
Graft-versus-Host Disease Modulation by Innate T Cells

 , and
, and
Abstract
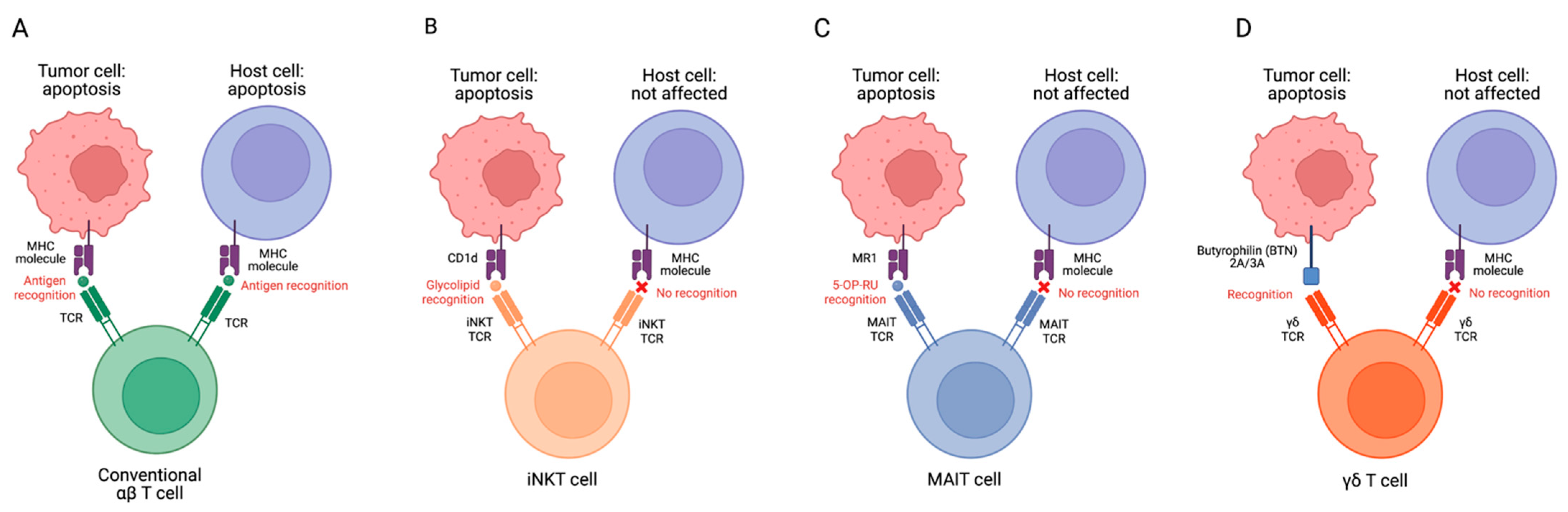
:1. Introduction
2. MAIT Cell Modulation of GvHD
3. iNKT Cells Modulation of GvHD
4. γδ T Cell Modulation of GvHD
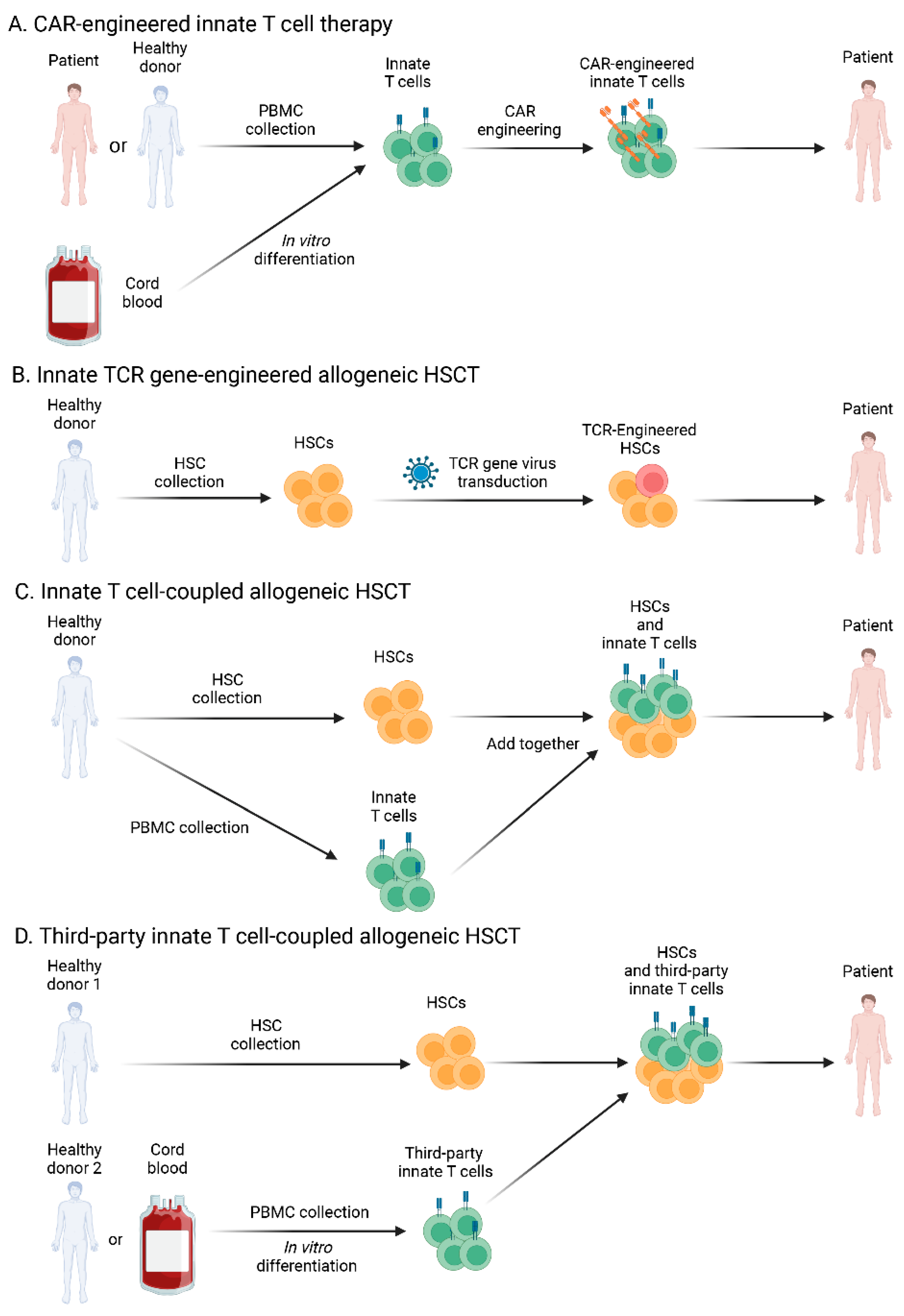
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrara, J.L.; Levine, J.E.; Reddy, P.; Holler, E. Graft-versus-host disease. Lancet 2009, 373, 1550–1561. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, A.; Labopin, M.; Bacigalupo, A.; Afanasyev, B.; Cornelissen, J.J.; Elmaagacli, A.; Itälä-Remes, M.; Blaise, D.; Meijer, E.; Koc, Y.; et al. Post-transplant cyclophosphamide for graft-versus-host disease prophylaxis in HLA matched sibling or matched unrelated donor transplant for patients with acute leukemia, on behalf of ALWP-EBMT. J. Hematol. Oncol. 2018, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Riesner, K.; Kalupa, M.; Shi, Y.; Elezkurtaj, S.; Penack, O. A preclinical acute GVHD mouse model based on chemotherapy conditioning and MHC-matched transplantation. Bone Marrow Transpl. 2016, 51, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Blazar, B.R.; Taylor, P.A.; Vallera, D.A. CD4+ and CD8+ T cells each can utilize a perforin-dependent pathway to mediate lethal graft-versus-host disease in major histocompatibility complex-disparate recipients. Transplantation 1997, 64, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, Y.; Zheng, H. The Effects of Interferons on Allogeneic T Cell Response in GVHD: The Multifaced Biology and Epigenetic Regulations. Front. Immunol. 2021, 12, 717540. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Joe, G.; Hexner, E.; Zhu, J.; Emerson, S.G. Alloreactive memory T cells are responsible for the persistence of graft-versus-host disease. J. Immunol. 2005, 174, 3051–3058. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.J.; Zhao, X.Y.; Huo, M.R.; Huang, X.J. Expression of CD62L on donor CD4(+) T cells in allografts: Correlation with graft-versus-host disease after unmanipulated allogeneic blood and marrow transplantation. J. Clin. Immunol. 2009, 29, 696–704. [Google Scholar] [CrossRef]
- Chen, B.J.; Cui, X.; Sempowski, G.D.; Liu, C.; Chao, N.J. Transfer of allogeneic CD62L− memory T cells without graft-versus-host disease. Blood 2004, 103, 1534–1541. [Google Scholar] [CrossRef] [Green Version]
