Oncol. 2020, 2020, 8341426. [Google Scholar] [CrossRef] [PubMed]
- Fanciulli, G.; Ruggeri, R.M.; Grossrubatscher, E.; Calzo, F.L.; Wood, T.D.; Faggiano, A.; Isidori, A.; Colao, A. Nike Serotonin pathway in carcinoid syndrome: Clinical, diagnostic, prognostic and therapeutic implications. Rev. Endocr. Metab. Disord. 2020, 21, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, G.; Mosca, A.; Frara, S.; Vitale, G.; Giustina, A. Somatostatin analogs in the treatment of neuroendocrine tumors: Current and emerging aspects. Expert. Opin. Pharmacother. 2017, 18, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Koumarianou, A.; Daskalakis, K.; Tsoli, M.; Kaltsas, G.; Pavel, M. Efficacy, safety and unmet needs of evolving medical treatments for carcinoid syndrome. J. Neuroendocrinol. 2022, 34, e13174. [Google Scholar] [CrossRef] [PubMed]
- Lamarca, A.; Barriuso, J.; McNamara, M.G.; Hubner, R.A.; Valle, J.W. Telotristat ethyl: A new option for the management of carcinoid syndrome. Expert. Opin. Pharmacother. 2016, 17, 2487–2498. [Google Scholar] [CrossRef] [PubMed]
- Dicitore, A.; Cantone, M.C. Targeting receptor tyrosine kinases in neuroendocrine neoplasm: What’s going on with lung carcinoids? Minerva Endocrinol. 2022, 47, 261–263. [Google Scholar] [CrossRef]
- Modlin, I.M.; Kidd, M.; Latich, I.; Zikusoka, M.N.; Shapiro, M.D. Current status of gastrointestinal carcinoids. Gastroenterology 2005, 128, 1717–1751. [Google Scholar] [CrossRef]
- Ear, P.H.; Li, G.; Wu, M.; Abusada, E.; Bellizzi, A.M.; Howe, J.R. Establishment and Characterization of Small Bowel Neuroendocrine Tumor Spheroids. J. Vis. Exp. 2019, 152. [Google Scholar] [CrossRef]
- Debons-Guillemin, M.C.; Launay, J.M.; Roseto, A.; Peries, J. Serotonin and histamine production by human carcinoid cells in culture. Cancer Res. 1982, 42, 1513–1516. [Google Scholar]
- Modlin, I.M.; Kidd, M.; Pfragner, R.; Eick, G.N.; Champaneria, M.C. The functional characterization of normal and neoplastic human enterochromaffin cells. J. Clin. Endocrinol. Metab. 2006, 91, 2340–2348. [Google Scholar] [CrossRef]
- Pfragner, R.; Wirnsberger, G.; Niederle, B.; Behmel, A.; Rinner, I.; Mandl, A.; Wawrina, F.; Luo, J.; Adamiker, D.; Hoger, H.; et al. Establishment of a continuous cell line from a human carcinoid of the small intestine (KRJ-I). Int. J. Oncol. 1996, 8, 513–520. [Google Scholar] [CrossRef]
- Siddique, Z.L.; Drozdov, I.; Floch, J.; Gustafsson, B.I.; Stunes, K.; Pfragner, R.; Kidd, M.; Modlin, I.M. KRJ-I and BON cell lines: Defining an appropriate enterochromaffin cell neuroendocrine tumor model. Neuroendocrinology 2009, 89, 458–470. [Google Scholar] [CrossRef]
- Kidd, M.; Schally, A.V.; Pfragner, R.; Malfertheiner, M.V.; Modlin, I.M. Inhibition of proliferation of small intestinal and bronchopulmonary neuroendocrine cell lines by using peptide analogs targeting receptors. Cancer 2008, 112, 1404–1414. [Google Scholar] [CrossRef]
- Svejda, B.; Kidd, M.; Kazberouk, A.; Lawrence, B.; Pfragner, R.; Modlin, I.M. Limitations in small intestinal neuroendocrine tumor therapy by mTor kinase inhibition reflect growth factor-mediated PI3K feedback loop activation via ERK1/2 and AKT. Cancer 2011, 117, 4141–4154. [Google Scholar] [CrossRef] [PubMed]
- Bold, R.J.; Ishizuka, J.; Townsend, C.M., Jr.; Thompson, J.C. All-trans-retinoic acid inhibits growth of human pancreatic cancer cell lines. Pancreas 1996, 12, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.R.; Pinchot, S.N.; Jaskula-Sztul, R.; Luo, J.; Kunnimalaiyaan, M.; Chen, H. Identification of a novel Raf-1 pathway activator that inhibits gastrointestinal carcinoid cell growth. Mol. Cancer Ther. 2010, 9, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Evers, B.M.; Ishizuka, J.; Townsend, C.M., Jr.; Thompson, J.C. The human carcinoid cell line, BON. A model system for the study of carcinoid tumors. Ann. N. Y. Acad. Sci. 1994, 733, 393–406. [Google Scholar] [CrossRef]
- Greenblatt, D.Y.; Ndiaye, M.; Chen, H.; Kunnimalaiyaan, M. Lithium inhibits carcinoid cell growth in vitro. Am. J. Transl. Res. 2010, 2, 248–253. [Google Scholar] [PubMed]
- Lopez, J.R.; Claessen, S.M.; Macville, M.V.; Albrechts, J.C.; Skogseid, B.; Speel, E.J. Spectral karyotypic and comparative genomic analysis of the endocrine pancreatic tumor cell line BON-1. Neuroendocrinology 2010, 91, 131–141. [Google Scholar] [CrossRef]
- Parekh, D.; Ishizuka, J.; Townsend, C.M., Jr.; Haber, B.E.; Beauchamp, R.D.; Rajaraman, S.; Karp, G.; Hsieh, J.; Thompson, J.C. Differential effects of sodium butyrate and hexamethylene bisacetamide on growth and secretion of cultured human endocrine tumor cells. Arch. Surg. 1991, 126, 467–472. [Google Scholar] [CrossRef]
- Pitt, S.C.; Chen, H.; Kunnimalaiyaan, M. Inhibition of phosphatidylinositol 3-kinase/Akt signaling suppresses tumor cell proliferation and neuroendocrine marker expression in GI carcinoid tumors. Ann. Surg. Oncol. 2009, 16, 2936–2942. [Google Scholar] [CrossRef]
- Somnay, Y.; Simon, K.; Harrison, A.D.; Kunnimalaiyaan, S.; Chen, H.; Kunnimalaiyaan, M. Neuroendocrine phenotype alteration and growth suppression through apoptosis by MK-2206, an allosteric inhibitor of AKT, in carcinoid cell lines in vitro. Anticancer Drugs 2013, 24, 66–72. [Google Scholar] [CrossRef]
