

4-Methylumbeliferone Treatment at a Dose of 1.2 g/kg/Day Is Safe for Long-Term Usage in Rats

 , , ,
, , ,  ,
,  and
and
Abstract
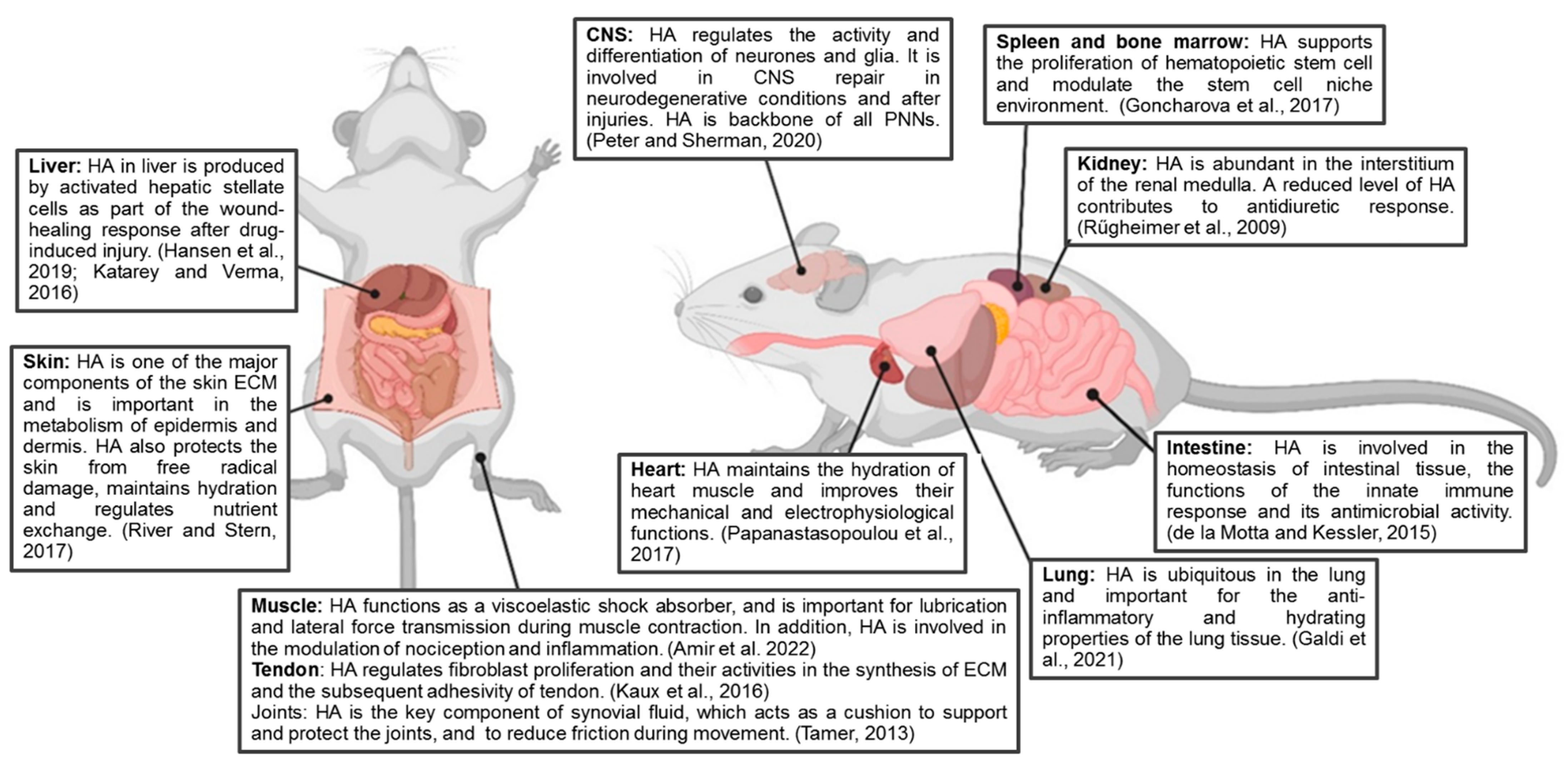
:1. Introduction
2. Results
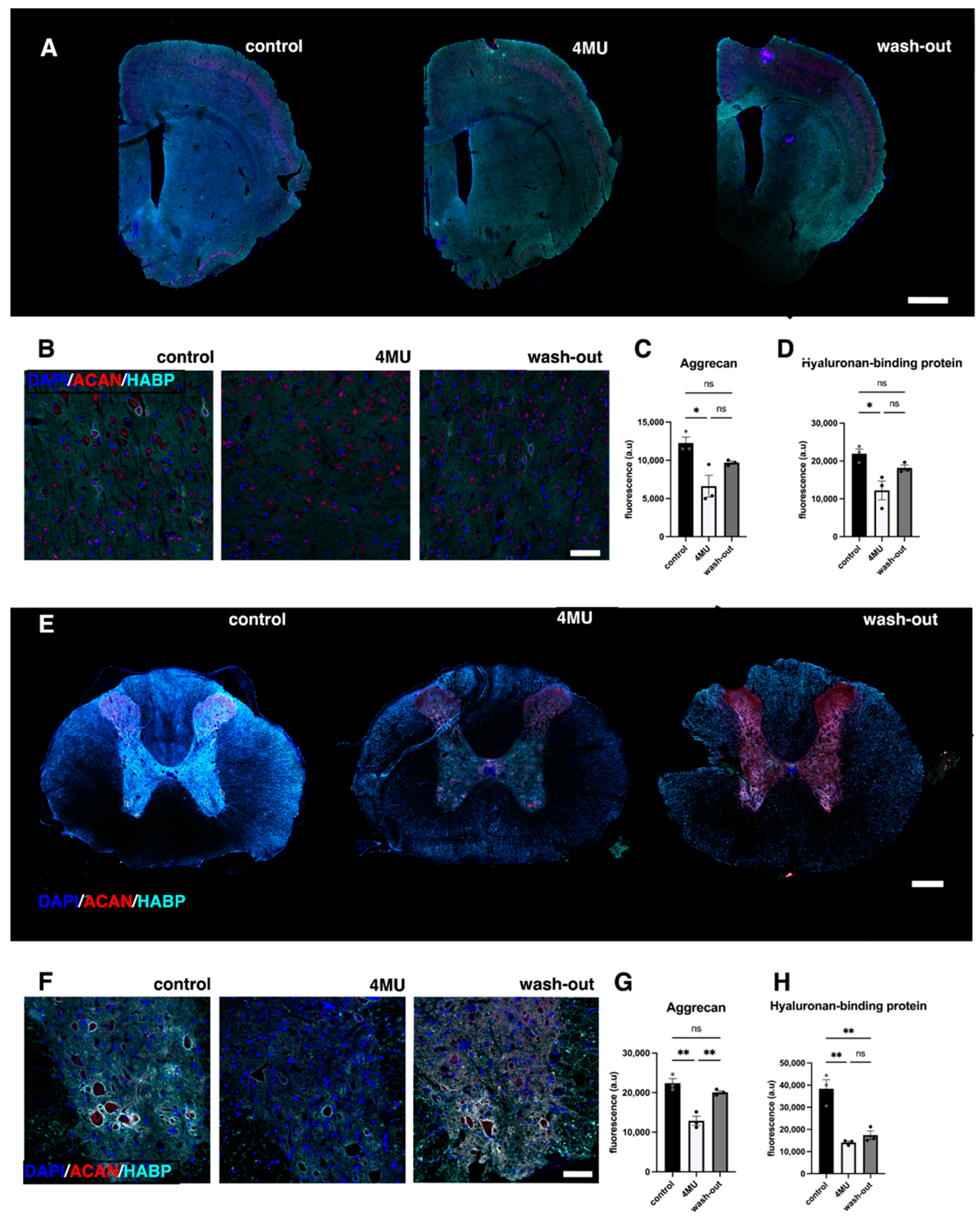
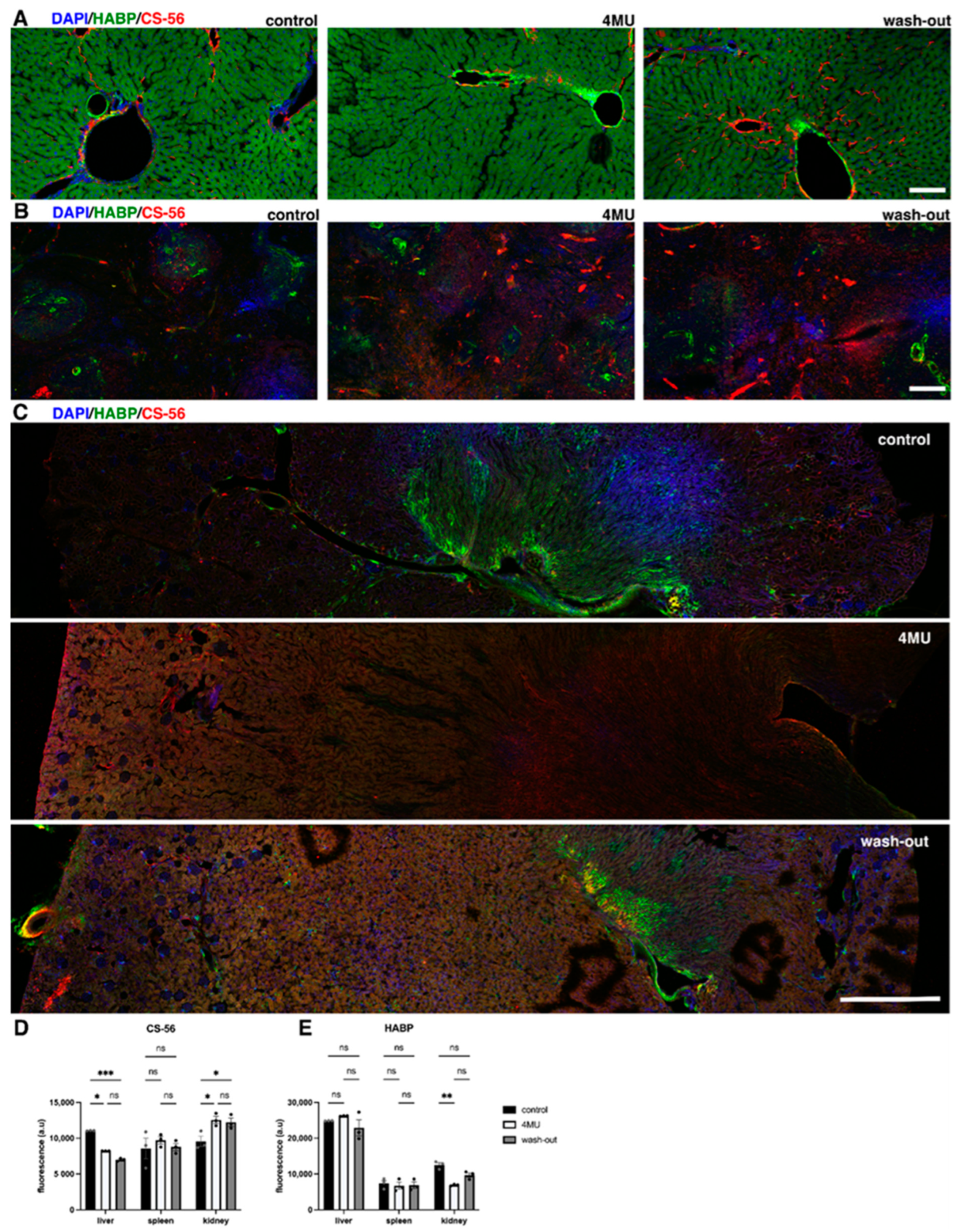
2.1. 1.2 g/kg/Day Dose of 4MU Downregulates HA throughout the Body
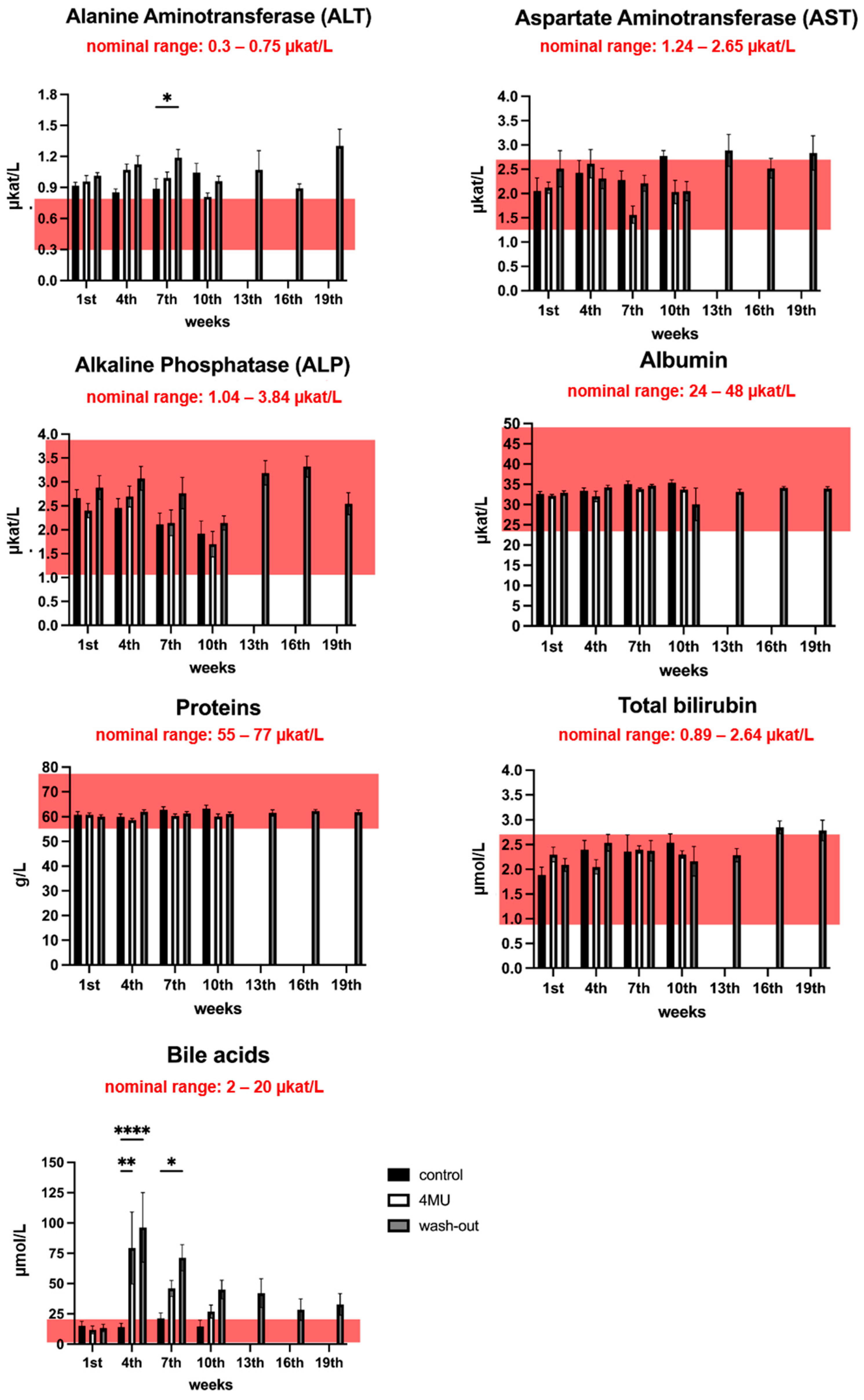
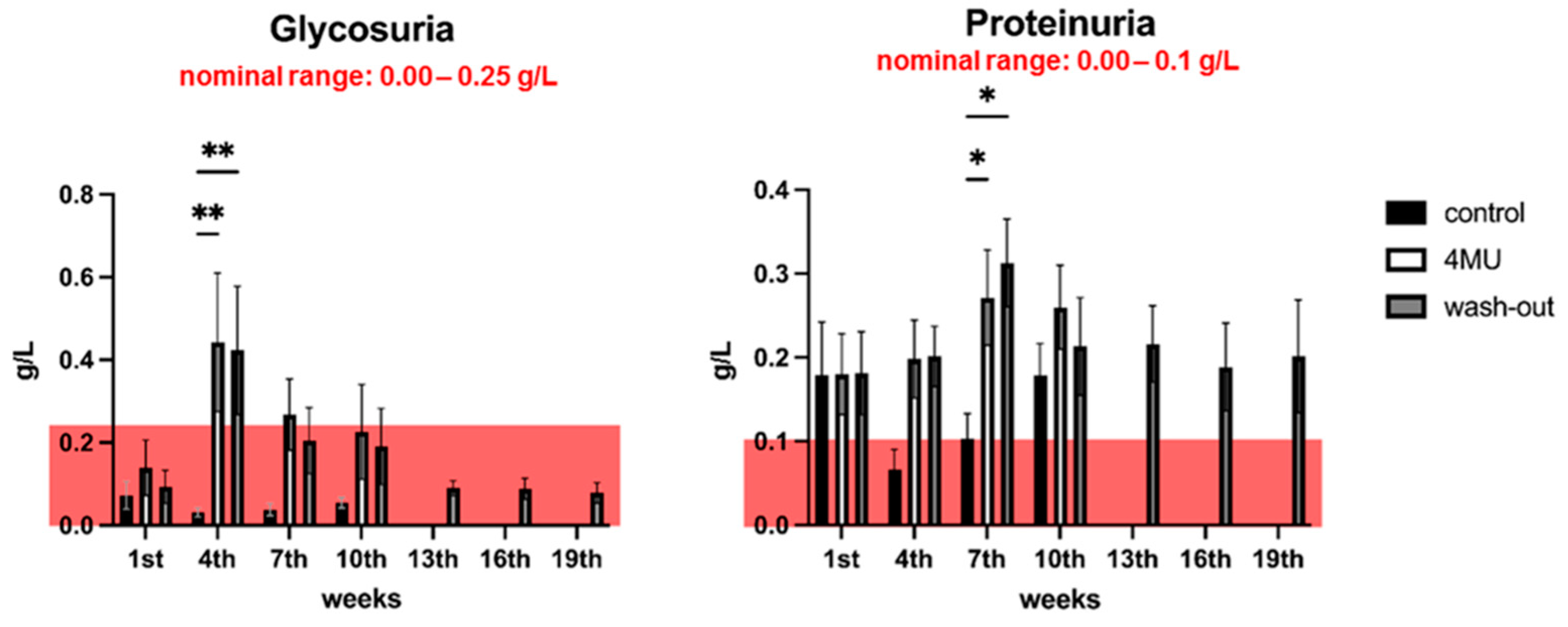
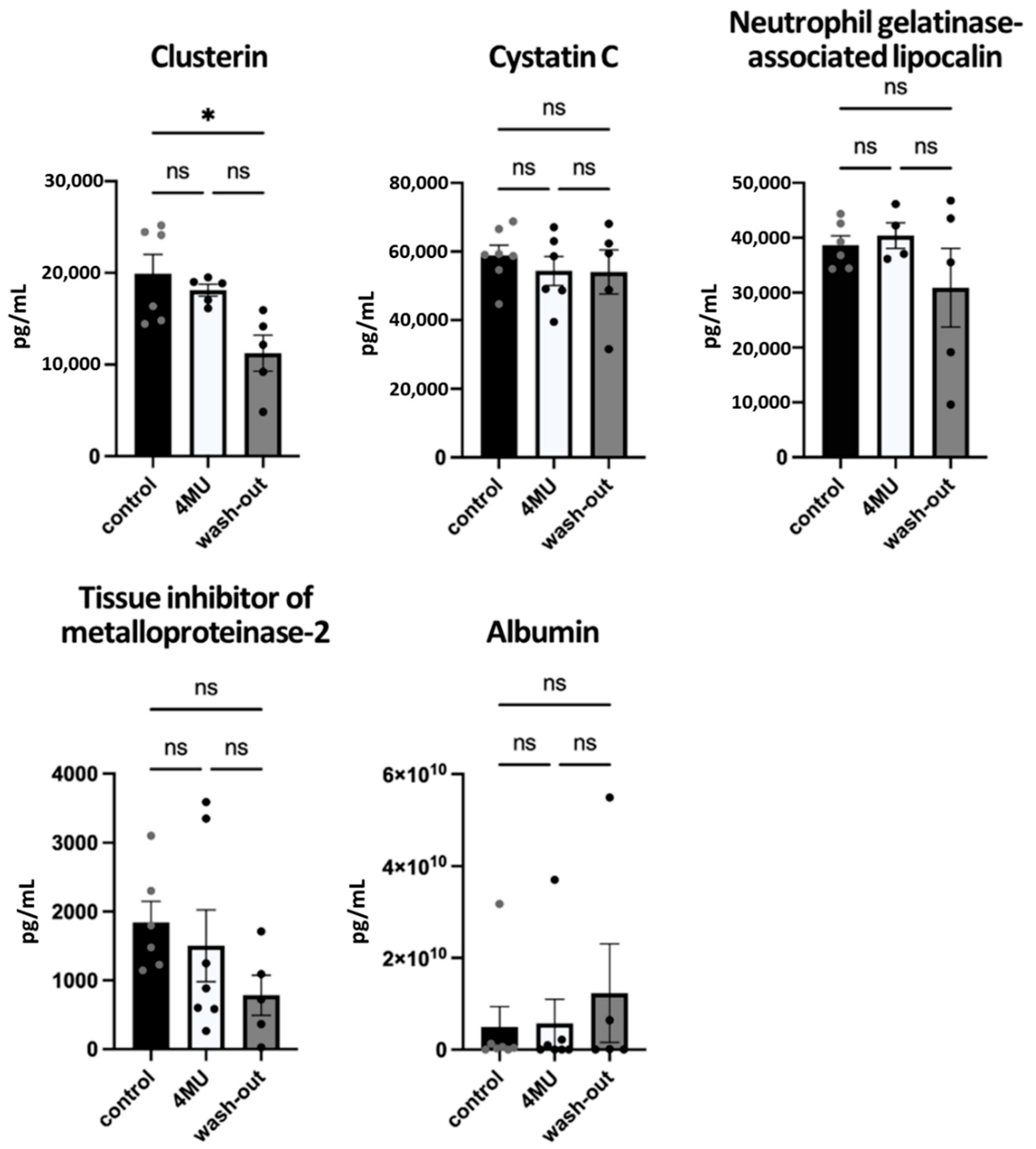
2.2. Neither Haematological nor Biochemical Parameters Indicate Adverse effects Caused by Long-Term Administration of 4MU at the Same Dose
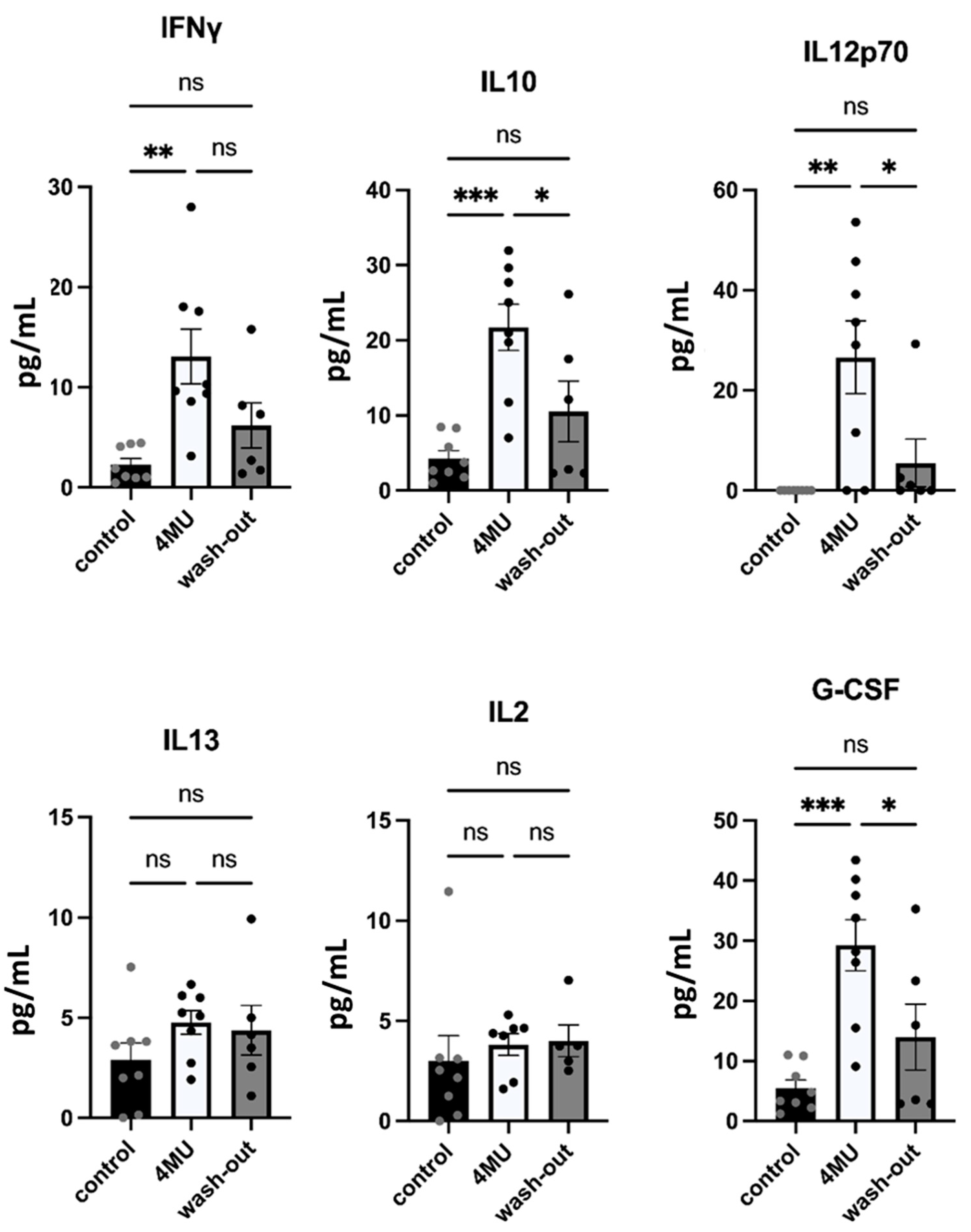
2.3. 1.2 g/kg/Day Dose of 4MU Increases Levels of IFN-γ, IL10 and IL12p70 in Blood Serum after 10 Weeks of Daily Administration but Values Returned Back to Control Levels during the Wash-Out Period
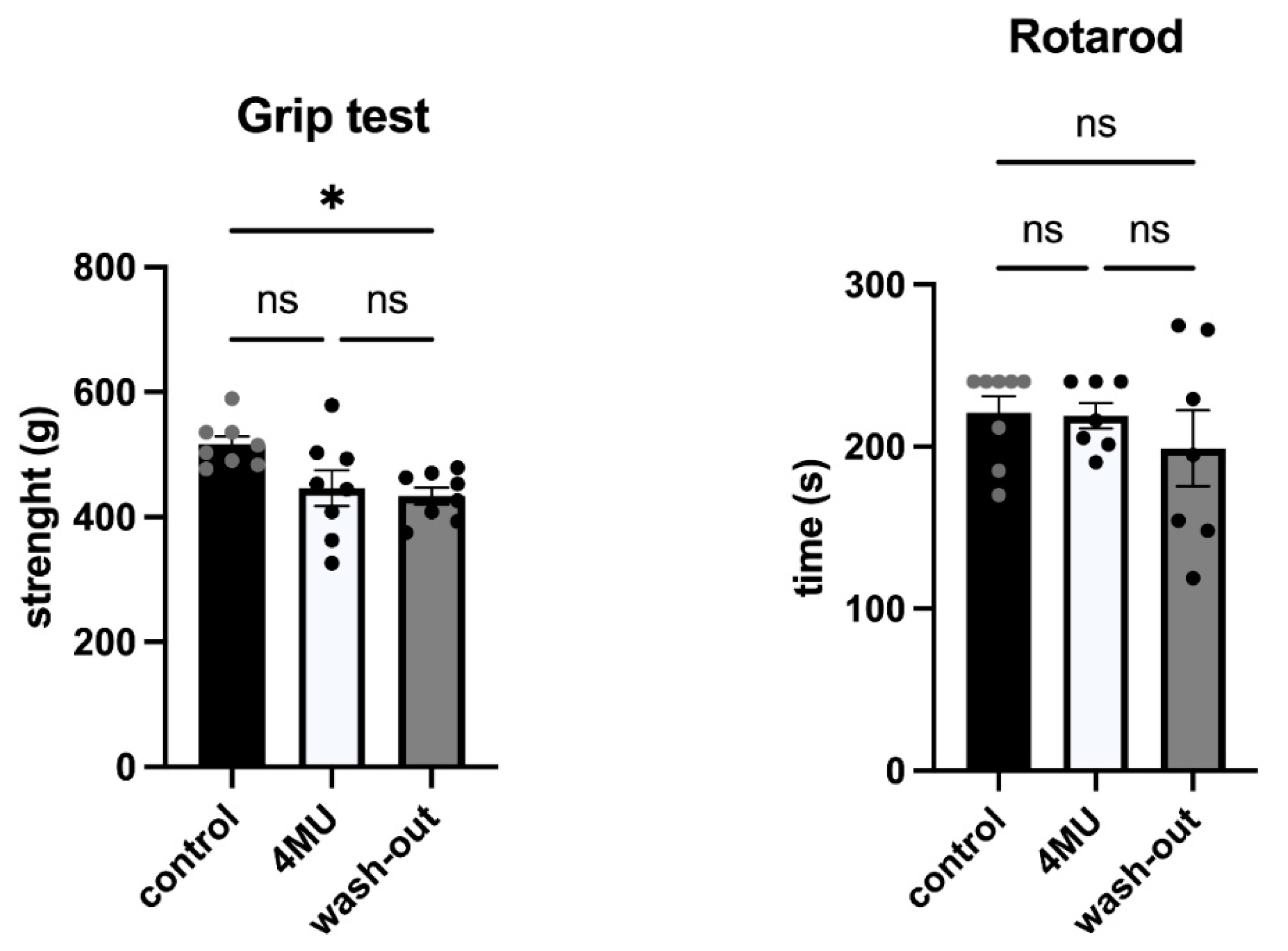
2.4. 1.2 g/kg/Day Dose of 4MU Does Not Affect Behavioural Performance of the Experimental Animals
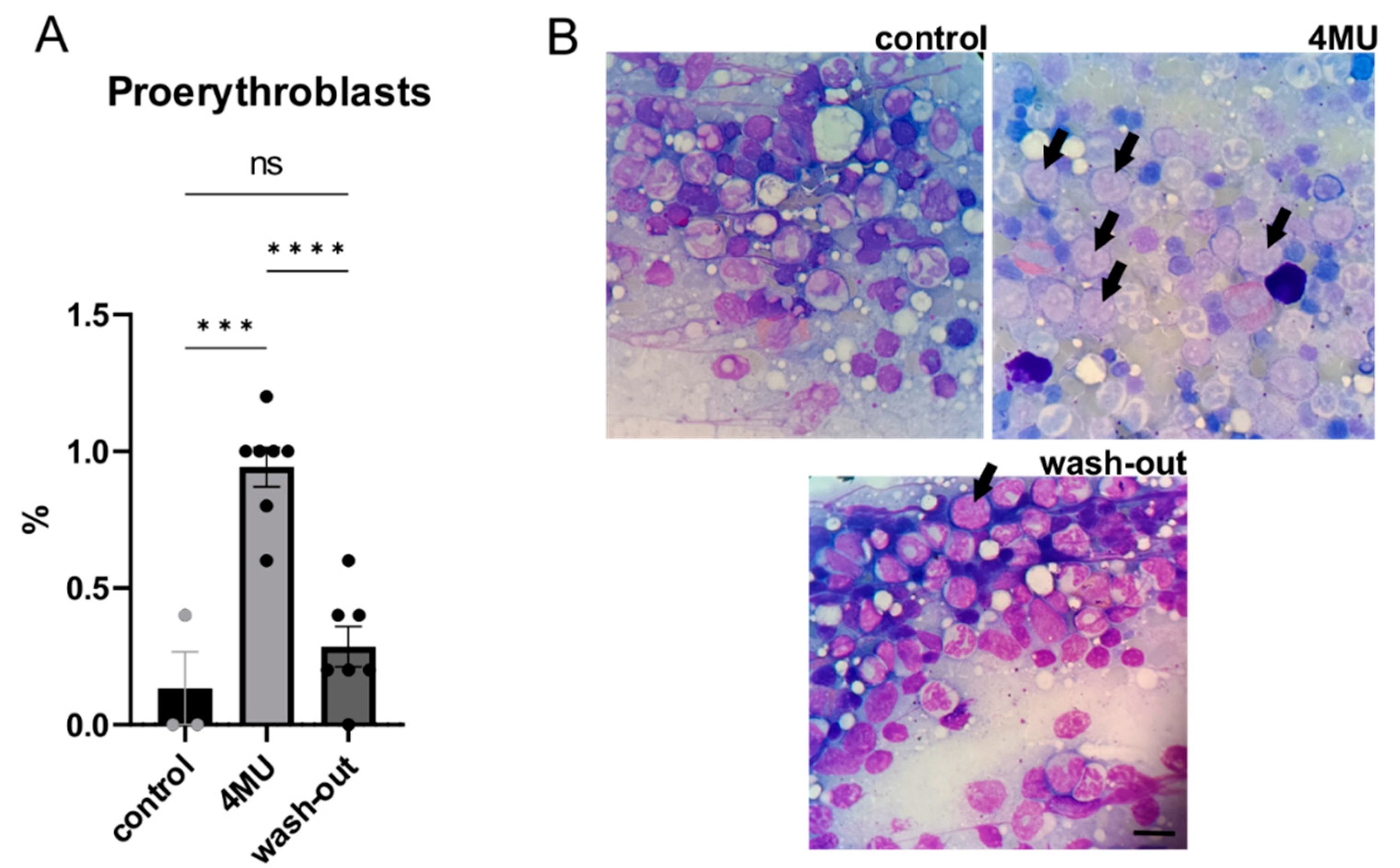
2.5. 1.2 g/kg/Day Dose of 4MU Increases the Relative Number of Proerythroblasts in Bone Marrow, but Returned to Normal after the Wash-Out Period
2.6. Long-Term 4MU Treatment at the Current Dose Does Not Affect the Biomechanical Properties of Tendons and Skin
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug Dosage
4.3. Haematology and Biochemistry
4.4. Immunohistochemistry
4.5. Proteomics
4.6. Behavioural Tests
4.7. Histopathological Evaluation
4.8. Bone Marrow Evaluation
4.9. Biomechanical Testing
4.9.1. Conditions and Equipment
4.9.2. Tensile Tests of Skin
4.9.3. Tensile Test of Tendons
4.9.4. Statistical Analysis of Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4MU | 4-methylumbelliferone |
| ACAN | Aggrecan |
| ALP | Alkaline phosphatase |
| ALT | Alanine transaminase |
| Apo-J | Apolipoprotein J |
| AST | Aspartate transaminase |
| BTM | Bone to muscle (deposition) |
| CNS | Central nervous system |
| CS-56 | Anti-chondroitin sulphate antibody |
| CS | Chondroitin sulphate |
| CSPGs | Chondroitin sulphate proteoglycans |
| Cys | Cystatin C |
| DAPI | 4’,6-diamidino-2-phenylindole, dihydrochloride |
| ECM | Extracellular matrix |
| FFPE | Formalin-fixed paraffin-embedded |
| GAG | Glycosaminoglycans |
| G-CSF | Granulocyte colony-stimulating factor |
| GlcA | D-glucuronic acid |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
| HA | Hyaluronan/hyaluronic acid |
| HAS | Hyaluronan synthase |
| HABP | Hyaluronan-binding protein |
| IFNγ | Interferon gamma |