- Dutt, S.; Tseng, D.; Ermann, J.; George, T.I.; Liu, Y.P.; Davis, C.R.; Fathman, C.G.; Strober, S. Naive and memory T cells induce different types of graft-versus-host disease. J. Immunol. 2007, 179, 6547–6554. [Google Scholar] [CrossRef] [Green Version]
- Delens, L.; Ehx, G.; Somja, J.; Vrancken, L.; Belle, L.; Seidel, L.; Grégoire, C.; Fransolet, G.; Ritacco, C.; Hannon, M.; et al. In Vitro Th17-Polarized Human CD4(+) T Cells Exacerbate Xenogeneic Graft-versus-Host Disease. Biol. Blood Marrow Transplant. 2019, 25, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Carlson, M.J.; West, M.L.; Coghill, J.M.; Panoskaltsis-Mortari, A.; Blazar, B.R.; Serody, J.S. In vitro-differentiated TH17 cells mediate lethal acute graft-versus-host disease with severe cutaneous and pulmonary pathologic manifestations. Blood 2009, 113, 1365–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.W.; Su, X.H.; Wang, M.Y.; Han, M.Z.; Feng, X.M.; Jiang, E.L. Regulatory T Cells in GVHD Therapy. Front. Immunol. 2021, 12, 697854. [Google Scholar] [CrossRef] [PubMed]
- Booth, C.; Lawson, S.; Veys, P. The current role of T cell depletion in paediatric stem cell transplantation. Br. J. Haematol. 2013, 162, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, V.A.; Kernan, N.A.; Boulad, F.; Cancio, M.; Allen, J.; Higman, M.; Margossian, S.P.; Mauguen, A.; Prockop, S.; Scaradavou, A.; et al. Low toxicity and favorable overall survival in relapsed/refractory B-ALL following CAR T cells and CD34-selected T-cell depleted allogeneic hematopoietic cell transplant. Bone Marrow Transpl. 2020, 55, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Bleakley, M.; Sehgal, A.; Seropian, S.; Biernacki, M.A.; Krakow, E.F.; Dahlberg, A.; Persinger, H.; Hilzinger, B.; Martin, P.J.; Carpenter, P.A.; et al. Naive T-Cell Depletion to Prevent Chronic Graft-Versus-Host Disease. J. Clin. Oncol. 2022, 40, 1174–1185. [Google Scholar] [CrossRef]
- Pierini, A.; Ruggeri, L.; Mancusi, A.; Carotti, A.; Falzetti, F.; Terenzi, A.; Martelli, M.F.; Velardi, A. T cell depletion and no post transplant immune suppression allow separation of graft versus leukemia from graft versus host disease. Bone Marrow Transpl. 2019, 54, 775–779. [Google Scholar] [CrossRef]
- Zafarani, A.; Taghavi-Farahabadi, M.; Razizadeh, M.H.; Amirzargar, M.R.; Mansouri, M.; Mahmoudi, M. The Role of NK Cells and Their Exosomes in Graft Versus Host Disease and Graft Versus Leukemia. Stem Cell Rev. Rep. 2023, 19, 26–45. [Google Scholar] [CrossRef]