- Veenstra, M.J.; van Koetsveld, P.M.; Dogan, F.; Farrell, W.E.; Feelders, R.A.; Lamberts, S.W.J.; de Herder, W.W.; Vitale, G.; Hofland, L.J. Epidrug-induced upregulation of functional somatostatin type 2 receptors in human pancreatic neuroendocrine tumor cells. Oncotarget 2018, 9, 14791–14802. [Google Scholar] [CrossRef]
- Yao, J.C.; Zhang, J.X.; Rashid, A.; Yeung, S.C.; Szklaruk, J.; Hess, K.; Xie, K.; Ellis, L.; Abbruzzese, J.L.; Ajani, J.A. Clinical and in vitro studies of imatinib in advanced carcinoid tumors. Clin. Cancer Res. 2007, 13, 234–240. [Google Scholar] [CrossRef]
- Zitzmann, K.; De Toni, E.N.; Brand, S.; Goke, B.; Meinecke, J.; Spottl, G.; Meyer, H.H.; Auernhammer, C.J. The novel mTOR inhibitor RAD001 (everolimus) induces antiproliferative effects in human pancreatic neuroendocrine tumor cells. Neuroendocrinology 2007, 85, 54–60. [Google Scholar] [CrossRef]
- Nolting, S.; Rentsch, J.; Freitag, H.; Detjen, K.; Briest, F.; Mobs, M.; Weissmann, V.; Siegmund, B.; Auernhammer, C.J.; Aristizabal Prada, E.T.; et al. The selective PI3Kalpha inhibitor BYL719 as a novel therapeutic option for neuroendocrine tumors: Results from multiple cell line models. PLoS ONE 2017, 12, e0182852. [Google Scholar] [CrossRef]
- Terracciano, F.; Capone, A.; Montori, A.; Rinzivillo, M.; Partelli, S.; Panzuto, F.; Pilozzi, E.; Arcidiacono, P.G.; Sette, C.; Capurso, G. MYC Upregulation Confers Resistance to Everolimus and Establishes Vulnerability to Cyclin-Dependent Kinase Inhibitors in Pancreatic Neuroendocrine Neoplasm Cells. Neuroendocrinology 2021, 111, 739–751. [Google Scholar] [CrossRef]
- Vitale, G.; de Herder, W.W.; van Koetsveld, P.M.; Waaijers, M.; Schoordijk, W.; Croze, E.; Colao, A.; Lamberts, S.W.; Hofland, L.J. IFN-beta is a highly potent inhibitor of gastroenteropancreatic neuroendocrine tumor cell growth in vitro. Cancer Res. 2006, 66, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; van Koetsveld, P.M.; de Herder, W.W.; van der Wansem, K.; Janssen, J.A.; Colao, A.; Lombardi, G.; Lamberts, S.W.; Hofland, L.J. Effects of type I interferons on IGF-mediated autocrine/paracrine growth of human neuroendocrine tumor cells. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E559–E566. [Google Scholar] [CrossRef] [PubMed]
- Cuny, T.; van Koetsveld, P.M.; Mondielli, G.; Dogan, F.; de Herder, W.W.; Barlier, A.; Hofland, L.J. Reciprocal Interactions between Fibroblast and Pancreatic Neuroendocrine Tumor Cells: Putative Impact of the Tumor Microenvironment. Cancers 2022, 14, 3481. [Google Scholar] [CrossRef] [PubMed]
- Doihara, H.; Nozawa, K.; Kojima, R.; Kawabata-Shoda, E.; Yokoyama, T.; Ito, H. QGP-1 cells release 5-HT via TRPA1 activation; a model of human enterochromaffin cells. Mol. Cell. Biochem. 2009, 331, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Dossing, K.B.V.; Kjaer, C.; Vikesa, J.; Binderup, T.; Knigge, U.; Culler, M.D.; Kjaer, A.; Federspiel, B.; Friis-Hansen, L. Somatostatin Analogue Treatment Primarily Induce miRNA Expression Changes and Up-Regulates Growth Inhibitory miR-7 and miR-148a in Neuroendocrine Cells. Genes 2018, 9, 337. [Google Scholar] [CrossRef]
- Brandt, D.W.; Pandol, S.J.; Deftos, L.J. Calcium-stimulated parathyroid hormone-like protein secretion: Potentiation through a protein kinase-C pathway. Endocrinology 1991, 128, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Pitt, S.C.; Chen, H.; Kunnimalaiyaan, M. Phosphatidylinositol 3-kinase-Akt signaling in pulmonary carcinoid cells. J. Am. Coll. Surg. 2009, 209, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Kidd, M.; Drozdov, I.; Joseph, R.; Pfragner, R.; Culler, M.; Modlin, I. Differential cytotoxicity of novel somatostatin and dopamine chimeric compounds on bronchopulmonary and small intestinal neuroendocrine tumor cell lines. Cancer 2008, 113, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Larsson, D.E.; Wickstrom, M.; Hassan, S.; Oberg, K.; Granberg, D. The cytotoxic agents NSC-95397, brefeldin A, bortezomib and sanguinarine induce apoptosis in neuroendocrine tumors in vitro. Anticancer Res. 2010, 30, 149–156. [Google Scholar]
- Hubbell, H.R.; Quinn, L.A.; Dolby, T.W. Cloning of a non-c-myc DNA fragment from the double minutes of a human colon carcinoid cell line. Cancer Genet. Cytogenet. 1987, 24, 17–31. [Google Scholar] [CrossRef]
- Lin, Y.L.; Liau, J.Y.; Yu, S.C.; Ou, D.L.; Lin, L.I.; Tseng, L.H.; Chang, Y.L.; Yeh, K.H.; Cheng, A.L. KRAS mutation is a predictor of oxaliplatin sensitivity in colon cancer cells. PLoS ONE 2012, 7, e50701. [Google Scholar] [CrossRef]
- Kolby, L.; Bernhardt, P.; Ahlman, H.; Wangberg, B.; Johanson, V.; Wigander, A.; Forssell-Aronsson, E.; Karlsson, S.; Ahren, B.; Stenman, G.; et al. A transplantable human carcinoid as model for somatostatin receptor-mediated and amine transporter-mediated radionuclide uptake. Am. J. Pathol. 2001, 158, 745–755. [Google Scholar] [CrossRef]
- Kirkland, S.C.; Ellison, M.L. Secretion of corticotrophin releasing factor-like activity by a human bronchial carcinoid cell line. J. Endocrinol. 1984, 103, 85–90. [Google Scholar] [CrossRef]
