
Co-Expression Analysis of Airway Epithelial Transcriptome in Asthma Patients with Eosinophilic vs. Non-Eosinophilic Airway Infiltration
 , , , ,
, , , , 

Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of the Patients
2.2. Functional and Structural Measures of Airway Remodeling Were Similar in EA and NEA
2.3. BAL and Blood Eosinophils Were Well Correlated
2.4. Blood and Bronchoalveolar Cytokine Profiles Were Separately Correlated with Eosinophilia in Each Compartment
2.5. Differentially Expressed Genes Are Involved in Processes Associated with Asthma Pathogenesis
2.6. Differentially Expressed Genes Correlate with Lung Function and Proxies of Airway Remodeling but Not Reticular Basement Membrane Thickness
2.7. Differential Co-Expression Analysis Reveals Genes Co-Regulated in Eosinophilic Asthma
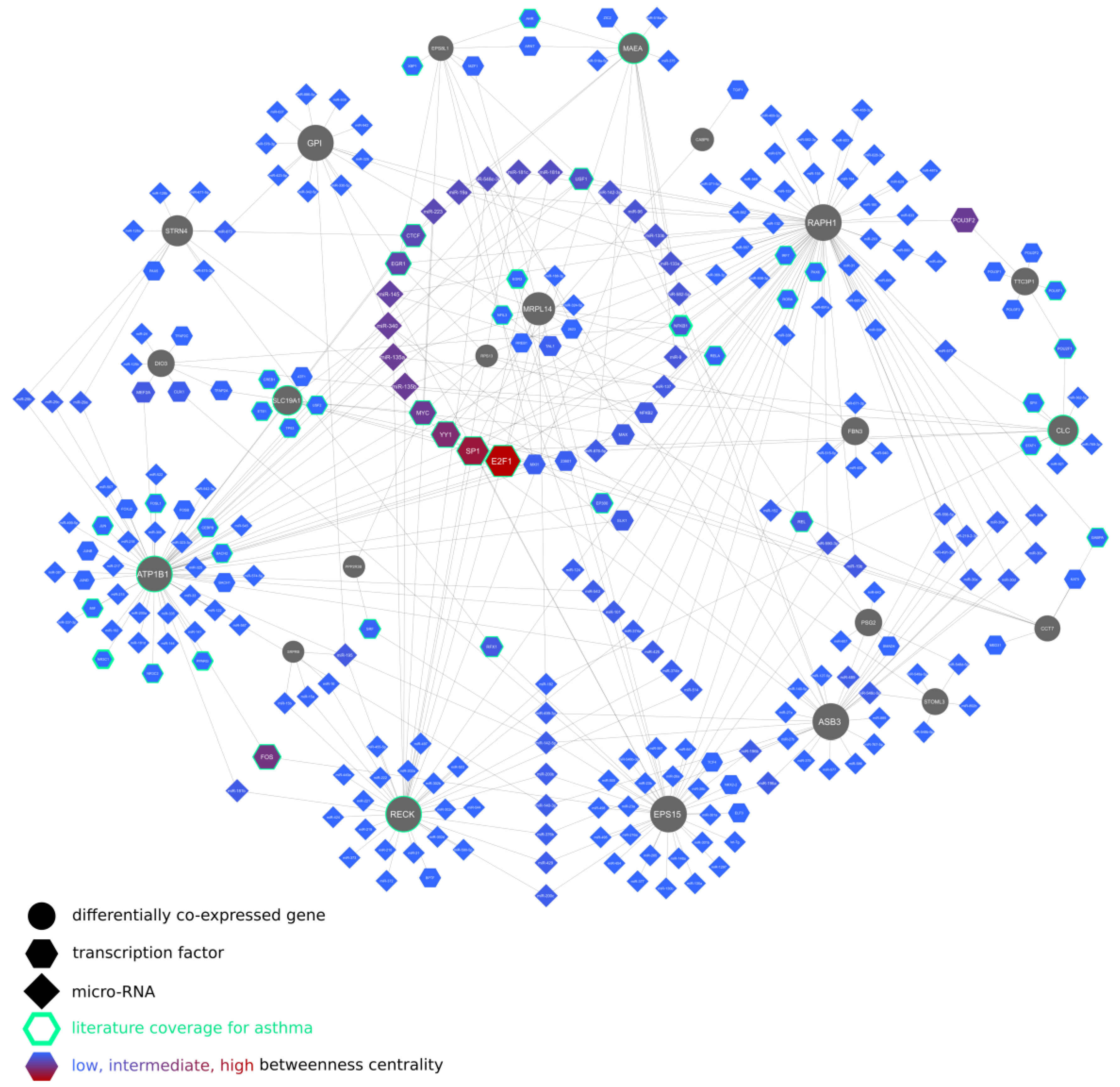
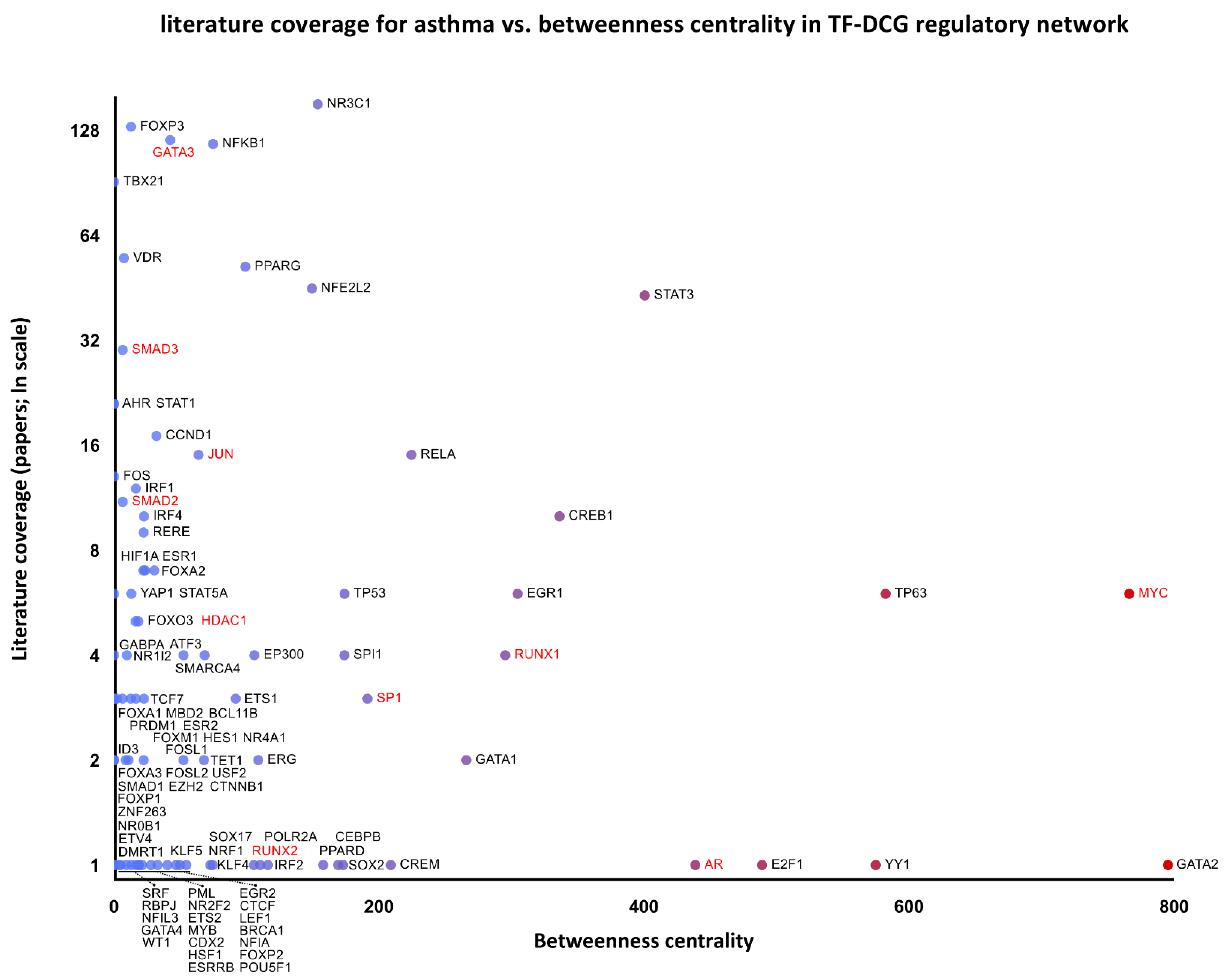
2.8. Regulatory Networks of the Differentially Co-Expressed Genes and the Inferred Signaling Pathways
2.9. Kinase Perturbation Studies
2.10. Cell-Type-Specific Histone Modifications Related to the Expression of Differentially Co-Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Characteristics of the Patients and Study Design
4.2. Airway Cross-Sectional Geometry in Lung Computed Tomography
4.3. Bronchofiberoscopy and Airway Sampling
4.4. Bronchial Biopsy Histology and Reticular Basement Membrane Measurement
4.5. Measurements of Immune Mediators in Serum Samples and BAL Supernatant
4.6. RNA Isolation and Microarray Processing
4.7. Retrieval of Microarray Data
4.8. Basic Statistical Analysis
4.9. Statistical Analysis of Differential Gene Expression and Co-Expression
4.10. Gene Set Enrichment, Interaction Networks, and Their Topological Robustness Analysis
- Gene set enrichment analysis (GSEA) to establish histone modifications and cell types matching differentially co-expressed genes’ expression pattern and associated biological processes, molecular functions, and signaling pathways, using Enrichr tool [76] and curated datasets (Supplement Table S6);
- Analysis of the networks of associated molecular entity interactions, i.e., (1) lung-specific binary protein–protein interactions of the proteins coded by the differentially co-expressed genes, (2) interactions between differentially co-expressed genes’ and their upstream regulators, and (3) interactions between differentially co-expressed genes, microRNAs, and associated transcription factors; the networks were constructed and analyzed using Cytoscape 3.7.1 [77] and NetworkAnalyst 3.0 [78];
- The topological robustness analysis of the above networks (graphs) by the principle of topological attack on biological network; betweenness centrality identified nodes with the highest network-disrupting potential as possible targets for pharmacological interventions.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kozlik, P.; Zuk, J.; Bartyzel, S.; Zarychta, J.; Okon, K.; Zareba, L.; Bazan, J.G.; Kosalka, J.; Soja, J.; Musial, J.; et al. The relationship of airway structural changes to blood and bronchoalveolar lavage biomarkers, and lung function abnormalities in asthma. Clin. Exp. Allergy 2020, 50, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Svenningsen, S.; Nair, P. Asthma Endotypes and an Overview of Targeted Therapy for Asthma. Front. Med. 2017, 4, 158. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G. Inflammatory phenotypes in adult asthma: Clinical applications. Clin. Respir. J. 2009, 3, 198–206. [Google Scholar] [CrossRef]
- Bazan-Socha, S.; Bukiej, A.; Marcinkiewicz, C.; Musial, J. Integrins in Pulmonary Inflammatory Diseases. Curr. Pharm. Des. 2005, 11, 893–901. [Google Scholar] [CrossRef]
- Pascual, R.M.; Peters, S.P. Airway remodeling contributes to the progressive loss of lung function in asthma: An overview. J. Allergy Clin. Immunol. 2005, 116, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Fukuda, Y.; Tanaka, A.; Sagara, H. Comparative Efficacy and Safety of Tezepelumab and Other Biologics in Patients with Inadequately Controlled Asthma According to Thresholds of Type 2 Inflammatory Biomarkers: A Systematic Review and Network Meta-Analysis. Cells 2022, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- Flood-Page, P.; Swenson, C.; Faiferman, I.; Matthews, J.; Williams, M.; Brannick, L.; Robinson, D.; Wenzel, S.; Busse, W.; Hansel, T.T.; et al. A Study to Evaluate Safety and Efficacy of Mepolizumab in Patients with Moderate Persistent Asthma. Am. J. Respir. Crit. Care Med. 2007, 176, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.; Brown, T.; Elliott, S.; Barber, C.; Gove, K.; Lau, L.; Rupani, H.; Chauhan, A.; Howarth, P.; Ono, J.; et al. Predictors of sputum and BAL eosinophilia in the Wessex Severe Asthma Cohort. Eur. Res. J. 2017, 50, PA4765. [Google Scholar] [CrossRef]
- Yancey, S.W.; Bradford, E.S.; Keene, O.N. Disease burden and efficacy of mepolizumab in patients with severe asthma and blood eosinophil counts of ≥150–300 cells/μL. Respir. Med. 2019, 151, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Hviid-Vyff, B.; Skjold, T.; Hoffmann, H.J. The usefulness of blood eosinophil count and FeNO to predict sputum eosinophilia in the diagnosis of severe eosinophilic asthma. Eur. Res. J. 2019, 54, OA2150. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.-H.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef] [PubMed]
