
Dipotassium Glycyrrhizininate Improves Skin Wound Healing by Modulating Inflammatory Process

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
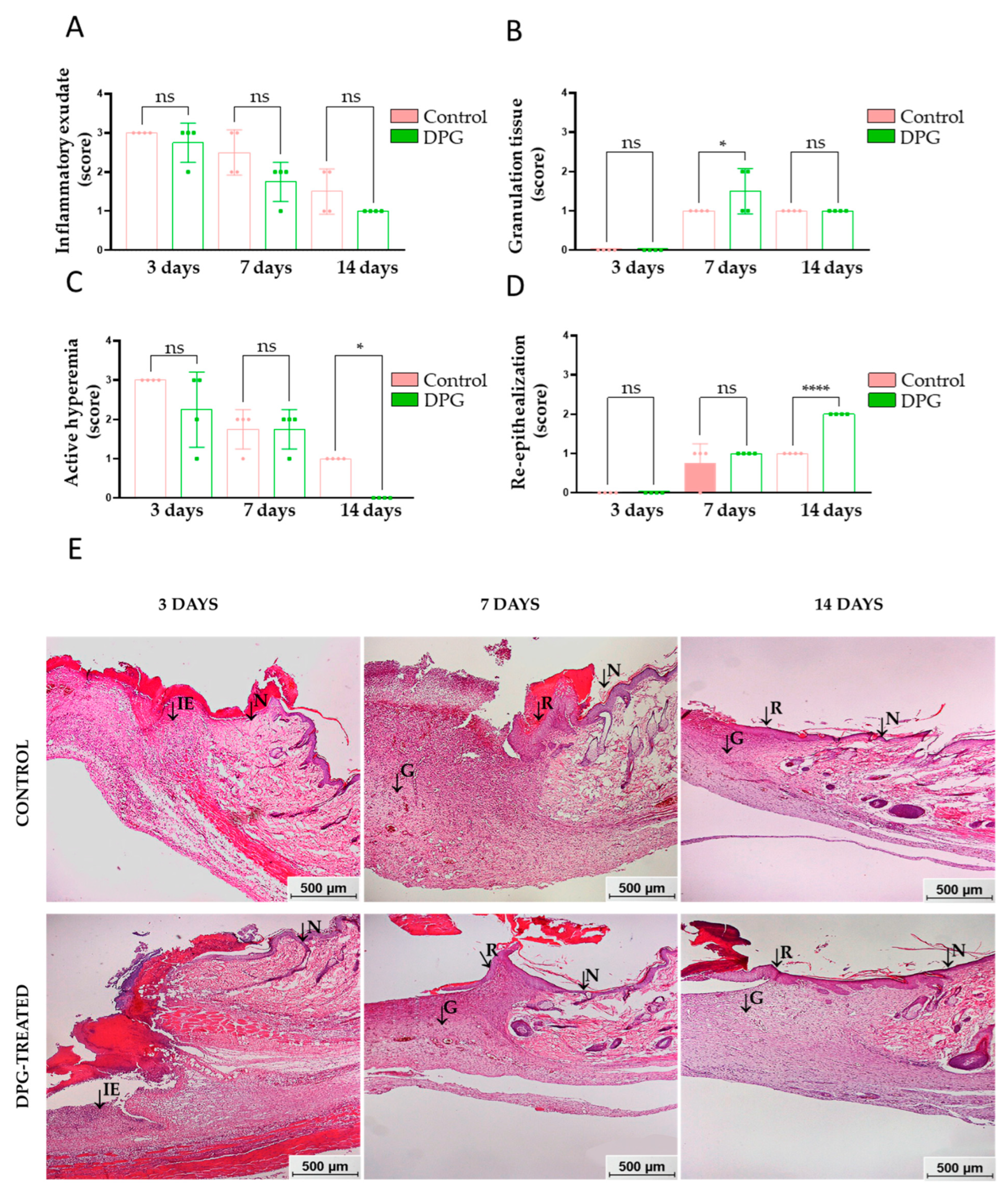
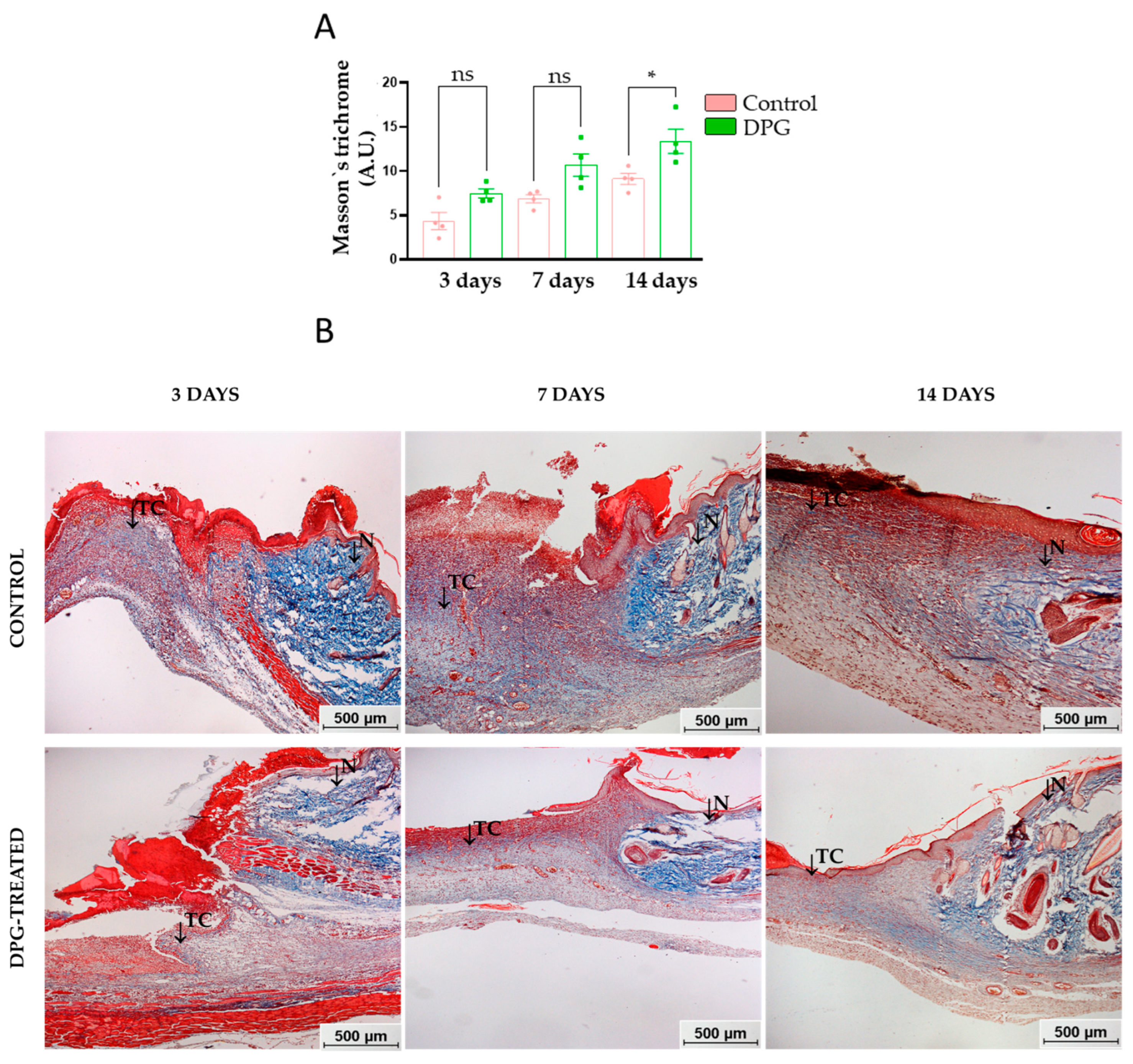
2.1. DPG Attenuates Inflammation, Promoting Formation of New Granulation Tissue and Tissue Re-Epithelialization
2.2. DPG Modulates the Expression of Pro- and Anti-Inflammatory, Proliferative, and Remodeling Genes
3. Materials and Methods
3.1. DPG Gel Cream
3.2. Animals and Experimental Groups
3.3. Excision Wound Model
3.4. Topical Treatment
3.5. Collection, Storage, and Processing of Samples
3.6. Macroscopic Analysis
3.7. Microscopic Analysis—Inflammatory and Proliferative Scores
3.8. RNA Extraction and Reverse Transcription Quantitative PCR (qPCR)
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shedoeva, A.; Leavesley, D.; Upton, Z.; Fan, C. Wound Healing and the Use of Medicinal Plants. Evid. Based. Complement. Altern. Med. 2019, 22, 2684108. [Google Scholar] [CrossRef]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Lindley, L.E.; Stojadinovic, O.; Pastar, I.; Tomic-Canic, M. Biology and biomarkers for wound healing. Plast. Reconstr. Surg. 2016, 138, 18–28. [Google Scholar] [CrossRef]
- Scully, D.; Sfyri, P.; Wilkinson, H.N.; Acebes-Huerta, A.; Verpoorten, S.; Muñoz-Turrillas, M.C.; Parnell, A.; Patel, K.; Hardman, M.J.; Gutiérrez, L.; et al. Optimising platelet secretomes to deliver robust tissue-specific regeneration. J. Tissue Eng. Regen. Med. 2020, 14, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Golebiewska, E.M.; Poole, A.W. Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef]
- Germolec, D.R.; Shipkowski, K.A.; Frawley, R.P.; Evans, E. Markers of Inflammation. Meth. Mol. Biol. 2018, 1803, 57–79. [Google Scholar] [CrossRef]
- Weiss, U. Inflammation. Nature 2008, 454, 7203. [Google Scholar] [CrossRef]
- Yuzhakova, D.V.; Shirmanova, M.V.; Bocharov, A.A.; Astrakhantseva, I.V.; Vasilenko, E.A.; Gorshkova, E.N.; Drutskaya, M.S.; Zagaynova, E.V.; Nedospasov, S.A.; Kruglov, A.A. Microbiota Induces Expression of Tumor Necrosis Factor in Postnatal Mouse Skin. Biochemistry 2016, 81, 1303–1308. [Google Scholar] [CrossRef]
- Macleod, T.; Berekmeri, A.; Bridgewood, C.; Stacey, M.; Mcgonagle, D.; Wittmann, M. The Immunological Impact of IL-1 Family Cytokines on the Epidermal Barrier. Front. Immunol. 2021, 23, 808012. [Google Scholar] [CrossRef]
- Tan, Q.; Huang, Q.; Ma, Y.L.; Mao, K.; Yang, G.; Luo, P.; Ma, G.; Mei, P.; Jin, Y. Potential roles of IL-1 subfamily members in glycolysis in disease. Cytokine Growth Factor Rev. 2018, 44, 18–27. [Google Scholar] [CrossRef]
- Martin, P.; Goldstein, J.D.; Mermoud, L.; Diaz-Barreiro, A.; Palmer, G. IL-1 Family Antagonists in Mouse and Human Skin Inflammation. Front. Immunol. 2021, 16, 652846. [Google Scholar] [CrossRef] [PubMed]
- Shaw, T.J.; Martin, P. Wound repair: A showcase for cell plasticity and migration. Curr. Opin. Cell Biol. 2016, 42, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Plikus, M.V.; Guerrero-Juarez, C.F.; Ito, M.; Li, Y.R.; Dedhia, P.H.; Zheng, Y.; Shao, M.; Gay, D.l.; Ramos, R.; Hsi, T.-C.; et al. Regeneration of fat cells from myofibroblasts during wound healing. Science 2017, 355, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, A.; Chattopadhyay, P.; Goyary, D.; Mazumder, P.M.; Veer, V. Eleutherine indica L. accelerates in vivo cutaneous wound healing by stimulating Smad-mediated collagen production. J. Ethnopharmacol. 2013, 146, 490–494. [Google Scholar] [CrossRef]
- Thomé, R.G.; Santos, H.B.; Santos, F.V.; Oliveira, R.J.S.O.; Camargos, L.F.; Pereira, M.N.; Longatti, T.R.; Souto, C.M.; Franco, C.S.; Schüffner, R.O.A.; et al. Evaluation of healing wound and genotoxicity potentials from extracts hydroalcoholic of Plantago major and Siparuna guianensis. Exp. Biol. Med. 2012, 237, 1379–1386. [Google Scholar] [CrossRef]
- Barone, A.; Cristiano, M.C.; Cilurzo, F.; Locatelli, M.; Iannotta, D.; Marzio, L.D.; Celia, C.; Paolino, D. Ammonium glycyrrhizate skin delivery from ultradeformable liposomes: A novel use as an anti-inflammatory agent in topical drug delivery. Colloids Surf. B. Biointerfaces 2020, 193, 111152. [Google Scholar] [CrossRef]
- Rizzato, G.; Scalabrin, E.; Radaelli, M.; Capodaglio, G.; Iccolo, O. A new exploration of licorice metabolome. Food Chem. 2017, 15, 959–968. [Google Scholar] [CrossRef]