- Herrera-Martinez, A.D.; van den Dungen, R.; Dogan-Oruc, F.; van Koetsveld, P.M.; Culler, M.D.; de Herder, W.W.; Luque, R.M.; Feelders, R.A.; Hofland, L.J. Effects of novel somatostatin-dopamine chimeric drugs in 2D and 3D cell culture models of neuroendocrine tumors. Endocr. Relat. Cancer 2019, 26, 585–599. [Google Scholar] [CrossRef]
- Ellis, L.M.; Samuel, S.; Sceusi, E. Varying opinions on the authenticity of a human midgut carcinoid cell line--letter. Clin. Cancer Res. 2010, 16, 5365–5366. [Google Scholar] [CrossRef]
- Bernhardt, P.; Oddstig, J.; Kolby, L.; Nilsson, O.; Ahlman, H.; Forssell-Aronsson, E. Effects of treatment with (177)Lu-DOTA-Tyr(3)-octreotate on uptake of subsequent injection in carcinoid-bearing nude mice. Cancer Biother. Radiopharm. 2007, 22, 644–653. [Google Scholar] [CrossRef]
- Musunuru, S.; Carpenter, J.E.; Sippel, R.S.; Kunnimalaiyaan, M.; Chen, H. A mouse model of carcinoid syndrome and heart disease. J. Surg. Res. 2005, 126, 102–105. [Google Scholar] [CrossRef]
- Jackson, L.N.; Chen, L.A.; Larson, S.D.; Silva, S.R.; Rychahou, P.G.; Boor, P.J.; Li, J.; Defreitas, G.; Stafford, W.L.; Townsend, C.M., Jr.; et al. Development and characterization of a novel in vivo model of carcinoid syndrome. Clin. Cancer Res. 2009, 15, 2747–2755. [Google Scholar] [CrossRef]
- Gustafsson, B.I.; Tommeras, K.; Nordrum, I.; Loennechen, J.P.; Brunsvik, A.; Solligard, E.; Fossmark, R.; Bakke, I.; Syversen, U.; Waldum, H. Long-term serotonin administration induces heart valve disease in rats. Circulation 2005, 111, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Elangbam, C.S.; Job, L.E.; Zadrozny, L.M.; Barton, J.C.; Yoon, L.W.; Gates, L.D.; Slocum, N. 5-hydroxytryptamine (5HT)-induced valvulopathy: Compositional valvular alterations are associated with 5HT2B receptor and 5HT transporter transcript changes in Sprague-Dawley rats. Exp. Toxicol. Pathol. 2008, 60, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Lancellotti, P.; Nchimi, A.; Hego, A.; Dulgheru, R.; Delvenne, P.; Drion, P.; Oury, C. High-dose oral intake of serotonin induces valvular heart disease in rabbits. Int. J. Cardiol. 2015, 197, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Nebigil, C.G.; Jaffre, F.; Messaddeq, N.; Hickel, P.; Monassier, L.; Launay, J.M.; Maroteaux, L. Overexpression of the serotonin 5-HT2B receptor in heart leads to abnormal mitochondrial function and cardiac hypertrophy. Circulation 2003, 107, 3223–3229. [Google Scholar] [CrossRef] [PubMed]
- Mekontso-Dessap, A.; Brouri, F.; Pascal, O.; Lechat, P.; Hanoun, N.; Lanfumey, L.; Seif, I.; Benhaiem-Sigaux, N.; Kirsch, M.; Hamon, M.; et al. Deficiency of the 5-hydroxytryptamine transporter gene leads to cardiac fibrosis and valvulopathy in mice. Circulation 2006, 113, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Gronstad, K.O.; Nilsson, O.; Dahlstrom, A.; Skolnik, G.; Ahlman, H. Adrenergic control of serotonin release from carcinoid tumor cells in vitro and in vivo. J. Surg. Res. 1987, 42, 141–146. [Google Scholar] [CrossRef]
- Welford, R.W.; Vercauteren, M.; Trebaul, A.; Cattaneo, C.; Eckert, D.; Garzotti, M.; Sieber, P.; Segrestaa, J.; Studer, R.; Groenen, P.M.; et al. Serotonin biosynthesis as a predictive marker of serotonin pharmacodynamics and disease-induced dysregulation. Sci. Rep. 2016, 6, 30059. [Google Scholar] [CrossRef] [PubMed]
- Hauso, O.; Gustafsson, B.I.; Loennechen, J.P.; Stunes, A.K.; Nordrum, I.; Waldum, H.L. Long-term serotonin effects in the rat are prevented by terguride. Regul. Pept. 2007, 143, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Contractor, T.; Clausen, R.; Harris, G.R.; Rosenfeld, J.A.; Carpizo, D.R.; Tang, L.; Harris, C.R. IGF2 drives formation of ileal neuroendocrine tumors in patients and mice. Endocr. Relat. Cancer 2020, 27, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Stevanovic, K.; Li, Z.; Yang, Q.M.; Oravecz, T.; Zambrowicz, B.; Jhaver, K.G.; Diacou, A.; Gershon, M.D. Pharmacological reduction of mucosal but not neuronal serotonin opposes inflammation in mouse intestine. Gut 2014, 63, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Wang, H.; Terc, J.D.; Zambrowicz, B.; Yang, Q.M.; Khan, W.I. Blocking peripheral serotonin synthesis by telotristat etiprate (LX1032/LX1606) reduces severity of both chemical- and infection-induced intestinal inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G455–G465. [Google Scholar] [CrossRef] [PubMed]
- Grozinsky-Glasberg, S.; Shimon, I.; Rubinfeld, H. The role of cell lines in the study of neuroendocrine tumors. Neuroendocrinology 2012, 96, 173–187. [Google Scholar] [CrossRef]
- Ferrans, V.J.; Roberts, W.C. The carcinoid endocardial plaque; an ultrastructural study. Hum. Pathol. 1976, 7, 387–409. [Google Scholar] [CrossRef]
- Fu, J.; Qiu, F.; Stolniceanu, C.R.; Yu, F.; Zang, S.; Xiang, Y.; Huang, Y.; Matovic, M.; Stefanescu, C.; Tang, Q.; et al. Combined use of (177) Lu-DOTATATE peptide receptor radionuclide therapy and fluzoparib for treatment of well-differentiated neuroendocrine tumors: A preclinical study. J. Neuroendocrinol. 2022, 34, e13109. [Google Scholar] [CrossRef]
- Johnbeck, C.B.; Munk Jensen, M.; Haagen Nielsen, C.; Fisker Hag, A.M.; Knigge, U.; Kjaer, A. 18F-FDG and 18F-FLT-PET imaging for monitoring everolimus effect on tumor-growth in neuroendocrine tumors: Studies in human tumor xenografts in mice. PLoS ONE 2014, 9, e91387. [Google Scholar] [CrossRef]