- van Veen, I.H.; Brinke, A.T.; Gauw, S.A.; Sterk, P.J.; Rabe, K.F.; Bel, E.H. Consistency of sputum eosinophilia in difficult-to-treat asthma: A 5-year follow-up study. J. Allergy Clin. Immunol. 2009, 124, 615–617.e2. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ovando, S.; Simpson, J.L.; Barker, D.; Baines, K.J.; Wark, P.A.B. Transcriptomics of biopsies identifies novel genes and pathways linked to neutrophilic inflammation in severe asthma. Clin. Exp. Allergy 2021, 51, 1279–1294. [Google Scholar] [CrossRef]
- Baines, K.J.; Simpson, J.L.; Wood, L.G.; Scott, R.J.; Fibbens, N.L.; Powell, H.; Cowan, D.C.; Taylor, D.R.; Cowan, J.O.; Gibson, P.G. Sputum gene expression signature of 6 biomarkers discriminates asthma inflammatory phenotypes. J. Allergy Clin. Immunol. 2014, 133, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Manabe, Y.; Shiga, M.; Kometani-Gunjigake, K.; Nakao-Kuroishi, K.; Mizuhara, M.; Toyono, T.; Seta, Y.; Kawamoto, T. Fibrillin-1 regulates periostin expression during maintenance of periodontal homeostasis. J. Dent. Sci. 2022, 17, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, J.; Dieckow, J.; Schröder, H.; Hampel, U.; Garreis, F.; Jacobi, C.; Milczarek, A.; Hsieh, K.L.; Pulli, B.; Chen, J.W.; et al. Plasma gelsolin promotes re-epithelialization. Sci. Rep. 2018, 8, 13140. [Google Scholar] [CrossRef]
- Suojalehto, H.; Ndika, J.; Lindström, I.; Airaksinen, L.; Karisola, P.; Alenius, H. Endotyping asthma related to 3 different work exposures. J. Allergy Clin. Immunol. 2021, 148, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Kalbe, B.; Knobloch, J.; Schulz, V.M.; Wecker, C.; Schlimm, M.; Scholz, P.; Jansen, F.; Stoelben, E.; Philippou, S.; Hecker, E.; et al. Olfactory Receptors Modulate Physiological Processes in Human Airway Smooth Muscle Cells. Front. Physiol. 2016, 7, 339. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-T.; Wu, K.-J. Epigenetic regulation of epithelial-mesenchymal transition: Focusing on hypoxia and TGF-β signaling. J. Biomed. Sci. 2020, 27, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, D.; Nico, J.; Johnston, A.D.; Tobias, T.A.M.; Jorge, Y.; Macian, F.; Greally, J.M. CDC42-related genes are upregulated in helper T cells from obese asthmatic children. J. Allergy Clin. Immunol. 2018, 141, 539–548.e7. [Google Scholar] [CrossRef]
- Mahn, K.; Hirst, S.J.; Ying, S.; Holt, M.R.; Lavender, P.; Ojo, O.O.; Siew, L.; Simcock, D.E.; McVicker, C.G.; Kanabar, V.; et al. Diminished sarco/endoplasmic reticulum Ca 2+ ATPase (SERCA) expression contributes to airway remodelling in bronchial asthma. Proc. Natl. Acad. Sci. USA 2009, 106, 10775–10780. [Google Scholar] [CrossRef]
- Hoang, T.T.; Sikdar, S.; Xu, C.-J.; Lee, M.K.; Cardwell, J.; Forno, E.; Imboden, M.; Jeong, A.; Madore, A.-M.; Qi, C.; et al. Epigenome-wide association study of DNA methylation and adult asthma in the Agricultural Lung Health Study. Eur. Respir. J. 2020, 56, 2000217. [Google Scholar] [CrossRef]
- Ingram, J.L.; Kraft, M. Metalloproteinases as modulators of allergic asthma: Therapeutic perspectives. Met. Med. 2015, 2, 61–74. [Google Scholar] [CrossRef]
- Katula, K.S.; Heinloth, A.N.; Paules, R.S. Folate deficiency in normal human fibroblasts leads to altered expression of genes primarily linked to cell signaling, the cytoskeleton and extracellular matrix. J. Nutr. Biochem. 2007, 18, 541–552. [Google Scholar] [CrossRef]
- Mercer, B.; Lemaitre, V.; Powell, C.; D’Armiento, J. The Epithelial Cell in Lung Health and Emphysema Pathogenesis. Curr. Respir. Med. Rev. 2006, 2, 101–142. [Google Scholar] [CrossRef]
- Dijk, F.N.; Xu, C.; Melén, E.; Carsin, A.-E.; Kumar, A.; Nolte, I.M.; Gruzieva, O.; Pershagen, G.; Grotenboer, N.S.; Savenije, O.E.; et al. Genetic regulation of IL1RL1 methylation and IL1RL1-a protein levels in asthma. Eur. Respir. J. 2018, 51, 1701377. [Google Scholar] [CrossRef]
- Hansmann, L.; Schmidl, C.; Kett, J.; Steger, L.; Andreesen, R.; Hoffmann, P.; Rehli, M.; Edinger, M. Dominant Th2 Differentiation of Human Regulatory T Cells upon Loss of FOXP3 Expression. J. Immunol. 2012, 188, 1275–1282. [Google Scholar] [CrossRef]
- McHugh, B.J.; Murdoch, A.; Haslett, C.; Sethi, T. Loss of the Integrin-Activating Transmembrane Protein Fam38A (Piezo1) Promotes a Switch to a Reduced Integrin-Dependent Mode of Cell Migration. PLoS ONE 2012, 7, e40346. [Google Scholar] [CrossRef]
- Kho, A.T.; Sharma, S.; Davis, J.S.; Spina, J.; Howard, D.; McEnroy, K.; Moore, K.; Sylvia, J.; Qiu, W.; Weiss, S.T.; et al. Circulating MicroRNAs: Association with Lung Function in Asthma. PLoS ONE 2016, 11, e0157998. [Google Scholar] [CrossRef] [Green Version]
- Li, M.J.; Wang, P.; Liu, X.; Lim, E.L.; Wang, Z.; Yeager, M.; Wong, M.P.; Sham, P.C.; Chanock, S.J.; Wang, J. GWASdb: A database for human genetic variants identified by genome-wide association studies. Nucleic Acids Res. 2012, 40, D1047–D1054. [Google Scholar] [CrossRef]
- Chen, Y.-G. Endocytic regulation of TGF-β signaling. Cell Res. 2009, 19, 58–70. [Google Scholar] [CrossRef]
