
What’s on the Other Side of the Gate: A Structural Perspective on DNA Gate Opening of Type IA and IIA DNA Topoisomerases


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
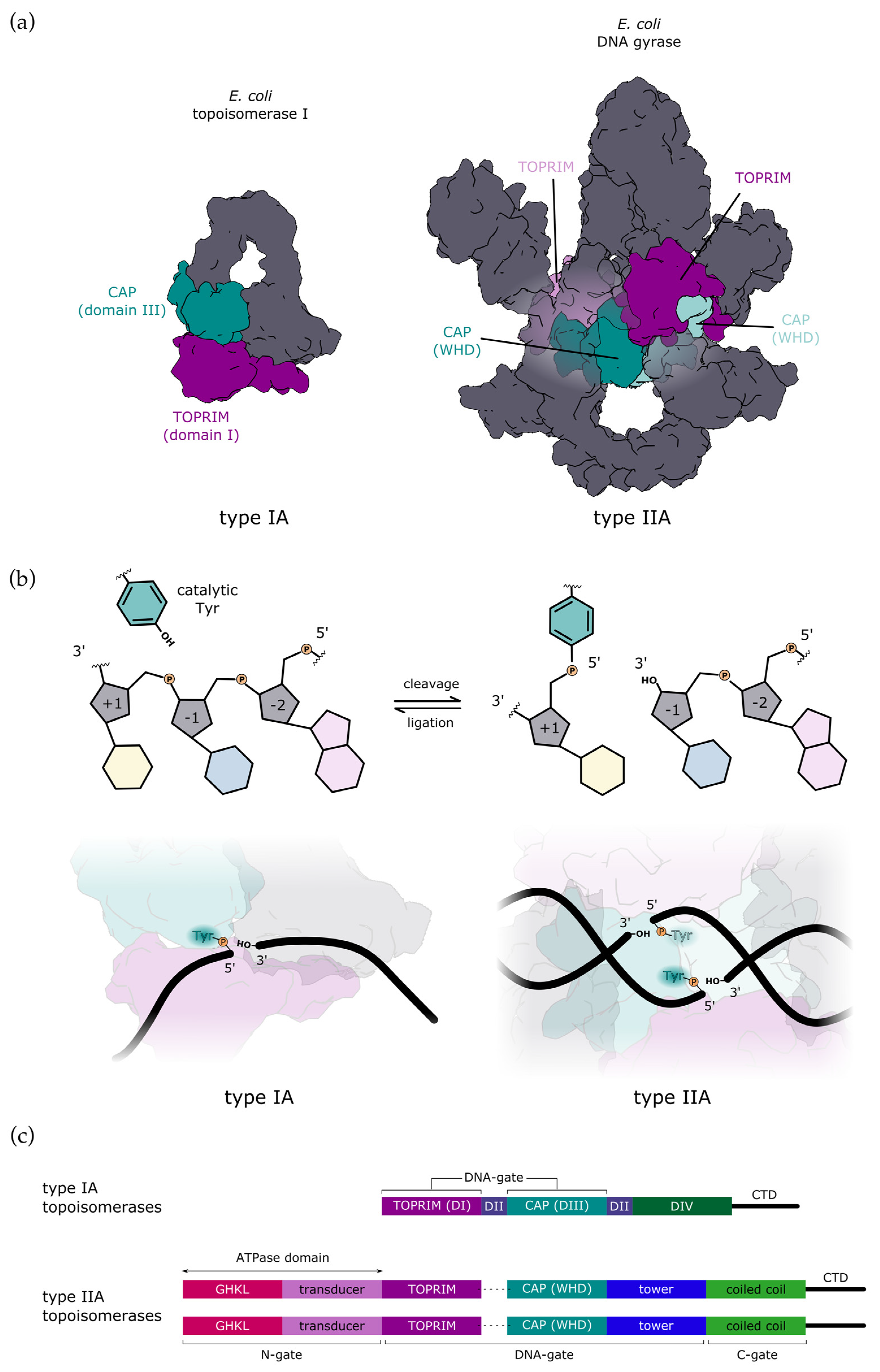
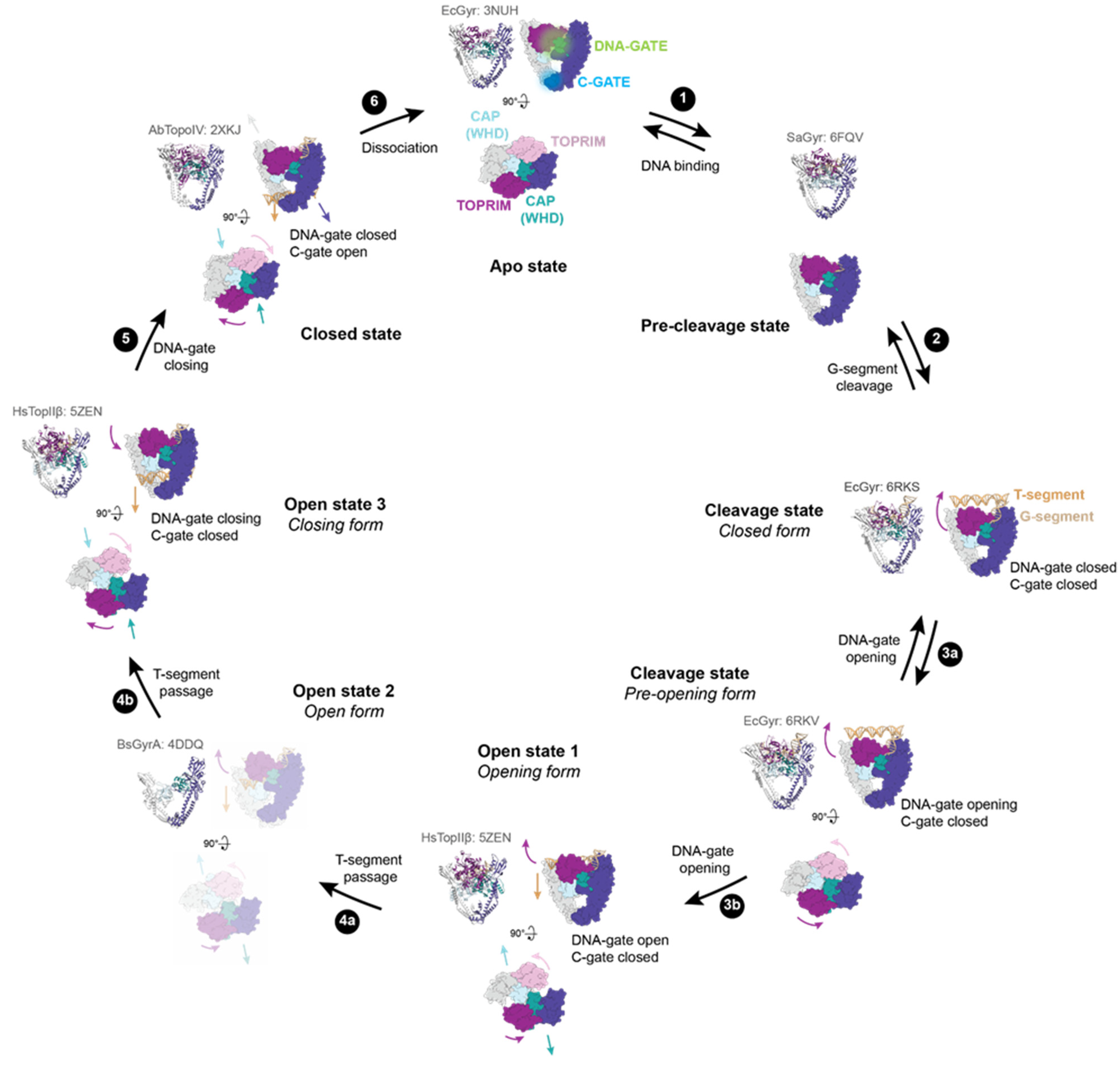
2. Mechanism of DNA-Gate Opening by Type IIA DNA Topoisomerases
2.1. Structural Rearrangements Leading to DNA-Gate Opening in Type IIA Topoisomerases

2.2. Regulation of DNA Gate Opening of Type IIA Topoisomerases
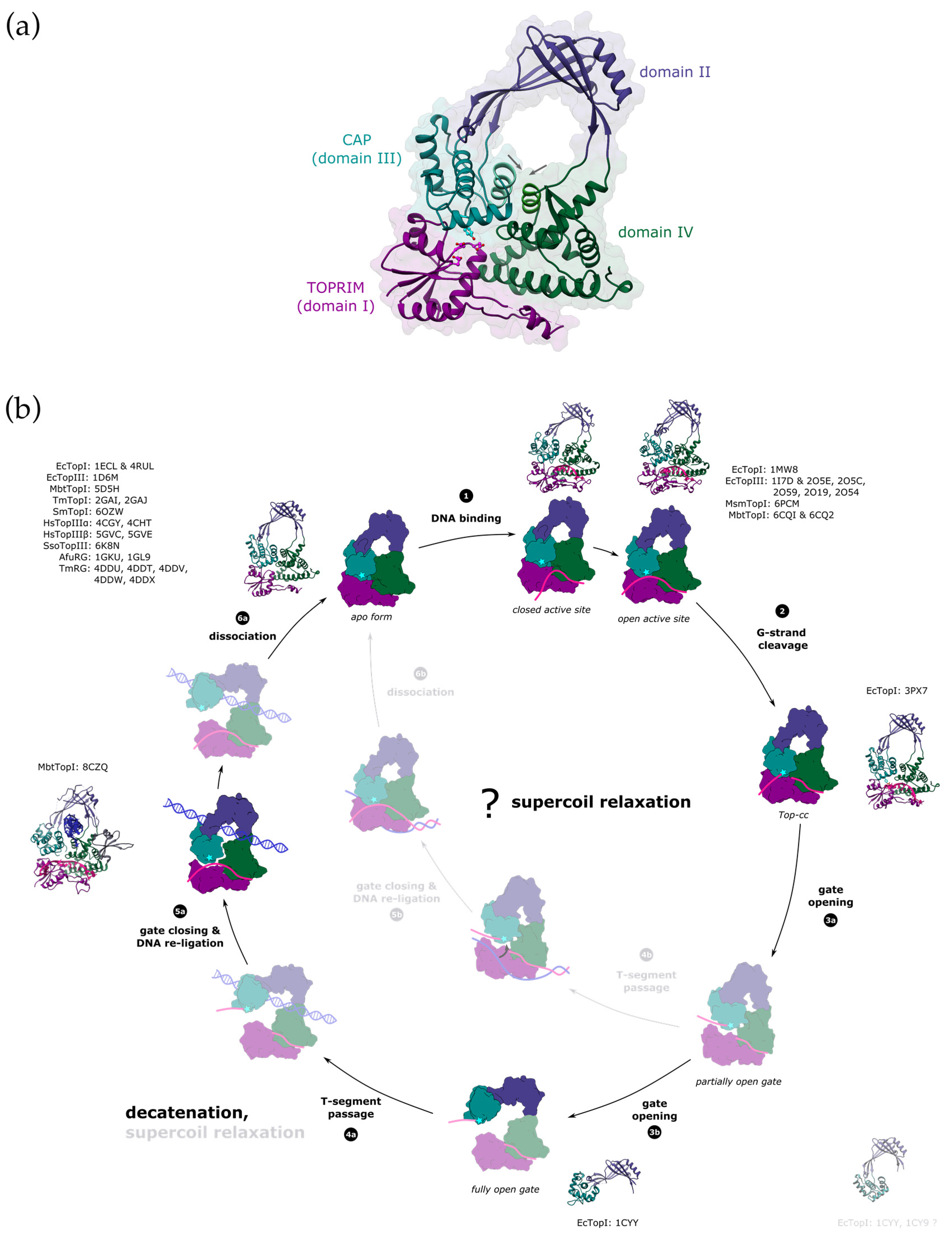
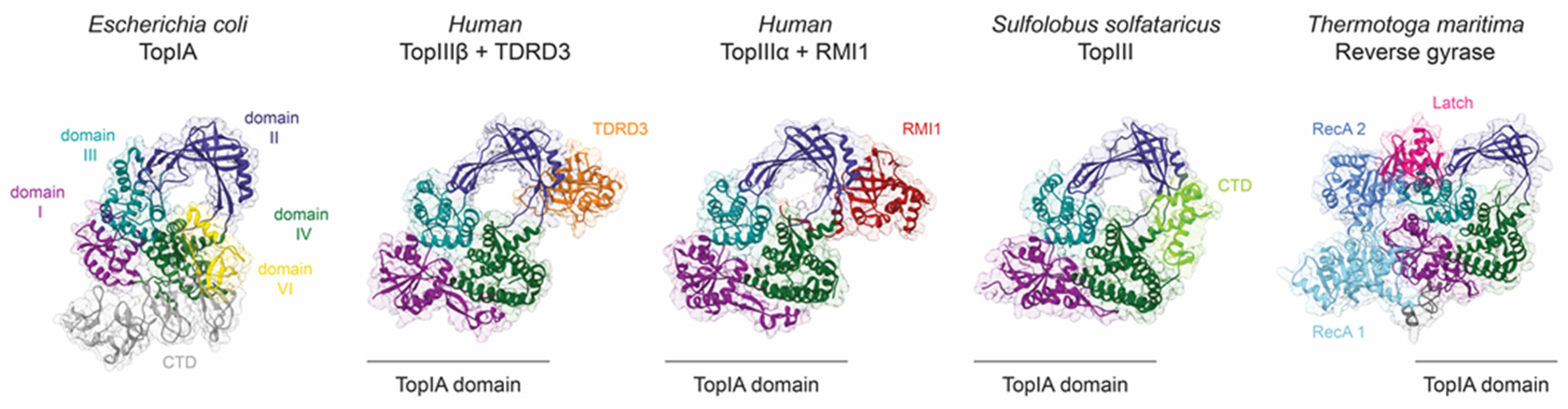
3. Mechanism of DNA Gate Opening by Type IA DNA Topoisomerases
3.1. Structural Mechanisms Leading to DNA-Gate Opening in Type IA Topoisomerases
3.2. Structural Regulation of DNA Gate Opening and Strand Passage in Type IA Topoisomerases

4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKie, S.J.; Neuman, K.C.; Maxwell, A. DNA Topoisomerases: Advances in Understanding of Cellular Roles and Multi-Protein Complexes via Structure-Function Analysis. BioEssays 2021, 43, 2000286. [Google Scholar] [CrossRef]
- Maxwell, A.; Bush, N.G.; Evans-Roberts, K. DNA Topoisomerases. EcoSal Plus 2015, 6. [Google Scholar] [CrossRef]
- Tse, Y.C.; Kirkegaard, K.; Wang, J.C. Covalent Bonds between Protein and DNA. Formation of Phosphotyrosine Linkage between Certain DNA Topoisomerases and DNA. J. Biol. Chem. 1980, 255, 5560–5565. [Google Scholar] [CrossRef]
- Schoeffler, A.J.; Berger, J.M. DNA Topoisomerases: Harnessing and Constraining Energy to Govern Chromosome Topology. Q. Rev. Biophys. 2008, 41, 41–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C. Interaction between DNA and an Escherichia Coli Protein ω. J. Mol. Biol. 1971, 55, 523-IN16. [Google Scholar] [CrossRef] [PubMed]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Nash, H.A. DNA Gyrase: An Enzyme That Introduces Superhelical Turns into DNA. Proc. Natl. Acad. Sci. USA 1976, 73, 3872–3876. [Google Scholar] [CrossRef] [Green Version]
- DiGate, R.J.; Marians, K.J. Molecular Cloning and DNA Sequence Analysis of Escherichia Coli TopB, the Gene Encoding Topoisomerase III. J. Biol. Chem. 1989, 264, 17924–17930. [Google Scholar]
- DiGate, R.J.; Marians, K.J. Escherichia Coli Topoisomerase III-Catalyzed Cleavage of RNA. J. Biol. Chem. 1992, 267, 20532–20535. [Google Scholar]
- LIU, L.; LIU, C.; ALBERTS, B. Type II DNA Topoisomerases: Enzymes That Can Unknot a Topologically Knotted DNA Molecule via a Reversible Double-Strand Break. Cell 1980, 19, 697–707. [Google Scholar] [CrossRef]
- Kreuzer, K.N.; Cozzarelli, N.R. Formation and Resolution of DNA Catenanes by DNA Gyrase. Cell 1980, 20, 245–254. [Google Scholar] [CrossRef]
- Schmidt, B.H.; Burgin, A.B.; Deweese, J.E.; Osheroff, N.; Berger, J.M. A Novel and Unified Two-Metal Mechanism for DNA Cleavage by Type II and IA Topoisomerases. Nature 2010, 465, 641–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, M.D. Structure and Function of an Archaeal Topoisomerase VI Subunit with Homology to the Meiotic Recombination Factor Spo11. EMBO J. 1999, 18, 6177–6188. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.D. Structure of the Topoisomerase VI-B Subunit: Implications for Type II Topoisomerase Mechanism and Evolution. EMBO J. 2003, 22, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, K.D.; Benedetti, P.; Berger, J.M. Holoenzyme Assembly and ATP-Mediated Conformational Dynamics of Topoisomerase VI. Nat. Struct. Mol. Biol. 2007, 14, 611–619. [Google Scholar] [CrossRef]