- Petersen, A.L.; Binderup, T.; Jolck, R.I.; Rasmussen, P.; Henriksen, J.R.; Pfeifer, A.K.; Kjaer, A.; Andresen, T.L. Positron emission tomography evaluation of somatostatin receptor targeted 64Cu-TATE-liposomes in a human neuroendocrine carcinoma mouse model. J. Control Release 2012, 160, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Zuetenhorst, J.M.; Bonfrer, J.M.; Korse, C.M.; Bakker, R.; van Tinteren, H.; Taal, B.G. Carcinoid heart disease: The role of urinary 5-hydroxyindoleacetic acid excretion and plasma levels of atrial natriuretic peptide, transforming growth factor-beta and fibroblast growth factor. Cancer 2003, 97, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Sari, Y.; Zhou, F.C. Serotonin and its transporter on proliferation of fetal heart cells. Int. J. Dev. Neurosci. 2003, 21, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Contractor, T.; Harris, C.R. Loss of copy of MIR1-2 increases CDK4 expression in ileal neuroendocrine tumors. Oncogenesis 2020, 9, 37. [Google Scholar] [CrossRef]
- Matthes, S.; Bader, M. Peripheral Serotonin Synthesis as a New Drug Target. Trends Pharmacol. Sci. 2018, 39, 560–572. [Google Scholar] [CrossRef]
- Cives, M.; Strosberg, J. Treatment Strategies for Metastatic Neuroendocrine Tumors of the Gastrointestinal Tract. Curr. Treat. Options Oncol. 2017, 18, 14. [Google Scholar] [CrossRef]
- Horsch, D.; Anthony, L.; Gross, D.J.; Valle, J.W.; Welin, S.; Benavent, M.; Caplin, M.; Pavel, M.; Bergsland, E.; Oberg, K.; et al. Long-Term Treatment with Telotristat Ethyl in Patients with Carcinoid Syndrome Symptoms: Results from the TELEPATH Study. Neuroendocrinology 2022, 112, 298–310. [Google Scholar] [CrossRef]
- Herrera-Martinez, A.D.; Feelders, R.A.; Van den Dungen, R.; Dogan-Oruc, F.; van Koetsveld, P.M.; Castano, J.P.; de Herder, W.W.; Hofland, L.J. Effect of the Tryptophan Hydroxylase Inhibitor Telotristat on Growth and Serotonin Secretion in 2D and 3D Cultured Pancreatic Neuroendocrine Tumor Cells. Neuroendocrinology 2020, 110, 351–363. [Google Scholar] [CrossRef]
- Shi, H.; Cui, Y.; Qin, Y. Discovery and characterization of a novel tryptophan hydroxylase 1 inhibitor as a prodrug. Chem. Biol. Drug Des. 2018, 91, 202–212. [Google Scholar] [CrossRef]
- Dillon, J.S.; Chandrasekharan, C. Telotristat ethyl: A novel agent for the therapy of carcinoid syndrome diarrhea. Future Oncol. 2018, 14, 1155–1164. [Google Scholar] [CrossRef]
- Kiesewetter, B.; Raderer, M. Ondansetron for diarrhea associated with neuroendocrine tumors. N. Engl. J. Med. 2013, 368, 1947–1948. [Google Scholar] [CrossRef] [PubMed]
- Waldum, H.; Wahba, A. Serotonin-A Driver of Progressive Heart Valve Disease. Front. Cardiovasc. Med. 2022, 9, 774573. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Grafton, G.; Powell, A.D.; Brock, K.; Chen, C.; Xie, D.; Huang, J.; Liu, S.; Cooper, A.J.; Brady, C.A.; et al. CSTI-300 (SMP-100); a Novel 5-HT3 Receptor Partial Agonist with Potential to Treat Patients with Irritable Bowel Syndrome or Carcinoid Syndrome. J. Pharmacol. Exp. Ther. 2020, 373, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Cozzolino, A.; Malandrino, P.; Minotta, R.; Puliani, G.; Saronni, D.; Faggiano, A.; Colao, A. Role of FGF System in Neuroendocrine Neoplasms: Potential Therapeutic Applications. Front. Endocrinol. 2021, 12, 665631. [Google Scholar] [CrossRef] [PubMed]
- Biasco, E.; Antonuzzo, A.; Galli, L.; Baldi, G.G.; Derosa, L.; Marconcini, R.; Farnesi, A.; Ricci, S.; Falcone, A. Small-bowel neuroendocrine tumor and retroperitoneal fibrosis: Efficacy of octreotide and tamoxifen. Tumori 2015, 101, e24–e28. [Google Scholar] [CrossRef]
- Luo, J.; Manning, B.D.; Cantley, L.C. Targeting the PI3K-Akt pathway in human cancer: Rationale and promise. Cancer Cell 2003, 4, 257–262. [Google Scholar] [CrossRef]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef]
- Voortman, J.; Lee, J.H.; Killian, J.K.; Suuriniemi, M.; Wang, Y.; Lucchi, M.; Smith, W.I., Jr.; Meltzer, P.; Wang, Y.; Giaccone, G. Array comparative genomic hybridization-based characterization of genetic alterations in pulmonary neuroendocrine tumors. Proc. Natl. Acad. Sci. USA 2010, 107, 13040–13045. [Google Scholar] [CrossRef]
- Valentino, J.D.; Li, J.; Zaytseva, Y.Y.; Mustain, W.C.; Elliott, V.A.; Kim, J.T.; Harris, J.W.; Campbell, K.; Weiss, H.; Wang, C.; et al. Cotargeting the PI3K and RAS pathways for the treatment of neuroendocrine tumors. Clin. Cancer Res. 2014, 20, 1212–1222. [Google Scholar] [CrossRef]
- Silva, S.R.; Zaytseva, Y.Y.; Jackson, L.N.; Lee, E.Y.; Weiss, H.L.; Bowen, K.A.; Townsend, C.M., Jr.; Evers, B.M. The effect of PTEN on serotonin synthesis and secretion from the carcinoid cell line BON. Anticancer Res. 2011, 31, 1153–1160. [Google Scholar]