- Satish, L.; O’Gorman, D.B.; Johnson, S.; Raykha, C.; Gan, B.S.; Wang, J.H.-C.; Kathju, S. Increased CCT-eta expression is a marker of latent and active disease and a modulator of fibroblast contractility in Dupuytren’s contracture. Cell Stress Chaperones 2013, 18, 397–404. [Google Scholar] [CrossRef]
- Génier, S.; Degrandmaison, J.; Moreau, P.; Labrecque, P.; Hébert, T.E.; Parent, J.-L. Regulation of GPCR expression through an interaction with CCT7, a subunit of the CCT/TRiC complex. Mol. Biol. Cell 2016, 27, 3800–3812. [Google Scholar] [CrossRef]
- Pascual, M.; Roa, S.; García-Sánchez, A.; Sanz, C.; Hernandez-Hernandez, L.; Greally, J.M.; Lorente, F.; Dávila, I.; Isidoro-García, M. Genome-wide expression profiling of B lymphocytes reveals IL4R increase in allergic asthma. J. Allergy Clin. Immunol. 2014, 134, 972–975. [Google Scholar] [CrossRef]
- Zong, M.; Lu, T.; Fan, S.; Zhang, H.; Gong, R.; Sun, L.; Fu, Z.; Fan, L. Glucose-6-phosphate isomerase promotes the proliferation and inhibits the apoptosis in fibroblast-like synoviocytes in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 100. [Google Scholar] [CrossRef]
- Mousavizadeh, R.; Hojabrpour, P.; Eltit, F.; McDonald, P.C.; Dedhar, S.; McCormack, R.G.; Duronio, V.; Jafarnejad, S.M.; Scott, A. β1 integrin, ILK and mTOR regulate collagen synthesis in mechanically loaded tendon cells. Sci. Rep. 2020, 10, 12644. [Google Scholar] [CrossRef]
- Liu, Y.; Huo, S.-G.; Xu, L.; Che, Y.-Y.; Jiang, S.-Y.; Zhu, L.; Zhao, M.; Teng, Y.-C. MiR-135b Alleviates Airway Inflammation in Asthmatic Children and Experimental Mice with Asthma via Regulating CXCL12. Immunol. Investig. 2022, 51, 496–510. [Google Scholar] [CrossRef]
- Huang, X.-P.; Qin, C.-Y.; Gao, Y.-M. miR-135a inhibits airway inflammatory response in asthmatic mice via regulating JAK/STAT signaling pathway. Braz. J. Med. Biol. Res. 2021, 54, e10023. [Google Scholar] [CrossRef]
- Roffel, M.; Boudewijn, I.; Guryev, V.; Brandsma, C.-A.; van den Berge, M.; Bracke, K.; Maes, T.; Heijink, I. Unraveling the role of miR-223-3p in the regulation of airway inflammation in asthma. Eur. Res. J. 2018, 52, PA4998. [Google Scholar] [CrossRef]
- Iwata, J.-I.; Tung, L.; Urata, M.; Hacia, J.G.; Pelikan, R.; Suzuki, A.; Ramenzoni, L.; Chaudhry, O.; Parada, C.; Sanchez-Lara, P.A.; et al. Fibroblast Growth Factor 9 (FGF9)-Pituitary Homeobox 2 (PITX2) Pathway Mediates Transforming Growth Factor β (TGFβ) Signaling to Regulate Cell Proliferation in Palatal Mesenchyme during Mouse Palatogenesis. J. Biol. Chem. 2012, 287, 2353–2363. [Google Scholar] [CrossRef]
- Shang, Y.; Doan, C.N.; Arnold, T.D.; Lee, S.; Tang, A.A.; Reichardt, L.F.; Huang, E.J. Transcriptional Corepressors HIPK1 and HIPK2 Control Angiogenesis Via TGF-β–TAK1–Dependent Mechanism. PLoS Biol. 2013, 11, e1001527. [Google Scholar] [CrossRef] [PubMed]
- Hurley, D.G.; Araki, H.; Tamada, Y.; Dunmore, B.; Sanders, D.; Humphreys, S.; Affara, M.; Imoto, S.; Yasuda, K.; Tomiyasu, Y.; et al. Gene network inference and visualization tools for biologists: Application to new human transcriptome datasets. Nucleic Acids Res. 2012, 40, 2377–2398. [Google Scholar] [CrossRef]
- Zhang, Y.; Lv, J.; Liu, H.; Zhu, J.; Su, J.; Wu, Q.; Qi, Y.; Wang, F.; Li, X. HHMD: The human histone modification database. Nucleic Acids Res. 2010, 38, D149–D154. [Google Scholar] [CrossRef]
- Matsumoto, H. Roles of Periostin in Asthma. Adv. Exp. Med. Biol. 2019, 1132, 145–159. [Google Scholar] [CrossRef]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 2005, 201, 233–240. [Google Scholar] [CrossRef]
- Pavlidis, S.; Takahashi, K.; Ng Kee Kwong, F.; Xie, J.; Hoda, U.; Sun, K.; Elyasigomari, V.; Agapow, P.; Loza, M.; Baribaud, F.; et al. “T2-high” in severe asthma related to blood eosinophil, exhaled nitric oxide and serum periostin. Eur. Respir. J. 2019, 53, 1800938. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, M.; Leimgruber, E.; Seguín-Estévez, Q.; Dunand-Sauthier, I.; Barras, E.; Reith, W. Expression of RAB4B, a protein governing endocytic recycling, is co-regulated with MHC class II genes. Nucleic Acids Res. 2006, 35, 595–605. [Google Scholar] [CrossRef]
- Do, A.N.; Chun, Y.; Grishina, G.; Grishin, A.; Rogers, A.J.; Raby, B.A.; Weiss, S.T.; Vicencio, A.; Schadt, E.E.; Bunyavanich, S. Network study of nasal transcriptome profiles reveals master regulator genes of asthma. J. Allergy Clin. Immunol. 2021, 147, 879–893. [Google Scholar] [CrossRef] [PubMed]
- Neznanov, N.; Neznanova, L.; Angres, B.; Gudkov, A.V. Serologically Defined Colon Cancer Antigen 3 Is Necessary for the Presentation of TNF Receptor 1 on Cell Surface. DNA Cell Biol. 2005, 24, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Howarth, P.H.; Babu, K.S.; Arshad, H.S.; Lau, L.; Buckley, M.; McConnell, W.; Beckett, P.; Al Ali, M.; Chauhan, A.; Wilson, S.J.; et al. Tumour necrosis factor (TNF) as a novel therapeutic target in symptomatic corticosteroid dependent asthma. Thorax 2005, 60, 1012–1018. [Google Scholar] [CrossRef]
- Tränkner, D.; Hahne, N.; Sugino, K.; Hoon, M.A.; Zuker, C. Population of sensory neurons essential for asthmatic hyperreactivity of inflamed airways. Proc. Natl. Acad. Sci. USA 2014, 111, 11515–11520. [Google Scholar] [CrossRef]