- Li, T.; Li, X.; Zamani, A.; Wang, W.; Lee, C.N.; Li, M.; Luo, G.; Eiler, E.; Sun, H.; Ghosh, S.; et al. c-Rel Is a Myeloid Checkpoint for Cancer Immunotherapy. Nat. Cancer 2020, 1, 507–517. [Google Scholar] [CrossRef]
- Demosthenous, C.; Sakellari, I.; Douka, V.; Papayanni, P.G.; Anagnostopoulos, A.; Gavriilaki, E. The Role of Myeloid-Derived Suppressor Cells (MDSCs) in Graft-versus-Host Disease (GVHD). J. Clin. Med. 2021, 10, 2050. [Google Scholar] [CrossRef]
- Ivanov, S.; Paget, C.; Trottein, F. Role of non-conventional T lymphocytes in respiratory infections: The case of the pneumococcus. PLoS Pathog. 2014, 10, e1004300. [Google Scholar] [CrossRef]
- Gao, M.G.; Hong, Y.; Zhao, X.Y.; Pan, X.A.; Sun, Y.Q.; Kong, J.; Wang, Z.D.; Wang, F.R.; Wang, J.Z.; Yan, C.H.; et al. The Potential Roles of Mucosa-Associated Invariant T Cells in the Pathogenesis of Gut Graft-Versus-Host Disease after Hematopoietic Stem Cell Transplantation. Front. Immunol. 2021, 12, 720354. [Google Scholar] [CrossRef] [PubMed]
- Andrlová, H.; Miltiadous, O.; Kousa, A.I.; Dai, A.; DeWolf, S.; Violante, S.; Park, H.Y.; Janaki-Raman, S.; Gardner, R.; El Daker, S.; et al. MAIT and Vδ2 unconventional T cells are supported by a diverse intestinal microbiome and correlate with favorable patient outcome after allogeneic HCT. Sci. Transl. Med. 2022, 14, eabj2829. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, K.; Umeda, K.; Hiejima, E.; Iwai, A.; Mikami, M.; Nodomi, S.; Saida, S.; Kato, I.; Hiramatsu, H.; Yasumi, T.; et al. Influence of post-transplant mucosal-associated invariant T cell recovery on the development of acute graft-versus-host disease in allogeneic bone marrow transplantation. Int. J. Hematol. 2018, 108, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Mengge, G.; Hong, Y.; Sun, Y.; Kong, J.; Yan, C.; Wang, Z.; Wang, Y.; Huang, X.; Zhao, X. The Low Number of Mucosal-Associated Invariant T Cells in the Graft Was Associated with Occurrence of Gut Graft-Versus-Host Disease. Blood 2019, 134, 2001. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Hanafi, L.A.; Sheih, A.; Golob, J.L.; Srinivasan, S.; Boeckh, M.J.; Pergam, S.A.; Mahmood, S.; Baker, K.K.; Gooley, T.A.; et al. Graft-Derived Reconstitution of Mucosal-Associated Invariant T Cells after Allogeneic Hematopoietic Cell Transplantation. Biol. Blood Marrow Transpl. 