| IL10 | Interleukin 10 |
| IL12p70 | Interleukin 12p70 |
| IL13 | Interleukin 13 |
| IL17a | Interleukin 17A |
| IL1α | Interleukin 1 alpha |
| IL1β | Interleukin 1 beta |
| IL2 | Interleukin 2 |
| IL4 | Interleukin 4 |
| IL5 | Interleukin 5 |
| IL6 | Interleukin 6 |
| IL10 | Interleukin 10 |
| JAK | Janus kinase |
| NGAL | Neutrophil gelatinase-associated lipocalin |
| PBS | Phosphate-buffered saline |
| PNNs | Perineuronal nets |
| RT | Room temperature |
| SCI | Spinal cord injury |
| STAT3 | Signal transducer and activator of transcription 3 |
| TIMP-1 | Tissue inhibitor of metalloproteinases 1 |
| TNFα | Tumour necrosis factor alpha |
| UDP-GlcA | UDP-glucuronic acid |
References
- Klos, C.; Paumgartner, G.; Reichen, J. Cation-anion gap and choleretic properties of rat bile. Am. J. Physiol. -Endocrinol. Metab. 1979, 236, E434. [Google Scholar] [CrossRef]
- Elferink, R.P.J.O.; Tytgat, G.N.J.; Groen, A.K. The role of mdr2 P-glycoprotein in hepatobiliary lipid transport. FASEB J. 1997, 11, 19–28. [Google Scholar] [CrossRef]
- Takeda, S.; Aburada, M. The choleretic mechanism of coumarin compounds and phenolic compounds. J. Pharm. 1981, 4, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Nagy, N.; Gurevich, I.; Kuipers, H.F.; Ruppert, S.M.; Marshall, P.L.; Xie, B.J.; Sun, W.; Malkovskiy, A.V.; Rajadas, J.; Grandoch, M.; et al. 4-Methylumbelliferyl glucuronide contributes to hyaluronan synthesis inhibition. J. Biol. Chem. 2019, 294, 7864–7877. [Google Scholar] [CrossRef]
- Dubisova, J.; Burianova, J.S.; Svobodova, L.; Makovicky, P.; Martinez-Varea, N.; Cimpean, A.; Fawcett, J.W.; Kwok, J.C.; Kubinova, S. Oral treatment of 4-methylumbelliferone reduced perineuronal nets and improved recognition memory in mice. Brain Res. Bull. 2022, 181, 144–156. [Google Scholar] [CrossRef]
- Irvine, S.F.; Gigout, S.; Štěpánková, K.; Varea, N.M.; Urdzíková, L.M.; Jendelová, P.; Kwok, J.C.F. 4-Methylumbelliferone enhances neuroplasticity in the central nervous system: Potential oral treatment for SCI. bioRxiv 2023, arXiv:2023.01.23.525137. [Google Scholar]
- Nagy, N.; Kuipers, H.F.; Frymoyer, A.R.; Ishak, H.D.; Bollyky, J.B.; Wight, T.N.; Bollyky, P.L. 4-Methylumbelliferone Treatment and Hyaluronan Inhibition as a Therapeutic Strategy in Inflammation, Autoimmunity, and Cancer. Front. Immunol. 2015, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Stepankova, K.; Chudickova, M.; Simkova, Z.; Martinez-Varea, N.; Kubinova, S.; Urdzikova, L.; Jendelova, P.; Kwok, J.C. Oral administration of 4-methylumbelliferone reduces glial scar and promotes anatomical plasticity. bioRxiv 2023, arXiv:2023.02.02.526565. [Google Scholar]