- Wang, Q.; Qian, Y.; Wang, Q.; Yang, Y.F.; Ji, S.; Song, W.; Qiao, X.; Guo, D.A.; Liang, H.; Ye, M. Metabolites identification of bioactive licorice compounds in rats. J. Pharm. Biomed. Anal. 2015, 10, 515–522. [Google Scholar] [CrossRef]
- Simmler, C.; Pauli, G.F.; Chen, S.N. Phytochemistry and biological properties of glabridin. Fitoterapia 2013, 90, 160–184. [Google Scholar] [CrossRef]
- Dastagir, G.; Rizvi, M.A. Review—Glycyrrhiza glabra L. (Liquorice). Pak. J. Pharm. Sci. 2016, 29, 727–1733. [Google Scholar]
- Yu, J.Y.; Ha, J.Y.; Kim, K.M.; Jung, Y.S.; Jung, J.C.; Oh, S. Anti-inflammatory activities of licorice extract and its active compounds, glycyrrhizic acid, liquiritin and liquiritigenin, in BV2 cells and mice liver. Molecules 2015, 20, 13041–13054. [Google Scholar] [CrossRef]
- Leite, C.S.; Bonafé, G.A.; Santos, J.C.; Martinez, C.A.R.M.; Ortega, M.M.; Ribeiro, M.L. The Anti-Inflammatory Properties of Licorice (Glycyrrhiza glabra) Derived Compounds in Intestinal Disorders. Int. J. Mol. Sci. 2022, 23, 4121. [Google Scholar] [CrossRef]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues; Oliveira, F.M.B.P.P. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef]
- Sun, Z.G.; Zhao, T.T.; Lu, N.; Yang, Y.A.; Zhu, H.L. Research Progress of Glycyrrhizic Acid on Antiviral Activity. Mini Rev. Med. Chem. 2019, 19, 826–832. [Google Scholar] [CrossRef]
- Salem, M.M.N.M.; Mohammed, A.A.W.; Al-Tameemi, J.A.; Sulaiman, G.M. Biological study of the effect of licorice roots extract on serum lipid profile, liver enzymes and kidney function tests in albino mice. Afr. J. Biotechnol. 2011, 12702–12706. [Google Scholar] [CrossRef]
- Vitali, R.; Palone, F.; Pierdomenico, M.; Negroni, A.; Cucchiara, S.; Aloi, M.; Oliva, S.; Stronati, L. Dipotassium glycyrrhizate via HMGB1 or AMPK signaling suppresses oxidative stress during intestinal inflammation. Biochem. Pharmacol. 2015, 97, 292–299. [Google Scholar] [CrossRef]
- Shim, J.M.; Yim, S.B.; Chung, J.H.; Hong, K.S. Antiplaque and antigingivitis effects of a mouthrinse containing cetylpyridinium chloride, triclosan and dipotassium glycyrrhizinate. J. Period. Implant. Sci. 2012, 42, 33–38. [Google Scholar] [CrossRef]
- Andersen, F.A. Final report on the safety assessment of glycyrrhetinic acid, potassium glycyrrhetinate, disodium succinoyl gly- cyrrhetinate, glyceryl glycyrrhetinate, glycyrrhetinyl stearate, stearyl glycyrrhetinate, glycyrrhizic acid, ammonium glycyrrhizate, dipotassium glycyrrhizate, disodium glycyrrhizate, trisodium glycyrrhizate, methyl glycyrrhizate, and potassium glycyrrhizinate. Int. J. Toxicol. 2007, 26, 79–112. [Google Scholar] [CrossRef]
- Li, S.; Zhu, J.H.; Cao, L.P.; Sun, Q.; Liu, H.D.; De Li, W.; Li, J.S.; Hang, C.H. Growth inhibitory in vitro effects of glycyrrhizic acid in U251 glioblastoma cell line. Neurol. Sci. 2014, 35, 1115–1120. [Google Scholar] [CrossRef]
- Bonafé, G.A.; Santos, J.S.; Ziegler, J.V.; Umezawa, K.; Ribeiro, M.L.; Rocha, T.; Ortega, M.M. Growth Inhibitory Effects of Dipotassium Glycyrrhizinate in Glioblastoma Cell Lines by Target MicroRNAs Through the NF-k B Signaling Pathway. Front. Cell. Neurosci. 2019, 13, 1–14. [Google Scholar] [CrossRef]
- Leite, C.S.; Pires, O.C.; Tenis, D.G.; Ziegler, J.V.N.; Priolli, D.G.; Rocha, T. Effects of dipotassium glycyrrhizinate on wound healing. Acta Cir. Bras. 2021, 36, 360801. [Google Scholar] [CrossRef]
- Huang, W.; Zhou, X. Anti-histamine effects of dipotassium glycyrrhizinate on lung fibroblasts, implicating its therapeutic mechanism for pulmonary fibrosis. J. Pharm. Pharmacol. 2022, 1241–1250. [Google Scholar] [CrossRef]
- Nosenko, M.A.; Ambaryan, S.G.; Drutskaya, M.S. Proinflammatory Cytokines and Skin Wound Healing in Mice. Mol. Biol. 2019, 53, 741–754. [Google Scholar] [CrossRef]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef]
- Siriwattanasatorn, M.; Itharat, A.; Thongdeeying, P.; Ooraikul, B. In Vitro Wound Healing Activities of Three Most Commonly Used Thai Medicinal Plants and Their Three Markers. Evid. Based Complement. Altern. Med. 2020, 2020, 6795383. [Google Scholar] [CrossRef]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The role of vascular endothelial growth factor in wound healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug. Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef]