- Vrielynck, N.; Chambon, A.; Vezon, D.; Pereira, L.; Chelysheva, L.; De Muyt, A.; Mézard, C.; Mayer, C.; Grelon, M. A DNA Topoisomerase VI–like Complex Initiates Meiotic Recombination. Science 2016, 351, 939–943. [Google Scholar] [CrossRef]
- Claeys Bouuaert, C.; Tischfield, S.E.; Pu, S.; Mimitou, E.P.; Arias-Palomo, E.; Berger, J.M.; Keeney, S. Structural and Functional Characterization of the Spo11 Core Complex. Nat. Struct. Mol. Biol. 2021, 28, 92–102. [Google Scholar] [CrossRef]
- McKie, S.J.; Desai, P.R.; Seol, Y.; Allen, A.M.; Maxwell, A.; Neuman, K.C. Topoisomerase VI Is a Chirally-Selective, Preferential DNA Decatenase. Elife 2022, 11, e67021. [Google Scholar] [CrossRef]
- Wigley, D.B.; Davies, G.J.; Dodson, E.J.; Maxwell, A.; Dodson, G. Crystal Structure of an N-Terminal Fragment of the DNA Gyrase B Protein. Nature 1991, 351, 624–629. [Google Scholar] [CrossRef]
- Berger, J.M.; Wang, J.C. Recent Developments in DNA Topoisomerase II Structure and Mechanism. Curr. Opin. Struct. Biol. 1996, 6, 84–90. [Google Scholar] [CrossRef]
- Aravind, L.; Leipe, D.D.; Koonin, E. V Toprim--a Conserved Catalytic Domain in Type IA and II Topoisomerases, DnaG-Type Primases, OLD Family Nucleases and RecR Proteins. Nucleic Acids Res. 1998, 26, 4205–4213. [Google Scholar] [CrossRef]
- Cabral, J.H.M.; Jackson, A.P.; Smith, C.V.; Shikotra, N.; Maxwell, A.; Liddington, R.C. Crystal Structure of the Breakage–Reunion Domain of DNA Gyrase. Nature 1997, 388, 903–906. [Google Scholar] [CrossRef]
- Roca, J.; Wang, J.C. DNA Transport by a Type II DNA Topoisomerase: Evidence in Favor of a Two-Gate Mechanism. Cell 1994, 77, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Roca, J.; Wang, J.C. The Probabilities of Supercoil Removal and Decatenation by Yeast DNA Topoisomerase II. Genes Cells 1996, 1, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.C.; Berger, J.M. Structural Basis for Gate-DNA Recognition and Bending by Type IIA Topoisomerases. Nature 2007, 450, 1201–1205. [Google Scholar] [CrossRef] [PubMed]
- Wendorff, T.J.; Schmidt, B.H.; Heslop, P.; Austin, C.A.; Berger, J.M. The Structure of DNA-Bound Human Topoisomerase II Alpha: Conformational Mechanisms for Coordinating Inter-Subunit Interactions with DNA Cleavage. J. Mol. Biol. 2012, 424, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Fass, D.; Bogden, C.E.; Berger, J.M. Quaternary Changes in Topoisomerase II May Direct Orthogonal Movement of Two DNA Strands. Nat. Struct. Biol. 1999, 6, 322–326. [Google Scholar] [CrossRef]
- Roca, J.; Wang, J.C. The Capture of a DNA Double Helix by an ATP-Dependent Protein Clamp: A Key Step in DNA Transport by Type II DNA Topoisomerases. Cell 1992, 71, 833–840. [Google Scholar] [CrossRef]