- Sorg, B.A.; Berretta, S.; Blacktop, J.M.; Fawcett, J.W.; Kitagawa, H.; Kwok, J.C.; Miquel, M. Casting a Wide Net: Role of Perineuronal Nets in Neural Plasticity. J. Neurosci. 2016, 36, 11459–11468. [Google Scholar] [CrossRef]
- Wang, D.; Fawcett, J. The perineuronal net and the control of CNS plasticity. Cell Tissue Res. 2012, 349, 147–160. [Google Scholar] [CrossRef]
- Ghorbani, S.; Yong, V.W. The extracellular matrix as modifier of neuroinflammation and remyelination in multiple sclerosis. Brain 2021, 144, 1958–1973. [Google Scholar] [CrossRef] [PubMed]
- Yong, N.; Guoping, C. Upregulation of matrix metalloproteinase-9 dependent on hyaluronan synthesis after sciatic nerve injury. Neurosci. Lett. 2008, 444, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, H.F.; Nagy, N.; Ruppert, S.M.; Sunkari, V.G.; Marshall, P.L.; A Gebe, J.; Ishak, H.D.; Keswani, S.G.; Bollyky, J.; Frymoyer, A.R.; et al. The pharmacokinetics and dosing of oral 4-methylumbelliferone for inhibition of hyaluronan synthesis in mice. Clin. Exp. Immunol. 2016, 185, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Garrett, E.R.; Venitz, J.; Eberst, K.; Cerda, J.J. Pharmacokinetics and bioavailabilities of hymecromone in human volunteers. Biopharm Drug Dispos 1993, 14, 13–39. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Sugiyama, Y.; Hanano, M. Pharmacokinetic study of 4-methylumbelliferone in rats: Influence of dose on its first-pass hepatic elimination. J. Pharm. 1986, 9, 117–124. [Google Scholar] [CrossRef]
- Kobayashi, T.; Chanmee, T.; Itano, N. Hyaluronan: Metabolism and Function. Biomolecules 2020, 10, 1525. [Google Scholar] [CrossRef]
- David, S.; Hamilton, J.P. Drug-induced Liver Injury. US Gastroenterol. Hepatol. Rev. 2010, 6, 73–80. [Google Scholar] [PubMed]
- Hoffmann, R.M.; Schwarz, G.; Pohl, C.; Ziegenhagen, D.J.; Kruis, W. [Bile acid-independent effect of hymecromone on bile secretion and common bile duct motility]. Dtsch. Med. Wochenschr. (1946) 2005, 130, 1938–1943. [Google Scholar] [CrossRef] [PubMed]
- Makris, K.; Markou, N.; Evodia, E.; Dimopoulou, E.; Drakopoulos, I.; Ntetsika, K.; Rizos, D.; Baltopoulos, G.; Haliassos, A. Urinary neutrophil gelatinase-associated lipocalin (NGAL) as an early marker of acute kidney injury in critically ill multiple trauma patients. Clin. Chem. Lab. Med. 2009, 47, 79–82. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Hong, Q.; Lin, H.; Zhu, H.; Liu, Q.; Wang, J.; Xie, Y.; Shang, X.; Shi, S.; et al. TIMP-1 promotes age-related renal fibrosis through upregulating ICAM-1 in human TIMP-1 transgenic mice. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 1130–1143. [Google Scholar] [CrossRef]