2018, 24, 242–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varelias, A.; Bunting, M.D.; Ormerod, K.L.; Koyama, M.; Olver, S.D.; Straube, J.; Kuns, R.D.; Robb, R.J.; Henden, A.S.; Cooper, L.; et al. Recipient mucosal-associated invariant T cells control GVHD within the colon. J. Clin. Investig. 2018, 128, 1919–1936. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.T.; Moreira-Teixeira, L.; Bachy, E.; Bouillié, M.; Milpied, P.; Coman, T.; Suarez, F.; Marcais, A.; Sibon, D.; Buzyn, A.; et al. Early posttransplantation donor-derived invariant natural killer T-cell recovery predicts the occurrence of acute graft-versus-host disease and overall survival. Blood 2012, 120, 2144–2154. [Google Scholar] [CrossRef]
- Malard, F.; Labopin, M.; Chevallier, P.; Guillaume, T.; Duquesne, A.; Rialland, F.; Derenne, S.; Peterlin, P.; Leauté, A.G.; Brissot, E.; et al. Larger number of invariant natural killer T cells in PBSC allografts correlates with improved GVHD-free and progression-free survival. Blood 2016, 127, 1828–1835. [Google Scholar] [CrossRef] [Green Version]
- Haraguchi, K.; Takahashi, T.; Hiruma, K.; Kanda, Y.; Tanaka, Y.; Ogawa, S.; Chiba, S.; Miura, O.; Sakamaki, H.; Hirai, H. Recovery of Valpha24+ NKT cells after hematopoietic stem cell transplantation. Bone Marrow Transpl. 2004, 34, 595–602. [Google Scholar] [CrossRef]
- Chaidos, A.; Patterson, S.; Szydlo, R.; Chaudhry, M.S.; Dazzi, F.; Kanfer, E.; McDonald, D.; Marin, D.; Milojkovic, D.; Pavlu, J.; et al. Graft invariant natural killer T-cell dose predicts risk of acute graft-versus-host disease in allogeneic hematopoietic stem cell transplantation. Blood 2012, 119, 5030–5036. [Google Scholar] [CrossRef] [Green Version]
- Pillai, A.B.; George, T.I.; Dutt, S.; Strober, S. Host natural killer T cells induce an interleukin-4-dependent expansion of donor CD4+CD25+Foxp3+ T regulatory cells that protects against graft-versus-host disease. Blood 2009, 113, 4458–4467. [Google Scholar] [CrossRef] [PubMed]
- Schneidawind, D.; Pierini, A.; Alvarez, M.; Pan, Y.; Baker, J.; Buechele, C.; Luong, R.H.; Meyer, E.H.; Negrin, R.S. CD4+ invariant natural killer T cells protect from murine GVHD lethality through expansion of donor CD4+CD25+FoxP3+ regulatory T cells. Blood 2014, 124, 3320–3328. [Google Scholar] [CrossRef] [PubMed]