- Hao, B.; Wang, X.; Ma, X.; Jin, Y.; Fan, W.; Laba, C.; Wujin, C.; Wang, Y.; Liang, J. Preparation of complex microcapsules of soluble polysaccharide from Glycyrrhiza uralensis and its application in wound repair and scar inhibition. Int. J. Biol. Macromol. 2020, 156, 906–917. [Google Scholar] [CrossRef]
- Kasuya, A.; Tokura, Y. Attempts to accelerate wound healing. J. Dermatol. Sci. 2014, 76, 169–172. [Google Scholar] [CrossRef]
- Ruh, A.C.; Fernandes, D.; Artoni, R.F.; Favero, G.M. Inflamação: Entre a regeneração e a cicatrização. Publ. UEPG Ciências Biológicas Saúde 2013, 19, 11–19. [Google Scholar] [CrossRef]
- Isaac, C.; Ladeira, P.R.S.; Rego, F.M.P.; Aldunate, J.C.B.; Ferreira, M.C. Processo de cura das feridas: Cicatrização fisiológica. Rev. Med. 2010, 89, 125–131. [Google Scholar] [CrossRef]
- Johnson, K.E.; Wilgus, T.A. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Kotian, S.; Bhat, K.; Pai, S.; Nayak, J.; Souza, A.; Gourisheti, K.; Padma, D. The Role of Natural Medicines on Wound Healing: A Biomechanical, Histological, Biochemical and Molecular Study. Ethiop. J. Health Sci. 2018, 6, 759–770. [Google Scholar] [CrossRef]
- Assar, D.H.; Elhabashi, N.; Mokhbatly, A.A.; Ragab, A.E.; Elbialy, Z.I.; Rizk, S.A.; Albalawi, A.E.; Althobaiti, N.A.; Al Jaouni, S.; Atiba, A. Wound healing potential of licorice extract in rat model: Antioxidants, histopathological, immunohistochemical and gene expression evidences. Biomed. Pharmacother. 2021, 143, 112151. [Google Scholar] [CrossRef]
- Reilly, D.M.; Lozano, J. Skin collagen through the lifestages: Importance for skin health and beauty. Plast. Aesthetic Res. 2021, 8, 22–24. [Google Scholar] [CrossRef]
- Nolte, S.V.; Xu, W.; Rennekampff, H.O.; Rodemann, H.P. Diversity of fibroblasts—A review on implications for skin tissue engineering. Cells Tissues Organs 2008, 187, 165–176. [Google Scholar] [CrossRef]
- San Antonio, J.D.; Jacenko, O.; Fertala, A.; Orgel, J.P.R.O. Collagen Structure-Function Mapping Informs Applications for Regenerative Medicine. Bioengineering 2020, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Sayed, K.S.; Mohammed, F.N.; Abdel Hay, R.M.; Amr, K.S.; Alorbani, A.M. Cyclooxygenase-2 Gene Polymorphisms -765G>C and -1195A>G and Mycosis Fungoides Risk. Dermatology 2021, 237, 17–21. [Google Scholar] [CrossRef]
- He, J.; Li, J.; Liu, H.; Yang, Z.; Zhou, F.; Wei, T.; Dong, Y.; Xue, H.; Tang, L.; Liu, M. Scandoside exerts anti-inflammatory effect via suppressing NF-κB and MAPK signaling pathways in LPS-induced RAW 264.7 macrophages. Int. J. Mol. Sci. 2018, 19, 457. [Google Scholar] [CrossRef]
- Avazi, D.O.; Awasum, A.C.; Hassan, A.Z.; Ayo, J.O.; Aluwong, T.; Muhammed, S.T.; Simon, A.Y.; Suleiman, M.H.; Kudi, A.C. Evaluation of levels of interleukin-6, interleukin-8 and some haematologic parameters of dogs with cutaneous wounds. Cytokine 2019, 113, 128–138. [Google Scholar] [CrossRef]
- Jiang, W.G.; Sanders, A.J.; Ruge, F.; Harding, K.G. Influence of interleukin-8 (IL-8) and IL-8 receptors on the migration of human keratinocytes, the role of PLC-γ and potential clinical implications. Exp. Ther. Med. 2012, 3, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.; Lin, E.J.; Tartar, D. Immunology of Wound Healing. Curr. Dermatol. Rep. 2018, 7, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Richmond, A. Chemokine Regulation of Neutrophil Infiltration of Skin Wounds. Adv. Wound Care 2015, 4, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Van der Vliet, A.; Janssen-Heininger, Y.M. Hydrogen peroxide as a damage signal in tissue injury and inflammation: Murderer, mediator, or messenger? J. Cell. Biochem. 2014, 115, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Park, E.J.; Jang, H.J.; Lee, S.J.; Park, C.S.; Yun, B.S.; Lee, S.W.; Rho, M.C. 1-Carbomethoxy-β-Carboline, derived from Portulaca oleracea L., Ameliorates LPS-mediated inflammatory response associated with MAPK signaling and nuclear translocation of NF-κB. Molecules 2019, 24, 4042. [Google Scholar] [CrossRef]
- Laulederkind, S.J.; Thompson-Jaeger, S.; Goorha, S.; Chen, Q.; Fu, A.; Rho, J.Y.; Ballou, L.R.; Raghow, R. Both constitutive and inducible prostaglandin H synthase affect dermal wound healing in mice. Lab. Investig. 2002, 82, 919–927. [Google Scholar] [CrossRef]
- Berman, B.; Maderal, A.; Raphael, B. Keloids and hypertrophic scars: Pathophysiology, classification, and treatment. Dermatol. Surg. 2017, 43, 3–18. [Google Scholar] [CrossRef]