- Goncharova, V.; Serobyan, N.; Iizuka, S.; Schraufstatter, I.; de Ridder, A.; Povaliy, T.; Wacker, V.; Itano, N.; Kimata, K.; Orlovskaja, I.A.; et al. Hyaluronan expressed by the hematopoietic microenvironment is required for bone marrow hematopoiesis. J. Biol. Chem. 2012, 287, 25419–25433. [Google Scholar] [CrossRef] [PubMed]
- Sikes, K.J.; Renner, K.; Li, J.; Grande-Allen, K.J.; Connell, J.P.; Cali, V.; Midura, R.J.; Sandy, J.D.; Plaas, A.; Wang, V.M. Knockout of hyaluronan synthase 1, but not 3, impairs formation of the retrocalcaneal bursa. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2018, 36, 2622–2632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermato-Endocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef]
- Nakamura, T.; Takagaki, K.; Shibata, S.; Tanaka, K.; Higuchi, T.; Endo, M. Hyaluronic-Acid-Deficient Extracellular Matrix Induced by Addition of 4-Methylumbelliferone to the Medium of Cultured Human Skin Fibroblasts. Biochem. Biophys. Res. Commun. 1995, 208, 470–475. [Google Scholar] [CrossRef]
- Kakizaki, I.; Kojima, K.; Takagaki, K.; Endo, M.; Kannagi, R.; Ito, M.; Maruo, Y.; Sato, H.; Yasuda, T.; Mita, S.; et al. A novel mechanism for the inhibition of hyaluronan biosynthesis by 4-methylumbelliferone. J. Biol. Chem. 2004, 279, 33281–33289. [Google Scholar] [CrossRef] [PubMed]
- Monslow, J.; Govindaraju, P.; Puré, E. Hyaluronan—A functional and structural sweet spot in the tissue microenvironment. Front Immunol. 2015, 6, 231. [Google Scholar] [CrossRef]
- Vigetti, D.; Rizzi, M.; Viola, M.; Karousou, E.; Genasetti, A.; Clerici, M.; Bartolini, B.; Hascall, V.C.; De Luca, G.; Passi, A. The effects of 4-methylumbelliferone on hyaluronan synthesis, MMP2 activity, proliferation, and motility of human aortic smooth muscle cells. Glycobiology 2009, 19, 537–546. [Google Scholar] [CrossRef]
- Dicker, K.T.; Gurski, L.A.; Pradhan-Bhatt, S.; Witt, R.L.; Farach-Carson, M.C.; Jia, X. Hyaluronan: A simple polysaccharide with diverse biological functions. Acta Biomater. 2014, 10, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Sherman, L.S. Diverse Roles for Hyaluronan and Hyaluronan Receptors in the Developing and Adult Nervous System. Int. J. Mol. Sci. 2020, 21, 5988. [Google Scholar] [CrossRef]