- Bjergbæk, L.; Jensen, S.; Westergaard, O.; Andersen, A.H. Using a Biochemical Approach to Identify the Primary Dimerization Regions in Human DNA Topoisomerase IIα. J. Biol. Chem. 1999, 274, 26529–26536. [Google Scholar] [CrossRef] [Green Version]
- Harkins, T.T.; Lewis, T.J.; Lindsley, J.E. Pre-Steady-State Analysis of ATP Hydrolysis by Saccharomyces Cerevisiae DNA Topoisomerase II. 2. Kinetic Mechanism for the Sequential Hydrolysis of Two ATP. Biochemistry 1998, 37, 7299–7312. [Google Scholar] [CrossRef]
- Schmidt, B.H.; Osheroff, N.; Berger, J.M. Structure of a Topoisomerase II-DNA-Nucleotide Complex Reveals a New Control Mechanism for ATPase Activity. Nat. Struct. Mol. Biol. 2012, 19, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Germe, T.; Vörös, J.; Jeannot, F.; Taillier, T.; Stavenger, R.A.; Bacqué, E.; Maxwell, A.; Bax, B.D. A New Class of Antibacterials, the Imidazopyrazinones, Reveal Structural Transitions Involved in DNA Gyrase Poisoning and Mechanisms of Resistance. Nucleic Acids Res. 2018, 46, 4114–4128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bax, B.D.; Chan, P.F.; Eggleston, D.S.; Fosberry, A.; Gentry, D.R.; Gorrec, F.; Giordano, I.; Hann, M.M.; Hennessy, A.; Hibbs, M.; et al. Type IIA Topoisomerase Inhibition by a New Class of Antibacterial Agents. Nature 2010, 466, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Wohlkonig, A.; Chan, P.F.; Fosberry, A.P.; Homes, P.; Huang, J.; Kranz, M.; Leydon, V.R.; Miles, T.J.; Pearson, N.D.; Perera, R.L.; et al. Structural Basis of Quinolone Inhibition of Type IIA Topoisomerases and Target-Mediated Resistance. Nat. Struct. Mol. Biol. 2010, 17, 1152–1153. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-F.; Huang, N.-L.; Lin, J.-H.; Wu, C.-C.; Wang, Y.-R.; Yu, Y.-J.; Gilson, M.K.; Chan, N.-L. Structural Insights into the Gating of DNA Passage by the Topoisomerase II DNA-Gate. Nat. Commun. 2018, 9, 3085. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-R.; Chen, S.-F.; Wu, C.-C.; Liao, Y.-W.; Lin, T.-S.; Liu, K.-T.; Chen, Y.-S.; Li, T.-K.; Chien, T.-C.; Chan, N.-L. Producing Irreversible Topoisomerase II-Mediated DNA Breaks by Site-Specific Pt(II)-Methionine Coordination Chemistry. Nucleic Acids Res. 2017, 45, 10920. [Google Scholar] [CrossRef] [PubMed]
- Zechiedrich, E.L.; Osheroff, N. Eukaryotic Topoisomerases Recognize Nucleic Acid Topology by Preferentially Interacting with DNA Crossovers. EMBO J. 1990, 9, 4555–4562. [Google Scholar] [CrossRef] [PubMed]
- Vologodskii, A.V.; Zhang, W.; Rybenkov, V.V.; Podtelezhnikov, A.A.; Subramanian, D.; Griffith, J.D.; Cozzarelli, N.R. Mechanism of Topology Simplification by Type II DNA Topoisomerases. Proc. Natl. Acad. Sci. USA 2001, 98, 3045–3049. [Google Scholar] [CrossRef] [Green Version]
- Vanden Broeck, A.; Lotz, C.; Ortiz, J.; Lamour, V. Cryo-EM Structure of the Complete E. Coli DNA Gyrase Nucleoprotein Complex. Nat. Commun. 2019, 10, 4935. [Google Scholar] [CrossRef] [Green Version]
- Vanden Broeck, A.; Lotz, C.; Drillien, R.; Haas, L.; Bedez, C.; Lamour, V. Structural Basis for Allosteric Regulation of Human Topoisomerase IIα. Nat. Commun. 2021, 12, 2962. [Google Scholar] [CrossRef]