- Schneidawind, D.; Baker, J.; Pierini, A.; Buechele, C.; Luong, R.H.; Meyer, E.H.; Negrin, R.S. Third-party CD4+ invariant natural killer T cells protect from murine GVHD lethality. Blood 2015, 125, 3491–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveson-Gower, D.B.; Olson, J.A.; Sega, E.I.; Luong, R.H.; Baker, J.; Zeiser, R.; Negrin, R.S. Low doses of natural killer T cells provide protection from acute graft-versus-host disease via an IL-4-dependent mechanism. Blood 2011, 117, 3220–3229. [Google Scholar] [CrossRef] [Green Version]
- Coman, T.; Rossignol, J.; D’Aveni, M.; Fabiani, B.; Dussiot, M.; Rignault, R.; Babdor, J.; Bouillé, M.; Herbelin, A.; Coté, F.; et al. Human CD4- invariant NKT lymphocytes regulate graft versus host disease. Oncoimmunology 2018, 7, e1470735. [Google Scholar] [CrossRef] [Green Version]
- Schmid, H.; Ribeiro, E.M.; Secker, K.A.; Duerr-Stoerzer, S.; Keppeler, H.; Dong, R.; Munz, T.; Schulze-Osthoff, K.; Hailfinger, S.; Schneidawind, C.; et al. Human invariant natural killer T cells promote tolerance by preferential apoptosis induction of conventional dendritic cells. Haematologica 2022, 107, 427–436. [Google Scholar] [CrossRef]
- Li, Y.R.; Zhou, Y.; Kim, Y.J.; Zhu, Y.; Ma, F.; Yu, J.; Wang, Y.C.; Chen, X.; Li, Z.; Zeng, S.; et al. Development of allogeneic HSC-engineered iNKT cells for off-the-shelf cancer immunotherapy. Cell Rep. Med. 2021, 2, 100449. [Google Scholar] [CrossRef]
- Du, J.; Paz, K.; Thangavelu, G.; Schneidawind, D.; Baker, J.; Flynn, R.; Duramad, O.; Feser, C.; Panoskaltsis-Mortari, A.; Negrin, R.S.; et al. Invariant natural killer T cells ameliorate murine chronic GVHD by expanding donor regulatory T cells. Blood 2017, 129, 3121–3125. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Zeng, S.; Dunn, Z.S.; Zhou, Y.; Li, Z.; Yu, J.; Wang, Y.C.; Ku, J.; Cook, N.; Kramer, A.; et al. Off-the-shelf third-party HSC-engineered iNKT cells for ameliorating GvHD while preserving GvL effect in the treatment of blood cancers. iScience 2022, 25, 104859. [Google Scholar] [CrossRef]
- Trujillo-Ocampo, A.; Cho, H.W.; Herrmann, A.C.; Ruiz-Vazquez, W.; Thornton, A.B.; He, H.; Li, D.; Qazilbash, M.A.; Ma, Q.; Porcelli, S.A.; et al. Rapid ex vivo expansion of highly enriched human invariant natural killer T cells via single antigenic stimulation for cell therapy to prevent graft-versus-host disease. Cytotherapy 2018, 20, 1089–1101. [Google Scholar] [CrossRef]