- Smith, C.C.; Mauricio, R.; Nobre, L.; Marsh, B.; Wüst, R.C.; Rossiter, H.B.; Ichiyama, R.M. Differential regulation of perineuronal nets in the brain and spinal cord with exercise training. Brain Res Bull. 2015, 111, 20–26. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Berretta, S. In Sickness and in Health: Perineuronal Nets and Synaptic Plasticity in Psychiatric Disorders. Neural Plast. 2016, 2016, 9847696. [Google Scholar] [CrossRef]
- De la Motte, C.A.; Kessler, S.P. The role of hyaluronan in innate defense responses of the intestine. Int. J. Cell Biol. 2015, 2015, 481301. [Google Scholar] [CrossRef] [Green Version]
- Galdi, F.; Pedone, C.; McGee, C.A.; George, M.; Rice, A.B.; Hussain, S.S.; Vijaykumar, K.; Boitet, E.R.; Tearney, G.J.; McGrath, J.A.; et al. Inhaled high molecular weight hyaluronan ameliorates respiratory failure in acute COPD exacerbation: A pilot study. Respir. Res. 2021, 22, 30. [Google Scholar] [CrossRef]
- Papanastasopoulou, C.; Papastamataki, M.; Karampatsis, P.; Anagnostopoulou, E.; Papassotiriou, I.; Sitaras, N. Cardiovascular Risk and Serum Hyaluronic Acid: A Preliminary Study in a Healthy Population of Low/Intermediate Risk. J. Clin. Lab. Anal. 2017, 31, e22010. [Google Scholar] [CrossRef]
- Amir, A.; Kim, S.; Stecco, A.; Jankowski, M.P.; Raghavan, P. Hyaluronan homeostasis and its role in pain and muscle stiffness. PM R J. Inj. Funct. Rehabil. 2022, 14, 1490–1496. [Google Scholar] [CrossRef]
- Kaux, J.F.; Samson, A.; Crielaard, J.M. Hyaluronic acid and tendon lesions. Muscles Ligaments Tendons J. 2015, 5, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Tamer, T.M. Hyaluronan and synovial joint: Function, distribution and healing. Interdiscip. Toxicol. 2013, 6, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Rivers, D.A.; Stern, R. Hyaluronan and the Process of Aging in Skin. In Textbook of Aging Skin; Farage, M., Miller, K., Maibach, H., Eds.; Springer: Berlin, Heidelberg, 2017. [Google Scholar]
- Hansen, J.F.; Christiansen, K.M.; Staugaard, B.; Moessner, B.K.; Lillevang, S.; Krag, A.; Christensen, P.B. Combining liver stiffness with hyaluronic acid provides superior prognostic performance in chronic hepatitis C. PLoS ONE 2019, 14, e0212036. [Google Scholar] [CrossRef]
- Katarey, D.; Verma, S. Drug-induced liver injury. Clin. Med. 2016, 16 (Suppl. 6), s104–s109. [Google Scholar] [CrossRef]
- Kakehi, K.; Kinoshita, M.; Yasueda, S.-i. Hyaluronic acid: Separation and biological implications. J. Chromatogr. B 2003, 797, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef]