- Chan, J.; Kulke, M. Targeting the mTOR signaling pathway in neuroendocrine tumors. Curr. Treat. Options Oncol. 2014, 15, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Orr-Asman, M.A.; Chu, Z.; Jiang, M.; Worley, M.; LaSance, K.; Koch, S.E.; Carreira, V.S.; Dahche, H.M.; Plas, D.R.; Komurov, K.; et al. mTOR Kinase Inhibition Effectively Decreases Progression of a Subset of Neuroendocrine Tumors that Progress on Rapalog Therapy and Delays Cardiac Impairment. Mol. Cancer Ther. 2017, 16, 2432–2441. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Martinez, A.D.; Pedraza-Arevalo, S.; L-López, F.; Gahete, M.D.; Galvez-Moreno, M.A.; Castano, J.P.; Luque, R.M. Type 2 Diabetes in Neuroendocrine Tumors: Are Biguanides and Statins Part of the Solution? J. Clin. Endocrinol. Metab. 2019, 104, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Das, S.; Miller, R.; Luque, L.; Cheuvront, S.N.; Cloud, J.; Tarter, Z.; Siddiqui, F.; Ramirez, R.A.; Anthony, L. Can an amino acid mixture alleviate gastrointestinal symptoms in neuroendocrine tumor patients? BMC Cancer 2021, 21, 580. [Google Scholar] [CrossRef]
- Yin, L.; Vijaygopal, P.; MacGregor, G.G.; Menon, R.; Ranganathan, P.; Prabhakaran, S.; Zhang, L.; Zhang, M.; Binder, H.J.; Okunieff, P.; et al. Glucose stimulates calcium-activated chloride secretion in small intestinal cells. Am. J. Physiol. Cell Physiol. 2014, 306, C687–C696. [Google Scholar] [CrossRef]
- Jacob, N.; Dasharathy, S.S.; Bui, V.; Benhammou, J.N.; Grody, W.W.; Singh, R.R.; Pisegna, J.R. Generalized Cytokine Increase in the Setting of a Multisystem Clinical Disorder and Carcinoid Syndrome Associated with a Novel NLRP12 Variant. Dig. Dis. Sci. 2019, 64, 2140–2146. [Google Scholar] [CrossRef]
- Cervini-Silva, J.; Nieto-Camacho, A.; Kaufhold, S.; Ufer, K.; Palacios, E.; Montoya, A.; Dath, W. Antiphlogistic effect by zeolite as determined by a murine inflammation model. Microporous Mesoporous Mater. 2016, 228, 207–214. [Google Scholar] [CrossRef]
- April-Monn, S.L.; Wiedmer, T.; Skowronska, M.; Maire, R.; Lena, M.S.; Trippel, M.; Di Domenico, A.; Muffatti, F.; Andreasi, V.; Capurso, G.; et al. Three-Dimensional Primary Cell Culture: A Novel Preclinical Model for Pancreatic Neuroendocrine Tumors. Neuroendocrinology 2021, 111, 273–287. [Google Scholar] [CrossRef]
- Detjen, K.; Hammerich, L.; Ozdirik, B.; Demir, M.; Wiedenmann, B.; Tacke, F.; Jann, H.; Roderburg, C. Models of Gastroenteropancreatic Neuroendocrine Neoplasms: Current Status and Future Directions. Neuroendocrinology 2021, 111, 217–236. [Google Scholar] [CrossRef]
- Gillette, A.A.; Babiarz, C.P.; VanDommelen, A.R.; Pasch, C.A.; Clipson, L.; Matkowskyj, K.A.; Deming, D.A.; Skala, M.C. Autofluorescence Imaging of Treatment Response in Neuroendocrine Tumor Organoids. Cancers 2021, 13, 1873. [Google Scholar] [CrossRef]
- Laskaratos, F.M.; Rombouts, K.; Caplin, M.; Toumpanakis, C.; Thirlwell, C.; Mandair, D. Neuroendocrine tumors and fibrosis: An unsolved mystery? Cancer 2017, 123, 4770–4790. [Google Scholar] [CrossRef] [PubMed]
- Svejda, B.; Kidd, M.; Giovinazzo, F.; Eltawil, K.; Gustafsson, B.I.; Pfragner, R.; Modlin, I.M. The 5-HT(2B) receptor plays a key regulatory role in both neuroendocrine tumor cell proliferation and the modulation of the fibroblast component of the neoplastic microenvironment. Cancer 2010, 116, 2902–2912. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, L.W.; Burn, T.C.; Brown, B.S.; Patterson, J.P.; Corjay, M.H.; Valentine, P.A.; Sun, J.H.; Link, J.R.; Abbaszade, I.; Hollis, J.M.; et al. Possible role of valvular serotonin 5-HT(2B) receptors in the cardiopathy associated with fenfluramine. Mol. Pharmacol. 2000, 57, 75–81. [Google Scholar] [PubMed]
- Setola, V.; Hufeisen, S.J.; Grande-Allen, K.J.; Vesely, I.; Glennon, R.A.; Blough, B.; Rothman, R.B.; Roth, B.L. 3,4-methylenedioxymethamphetamine (MDMA, “Ecstasy”) induces fenfluramine-like proliferative actions on human cardiac valvular interstitial cells in vitro. Mol. Pharmacol. 2003, 63, 1223–1229. [Google Scholar] [CrossRef]
- Herring, B.; Jang, S.; Whitt, J.; Goliwas, K.; Aburjania, Z.; Dudeja, V.; Ren, B.; Berry, J.; Bibb, J.; Frost, A.; et al. Ex Vivo Modeling of Human Neuroendocrine Tumors in Tissue Surrogates. Front. Endocrinol. 2021, 12, 710009. [Google Scholar] [CrossRef]
- Carter, A.M.; Kumar, N.; Herring, B.; Tan, C.; Guenter, R.; Telange, R.; Howse, W.; Viol, F.; McCaw, T.R.; Bickerton, H.H.; et al. Cdk5 drives formation of heterogeneous pancreatic neuroendocrine tumors. Oncogenesis 2021, 10, 83. [Google Scholar] [CrossRef]
- Pozo, K.; Castro-Rivera, E.; Tan, C.; Plattner, F.; Schwach, G.; Siegl, V.; Meyer, D.; Guo, A.; Gundara, J.; Mettlach, G.; et al. The role of Cdk5 in neuroendocrine thyroid cancer. Cancer Cell 2013, 24, 499–511. [Google Scholar] [CrossRef]
- Biran, J.; Blechman, J.; Wircer, E.; Levkowitz, G. Development and Function of the Zebrafish Neuroendocrine System. In Model Animals in Neuroendocrinology: From Worm to Mouse to Man; Ludwig, M., Levkowitz, G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Gaudenzi, G.; Carra, S.; Dicitore, A.; Cantone, M.C.; Persani, L.; Vitale, G. Fishing for neuroendocrine tumors. Endocr. Relat. Cancer 2020, 27, R163–R176. [Google Scholar] [CrossRef]
- Njagi, J.; Ball, M.; Best, M.; Wallace, K.N.; Andreescu, S. Electrochemical quantification of serotonin in the live embryonic zebrafish intestine. Anal. Chem. 2010, 82, 1822–1830. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Yao, T.; Jia, R. Benefits of Zebrafish Xenograft Models in Cancer Research. Front. Cell Dev. Biol. 2021, 9, 616551. [Google Scholar] [CrossRef]