- Bedez, C.; Lotz, C.; Batisse, C.; Broeck, A.V.; Stote, R.H.; Howard, E.; Pradeau-Aubreton, K.; Ruff, M.; Lamour, V. Post-Translational Modifications in DNA Topoisomerase 2α Highlight the Role of a Eukaryote-Specific Residue in the ATPase Domain. Sci. Rep. 2018, 8, 9272. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, A.C.; Hawaz, M.G.; Hoang, K.G.; Trac, J.; Keck, J.M.; Ayes, C.; Deweese, J.E. Exploration of the Role of the C-Terminal Domain of Human DNA Topoisomerase IIα in Catalytic Activity. ACS Omega 2021, 6, 25892–25903. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-J.; Yen, T.-J.; Lin, T.-S.; Chang, H.-T.; Huang, S.-Y.; Hsu, C.-H.; Farh, L.; Chan, N.-L. Twisting of the DNA-Binding Surface by a β-Strand-Bearing Proline Modulates DNA Gyrase Activity. Nucleic Acids Res. 2010, 38, 4173–4181. [Google Scholar] [CrossRef] [Green Version]
- Ruthenburg, A.J.; Graybosch, D.M.; Huetsch, J.C.; Verdine, G.L. A Superhelical Spiral in the Escherichia Coli DNA Gyrase A C-Terminal Domain Imparts Unidirectional Supercoiling Bias. J. Biol. Chem. 2005, 280, 26177–26184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, K.D.; Shultzaberger, R.K.; Berger, J.M. The C-Terminal Domain of DNA Gyrase A Adopts a DNA-Bending β-Pinwheel Fold. Proc. Natl. Acad. Sci. USA 2004, 101, 7293–7298. [Google Scholar] [CrossRef] [Green Version]
- Corbett, K.D.; Schoeffler, A.J.; Thomsen, N.D.; Berger, J.M. The Structural Basis for Substrate Specificity in DNA Topoisomerase IV. J. Mol. Biol. 2005, 351, 545–561. [Google Scholar] [CrossRef] [PubMed]
- Bouige, A.; Darmon, A.; Piton, J.; Roue, M.; Petrella, S.; Capton, E.; Forterre, P.; Aubry, A.; Mayer, C. Mycobacterium Tuberculosis DNA Gyrase Possesses Two Functional GyrA-Boxes. Biochem. J. 2013, 455, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampranis, S.C.; Maxwell, A. Conversion of DNA Gyrase into a Conventional Type II Topoisomerase. Proc. Natl. Acad. Sci. USA 1996, 93, 14416–14421. [Google Scholar] [CrossRef] [Green Version]
- Tretter, E.M.; Berger, J.M. Mechanisms for Defining Supercoiling Set Point of DNA Gyrase Orthologs: I. A Nonconserved Acidic C-Terminal Tail Modulates Escherichia Coli Gyrase Activity. J. Biol. Chem. 2012, 287, 18636–18644. [Google Scholar] [CrossRef] [Green Version]
- Tretter, E.M.; Berger, J.M. Mechanisms for Defining Supercoiling Set Point of DNA Gyrase Orthologs: II. The Shape of the GyrA Subunit C-Terminal Domain (CTD) Is Not a Sole Determinant for Controlling Supercoiling Efficiency. J. Biol. Chem. 2012, 287, 18645–18654. [Google Scholar] [CrossRef] [Green Version]
- Lanz, M.A.; Klostermeier, D. The GyrA-Box Determines the Geometry of DNA Bound to Gyrase and Couples DNA Binding to the Nucleotide Cycle. Nucleic Acids Res. 2012, 40, 10893–10903. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.D.; Wang, J.C.; Mondragón, A. Three-Dimensional Structure of the 67K N-Terminal Fragment of E. Coli DNA Topoisomerase I. Nat. 1994 3676459 1994, 367, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.; Tse-Dinh, Y.