- Fenderson, B.A.; Stamenkovic, I.; Aruffo, A. Localization of hyaluronan in mouse embryos during implantation, gastrulation and organogenesis. Differ. Res. Biol. Divers. 1993, 54, 85–98. [Google Scholar]
- Tammi, M.; Seppälä, P.O.; Lehtonen, A.; Möttönen, M. Connective tissue components in normal and atherosclerotic human coronary arteries. Atherosclerosis 1978, 29, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Smadja-Joffe, F.; Legras, S.; Girard, N.; Li, Y.; Delpech, B.; Bloget, F.; Morimoto, K.; Le Bousse-Kerdilès, C.; Clay, D.; Jasmin, C.; et al. CD44 and hyaluronan binding by human myeloid cells. Leuk. Lymphoma 1996, 21, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Drózdz, M.; Kula, B.; Wardas, M.; Weglarz, L. Hyaluronic acid content and hyaluronidase activity in liver and spleen of rats with hydralazine-induced collagen-like syndrome. Biomed. Biochim. Acta 1988, 47, 247–250. [Google Scholar] [PubMed]
- Andreutti, D.; Geinoz, A.; Gabbiani, G. Effect of hyaluronic acid on migration, proliferation and alpha-smooth muscle actin expression by cultured rat and human fibroblasts. J. Submicrosc. Cytol. Pathol. 1999, 31, 173–177. [Google Scholar]
- Klamer, S.; Voermans, C. The role of novel and known extracellular matrix and adhesion molecules in the homeostatic and regenerative bone marrow microenvironment. Cell Adhes. Migr. 2014, 8, 563–577. [Google Scholar] [CrossRef]
- Köpke-Aguiar, L.A.; Martins, J.R.; Passerotti, C.C.; Toledo, C.F.; Nader, H.B.; Borges, D.R. Serum hyaluronic acid as a comprehensive marker to assess severity of liver disease in schistosomiasis. Acta Trop. 2002, 84, 117–126. [Google Scholar] [CrossRef]
- Malakouti, M.; Kataria, A.; Ali, S.K.; Schenker, S. Elevated Liver Enzymes in Asymptomatic Patients-What Should I Do? J. Clin. Transl. Hepatol. 2017, 5, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Dufour, D.R.; Lott, J.A.; Nolte, F.S.; Gretch, D.R.; Koff, R.S.; Seeff, L.B. Diagnosis and monitoring of hepatic injury. II. Recommendations for use of laboratory tests in screening, diagnosis, and monitoring. Clin. Chem. 2000, 46, 2050–2068. [Google Scholar] [CrossRef]
- Miura, Y. [Aspartate aminotransferase (AST) and alanine aminotransferase (ALT)]. Nihon Rinsho Jpn. J. Clin. Med. 1995, 53 Pt 1, 266–271. [Google Scholar]
- Kvietys, P.R.; Granger, D.N. Role of intestinal lymphatics in interstitial volume regulation and transmucosal water transport. Ann. N. Y. Acad. Sci. 2010, 1207 (Suppl. 1), E29–E43. [Google Scholar] [CrossRef] [PubMed]