- Fior, R.; Povoa, V.; Mendes, R.V.; Carvalho, T.; Gomes, A.; Figueiredo, N.; Ferreira, M.G. Single-cell functional and chemosensitive profiling of combinatorial colorectal therapy in zebrafish xenografts. Proc. Natl. Acad. Sci. USA 2017, 114, E8234–E8243. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Brunson, D.C.; Tang, Q.; Do, D.; Iftimia, N.A.; Moore, J.C.; Hayes, M.N.; Welker, A.M.; Garcia, E.G.; Dubash, T.D.; et al. Visualizing Engrafted Human Cancer and Therapy Responses in Immunodeficient Zebrafish. Cell 2019, 177, 1903–1914.e14. [Google Scholar] [CrossRef] [PubMed]
- Carra, S.; Gaudenzi, G. New perspectives in neuroendocrine neoplasms research from tumor xenografts in zebrafish embryos. Minerva Endocrinol. 2020, 45, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Carra, S.; Gaudenzi, G.; Dicitore, A.; Cantone, M.C.; Plebani, A.; Saronni, D.; Zappavigna, S.; Caraglia, M.; Candeo, A.; Bassi, A.; et al. Modeling Lung Carcinoids with Zebrafish Tumor Xenograft. Int. J. Mol. Sci. 2022, 23, 8126. [Google Scholar] [CrossRef]
- Carra, S.; Gaudenzi, G.; Dicitore, A.; Saronni, D.; Cantone, M.C.; Plebani, A.; Ghilardi, A.; Borghi, M.O.; Hofland, L.J.; Persani, L.; et al. Vandetanib versus Cabozantinib in Medullary Thyroid Carcinoma: A Focus on Anti-Angiogenic Effects in Zebrafish Model. Int. J. Mol. Sci. 2021, 22, 3031. [Google Scholar] [CrossRef]
- Dicitore, A.; Saronni, D.; Gaudenzi, G.; Carra, S.; Cantone, M.C.; Borghi, M.O.; Persani, L.; Vitale, G. Long-term effects of somatostatin analogues in rat GH-secreting pituitary tumor cell lines. J. Endocrinol. Investig. 2022, 45, 29–41. [Google Scholar] [CrossRef]
- Gaudenzi, G.; Albertelli, M.; Dicitore, A.; Wurth, R.; Gatto, F.; Barbieri, F.; Cotelli, F.; Florio, T.; Ferone, D.; Persani, L.; et al. Patient-derived xenograft in zebrafish embryos: A new platform for translational research in neuroendocrine tumors. Endocrine 2017, 57, 214–219. [Google Scholar] [CrossRef]
- Peverelli, E.; Giardino, E.; Treppiedi, D.; Meregalli, M.; Belicchi, M.; Vaira, V.; Corbetta, S.; Verdelli, C.; Verrua, E.; Serban, A.L.; et al. Dopamine receptor type 2 (DRD2) and somatostatin receptor type 2 (SSTR2) agonists are effective in inhibiting proliferation of progenitor/stem-like cells isolated from nonfunctioning pituitary tumors. Int. J. Cancer 2017, 140, 1870–1880. [Google Scholar] [CrossRef]
- Vitale, G.; Gaudenzi, G.; Circelli, L.; Manzoni, M.F.; Bassi, A.; Fioritti, N.; Faggiano, A.; Colao, A.; Group, N. Animal models of medullary thyroid cancer: State of the art and view to the future. Endocr. Relat. Cancer 2017, 24, R1–R12. [Google Scholar] [CrossRef]
- Vitale, G.; Gaudenzi, G.; Dicitore, A.; Cotelli, F.; Ferone, D.; Persani, L. Zebrafish as an innovative model for neuroendocrine tumors. Endocr. Relat. Cancer 2014, 21, R67–R83. [Google Scholar] [CrossRef]
- Saronni, D.; Gaudenzi, G.; Dicitore, A.; Carra, S.; Cantone, M.C.; Borghi, M.O.; Barbieri, A.; Mignani, L.; Hofland, L.J.; Persani, L.; et al. Preclinical Evaluation of Novel Tyrosine-Kinase Inhibitors in Medullary Thyroid Cancer. Cancers 2022, 14, 4442. [Google Scholar] [CrossRef] [PubMed]
- Lawson, N.D.; Weinstein, B.M. In Vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Monazzam, A.; Li, S.C.; Wargelius, H.; Razmara, M.; Bajic, D.; Mi, J.; Bergquist, J.; Crona, J.; Skogseid, B. Generation and characterization of CRISPR/Cas9-mediated MEN1 knockout BON1 cells: A human pancreatic neuroendocrine cell line. Sci. Rep. 2020, 10, 14572. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rosa, J.M. Zebrafish Models of Cardiac Disease: From Fortuitous Mutants to Precision Medicine. Circ. Res. 2022, 130, 1803–1826. [Google Scholar] [CrossRef] [PubMed]
- Gunawan, F.; Gentile, A.; Fukuda, R.; Tsedeke, A.T.; Jimenez-Amilburu, V.; Ramadass, R.; Iida, A.; Sehara-Fujisawa, A.; Stainier, D.Y.R. Focal adhesions are essential to drive zebrafish heart valve morphogenesis. J. Cell. Biol. 2019, 218, 1039–1054. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.T.; Bartman, T. Analysis of heart valve development in larval zebrafish. Dev. Dyn. 2009, 238, 1796–1802. [Google Scholar] [CrossRef]
- Yelon, D.; Horne, S.A.; Stainier, D.Y. Restricted expression of cardiac myosin genes reveals regulated aspects of heart tube assembly in zebrafish. Dev. Biol. 1999, 214, 23–37. [Google Scholar] [CrossRef]
- Wang, L.W.; Huttner, I.G.; Santiago, C.F.; Kesteven, S.H.; Yu, Z.Y.; Feneley, M.P.; Fatkin, D. Standardized echocardiographic assessment of cardiac function in normal adult zebrafish and heart disease models. Dis. Model Mech. 2017, 10, 63–76. [Google Scholar] [CrossRef]
- Wang, X.; Copmans, D.; de Witte, P.A.M. Using Zebrafish as a Disease Model to Study Fibrotic Disease. Int. J. Mol. Sci. 2021, 22, 6404. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cell Line | Source | Biochemical and Molecular Properties | Drug Sensitivity | References |
|---|---|---|---|---|
| CGP | Jejunum | Low proliferation rate; synthesis, store, and release of 5-HT and histamine | Unknown | [19] |
| KRJ-I | Small intestine | Secretion of 5-HT, noradrenaline and pituitary adenylate cyclase; expression of CgA, NSE, Ki-67, TPH-1, substance P and guanylin | High resistance to octreotide-mediated 5-HT secretion inhibition; sensitivity to octreotide + RAD001 co-treatment and AN-238 | [20,21,22,23,24] |