-C.; Neuman, K.C. Direct Observation of Topoisomerase IA Gate Dynamics. Nat. Struct. Mol. Biol. 2018, 25, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Gunn, K.H.; Marko, J.F.; Mondragón, A. An Orthogonal Single-Molecule Experiment Reveals Multiple-Attempt Dynamics of Type IA Topoisomerases. Nat. Struct. Mol. Biol. 2017, 24, 484–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Mondragón, A.; DiGate, R.J. The Mechanism of Type IA Topoisomerase-Mediated DNA Topological Transformations. Mol. Cell 2001, 7, 301–307. [Google Scholar] [CrossRef]
- Leelaram, M.N.; Bhat, A.G.; Godbole, A.A.; Bhat, R.S.; Manjunath, R.; Nagaraja, V. Type IA Topoisomerase Inhibition by Clamp Closure. FASEB J. 2013, 27, 3030–3038. [Google Scholar] [CrossRef]
- Ferdous, S.; Dasgupta, T.; Annamalai, T.; Tan, K.; Tse-Dinh, Y.-C. The Interaction between Transport-Segment DNA and Topoisomerase IA—Crystal Structure of MtbTOP1 in Complex with Both G- and T-Segments. Nucleic Acids Res. 2022, 51, 349–364. [Google Scholar] [CrossRef]
- Chen, L.; Huang, L. Oligonucleotide Cleavage and Rejoining by Topoisomerase III from the Hyperthermophilic Archaeon Sulfolobus Solfataricus: Temperature Dependence and Strand Annealing-Promoted DNA Religation. Mol. Microbiol. 2006, 60, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Mondragón, A.; DiGate, R. The Structure of Escherichia Coli DNA Topoisomerase III. Structure 1999, 7, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.; Cao, N.; Cheng, B.; Joachimiak, A.; Tse-Dinh, Y.-C. Insights from the Structure of Mycobacterium Tuberculosis Topoisomerase I with a Novel Protein Fold. J. Mol. Biol. 2016, 428, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changela, A.; DiGate, R.J.; Mondragón, A. Structural Studies of E. Coli Topoisomerase III-DNA Complexes Reveal A Novel Type IA Topoisomerase-DNA Conformational Intermediate. J. Mol. Biol. 2007, 368, 105. [Google Scholar] [CrossRef] [Green Version]
- Cao, N.; Tan, K.; Annamalai, T.; Joachimiak, A.; Tse-Dinh, Y.C. Investigating Mycobacterial Topoisomerase I Mechanism from the Analysis of Metal and DNA Substrate Interactions at the Active Site. Nucleic Acids Res. 2018, 46, 7296–7308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, N.; Tan, K.; Zuo, X.; Annamalai, T.; Tse-Dinh, Y.C. Mechanistic Insights from Structure of Mycobacterium Smegmatis Topoisomerase I with SsDNA Bound to Both N- and C-Terminal Domains. Nucleic Acids Res. 2020, 48, 4448. [Google Scholar] [CrossRef] [PubMed]
- Perry, K.; Mondragón, A. Structure of a Complex between E. Coli DNA Topoisomerase I and Single-Stranded DNA. Structure 2003, 11, 1349–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changela, A.; DiGate, R.J.; Mondragón, A. Crystal Structure of a Complex of a Type IA DNA Topoisomerase with a Single-Stranded DNA Molecule. Nature 2001, 411, 1077–1081. [Google Scholar] [CrossRef]
- Narula, G.; Tse-Dinh, Y.-C. Residues of E. Coli Topoisomerase I Conserved for Interaction with a Specific Cytosine Base to Facilitate DNA Cleavage. Nucleic Acids Res. 2012, 40, 9233–9243. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Saha, S.; Yang, W.; Neuman, K.C.; Pommier, Y. Structural and Biochemical Basis for DNA and RNA Catalysis by Human Topoisomerase 3β. Nat. Commun. 2022, 13, 4656. [Google Scholar] [CrossRef]