- Sato, Y.; Ohshima, T.; Kondo, T. Regulatory role of endogenous interleukin-10 in cutaneous inflammatory response of murine wound healing. Bioche. Biophys. Res.Commun. 1999, 265, 194–199. [Google Scholar] [CrossRef]
- Wise, L.M.; Stuart, G.S.; Real, N.C.; Fleming, S.B.; Mercer, A.A. Orf virus IL-10 accelerates wound healing while limiting inflammation and scarring. Wound Repair Regen. 2014, 22, 356–367. [Google Scholar] [CrossRef]
- Kumar, S.; Joos, G.; Boon, L.; Ttournoy, K.; Provoost, S.; Maes, T. Role of tumor necrosis factor-α and its receptors in diesel exhaust particle-induced pulmonary inflammation. Sci. Rep. 2017, 7, 11508. [Google Scholar] [CrossRef]
- Feldmann, M. Development of anti-TNF therapy for rheumatoid arthritis. Nat. Rev. Immunol. 2002, 2, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Kankaanranta, H.; Ilmarinen, P.; Zhang, X.; Adcock, I.M.; Lahti, A.; Barnes, P.J.; Giembycz, M.A.; Lindsay, M.A.; Moilanen, E. Tumour necrosis factor-α regulates human eosinophil apoptosis via ligation of TNF-receptor 1 and balance between NF-κB and AP-1. PLoS ONE 2014, 9, 90298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, X.; Chen, H.; Wang, L.; Liang, J.; Luo, D.; Liu, Y.; Yang, H.; Li, Y.; Xie, J.; et al. Anti-inflammatory activity of beta-patchoulene isolated from patchouli oil in mice. Eur. J. Pharmacol. 2016, 781, 229–238. [Google Scholar] [CrossRef]
- Kalliolias, G.D.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Beserra, F.P.; Gushiken, L.F.S.; Vieira, A.J.; Bérgamo, D.A.; Bérgamo, P.L.; Souza, M.O.; Hussni, C.A.; Takahira, R.K.; Nóbrega, R.H.; Martinez, E.R.M.; et al. From Inflammation to Cutaneous Repair: Topical Application of Lupeol Improves Skin Wound Healing in Rats by Modulating the Cytokine Levels, NF-κB, Ki-67, Growth Factor Expression, and Distribution of Collagen Fibers. Int. J. Mol. Sci. 2020, 21, 4952. [Google Scholar] [CrossRef]
- Vitali, R.; Palone, F.; Cucchiara, S.; Negroni, A.; Cavone, L.; Costanzo, M.; Aloi, M.; Dilillo, A.; Stronati, L. Dipotassium Glycyrrhizate Inhibits HMGB1-Dependent Inflammation and Ameliorates Colitis in Mice. PLoS ONE 2013, 6, 66527. [Google Scholar] [CrossRef]
- Lv, T.; Wu, D.; Song, N.; Zhang, S.; He, S.; Wang, J.; Ding, Y.; Zhang, W.; Cao, Y. Dipotassium glycyrrhizinate relieves leptospira-induced nephritis in vitro and in vivo. Microb. Pathog. 2021, 152, 104770. [Google Scholar] [CrossRef]
- Shinozaki, M.; Okada, Y.; Kitano, A.; Ikeda, K.; Saika, S.; Shinozaki, M. Impaired cutaneous wound healing with excess granulation tissue formation in TNFalpha-null mice. Arch. Dermatol. Res. 2009, 301, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Xu, Y.; Tan, G.; Han, Y.; Tang, Z.; Xu, W.; Zeng, F.; Guo, Q. Glycyrrhizin ameliorates imiquimod-induced psoriasis-like skin lesions in BALB/c mice and inhibits TNF-α-induced ICAM-1 expression via NF-κB/MAPK in HaCaT cells. Cell Physiol. Biochem. 2015, 4, 1335–1346. [Google Scholar] [CrossRef]
- Ambrozova, N.; Ulrichova, J.; Galandakova, A. Models for the Study of Skin Wound Healing. The Role of Nrf2 and NF-κB. Biomedical Papers of the Medical Faculty of Palacky University in Olomouc; Palacky University in Olomouc: Olomouc, Czech Republic, 2017; Volume 161, pp. 1–13. [Google Scholar] [CrossRef]
- Bagaev, A.V.; Garaeva, A.Y.; Lebedeva, E.S.; Pichugin, A.V.; Ataullakhanov, R.I.; Ataullakhanov, F.I. Elevated pre-activation basal level of nuclear NF-κB in native macrophages accelerates LPS-induced translocation of cytosolic NF-κB into the cell nucleus. Sci. Rep. 2019, 9, 4563. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef]
- Wullaert, A.; Bonnet, M.C.; Pasparakis, M. NF-κB in the regulation of epithelial homeostasis and inflammation. Cell Res. 2011, 21, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Gao, Z.; Lu, J.; Wang, T.; Wang, W.; Fan, L.; Xi, J.; Han, B. Ferrous iron-induced formation of glycyrrhizic acid hydrogels for Staphylococcus aureus-infected wound healing. Colloids Surf. B. Biointerfaces 2023, 221, 112977. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Jia, S.; Xie, P.; Zhong, A.; Galiano, R.D.; Mustoe, T.A.; Hong, S.J. The expression of proinflammatory genes in epidermal keratinocytes is regulated by hydration status. J. Investig. Dermatol. 2014, 134, 1044–1055. [Google Scholar] [CrossRef]