- Oguchi, M.E.; Noguchi, K.; Fukuda, M. TBC1D12 is a novel Rab11-binding protein that modulates neurite outgrowth of PC12 cells. PLoS ONE 2017, 12, e0174883. [Google Scholar] [CrossRef] [PubMed]
- Kurundkar, A.R.; Kurundkar, D.; Rangarajan, S.; Locy, M.L.; Zhou, Y.; Liu, R.; Zmijewski, J.; Thannickal, V.J. The matricellular protein CCN1 enhances TGF-β1/SMAD3-dependent profibrotic signaling in fibroblasts and contributes to fibrogenic responses to lung injury. FASEB J. 2016, 30, 2135–2150. [Google Scholar] [CrossRef] [PubMed]
- Zuyderduyn, S.; Sukkar, M.; Fust, A.; Dhaliwal, S.; Burgess, J. Treating asthma means treating airway smooth muscle cells. Eur. Respir. J. 2008, 32, 265–274. [Google Scholar] [CrossRef]
- Oshimori, N.; Fuchs, E. Paracrine TGF-β Signaling Counterbalances BMP-Mediated Repression in Hair Follicle Stem Cell Activation. Cell Stem Cell 2012, 10, 63–75. [Google Scholar] [CrossRef]
- Formentini, L.; Sánchez-Aragó, M.; Sánchez-Cenizo, L.; Cuezva, J.M. The Mitochondrial ATPase Inhibitory Factor 1 Triggers a ROS-Mediated Retrograde Prosurvival and Proliferative Response. Mol. Cell 2012, 45, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Bazan-Socha, S.; Zuk, J.; Plutecka, H.; Marcinkiewicz, C.; Zareba, L.; Musial, J. Collagen Receptors α(1)β(1) and α(2)β(1) Integrins Are Involved in Transmigration of Peripheral Blood Eosinophils, but Not Mononuclear Cells through Human Microvascular Endothelial Cells Monolayer. J. Physiol. Pharmacol. 2012, 63, 373–379. [Google Scholar]
- Seo, G.; Han, H.; Vargas, R.E.; Yang, B.; Li, X.; Wang, W. MAP4K Interactome Reveals STRN4 as a Key STRIPAK Complex Component in Hippo Pathway Regulation. Cell Rep. 2020, 32, 107860. [Google Scholar] [CrossRef]
- Chuang, H.-C.; Sheu, W.H.-H.; Lin, Y.-T.; Tsai, C.-Y.; Yang, C.-Y.; Cheng, Y.-J.; Huang, P.-Y.; Li, J.-P.; Chiu, L.-L.; Wang, X.; et al. HGK/MAP4K4 deficiency induces TRAF2 stabilization and Th17 differentiation leading to insulin resistance. Nat. Commun. 2014, 5, 4602. [Google Scholar] [CrossRef] [PubMed]
- Zein, J.G.; McManus, J.M.; Sharifi, N.; Erzurum, S.C.; Marozkina, N.; Lahm, T.; Giddings, O.; Davis, M.D.; DeBoer, M.D.; Comhair, S.A.; et al. Benefits of Airway Androgen Receptor Expression in Human Asthma. Am. J. Respir. Crit. Care Med. 2021, 204, 285–293. [Google Scholar] [CrossRef]
- Castro-Giner, F.; Bustamante, M.; González, J.R.; Kogevinas, M.; Jarvis, D.; Heinrich, J.; Antó, J.-M.; Wjst, M.; Estivill, X.; de Cid, R. A pooling-based genome-wide analysis identifies new potential candidate genes for atopy in the European Community Respiratory Health Survey (ECRHS). BMC Med. Genet. 2009, 10, 128. [Google Scholar] [CrossRef]
- Wnuk, D.; Paw, M.; Ryczek, K.; Bochenek, G.; Sładek, K.; Madeja, Z.; Michalik, M. Enhanced asthma-related fibroblast to myofibroblast transition is the result of profibrotic TGF-β/Smad2/3 pathway intensification and antifibrotic TGF-β/Smad1/5/(8)9 pathway impairment. Sci. Rep. 2020, 10, 16492. [Google Scholar] [CrossRef]
- Jia, X.-X.; Zhu, T.-T.; Huang, Y.; Zeng, X.-X.; Zhang, H.; Zhang, W.-X. Wnt/β-catenin signaling pathway regulates asthma airway remodeling by influencing the expression of c-Myc and cyclin D1 via the p38 MAPK-dependent pathway. Exp. Ther. Med. 2019, 18, 3431–3438. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, P.; Etcheverry, A.; Aubry, M.; Missey, A.; Lachat, C.; Perrard, J.; Hendrick, E.; Delage-Mourroux, R.; Mosser, J.; Borg, C.; et al. EMT is associated with an epigenetic signature of ECM remodeling genes. Cell Death Dis. 2019, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Reddel, H.K.; Bacharier, L.B.; Bateman, E.D.; Brightling, C.E.; Brusselle, G.G.; Buhl, R.; Cruz, A.A.; Duijts, L.; Drazen, J.M.; FitzGerald, J.M.; et al. Global Initiative for Asthma Strategy 2021: Executive summary and rationale for key changes. Eur. Respir. J. 2022, 59, 2102730. [Google Scholar] [CrossRef]
- Kuo, C.-H.S.; Pavlidis, S.; Loza, M.; Baribaud, F.; Rowe, A.; Pandis, I.; Sousa, A.; Corfield, J.; Djukanovic, R.; Lutter, R.; et al. T-helper cell type 2 (Th2) and non-Th2 molecular phenotypes of asthma using sputum transcriptomics in U-BIOPRED. Eur. Respir. J. 2017, 49, 1602135. [Google Scholar] [CrossRef]
- Sokolowski, J.W.; Burgher, L.W.; Jones, F.L.; Patterson, J.R.; Selecky, P.A. Position Paper on Guidelines for Fiberoptic Bronchoscopy in Adults. Am. Rev. Respir. Dis. 1987, 136, 1066. [Google Scholar] [CrossRef]
- Bazan-Socha, S.; Jakiela, B.; Zuk, J.; Zarychta, J.; Soja, J.; Okon, K.; Dziedzina, S.; Zareba, L.; Dropinski, J.; Wojcik, K.; et al. Interactions via α2β1 Cell Integrin May Protect against the Progression of Airway Structural Changes in Asthma. Int. J. Mol. Sci. 2021, 22, 6315. [Google Scholar] [CrossRef]
- Ferrando, R.E.; Nyengaard, J.R.; Hays, S.R.; Fahy, J.V.; Woodruff, P.G. Applying stereology to measure thickness of the basement membrane zone in bronchial biopsy specimens. J. Allergy Clin. Immunol. 2003, 112, 1243–1245. [Google Scholar] [CrossRef]
- Rueda, L. Microarray Image and Data Analysis; Rueda, L., Ed.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781466586871. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Hu, J.; Zou, F.; Wright, F.A. Practical FDR-based sample size calculations in microarray experiments. Bioinformatics 2005, 21, 3264–3272. [Google Scholar] [CrossRef] [Green Version]