- De la Motte, C.A.; Hascall, V.C.; Drazba, J.; Bandyopadhyay, S.K.; Strong, S.A. Mononuclear leukocytes bind to specific hyaluronan structures on colon mucosal smooth muscle cells treated with polyinosinic acid:polycytidylic acid: Inter-alpha-trypsin inhibitor is crucial to structure and function. Am. J. Pathol. 2003, 163, 121–133. [Google Scholar] [CrossRef]
- Kaul, A.; Singampalli, K.L.; Parikh, U.M.; Yu, L.; Keswani, S.G.; Wang, X. Hyaluronan, a double-edged sword in kidney diseases. Pediatr. Nephrol. 2022, 37, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Colombaro, V.; Declèves, A.E.; Jadot, I.; Voisin, V.; Giordano, L.; Habsch, I.; Nonclercq, D.; Flamion, B.; Caron, N. Inhibition of hyaluronan is protective against renal ischaemia-reperfusion injury. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2013, 28, 2484–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, J.R.; Laurent, T.C.; Laurent, U.B. Hyaluronan: Its nature, distribution, functions and turnover. J. Intern. Med. 1997, 242, 27–33. [Google Scholar] [CrossRef]
- Laurent, C.; Johnson-Wells, G.; Hellström, S.; Engström-Laurent, A.; Wells, A.F. Localization of hyaluronan in various muscular tissues. A morphological study in the rat. Cell Tissue Res. 1991, 263, 201–205. [Google Scholar] [CrossRef]
- Stecco, C.; Stern, R.; Porzionato, A.; Macchi, V.; Masiero, S.; Stecco, A.; De Caro, R. Hyaluronan within fascia in the etiology of myofascial pain. Surg. Radiol. Anat. SRA 2011, 33, 891–896. [Google Scholar] [CrossRef]
- Purslow, P.P. Muscle fascia and force transmission. J. Bodyw. Mov. Ther. 2010, 14, 411–417. [Google Scholar] [CrossRef]
- Hunter, S.K.; Pereira, H.M.; Keenan, K.G. The aging neuromuscular system and motor performance. J. Appl. Physiol. 2016, 121, 982–995. [Google Scholar] [CrossRef]
- Stern, R.; Maibach, H.I. Hyaluronan in skin: Aspects of aging and its pharmacologic modulation. Clin. Dermatol. 2008, 26, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Longaker, M.T.; Chiu, E.S.; Adzick, N.S.; Stern, M.; Harrison, M.R.; Stern, R. Studies in fetal wound healing. V. A prolonged presence of hyaluronic acid characterizes fetal wound fluid. Ann. Surg. 1991, 213, 292–296. [Google Scholar] [CrossRef]