- Sutormin, D.; Galivondzhyan, A.; Musharova, O.; Travin, D.; Rusanova, A.; Obraztsova, K.; Borukhov, S.; Severinov, K. Interaction between Transcribing RNA Polymerase and Topoisomerase I Prevents R-Loop Formation in E. Coli. Nat. Commun. 2022, 13, 4524. [Google Scholar] [CrossRef]
- Zhang, Z.; Cheng, B.; Tse-Dinh, Y.-C. Crystal Structure of a Covalent Intermediate in DNA Cleavage and Rejoining by Escherichia Coli DNA Topoisomerase I. Proc. Natl. Acad. Sci. USA 2011, 108, 6939–6944. [Google Scholar] [CrossRef] [Green Version]
- Corbett, K.D.; Berger, J.M. Structure, Molecular Mechanisms, and Evolutionary Relationships in DNA Topoisomerases. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Bocquet, N.; Bizard, A.H.; Abdulrahman, W.; Larsen, N.B.; Faty, M.; Cavadini, S.; Bunker, R.D.; Kowalczykowski, S.C.; Cejka, P.; Hickson, I.D.; et al. Structural and Mechanistic Insight into Holliday Junction Dissolution by Topoisomerase IIIα and RMI1. Nat. Struct. Mol. Biol. 2014, 21, 261. [Google Scholar] [CrossRef] [Green Version]
- Goto-Ito, S.; Yamagata, A.; Takahashi, T.S.; Sato, Y.; Fukai, S. Structural Basis of the Interaction between Topoisomerase IIIβ and the TDRD3 Auxiliary Factor. Sci. Rep. 2017, 7, 42123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, P.O.; Cozzarelli, N.R. Catenation and Knotting of Duplex DNA by Type 1 Topoisomerases: A Mechanistic Parallel with Type 2 Topoisomerases. Proc. Natl. Acad. Sci. USA 1981, 78, 843–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Shen, W.; Li, W.; Xue, Y.; Zou, S.; Xu, D.; Wang, W. Topoisomerase 3β Is the Major Topoisomerase for MRNAs and Linked to Neurodevelopment and Mental Dysfunction. Nucleic Acids Res. 2016, 45, gkw1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Xue, Y.; Lee, S.K.; Martindale, J.L.; Shen, W.; Li, W.; Zou, S.; Ciaramella, M.; Debat, H.; Nadal, M.; et al. RNA Topoisomerase Is Prevalent in All Domains of Life and Associates with Polyribosomes in Animals. Nucleic Acids Res. 2016, 44, 6335–6349. [Google Scholar] [CrossRef] [Green Version]
- Cejka, P.; Plank, J.L.; Dombrowski, C.C.; Kowalczykowski, S.C. Decatenation of DNA by the S. Cerevisiae Sgs1-Top3-Rmi1 and RPA Complex: A Mechanism for Disentangling Chromosomes. Mol. Cell 2012, 47, 886–896. [Google Scholar] [CrossRef] [Green Version]
- Terekhova, K.; Marko, J.F.; Mondragón, A.; Mondragón, M. Single-Molecule Analysis Uncovers the Difference between the Kinetics of DNA Decatenation by Bacterial Topoisomerases I and III. Nucleic Acids Res. 2014, 42, 11657–11667. [Google Scholar] [CrossRef] [Green Version]
- Phoenix, P.; Raymond, M.A.; Massé, É.; Drolet, M. Roles of DNA Topoisomerases in the Regulation of R-Loop Formation in Vitro. J. Biol. Chem. 1997, 272, 1473–1479. [Google Scholar] [CrossRef] [Green Version]
- Jeanne Dit Fouque, K.; Garabedian, A.; Leng, F.; Tse-Dinh, Y.-C.; Fernandez-Lima, F. Microheterogeneity of Topoisomerase IA/IB and Their DNA-Bound States. ACS Omega 2019, 4, 3619–3626. [Google Scholar] [CrossRef]