- Farahbod, M.; Pavlidis, P. Differential coexpression in human tissues and the confounding effect of mean expression levels. Bioinformatics 2019, 35, 55–61. [Google Scholar] [CrossRef]
- Watson, M. CoXpress: Differential co-expression in gene expression data. BMC Bioinform. 2006, 7, 509. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample’s Clinical Features | EA | NEA | Difference p-Value * |
|---|---|---|---|
| Clinical features, confounding diseases | |||
| Sample size | n = 20 | n = 20 | |
| Gender, n (%) female | 13 (65%) | 16 (80%) | 0.240 |
| Age, years ** | 58 (16) | 52 (23) | 0.096 |
| BMI, kg/m2 ** | 25.4 (3.1) | 30.4 (11.4) | 0.020 |
| Atopy, n (%) | 11 (58%) | 9 (53%) | 0.515 |
| Allergic rhinitis, n (%) | 15 (88%) | 17 (89%) | 0.655 |
| Gastroesophageal reflux disease, n (%) | 8 (44%) | 10 (58%) | 0.305 |
| Nicotinism, past, n (%) | 13 (65%) | 16 (80%) | 0.563 |
| Asthma severity | |||
| ACT, points ** | 14 (8) | 16 (9) | 0.227 |
| Severe asthma (GINA class 4), n (%) | 9 (45%) | 7 (35%) | 0.519 |
| Asthma phenotype by different criteria | |||
| BAL eosinophilia ≥ 1% | 20 (100%) | none | n/a |
| BAL eosinophilia ≥ 2.5% | 11 (55%) | none | n/a |
| BAL eosinophilia > 3% | 8 (40%) | none | n/a |
| Blood eosinophilia ≥ 350/mcL | 11 (55%) | 4 (20%) | 0.008 |
| BAL neutrophilia > 40% | 1 (5%) | none | n/a |
| BAL neutrophilia ≤ 40% and BAL eosinophilia < 1% | none | 20 (100%) | n/a |
| Medications used | |||
| Oral steroid, n (%) | 5 (25%) | 2 (10%) | 0.237 |
| Inhaled steroid, n (%) | 15 (75%) | 16 (80%) | 0.962 |
| LABA, n (%) | 16 (80%) | 15 (75%) | 0.412 |
| LAMA, n (%) | 1 (5%) | 4 (20%) | 0.135 |
| Montelukast, n (%) | 2 (10%) | 3 (15%) | 0.677 |
| Theophylline, n (%) | 3 (15%) | 2 (10%) | 0.630 |
| Acetylsalicylic acid/thienopyridines, n (%) | 4 (20%) | 8 (40%) | 0.630 |
| ACEI/ARB, n (%) | 2 (10%) | 6 (30%) | 0.197 |
| Beta-blockers, n (%) | 3 (15%) | 5 (25%) | 0.110 |
| Calcium channel blockers, n (%) | 4 (20%) | 7 (35%) | 0.331 |
| Diuretics, n (%) | 4 (20%) | 7 (35%) | 0.331 |
| Statins, n (%) | 11 (55%) | 9 (45%) | 0.278 |
| Proton pump inhibitors, n (%) | 1 (5%) | 4 (20%) | 0.402 |
| Proxies of Airway Remodeling | EA | NEA | Difference p-Value * |
|---|---|---|---|
| Spirometry (with reversibility test) and body plethysmography | |||
| FEV1, L | 2.1 (0.79) | 2.28 (1.26) | 0.196 |
| FEV1, % predicted | 77.4 (40.2) | 89.7 (31.1) | 0.421 |
| VC, L | 3.33 (1.78) | 3.36 (1.11) | 0.292 |
| FEV1/VC | 62.86 (16.86) | 67.8 (17.1) | 0.267 |
| ∆FEV1, L | 0.21 (0.23) | 0.17 (0.22) | 0.506 |
| ∆FEV1, % | 0.118 (0.143) | 0.07 (0.13) | 0.419 |
| TLC, L | 6.65 (2.13) | 5.63 (1.51) | 0.419 |
| TLC, % predicted | 114.25 (18.5) | 111.9 (22.5) | 0.295 |
| RV% TLC | 30.63 (3.68) | 30.45 (11.48) | 0.913 |
| High-resolution computed tomography (at RB1 bronchus) | |||
| Wall thickness (mm, average) | 1.216 (0.32) | 1.078 (0.22) | 0.033 |
| Lumen diameter (mm, average) | 2.719 (0.84) | 2.344 (0.47) | 0.077 |
| Airway diameter (mm, average) | 5.238 (1.58) | 4.593 (0.68) | 0.044 |
| Wall thickness ratio | 14.692 (2.2) | 14.817 (1.94) | 0.879 |
| Wall area (mm2) | 25.365 (12.81) | 18.708 (6.56) | 0.033 |
| Wall area ratio | 44.24 (4.99) | 45.290 (4.98) | 0.676 |
| Serum and BAL Studies | EA Group | NEA Group | Difference p-Value * |
|---|---|---|---|
| Serum cytokines, pg/mL (mean ± SD) | |||
| IFN-γ | 0.575 ± 1.406 | 0.405 ± 1.355 | 0.490 |
| IL- | 0.005 ± 0 | 0.005 ± 0 | n/a |
| IL-5 | 0.005 ± 0 | 0.946 ± 4.210 | 0.797 |
| IL-6 | 2.058 ± 2.059 | 1.366 ± 1.508 | 0.387 |
| IL-10 | 1.260 ± 1.074 | 0.602 ± 0.807 | 0.017 |
| IL-12p70 | 1.941 ± 3.777 | 1.255 ± 2.640 | 0.946 |
| IL-17A | 0.188 ± 0.579 | 0.171 ± 0.450 | 0.860 |
| IL-23 | 15.81 ± 25.11 | 48.53 ± 98.36 | 0.323 |
| Periostin | 1.284 ± 3.038 | 0.329 ± 0.086 | 0.340 |
| ADAM33 (ng/mL) | 2.229 ± 1.612 | 1.462 ± 0.944 | 0.126 |
| IgE (IU/mL) | 161 (582) | 32 (124) | 0.018 |
| BAL cytokines, pg/mL (mean ± SD) | |||
| IFN-γ | 0.005 ± 0 | 0.005 ± 0 | n/a |
| IL-4 | 0.645 ± 1.832 | 0.379 ± 1.276 | 0.931 |
| IL-5 | 0.342 ± 1.509 | 0.005 ± 0 | 0.342 |
| IL-6 | 0.586 ± 0.583 | 0.832 ± 0.624 | 0.197 |
| IL-10 | 0.005 ± 0 | 0.005 ± 0 | n/a |
| IL-12p70 | 0.068 ± 0.021 | 0.063 ± 0.033 | 0.685 |
| IL-17A | 0.005 ± 0 | 0.005 ± 0 | n/a |
| IL-23 | 0.245 ± 1.076 | 0.005 ± 0 | 0.342 |
| Periostin | 0.741 ± 0.210 | 0.911 ± 0.143 | 0.011 |
| Blood cell count, cells/µL (mean ± SD) | |||
| Neutrophils | 3580 (2100) | 3480 (1260) | 0.370 |
| Lymphocytes | 2240 (1110) | 1930 (730) | 0.132 |
| Eosinophils | 480 (880) | 140 (260) | 0.008 |
| Basophils | 40 (30) | 20 (20) | 0.012 |
| Monocytes | 700 (370) | 505 (110) | 0.020 |
| BAL cell %, median (range) | |||
| Neutrophiles | 4.0 (0.0–85.5) | 3.0 (0.0–17.0) | 0.211 |
| Lymphocytes | 8.0 (0.1–67.0) | 4.0 (1.0–41.0) | 0.052 |
| Eosinophils | 2.3 (1.0–42.0) | 0.0 (0.0–0.5) | <0.001 |
| Macrophages | 80 (0.0–92.0) | 92.8 (53.5–98.5) | <0.001 |
| Cellularity, per ml | 13,920 (253–42,250) | 15,019 (6020–42,750) | 0.157 |
| Gene Name | Encoded Protein | log2FC | Adjusted p-Value | Gene Function and Relevance in Asthma |
|---|---|---|---|---|
| Genes up-regulated in eosinophilic asthma | ||||