| BON-1 | Pancreas | Synthesis of 5-HT, CgA, neurotensin and pancreastatin; expression of gastrin, somatostatin, 5-HT, and acetylcholine receptors; expression of IGF and IGFR | Sensitivity to imatinib, leflunomide, sodium butyrate, hexamethylene bisacetamide, everolimus, pasireotide, LY294002, MK-2206, octreotide, IFN-β and BYL719 | [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39] |
| QGP-1 | Pancreas | Expression of TPH-1, CgA, synaptophysin, VMAT1, mGluR4, ADB1, ACM4, substance P, SERT and guanylin | Sensitivity to everolimus, octreotide and BYL719 | [33,36,40,41] |
| NCI-H727 | Bronchus (typical) | Expression of p53, SST-2 and SST-5; secretion of calcium-stimulated PTHLP | Sensitivity to EGFR monoclonal antibodies, LY294002, SSA, BYL719, BIM23120 and BIM23A779 | [36,42,43,44] |
| NCI-H720 | Bronchus (atypical) | Expression of SST-2 and SST-5 | Sensitivity to NSC 95397, brefeldin A, bortezomib, lanreotide, BIM23206 and BIM23120 | [45,46] |
| COLO 320 DM | Colon | Synthesis of 5-HT, parathyroid hormone, ACTH, norepinephrine and epinephrine | Sensitivity to oxaliplatin | [47,48] |
| GOT1 | Ileum | Expression of all recognized somatostatin receptors, VMAT1 and VMAT2 | Binding to radiolabeled SSA | [49] |
| Animal Model | Laboratory, Molecular and Biological Properties | Tested Drugs | Results | References |
|---|---|---|---|---|
| GOT1-bearing nude mice | - Increased plasma 5-HT levels and 5-HIAA urine levels. - Tumors express somatostatin receptors and VMAT1 and VMAT2. | All xenografted tumors could be visualized scintigraphically using the SSA 111In-octreotide and 123I-MIBG. | [49] | |
| 177Lu-DOTATATE and then111In-DOTATATE. | Suboptimal therapeutic amounts of 177Lu-DOTATATE caused an increased uptake of the second injection (111In-DOTATATE). | [53] | ||
| BON-1-bearing athymic nude mice | - Liver metastases in 65% with elevated platelet 5-HT levels and fibrosis on the valvular tissue (above all on tricuspid valve). | [54] | ||
| BON-1- bearing athymic nude mice | - Increased plasma 5-HT levels and 5-HIAA urine levels. - Liver metastasis with positive staining for 5-HT and CgA. - Fibrosis, diarrhea and fibrotic cardiac valvular disease (tricuspid and mitral thickening). | Octreotide/bevacizumab | Octreotide/bevacizumab reduce liver metastasis and manifestation of CS, including valvular heart disease. | [55] |
| Sprague–Dawley rats | Subcutaneous injections of 5-HT daily for 3 months | - Increased plasma 5-HT levels. - Flushing, loose stools and anorexia. - Cardiac disease with pathological echocardiographs and histopathological changes (shortened and thickened aortic cusps with an increased cellularity of myofibroblasts in a collagenous matrix). | [56] | |
| Sprague–Dawley rats | Subcutaneous injections of 5-HT daily for 7 days | - Higher amount of glycosaminoglycans and a lower amount of collagen. - Thickening and compositional alteration of aortic and mitral valves. | [57] | |
| New Zealand white rabbits | Long term oral administration of 5-HT | - Increased plasma 5-HT levels and 5-HIAA urine levels. - Valvular heart disease with thickened aortic, mitral and tricuspid leaflets and several areas of chondroid metaplasia. | [58] | |
| Transgenic mice overexpressing the Gq-coupled 5-HT2BR specifically in the heart | - Proliferation of the mitochondria. - Hypertrophic cardiomyopathy. | [59] | ||
| 5-HTT-deficient mice | - Increased and persistent interactions between 5-HT and 5-HTR and valvular mitogenic activity with extracellular matrix production. - Structural and functional cardiac abnormalities and valvulopathy. | [60] | ||
| Sprague–Dawley rats transplanted intraocularly with midgut carcinoid tumors secreting 5-HT | Application of adrenoceptor agonists locally to the eye | - The activation of beta-adrenoceptors (Isoprenaline) causes release of 5-HT from carcinoid tumor cells. - The stimulation with alpha-adrenoceptors (Norepinephrine) did not elicit any 5-HT release. | [61] | |
| Wistar rats | Oral administration of PCPA and PEPA | Reduced plasma 5-HT levels and 5-HIAA urine levels. | [62] | |
| Sprague–Dawley rats | Subcutaneous injections of 5-HT daily for 4 months and then also with Terguride | - Vasodilatation and decreased heart rate. Block of serotonin-induced changes in the skin (achantosis). - Not heart/liver/stomach weight gain or right-sided echocardiographic changes. - Flushing. | [63] | |
| Transgenic RT2 mice with B6AF1 genetic background | - Loss of imprinting of IGF2 with its overexpression. - Development of ileal NETs. - Secretion of 5-HT in 22% of ileal NETs. | IGF2 is considered as the first genetic driver of ileal neuroendocrine tumorigenesis. | [64] | |
| C57BL/6 and C57 albino mice | Oral administration of LP-920540 and LX1032 | - Reduced 5-HT levels in the intestinal mucosa and in plasma. Neither brain 5-HT nor 5-HIAA urine levels were affected significantly. - Improvement of colonic motility. | [65] | |
| C57BL/6 mice models of intestinal inflammation | Oral administration of LX1032/LX1606 | - Decreased pro-inflammatory cytokine levels and 5-HT intestinal levels. - Reduced colitis severity and diarrhea episodes. | [66] |
| Model | Pros | Cons |
|---|---|---|