- Acheva, A.; Schettino, G.; Prise, K.M. Pro-inflammatory Signaling in a 3D Organotypic Skin Model after Low LET Irradiation-NF-κB, COX-2 Activation, and Impact on Cell Differentiation. Front. Immunol. 2017, 10, 82. [Google Scholar] [CrossRef]
- Abbas, S.; Alam, S.; Pal, A.; Kumar, M.; Singh, D.; Ansari, K.M. UVB exposure enhanced benzanthrone-induced inflammatory responses in SKH-1 mouse skin by activating the expression of COX-2 and iNOS through MAP kinases/NF-κB/AP-1 signalling pathways. Food Chem. Toxicol. 2016, 96, 183–190. [Google Scholar] [CrossRef]
- Hur, S.; Lee, Y.S.; Yoo, H.; Yang, J.H.; Kim, T.Y. Homoisoflavanone inhibits UVB-induced skin inflammation through reduced cyclooxygenase-2 expression and NF-kappaB nuclear localization. J. Dermatol. Sci. 2010, 59, 163–169. [Google Scholar] [CrossRef]
- Futagami, A.; Ishizaki, M.; Fukuda, Y.; Kawana, S.; Yamanaka, N. Wound healing involves induction of cyclooxygenase-2 expression in rat skin. Lab. Investig. 2002, 82, 1503–1513. [Google Scholar] [CrossRef]
- Meizarini, A.; Aryati; Riawan, W.; Puteri, A. Anti-inflammatory properties of a wound dressing combination of zinc oxide and turmeric extract. Vet. World 2018, 11, 25–29. [Google Scholar] [CrossRef]
- Romana-Souza, B.; Santos, J.S.; Bandeira, L.G.; Monte-Alto-Costa, A. Selective inhibition of COX-2 improves cutaneous wound healing of pressure ulcers in mice through reduction of iNOS expression. Life Sci. 2016, 15, 82–92. [Google Scholar] [CrossRef]
- Goren, I.; Lee, S.Y.; Maucher, D.; Nüsing, R.; Schlich, T.; Pfeilschifter, J.; Frank, S. Inhibition of cyclooxygenase-1 and -2 activity in keratinocytes inhibits PGE2 formation and impairs vascular endothelial growth factor release and neovascularisation in skin wounds. Int. Wound J. 2017, 14, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Vabeiryureilai, M.; Lalrinzuali, K.; Jagetia, G.C. NF-κB and COX-2 repression with topical application of hesperidin and naringin hydrogels augments repair and regeneration of deep dermal wounds. Burns 2022, 48, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhao, Y.; Simonenko, V.; Xu, J.J.; Liu, K.; Wang, D.; Shi, J.; Zhong, T.; Zhang, L.; Zeng, L.; et al. Simultaneous silencing of TGF-β1 and COX-2 reduces human skin hypertrophic scar through activation of fibroblast apoptosis. Oncotarget 2017, 8, 80651–80665. [Google Scholar] [CrossRef] [PubMed]
- Stratton, R.; Shiwen, X. Role of prostaglandins in fibroblast activation and fibrosis. J. Cell Commun. Signal. 2010, 4, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Brat, D.J.; Bellail, A.C.; Van Meir, E.G. The role of interleukin-8 and its receptors in gliomagenesis and tumoral angiogenesis. Neuro-Oncol 2005, 7, 122–133. [Google Scholar] [CrossRef]
- Zinkernagel, A.S.; Timmer, A.M.; Pence, M.A.; Locke, J.B.; Buchanan, J.T.; Turner, C.E.; Mishalian, I.; Sriskandan, S.; Hanski, E.; Nizet, V. The IL-8 protease SpyCEP/ScpC of group A Streptococcus promotes resistance to neutrophil killing. Cell Host. Microbe 2008, 4, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, K.; Yang, D.; Oppenheim, J.J. Interleukin-8: An evolving chemokine. Cytokine 2022, 153, 155828. [Google Scholar] [CrossRef] [PubMed]
- Basso, F.G.; Pansani, T.N.; Turrioni, A.P.; Soares, D.G.; De Souza Costa, C.A.; Hebling, J. Tumor Necrosis Factor-α and Interleukin (IL)-1β, IL-6, and IL-8 Impair In Vitro Migration and Induce Apoptosis of Gingival Fibroblasts and Epithelial Cells, Delaying Wound Healing. J. Periodontol. 2016, 87, 990–996. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold. Spring. Harb. Perspect. Biol. 2014, 6, 016295. [Google Scholar] [CrossRef]
- Akdis, M.; Aab, A.; Altunbulakli, C.; Azkur, K.; Costa, R.A.; Crameri, R.; Duan, S.; Eiwegger, T.; Eljaszewicz, A.; Ferstl, R.; et al. Interleukins (from IL-1 to IL-38), interferons, transforming growth factor β, and TNF-α: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2016, 138, 984–1010. [Google Scholar] [CrossRef] [Green Version]
- Bandara, M.; Arun, S.J.; Allanson, M.; Widyarini, S.; Chai, Z.; Reeve, V.E. Topical isoflavonoids reduce experimental cutaneous inflammation in mice. Immunol. Cell Biol. 2010, 88, 727–733. [Google Scholar] [CrossRef]
- Jain, A.; Kaczanowska, S.; Davila, E. IL-1 Receptor-Associated Kinase Signaling and Its Role in Inflammation, Cancer Progression, and Therapy Resistance. Front. Immunol. 2014, 17, 553. [Google Scholar] [CrossRef]