| CPLX2 | Complexin 2 | 0.077 | 0.00016 | Regulates the formation of synaptic vesicles; likely plays a role in immunoglobulin secretion from plasmocytes. |
| RTN4RL1 | Reticulon 4 Receptor Like 1 | 0.105 | 0.01339 | Surface receptor found on vagal sensory neurons, likely involved in bronchial hyperreactivity. |
| KRTAP10-1 | Keratin Associated Protein 10-1 | 0.070 | 0.01105 | Forms a matrix of keratin intermediate filaments; also involved in nervous system development. |
| KIR3DS1 | Killer Cell Immunoglobulin-Like Receptor, Three Ig Domains And Short Cytoplasmic Tail 1 | 0.088 | 0.01721 | Natural killer cell immunoglobulin-like receptor for MHC class I molecules transducing activating signals; triggers degranulation and antiviral cytokine secretion upon interaction. |
| AL137028.1 | IQ Motif And Sec7 Domain 3 Pseudogene 3 | 0.110 | 0.00348 | A pseudogene of undetermined function. |
| FBLN1 | Fibulin 1 | 0.075 | 0.01134 | A structural protein stabilizing ECM proteins; regulates the biological availability of latent TGF-β [16]. |
| CDH23 | Cadherin Related 23 | 0.120 | 0.01721 | Member of cadherin superfamily playing a role in cell–cell adhesion. |
| SOX14 | SRY-Box Transcription Factor 14 | 0.055 | 0.01028 | Transcription factor involved in angiogenesis, tissue injury, and wound healing. |
| TMEFF1 | Transmembrane Protein With EGF Like And Two Follistatin Like Domains 1 | 0.081 | 0.00895 | Found to be expressed upon stimulation with TGF-β as direct target gene of TGF-β2/Smad2/3; involved in airway remodeling. |
| NDUFAF8 | NADH:Ubiquinone Oxidoreductase Complex Assembly Factor 8 | 0.075 | 0.00895 | Involved in the assembly of mitochondrial NADH: ubiquinone oxidoreductase complex participating in reactive oxygen species production. |
| ENTR1 | Endosome Associated Trafficking Regulator 1 | 0.106 | 0.0008 | An endosomal protein that functions in actin cytoskeleton remodeling, protein trafficking, and secretion, cytokinesis, and apoptosis. |
| CNN1 | Calponin 1 | 0.072 | 0.01928 | Filament-associated protein implicated in the regulation of smooth muscle contraction. |
| GSN | Gelsolin | 0.154 | 0.00096 | Implicated in TGF-β-dependent smooth muscle actin synthesis in myofibroblasts and epithelial–mesenchymal transition [17]. |
| FMNL1 | Formin Like 1 | 0.065 | 0.00703 | Plays a role in regulation of cell morphology and cytoskeletal organization, modifies macrophage motility. |
| SUN5 | Sad1 and UNC84 Domain-Containing 5 | 0.062 | 0.01570 | SUN family builds a bridge between the nucleoskeleton and the cytoskeleton, mediating nuclear dynamics during cell division. |
| GYS2 | Glycogen Synthase 2 | 0.085 | 0.01267 | Up-regulated upon signaling through PPARα, a major regulator of airway inflammation. |
| SLC22A18AS | Solute Carrier Family 22 Member 18 Antisense | 0.102 | 0.00895 | Antisense partner of an SLC22A18 gene; its up-regulation may suppress LC22A18, an important tumor-suppressor gene, increasing cell proliferation. |
| ANK3 | Ankyrin G | 0.092 | 0.00895 | The integral membrane protein that plays a role in cell motility, activation, proliferation, and contact. |
| ATPIF1 | ATP Synthase Inhibitory Factor Subunit 1 | 0.071 | 0.01339 | Its overexpression triggers reactive oxygen species production and promotes transcriptional activation of NFκB, resulting in a proliferative response involved in tissue remodeling. |
| RAB4B | Ras-Related GTP-Binding Protein 4b | 0.090 | 0.00096 | Plays a role in the regulation of vesicular trafficking. |
| Genes down-regulated in eosinophilic asthma | ||||
| DNAJA1 | DnaJ Heat Shock Protein Family (Hsp40) Member A1 | −0.075 | 0.01339 | Down-regulation of DNAJA1 was indicated as one of the biomarkers of isocyanate occupational asthma [18]. |
| GJB1 | Gap Junction Protein Beta 1 | −0.040 | 0.03043 | A gap junction channel protein; its absence inhibits TNF-α-induced extrinsic apoptosis pathway. |
| TBC1D12 | TBC1 Domain Family Member 12 | −0.088 | 0.00096 | Negatively regulates neurite outgrowth; down-regulation in eosinophilic asthma may result in increased neurite sprouting in the airways, a feature of airway remodeling in asthma. |
| GDPD1 | Glycerophosphodiester Phosphodiesterase Domain Containing 1 | −0.064 | 0.00703 | One of the proteins involved in glucocorticoid signaling pathways (PAHS-154Z); down-regulation may be associated with decreased sensitivity to glucocorticoids or prolonged glucocorticoid therapy. |
| OR10H5 | Olfactory Receptor Family 10 Subfamily H Member 5 | −0.067 | 0.01801 | An olfactory receptor regulating cytoskeletal remodeling and ASM cell proliferation [19]. |
| DMRTC2 | DMRT Like Family C2 | −0.049 | 0.01301 | A transcription factor involved in H3K9 methylation pattern and TGF-β epithelial–mesenchymal transition in airway remodeling [20]. |
| SIN3A | SIN3 Transcription Regulator Family Member A | −0.057 | 0.01268 | A transcriptional repressor antagonizes the effects of MYC. Regulates the vast majority of the transcriptional response to hypoxia. |
| HYMAI | Hydatidiform Mole Associated And Imprinted | −0.055 | 0.01301 | A non-protein coding gene of unknown function in airway pathology. |
| UGT2A3 | UDP Glucuronosyltransferase Family 2 Member A3 | −0.057 | 0.03012 | One of the genes implicated in the metabolism of xenobiotics, including nicotine and tobacco carcinogens. |