| Immortalized NET cell lines | Useful for the study of disease mechanisms and drug efficacy in a controlled environment. Reliable and reproducible results. They provide a means for testing a large number of compounds or interventions in a relatively short time frame. | They may not accurately represent the complexity and heterogeneity of the disease in humans. They may not predict the efficacy or toxicity of a drug in patients with NETs. They may lose characteristics of the original tumor (e.g., secretory function). |
| Primary cultures of human NETs | A more accurate representation than immortalized cell lines that have been passaged multiple times in culture. Representative of the tumor and the host microenvironment. Cost-effective compared to in vivo models. | Difficult to establish and maintain. High variability. Limited lifespan due to the onset of senescence, which makes them difficult to be used for long-term studies. |
| Xenotransplantation of NET cells in nude mice | Most of these animals developed liver metastases with elevated platelet 5-HT levels. Most of these animals developed mesenteric fibrosis, diarrhea and fibrotic cardiac valvular disease reminiscent of CS. They provide a more accurate representation of the disease than cell culture models, allowing for the study of the interactions between the tumor and the host and of drug efficacy and toxicity in a whole organism. | They may not accurately represent the complexity and heterogeneity of the disease in humans as the xenografts are grown in an immunodeficient mouse, which does not have a human immune system. Cons related to the injection of immortalized cell lines previously reported. PDX are extremely difficult to be developed, due to the low proliferation rate of primary cultures. Expensive. |
| Administration of 5-HT in rabbit models or transgenic mouse models with alterations of 5-HT pathway | 5-HT injections induced clinical signs observed in patients with CS (flushing, loose stools and anorexia). Long-term 5-HT overload can cause valvular heart disease, similar to that reported in patients with CHD. | They may not accurately represent the complexity and heterogeneity of the disease in humans. They may not predict the efficacy or toxicity of a drug in patients with NETs. |
| Mouse models of intestinal inflammation | Adopted to test the effects of TPH inhibitors on 5-HT intestinal levels, colitis severity and diarrhea episodes. | They may not accurately represent the complexity and heterogeneity of the disease in humans because they are not specific models of CS/CHD. |
| Primary 3D-culture and co-culture models | Maintenance of 3D architecture of the tissue, providing a faithful representation of stromal and extracellular matrix contributions. Possibility to study the crosstalk between selected cell populations involved in CS. Possibility to study the secretion of different factors from primary cells derived from CS patients. Possibility to perform drug screening. Omic technologies could be combined with these platforms to dissect specific molecular mechanisms, with the aim to discover novel biomarkers and therapeutic targets for the CS. | Difficulties in cell culture due to the low proliferation rate of primary NET cells. Limited representation of the disease due to the loss of interactions between tumor cells and the whole organism. Models to be validated for CS. |
| Zebrafish xenograft models | Tumor implant with low number of cells. No tumor cell rejection in embryos and larvae due to the lack of a fully developed immune system. High number of embryos are available for each experiment. High engraftment rate. In vivo and real-time visualization of tumor implants. Embryo tissue permeability to small molecules, useful for drug-screening. | Differences between the optimal temperature for zebrafish development and mammalian cell metabolism. Difficulties in recapitulating all clinical CS manifestations. Difficulties for long-term analyses. Difficulties in administration of molecules with low water solubility. Models to be validated for CS. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitale, G.; Carra, S.; Alessi, Y.; Campolo, F.; Pandozzi, C.; Zanata, I.; Colao, A.; Faggiano, A.; on behalf of the NIKE Group. Carcinoid Syndrome: Preclinical Models and Future Therapeutic Strategies. Int. J. Mol. Sci. 2023, 24, 3610. https://doi.org/10.3390/ijms24043610
Vitale G, Carra S, Alessi Y, Campolo F, Pandozzi C, Zanata I, Colao A, Faggiano A, on behalf of the NIKE Group. Carcinoid Syndrome: Preclinical Models and Future Therapeutic Strategies. International Journal of Molecular Sciences. 2023; 24(4):3610. https://doi.org/10.3390/ijms24043610
Chicago/Turabian StyleVitale, Giovanni, Silvia Carra, Ylenia Alessi, Federica Campolo, Carla Pandozzi, Isabella Zanata, Annamaria Colao, Antongiulio Faggiano, and on behalf of the NIKE Group. 2023. "Carcinoid Syndrome: Preclinical Models and Future Therapeutic Strategies" International Journal of Molecular Sciences 24, no. 4: 3610. https://doi.org/10.3390/ijms24043610
APA StyleVitale, G., Carra, S., Alessi, Y., Campolo, F., Pandozzi, C., Zanata, I., Colao, A., Faggiano, A., & on behalf of the NIKE Group. (2023). Carcinoid Syndrome: Preclinical Models and Future Therapeutic Strategies. International Journal of Molecular Sciences, 24(4), 3610. https://doi.org/10.3390/ijms24043610