- Cheng, B.; Feng, J.; Gadgil, S.; Tse-Dinh, Y.-C.C. Flexibility at Gly-194 Is Required for DNA Cleavage and Relaxation Activity of Escherichia Coli DNA Topoisomerase I. J. Biol. Chem. 2004, 279, 8648–8654. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.C.; Stock, D. Crystal Structure of Reverse Gyrase: Insights into the Positive Supercoiling of DNA. EMBO J. 2002, 21, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, M.G.; del Toro Duany, Y.; Jungblut, S.P.; Ganguly, A.; Klostermeier, D. Crystal Structures of Thermotoga Maritima Reverse Gyrase: Inferences for the Mechanism of Positive DNA Supercoiling. Nucleic Acids Res. 2013, 41, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Duguet, M.; Serre, M.C.; Bouthier de La Tour, C. A Universal Type IA Topoisomerase Fold. J. Mol. Biol. 2006, 359, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Bakx, J.A.M.; Biebricher, A.S.; King, G.A.; Christodoulis, P.; Sarlós, K.; Bizard, A.H.; Hickson, I.D.; Wuite, G.J.L.; Peterman, E.J.G. Duplex DNA and BLM Regulate Gate Opening by the Human TopoIIIα-RMI1-RMI2 Complex. Nat. Commun. 2022, 13, 584. [Google Scholar] [CrossRef] [PubMed]
- Soczek, K.M.; Grant, T.; Rosenthal, P.B.; Mondragón, A. CryoEM Structures of Open Dimers of Gyrase A in Complex with DNA Illuminate Mechanism of Strand Passage. Elife 2018, 7, e41215. [Google Scholar] [CrossRef]
- Tan, K.; Zhou, Q.; Cheng, B.; Zhang, Z.; Joachimiak, A.; Tse-Dinh, Y.-C.C. Structural Basis for Suppression of Hypernegative DNA Supercoiling by E. Coli Topoisomerase I. Nucleic Acids Res. 2015, 43, 11031–11046. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, J.; Gao, Z.; Zheng, X.; Zhu, K.; Zhang, Z.; Zhang, Z.; Dong, Y.; Huang, L.; Gong, Y. Crystal Structure of Sulfolobus Solfataricus Topoisomerase III Reveals a Novel Carboxyl-Terminal Zinc Finger Domain Essential for Decatenation Activity. bioRxiv 2021. [Google Scholar] [CrossRef] [Green Version]
- Feinberg, H.; Lima, C.D.; Mondragon, A. Conformational Changes in E. Coli DNA Topoisomerase I. Nat. Struct. Biol. 1999, 6, 918–922. [Google Scholar] [CrossRef]
- Terekhova, K.; Gunn, K.H.; Marko, J.F.; Mondragón, A. Bacterial Topoisomerase I and Topoisomerase III Relax Supercoiled DNA via Distinct Pathways. Nucleic Acids Res. 2012, 40, 10432–10440. [Google Scholar] [CrossRef] [Green Version]
- Viard, T.; de la Tour, C.B. Type IA Topoisomerases: A Simple Puzzle? Biochimie 2007, 89, 456–467. [Google Scholar] [CrossRef]
- Bizard, A.H.; Yang, X.; Débat, H.; Fogg, J.M.; Zechiedrich, L.; Strick, T.R.; Garnier, F.; Nadal, M. TopA, the Sulfolobus Solfataricus Topoisomerase III, Is a Decatenase. Nucleic Acids Res. 2018, 46, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Kirkegaard, K.; Wang, J.C. Bacterial DNA Topoisomerase I Can Relax Positively Supercoiled DNA Containing a Single-Stranded Loop. J. Mol. Biol. 1985, 185, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Dekker, N.H.; Rybenkov, V.V.; Duguet, M.; Crisona, N.J.; Cozzarelli, N.R.; Bensimon, D.; Croquette, V. The