| CREB5 | CAMP Responsive Element Binding Protein 5 | −0.038 | 0.01350 | Recently identified as a common regulon in asthma exacerbations, expressed in macrophages and dendritic cells. |
| ITM2B | Integral Membrane Protein 2B | −0.100 | 0.00895 | Associated with pediatric obesity-related asthma cases [21]. |
| CDC37P1 | Cell Division Cycle 37 Pseudogene 1 | −0.057 | 0.00895 | A pseudogene of undetermined function. |
| C3orf23 | T Cell Activation Inhibitor, Mitochondrial | −0.041 | 0.02436 | A mitochondrial protein implicated in apoptosis of T-cells. |
| Proxy of Airway Remodeling | Differentially Expressed Gene | Pearson’s Correlation Coefficient | Correlation p-Value |
|---|---|---|---|
| Spirometry (with reversibility test) and body plethysmography | |||
| FEV1, L | RAB4B | −0.64 | 0.001 |
| FMNL1 | −0.51 | 0.002 | |
| AL137028.1 | −0.51 | 0.002 | |
| GSN | −0.45 | 0.008 | |
| ANK3 | −0.40 | 0.018 | |
| SDCCAG3 | −0.39 | 0.024 | |
| NDUFAF8 | −0.36 | 0.036 | |
| KRTAP10.1 | −0.36 | 0.036 | |
| TBC1D12 | 0.41 | 0.017 | |
| FEV1/VC | GSN | −0.53 | 0.001 |
| AL137028.1 | −0.49 | 0.004 | |
| KIR3DS1 | −0.48 | 0.004 | |
| NDUFAF8 | −0.47 | 0.005 | |
| CDH23 | −0.44 | 0.010 | |
| KRTAP10.1 | −0.42 | 0.013 | |
| RTN4RL1 | −0.38 | 0.028 | |
| TBC1D12 | 0.41 | 0.017 | |
| TLC, % predicted | SIN3A | −0.41 | 0.032 |
| GSN | 0.42 | 0.028 | |
| SUN5 | 0.44 | 0.018 | |
| FMNL1 | 0.45 | 0.016 | |
| RV, %TLC | DMRTC2 | −0.41 | 0.033 |
| High-resolution computed tomography (at RB1 bronchus) | |||
| Wall thickness (mm, average) | ATPIF1 | 0.49 | 0.006 |
| Airway diameter (mm, average) | ATPIF1 | 0.48 | 0.007 |
| Wall area (mm2) | ATPIF1 | 0.51 | 0.004 |
| Wall area ratio | RAB4B | 0.42 | 0.023 |
| GSN | 0.39 | 0.034 | |
| NDUFAF8 | 0.38 | 0.041 | |
| Gene Name | Encoded Protein | log2FC | Gene Function and Relevance in Asthma |
|---|---|---|---|
| Genes up-regulated in eosinophilic asthma | |||
| ATP1B1 | Sodium/potassium-transporting ATPase subunit beta-1 | 0.534 | A ubiquitous protein regulating the location and function of sodium–potassium ATPase with possible involvement in epithelial sheathing, CMV and RSV infection, Th17 polarization, activation of inflammatory and airway smooth muscle cells, mucus secretion, and airway hyperreactivity [22]. |
| STRN4 | Striatin-4 | 0.481 | A regulatory subunit of the STRIPAK complex involved in cell cycle control, cell adhesion, migration, epithelial integrity, and epithelial–mesenchymal transition; associated with asthma in an EWAS study [23]. |
| GPI | Glucose-6-phosphate isomerase | 0.349 | An enzyme exhibiting the function of neurotrophic factor, a lymphokine inducing immunoglobulin secretion and an angiogenic factor, involved in secretion of TNF-α and IL-1b. |
| ANKRD26P1 | Ankyrin repeat domain 26 pseudogene 1 | 0.259 | A pseudogene of little-known function. |
| RAPH1 | Ras association (RalGDS/AF-6) and pleckstrin homology domains | 0.245 | An adaptor protein regulating actin dynamics. |
| RP3-473B4.3 | (lincRNA; not named yet) | 0.145 | No direct association with asthma, inflammation, or airway remodeling is known. |
| CLC | Charcot–Leyden crystal protein | 0.124 | An atypical galectin with an activity of lysophospholipase; taking part in vesicular transport of eosinophilic granule ribonucleases; ubiquitous in sputum of eosinophilic asthma patients. |
| RADX | RPA1 related single stranded DNA binding protein, X-linked | 0.107 | One of the genes up-regulated upon transition from pericytes into mesenchymal cells; may indicate an epithelial–mesenchymal transition in airway remodeling. |
| RECK | Reversion inducing cysteine rich protein with kazal motifs | 0.084 | A membrane-anchored protein negatively regulating a matrix metalloproteinase-9, a major metalloproteinase involved in asthma pathology [24]. |
| DGLUCY | D-glutamate cyclase | 0.076 | A mitochondrial enzyme known to up-regulate E-cadherin and repress signaling through ERK, possibly limiting epithelial–mesenchymal transition. |
| SLC19A1 | Solute carrier family 19 member 1 | 0.068 | Takes part in folate, homocysteine, nitric oxide, and reactive oxygen species metabolism and DNA methylation; possibly linked with asthma through effects of folate deficiency resulting in aggravated asthma symptoms, increased CD4/CD8 T-cell ratio, extracellular matrix remodeling, and signalling through β-catening/Wnt pathway implied in airway remodeling. [25] |
| TTC3P1 | Tetratricopeptide repeat domain 3 pseudogene 1 | 0.060 | As a pseudogene, it may interfere with TTC3, a gene for E3 ubiquitin-protein ligase. |
| RP11-321E2.2 | (lincRNA; not named yet) | 0.052 | No direct association with asthma, inflammation, or airway remodeling is known. |
| MAEA | macrophage erythroblast attacher | 0.049 | A ubiquitin ligase up-regulated upon exposure to cigarette smoke extract, RSV and MMP-9 [26]; an EWAS study found an association between trans-CpG site in MAEA and IL1RL1 [27]. |
| FBN3 | Fibrillin-3 | 0.035 | A component of extracellular calcium-binding myofibrils occurring with elastin- or elastin-free bundles important for ECM integrity; may be involved in the regulation of TGF-β signalling through association with latent TFG-β-binding proteins. |