- Ringwood, L.; Li, L. The involvement of the interleukin-1 receptor-associated kinases (IRAKs) in cellular signaling networks controlling inflammation. Cytokine 2008, 42, 1–7. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, W.; Xu, L.; Zhang, H.; Liu, Q.; Yin, W. Identification of a Constitutively Active Mutant Mouse IRAK2 by Retroviral Expression Screening. Mol. Biotechnol. 2018, 60, 245–250. [Google Scholar] [CrossRef]
- Conner, J.R.; Smirnova, I.I.; Poltorak, A. A mutation in Irak2c identifies IRAK-2 as a central component of the TLR regulatory network of wild-derived mice. J. Exp. Med. 2009, 206, 1615–1631. [Google Scholar] [CrossRef]
- Shao, S.; Tsoi, L.C.; Swindell, W.R.; Chen, J.; Uppala, R.; Billi, A.C.; Xing, X.; Zeng, C.; Sarkar, M.K.; Wasikowski, R.; et al. IRAK2 Has a Critical Role in Promoting Feed-Forward Amplification of Epidermal Inflammatory Responses. J. Investig. Dermatol. 2021, 141, 2436–2448. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Nograles, K.E.; Krueger, J.G. Contrasting pathogenesis of atopic dermatitis and psoriasis--part I: Clinical and pathologic concepts. J. Allergy Clin. Immunol. 2011, 127, 1110–1118. [Google Scholar] [CrossRef]
- Kawagoe, T.; Sato, S.; Matsushita, K.; Kato, H.; Matsui, K.; Kumagai, Y.; Saitoh, T.; Kawai, T.; Takeuchi, O.; Akira, S. Sequential control of toll-like receptor-dependent responses by IRAK1 and IRAK2. Nat. Immunol. 2008, 9, 684–691. [Google Scholar] [CrossRef]
- Wan, Y.; Xiao, H.; Affolter, J.; Kim, T.W.; Bulek, K.; Chaudhuri, S.; Carlson, D.; Hamilton, T.; Mazumder, B.; Stark, G.R.; et al. Interleukin-1 receptor-associated kinase 2 is critical for lipopolysaccharide-mediated post-transcriptional control. J. Biol. Chem. 2009, 284, 10367–10375. [Google Scholar] [CrossRef]
- Cavalli, G.; Colafrancesco, S.; Emmi, E.; Imazio, M.; Lopalco, G.; Maggio, M.C.; Sota, J.; Dinarello, C.A. Interleukin 1α: A comprehensive review on the role of IL-1α in the pathogenesis and treatment of autoimmune and inflammatory diseases. Autoimmun. Rev. 2021, 20, 102763. [Google Scholar] [CrossRef]
- Yazdi, A.S.; Goreschi, K. The Interleukin-1 Family. Adv. Exp. Med. Biol. 2016, 941, 21–29. [Google Scholar] [CrossRef]
- Singh, A.K.; Fechtner, S.; Chourasia, M.; Sicalo, J.; Ahmed, S. Critical role of IL-1α in IL-1β-induced inflammatory responses: Cooperation with NF-κBp65 in transcriptional regulation. FASEB J. 2019, 33, 2526–2536. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the cross- roads of cell signaling and inflammatory dis- ease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Ogata, S.; Kubota, Y.; Yamashiro, T.; Takeuchi, H.; Ninomiya, T.; Suyama, Y.; Shirasuna, K. Signaling pathways regulating IL-1alpha-induced COX-2 expression. J. Dent. Res. 2007, 86, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Bi, Z.G. UVB-irradiated human keratinocytes and interleukin-1alpha indirectly increase MAP kinase/AP-1 activation and MMP-1 production in UVA-irradiated dermal fibroblasts. Chin. Med. J. 2006, 119, 827–831. [Google Scholar] [CrossRef]
- Jia, Y.; Chen, X.; Sun, J. Apremilast ameliorates IL-1α-induced dysfunction in epidermal stem cells. Aging 2021, 13, 19293–19305. [Google Scholar] [CrossRef]
- El Ayadi, A.; Jay, J.W.; Prasai, A. Current Approaches Targeting the Wound Healing Phases to Attenuate Fibrosis and Scarring. Int. J. Mol. Sci. 2020, 21, 1105. [Google Scholar] [CrossRef]
- He, L.; Marneros, A.G. Macrophages are essential for the early wound healing response and the formation of a fibrovascular scar. Am. J. Pathol. 2013, 2407–2417. [Google Scholar] [CrossRef]
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat. Immunol. 2011, 12, 1035–1044. [Google Scholar] [CrossRef]
- Yanez, D.A.; Lacher, R.K.; Vidyarthi, A.; Colegio, O.R. The role of macrophages in skin homeostasis. Pflügers Arch. Eur. J. Physiol. 2017, 469, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Coradin, T.; Hélary, C. Modulating inflammation in a cutaneous chronic wound model by IL-10 released from collagen– silica nanocomposites via gene delivery. Biomater. Sci. 2018, 6, 398–406. [Google Scholar] [CrossRef]