Mechanism of Type IA Topoisomerases. Proc. Natl. Acad. Sci. USA 2002, 99, 12126–12131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viard, T.; Lamour, V.; Duguet, M.; Bouthier de la Tour, C. Hyperthermophilic Topoisomerase I from Thermotoga Maritima. J. Biol. Chem. 2001, 276, 46495–46503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Shen, W.; Guo, R.; Xue, Y.; Peng, W.; Sima, J.; Yang, J.; Sharov, A.; Srikantan, S.; Yang, J.; et al. Top3β Is an RNA Topoisomerase That Works with Fragile X Syndrome Protein to Promote Synapse Formation HHS Public Access Author Manuscript. Nat. Neurosci. 2013, 16, 1238–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahumada, A.; Tse-Dinh, Y.C. The Zn(II) Binding Motifs of E. Coli DNA Topoisomerase I Is Part of a High-Affinity DNA Binding Domain. Biochem. Biophys. Res. Commun. 1998, 251, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Viard, T.; Cossard, R.; Duguet, M.; de La Tour, C.B. Thermotoga Maritima-Escherichia Coli Chimeric Topoisomerases. J. Biol. Chem. 2004, 279, 30073–30080. [Google Scholar] [CrossRef] [Green Version]
- Ahumada, A.; Tse-Dinh, Y.-C. The Role of the Zn(II) Binding Domain in the Mechanism of E. Coli DNA Topoisomerase I. BMC Biochem. 2002, 3, 13. [Google Scholar] [CrossRef]
- Strzałka, A.; Szafran, M.J.; Strick, T.; Jakimowicz, D. C-Terminal Lysine Repeats in Streptomyces Topoisomerase I Stabilize the Enzyme–DNA Complex and Confer High Enzyme Processivity. Nucleic Acids Res. 2017, 45, 11908–11924. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, W.; Bhat, A.G.; Leelaram, M.N.; Menon, S.; Nagaraja, V. Carboxyl Terminal Domain Basic Amino Acids of Mycobacterial Topoisomerase I Bind DNA to Promote Strand Passage. Nucleic Acids Res. 2013, 41, 7462–7471. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Garnier, F.; Débat, H.; Strick, T.R.; Nadal, M. Direct Observation of Helicase-Topoisomerase Coupling within Reverse Gyrase. Proc. Natl. Acad. Sci. USA 2020, 117, 10856–10864. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidmar, V.; Vayssières, M.; Lamour, V. What’s on the Other Side of the Gate: A Structural Perspective on DNA Gate Opening of Type IA and IIA DNA Topoisomerases. Int. J. Mol. Sci. 2023, 24, 3986. https://doi.org/10.3390/ijms24043986
Vidmar V, Vayssières M, Lamour V. What’s on the Other Side of the Gate: A Structural Perspective on DNA Gate Opening of Type IA and IIA DNA Topoisomerases. International Journal of Molecular Sciences. 2023; 24(4):3986. https://doi.org/10.3390/ijms24043986
Chicago/Turabian StyleVidmar, Vita, Marlène Vayssières, and Valérie Lamour. 2023. "What’s on the Other Side of the Gate: A Structural Perspective on DNA Gate Opening of Type IA and IIA DNA Topoisomerases" International Journal of Molecular Sciences 24, no. 4: 3986. https://doi.org/10.3390/ijms24043986
APA StyleVidmar, V., Vayssières, M., & Lamour, V. (2023). What’s on the Other Side of the Gate: A Structural Perspective on DNA Gate Opening of Type IA and IIA DNA Topoisomerases. International Journal of Molecular Sciences, 24(4), 3986. https://doi.org/10.3390/ijms24043986