| PSG2 | Pregnancy-specific beta-1-glycoprotein 2 | 0.031 | Stimulates transcription of FOXP3 in mononuclear and CD4+ T-cells providing a signal for T-reg and Th17 differentiation [28]. |
| PPP2R3B | Protein phosphatase 2 regulatory subunit B beta | 0.024 | A positive regulator of cell cycle progression. |
| GOLGA2P3Y | Golgin A2 Pseudogene 3 | 0.023 | No direct association with asthma, inflammation, or airway remodeling is known. |
| STOML3 | Stomatin like 3 | 0.025 | Plays a role in mechanotransduction through interaction with PIEZO1 with possible implications for airway hyperreactivity, cell adhesion, and migratory capacity of epithelial cells through inactivation of β1 integrin affinity [29]. |
| DIO3 | Deiodinase iodothyronine type III | 0.019 | An enzyme inactivating thyroid hormones; induced by TGF-β and oxidative stress. |
| FADS6 | Fatty acid desaturase 6 | 0.017 | A peroxisomal enzyme taking part in polyunsaturated fatty acids biosynthesis; contributes to arachidonic acid synthesis; regulated by miR-331-3p, a post-transcriptional regulator associated with lung function in asthma [30]. |
| EPS8L1 | EPS8 like 1 | 0.012 | A protein related to Eps8 involved in actin remodeling; plays a role in T-cell receptor binding, membrane ruffling, and remodeling of the actin cytoskeleton by CDC42 relevant for cell migration. |
| RPS13 | Ribosomal protein S13 | 0.013 | A housekeeping gene forming the TNF-α/NFκB signalling complex of established role in asthma. |
| ASB3 | Ankyrin Repeat And SOCS Box Containing 3 | 0.008 | Part of the ASB gene family involved in Erk1/2 and PI3K/Akt signal transduction pathways, implied in airway remodeling. |
| MRPL14 | Mitochondrial ribosomal protein L14 | 0.009 | Controlled by MYC transcription factor; related to asthma in a GWAS study [31]. |
| Genes down-regulated in eosinophilic asthma | |||
| EPS15 | Epidermal growth factor receptor pathway substrate 15 | −0.004 | A protein involved in internalization of tyrosine kinase receptors (including TGF-β), integrins (including integrin β1 and E-cadherin), and receptors relevant for bronchoconstriction (β2-adrenergic and M3-muscarinic) [32]. |
| RHOBTB2 | Rho related BTB domain containing 2 | −0.010 | Required for expression of CXCL14, a chemoattractant controlling dendritic cell activation, leukocyte migration, angiogenesis, and an autocrine growth factor for fibroblasts; differentially expressed after bronchial thermoplasty. |
| SRCRB4D | Scavenger Receptor Cysteine Rich Family Member With 4 Domains | −0.016 | Possibly associated with asthma as one of the target genes of BACH1 involved in response to oxidative stress and associated with asthma in an EWAS study [23]. |
| OR52I1 | Olfactory receptor family 52 subfamily I member 1 | −0.098 | An olfactory receptor possibly involved in airway hyperreactivity. |
| CCT7 | Chaperonin containing TCP1 subunit 7 | −0.141 | A molecular chaperon up-regulated in fibrotic wound healing and essential for accumulation of α-smooth muscle actin in fibroblasts and differentiation to myofibroblasts [33]; recently found to coimmunoprecipitate with thromboxane A2 receptor and β2-adrenergic receptor [34]; its depletion resulted in reduced expression of both receptors involved in asthma pathogenesis. |
| SRPRB | Signal recognition particle receptor beta subunit | −0.162 | Involved in intracellular trafficking of proteins. |
| CABP5 | Calcium-binding protein 5 | −0.602 | Reported in T-cells and differentially expressed in allergic asthma patients [35]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozlik-Siwiec, P.; Buregwa-Czuma, S.; Zawlik, I.; Dziedzina, S.; Myszka, A.; Zuk-Kuwik, J.; Siwiec-Kozlik, A.; Zarychta, J.; Okon, K.; Zareba, L.; et al. Co-Expression Analysis of Airway Epithelial Transcriptome in Asthma Patients with Eosinophilic vs. Non-Eosinophilic Airway Infiltration. Int. J. Mol. Sci. 2023, 24, 3789. https://doi.org/10.3390/ijms24043789
Kozlik-Siwiec P, Buregwa-Czuma S, Zawlik I, Dziedzina S, Myszka A, Zuk-Kuwik J, Siwiec-Kozlik A, Zarychta J, Okon K, Zareba L, et al. Co-Expression Analysis of Airway Epithelial Transcriptome in Asthma Patients with Eosinophilic vs. Non-Eosinophilic Airway Infiltration. International Journal of Molecular Sciences. 2023; 24(4):3789. https://doi.org/10.3390/ijms24043789
Chicago/Turabian StyleKozlik-Siwiec, Pawel, Sylwia Buregwa-Czuma, Izabela Zawlik, Sylwia Dziedzina, Aleksander Myszka, Joanna Zuk-Kuwik, Andzelika Siwiec-Kozlik, Jacek Zarychta, Krzysztof Okon, Lech Zareba, and et al. 2023. "Co-Expression Analysis of Airway Epithelial Transcriptome in Asthma Patients with Eosinophilic vs. Non-Eosinophilic Airway Infiltration" International Journal of Molecular Sciences 24, no. 4: 3789. https://doi.org/10.3390/ijms24043789
APA StyleKozlik-Siwiec, P., Buregwa-Czuma, S., Zawlik, I., Dziedzina, S., Myszka, A., Zuk-Kuwik, J., Siwiec-Kozlik, A., Zarychta, J., Okon, K., Zareba, L., Soja, J., Jakiela, B., Kepski, M., Bazan, J. G., & Bazan-Socha, S. (2023). Co-Expression Analysis of Airway Epithelial Transcriptome in Asthma Patients with Eosinophilic vs. Non-Eosinophilic Airway Infiltration. International Journal of Molecular Sciences, 24(4), 3789. https://doi.org/10.3390/ijms24043789