- Willenborg, S.; Lucas, T.; Van Loo, G.; Knipper, J.A.; Krieg, T.; Haase, I.; Brachvogel, B.; Hammerschmidt, M.; Nagy, A.; Ferrara, N.; et al. CCR2 recruits an inflammatory macrophage subpopulation critical for angiogenesis in tissue repair. Blood 2012, 120, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Harsono, A.D.; Prasetyono, T.O.H.; Dilogo, I.H. The Role of Interleukin 10 in Keloid Therapy: A Literature Review. Ann. Plast. Surg. 2022, 88, 617–621. [Google Scholar] [CrossRef]
- Balaji, S.; Wang, X.; King, A.; Le, L.D.; Bhattacharya, S.S.; Moles, C.M.; Butte, M.J.; De Jesus Perez, V.A.; Liechty, K.W.; Wight, T.N.; et al. Interleukin-10-mediated regenerative postnatal tissue repair is dependent on regulation of hyaluronan metabolism via fibroblast-specific STAT3 signaling. FASEB J. 2017, 31, 868–881. [Google Scholar] [CrossRef] [PubMed]
- Singampalli, K.L.; Balaji, S.; Wang, X.; Parikh, U.M.; Kaul, A.; Gilley, J.; Birla, R.K.; Bollyky, P.L.; Keswani, S.G. The Role of an IL-10/Hyaluronan Axis in Dermal Wound Healing. Front. Cell Dev. Biol. 2020, 7, 636. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Shi, S.; Xie, W.; Zhao, M.; Li, Y.; Zhang, J.; Li, N.; Bai, X.; Cai, W.; Hu, X.; et al. IL-10 alleviates lipopolysaccharide-induced skin scarring via IL-10R/STAT3 axis regulating TLR4/NF-κB pathway in dermal fibroblasts. J. Cell. Mol. Med. 2021, 25, 1554–1567. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Suşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar] [PubMed]
- Shibuya, M. VEGF-VEGFR System as a Target for Suppressing Inflammation and other Diseases. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 135–144. [Google Scholar] [CrossRef]
- Nogami, M.; Hoshi, T.; Kinoshita, M.; Arai, T.; Takama, M.; Takahashi, I. Vascular endothelial growth factor expression in rat skin incision wound. Med. Mol. Morphol. 2007, 40, 82–87. [Google Scholar] [CrossRef]
- Brown, L.F.; Yeo, K.T.; Berse, B.; Yeo, T.K.; Senger, D.R.; Dvorak, H.F.; Van de Water, L. Expression of vascular permeability factor (vascular endothelial growth factor) by epidermal keratinocytes during wound healing. J. Exp. Med. 1992, 176, 1375–1379. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, M.; Sricholpech, M.; Terajima, M.; Tomer, K.B.; Perdivara, I. Glycosylation of Type I Collagen. Methods Mol. Biol. 2019, 1934, 127–144. [Google Scholar] [CrossRef]
- Rittié, L. Type I Collagen Purification from Rat Tail Tendons. Methods Mol. Biol. 2017, 1627, 287–308. [Google Scholar] [CrossRef]
- Varma, S.; Orgel, J.P.; Schieber, J.D. Nanomechanics of Type I Collagen. Biophys. J. 2016, 111, 50–56. [Google Scholar] [CrossRef]
- Wang, T.; Gu, Q.; Zhao, J.; Mei, J.; Shao, M.; Pan, Y.; Zhang, J.; Wu, H.; Zhang, Z.; Liu, F. Calcium alginate enhances wound healing by up-regulating the ratio of collagen types I/III in diabetic rats. Int. J. Clin. Exp. Pathol. 2015, 8, 6636–6645. [Google Scholar] [PubMed]
- Asfour, H.Z.; Alhakamy, N.A.; Ahmed, O.A.A.; Fahmy, U.A.; El-Moselhy, M.A.; Rizg, W.Y.; Alghaith, A.F.; Eid, B.G.; Abdel-Naim, A.B. Amitriptyline-Based Biodegradable PEG-PLGA Self-Assembled Nanoparticles Accelerate Cutaneous Wound Healing in Diabetic Rats. Pharmaceutics 2022, 14, 1792. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.Y.; Chen, Z.P.; Hong, Z.P.; Zhang, L.; Liang, X.; Li, Y.; Duan, X.; Luo, H.; Peng, J.; Guo, J. Injectable thermo-sensitive and wide-crack self-healing hydrogel loaded with antibacterial anti-inflammatory dipotassium glycyrrhizate for full-thickness skin wound repair. Acta Biomater. 2022, 143, 203–215. [Google Scholar] [CrossRef]
- Wang, Z.C.; Zhao, W.Y.; Cao, Y.; Liu, Y.Q.; Sun, Q.; Shi, P.; Cai, J.Q.; Shen, X.Z.; Tan, W.Q. The Roles of Inflammation in Keloid and Hypertrophic Scars. Front. Immunol. 2020, 4, 603187. [Google Scholar] [CrossRef]
- Ogawa, R. Keloid and Hypertrophic Scars Are the Result of Chronic Inflammation in the Reticular Dermis. Int. J. Mol. Sci. 2017, 18, 606. [Google Scholar] [CrossRef]
- Syed, F.; Ahmadi, E.; Iqbal, S.A.; Singh, S.; McGrouther, D.A.; Bayat, A. Fibroblasts from the growing margin of keloid scars produce higher levels of collagen I and III compared with intralesional and extralesional sites: Clinical implications for lesional site-directed therapy. Br. J. Dermatol. 2011, 1, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.V.; Hawkins, H.K.; Chinkes, D.; Burke, A.; Tavares, A.L.; Ramos-e-Silva, M.; Albrecht, T.B.; Kitten, G.T.; Herndon, D.N. Hypertrophic versus non hypertrophic scars compared by immunohistochemistry and laser confocal microscopy: Type I and III collagens. Int. Wound J. 2009, 6, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Peseto, D.C.; Carmona, E.V.; Silva, K.C.; Guedes, F.R.V.; Hummel Filho, F.; Martinez, N.P.; Pereira, J.A.; Rocha, T.; Priolli, D.G. Effects of tretinoin on wound healing in aged skin. Wound Rep. Regen. 2016, 24, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.V.P. Eutanásia. In Animais de Laboratório: Criação e Experimentação; Andrade, A., Pinto, S.C., Oliveira, R.S., Eds.; Fiocruz: Rio de Janeiro, Brazil, 2002; pp. 275–279. [Google Scholar]