- Schachtschabel, D.O.; Wever, J. Age-related decline in the synthesis of glycosaminoglycans by cultured human fibroblasts (WI-38). Mech. Ageing Dev. 1978, 8, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.S.; Kemparaju, K. The magic glue hyaluronan and its eraser hyaluronidase: A biological overview. Life Sci. 2007, 80, 1921–1943. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, M.R.; Burdick, M.D.; Strieter, R.M.; Bao, C.; Noble, P.W. Regulation of hyaluronan-induced chemokine gene expression by IL-10 and IFN-gamma in mouse macrophages. J. Immunol. 1998, 160, 3023–3030. [Google Scholar] [CrossRef]
- King, A.; Balaji, S.; Le, L.D.; Crombleholme, T.M.; Keswani, S.G. Regenerative Wound Healing: The Role of Interleukin-10. Adv. Wound Care 2014, 3, 315–323. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Skin | ||
| Young modulus E5 (5%)/MPa | control | 9.326 ± 1.113 |
| 4MU | 13.57 ± 1.856 | |
| wash-out | 10.43 ± 1.240 | |
| Young modulus E10 (10%)/MPa | control | 16.43 ± 2.910 |
| 4MU | 16.06 ± 2.702 | |
| wash-out | 16.79 ± 3.704 | |
| Tendons | ||
| Maximum force Fmax/N | control | 20.11 ± 2.673 |
| 4MU | 14.75 ± 1.746 | |
| wash-out | 23.56 ± 3.708 | |
| Maximum stress σmax/Nm−2 | control | 23.2 ± 3.015 |
| 4MU | 22.51 ± 3.531 | |
| wash-out | 38.83 ± 7.662 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štěpánková, K.; Mareková, D.; Kubášová, K.; Sedláček, R.; Turnovcová, K.; Vacková, I.; Kubinová, Š.; Makovický, P.; Petrovičová, M.; Kwok, J.C.F.; et al. 4-Methylumbeliferone Treatment at a Dose of 1.2 g/kg/Day Is Safe for Long-Term Usage in Rats. Int. J. Mol. Sci. 2023, 24, 3799. https://doi.org/10.3390/ijms24043799
Štěpánková K, Mareková D, Kubášová K, Sedláček R, Turnovcová K, Vacková I, Kubinová Š, Makovický P, Petrovičová M, Kwok JCF, et al. 4-Methylumbeliferone Treatment at a Dose of 1.2 g/kg/Day Is Safe for Long-Term Usage in Rats. International Journal of Molecular Sciences. 2023; 24(4):3799. https://doi.org/10.3390/ijms24043799
Chicago/Turabian StyleŠtěpánková, Kateřina, Dana Mareková, Kristýna Kubášová, Radek Sedláček, Karolína Turnovcová, Irena Vacková, Šárka Kubinová, Pavol Makovický, Michaela Petrovičová, Jessica C. F. Kwok, and et al. 2023. "4-Methylumbeliferone Treatment at a Dose of 1.2 g/kg/Day Is Safe for Long-Term Usage in Rats" International Journal of Molecular Sciences 24, no. 4: 3799. https://doi.org/10.3390/ijms24043799
APA StyleŠtěpánková, K., Mareková, D., Kubášová, K., Sedláček, R., Turnovcová, K., Vacková, I., Kubinová, Š., Makovický, P., Petrovičová, M., Kwok, J. C. F., Jendelová, P., & Machová Urdzíková, L. (2023). 4-Methylumbeliferone Treatment at a Dose of 1.2 g/kg/Day Is Safe for Long-Term Usage in Rats. International Journal of Molecular Sciences, 24(4), 3799. https://doi.org/10.3390/ijms24043799