- Alves Junior, A.J.T.; Pereira, J.A.; Ávila, M.G.; Domingues, F.A.; Sato, D.T.; Martinez, C.A.R. Tissue content of metalloproteinase-9 and collagen in the colon with and without fecal stream after intervention with infliximab in rats subjected to Hartmann’s surgery. Acta Cir. Bras. 2021, 36, 360401. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | % | Function | INCI Name | CAS Number |
|---|---|---|---|---|
| Aqua | 93.600 | Solvent | Aqua | 7732-18-5 |
| Sepigel 305 | 4.000 | Viscosity controlling | Polyacrylamide, C13-14 Isoparaffin (and) Laureth 7 | 9003-05-8/246538-79-4/68439-50-9/9002-92-0/7732-18-5 |
| Dipotassium Glycyrrhizinate | 2.000 | Active | Dipotassium Glycyrrhizinate | 68797-35-3 |
| Euxyl PE 9010 | 0.400 | Preservative | Phenoxyethanol (and) Ethylhexylglycerin | 122-99-6/70445-33-9 |
| Total | 100.000 |
| Degree | Score | Inflammatory Exudate | Active Hyperemia |
|---|---|---|---|
| Absent | 0 | No leukocyte tissue infiltration (0%) | No tissue hyperemia (0%) |
| Light | 1 | Leukocyte infiltration (<50%) | Presence of hyperemia (<50%) |
| Moderate | 2 | Leukocyte infiltration (≥51%) | Presence of hyperemia (≥51%) |
| Intense | 3 | Abundant leukocyte infiltration (≥75%) | Abundant hyperemia (≥75%) |
| Degree | Score | Granulation Tissue | Re-Epithelialization |
|---|---|---|---|
| Absent | 0 | Neoformed tissue absent (0%) | Re-epithelialization tissue absent (0%) |
| Light | 1 | Presence of neoformed tissue (<50%) | Presence of re-epithelialization tissue (<50%) |
| Moderate | 2 | Presence of neoformed tissue (≥51%) | Presence of re-epithelialization tissue (≥51%) |
| Intense | 3 | Abundant neoformed tissue (≥75%) | Presence of re-epithelialization tissue (≥75%) |
| Gene | Primers Sequence 5′-3′ |
|---|---|
| Cox-2 | FW: AACAACATTCCCTTCCTTCG RV: AAGTTGGTGGGCTGTCAATC |
| Tnf-α | FW: GGGCTCCCTCTCATCAGTT RV: TTGCTACGACGTGGGCTAC |
| Nf-kb | FW: CAGCTCTTCTCAAAGCAGCA RV: AGCCTTCTCCCAAGAGTCG |
| Il-1α | FW: GGCCATAGCCCATGATTTAG RV: TGATGAACTCCTGCTTGACG |
| Irak-2 | FW: TCAAGAGGCTCAGGGAGGT RV: CCCAGCAGAGGTAGGATGTT |
| Il-8rb | FW: ATCTTTGCTGTGGTCCTCGT RV: GGTCTCCTTGATCAGCTTGG |
| Il-10 | FW: AGCCTTGCAGAAAACAGAGC RV: GCCTTTGCTGGTCTTCACTC |
| Vegf | FW: CGGAGAGCAACGTCACTATG RV: GCTGCAGGAAGCTCATCTCT |
| Col-1 | FW: GGAATGAAGGGACACAGAGG RV: AGGCTCTCCCTTAGGACCAG |
| 18s | FW: CGCGGTTCTATTTTGTTGGT RV: CGGTCCAAGAATTTCACCTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leite, C.d.S.; Bonafé, G.A.; Pires, O.C.; Santos, T.W.d.; Pereira, G.P.; Pereira, J.A.; Rocha, T.; Martinez, C.A.R.; Ortega, M.M.; Ribeiro, M.L. Dipotassium Glycyrrhizininate Improves Skin Wound Healing by Modulating Inflammatory Process. Int. J. Mol. Sci. 2023, 24, 3839. https://doi.org/10.3390/ijms24043839
Leite CdS, Bonafé GA, Pires OC, Santos TWd, Pereira GP, Pereira JA, Rocha T, Martinez CAR, Ortega MM, Ribeiro ML. Dipotassium Glycyrrhizininate Improves Skin Wound Healing by Modulating Inflammatory Process. International Journal of Molecular Sciences. 2023; 24(4):3839. https://doi.org/10.3390/ijms24043839
Chicago/Turabian StyleLeite, Camila dos Santos, Gabriel Alves Bonafé, Oscar César Pires, Tanila Wood dos Santos, Geovanna Pacciulli Pereira, José Aires Pereira, Thalita Rocha, Carlos Augusto Real Martinez, Manoela Marques Ortega, and Marcelo Lima Ribeiro. 2023. "Dipotassium Glycyrrhizininate Improves Skin Wound Healing by Modulating Inflammatory Process" International Journal of Molecular Sciences 24, no. 4: 3839. https://doi.org/10.3390/ijms24043839
APA StyleLeite, C. d. S., Bonafé, G. A., Pires, O. C., Santos, T. W. d., Pereira, G. P., Pereira, J. A., Rocha, T., Martinez, C. A. R., Ortega, M. M., & Ribeiro, M. L. (2023). Dipotassium Glycyrrhizininate Improves Skin Wound Healing by Modulating Inflammatory Process. International Journal of Molecular Sciences, 24(4), 3839. https://doi.org/10.3390/ijms24043839