- Pillai, A.B.; George, T.I.; Dutt, S.; Teo, P.; Strober, S. Host NKT cells can prevent graft-versus-host disease and permit graft antitumor activity after bone marrow transplantation. J. Immunol. 2007, 178, 6242–6251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas-Bauer, K.; Lohmeyer, J.K.; Hirai, T.; Ramos, T.L.; Fazal, F.M.; Litzenburger, U.M.; Yost, K.E.; Ribado, J.V.; Kambham, N.; Wenokur, A.S.; et al. Invariant natural killer T-cell subsets have diverse graft-versus-host-disease-preventing and antitumor effects. Blood 2021, 138, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Minculescu, L.; Marquart, H.V.; Ryder, L.P.; Andersen, N.S.; Schjoedt, I.; Friis, L.S.; Kornblit, B.T.; Petersen, S.L.; Haastrup, E.; Fischer-Nielsen, A.; et al. Improved Overall Survival, Relapse-Free-Survival, and Less Graft-vs.-Host-Disease in Patients with High Immune Reconstitution of TCR Gamma Delta Cells 2 Months after Allogeneic Stem Cell Transplantation. Front. Immunol. 2019, 10, 1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pabst, C.; Schirutschke, H.; Ehninger, G.; Bornhäuser, M.; Platzbecker, U. The graft content of donor T cells expressing gamma delta TCR+ and CD4+foxp3+ predicts the risk of acute graft versus host disease after transplantation of allogeneic peripheral blood stem cells from unrelated donors. Clin. Cancer Res. 2007, 13, 2916–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minculescu, L.; Sengeløv, H. The role of gamma delta T cells in haematopoietic stem cell transplantation. Scand. J. Immunol. 2015, 81, 459–468. [Google Scholar] [CrossRef]
- Gao, L.; Xuan, L.; Wu, X.; Fan, Z.; Huang, F.; Yi, Z.; Li, Y.; Liu, Q. Increase of regulatory γδ T cells reduces the incidence of acute graft-versus-host disease after allogeneic hematopoietic stem cell transplantation. Blood 2016, 128, 2230. [Google Scholar] [CrossRef]
- Xuan, L.; Wu, X.; Qiu, D.; Gao, L.; Liu, H.; Fan, Z.; Huang, F.; Jin, Z.; Sun, J.; Li, Y.; et al. Regulatory γδ T cells induced by G-CSF participate in acute graft-versus-host disease regulation in G-CSF-mobilized allogeneic peripheral blood stem cell transplantation. J. Transl. Med. 2018, 16, 144. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.R.; Brown, J.; Yu, Y.; Lee, D.; Zhou, K.; Dunn, Z.S.; Hon, R.; Wilson, M.; Kramer, A.; Zhu, Y.; et al. Targeting Immunosuppressive Tumor-Associated Macrophages Using Innate T Cells for Enhanced Antitumor Reactivity. Cancers 2022, 14, 2749. [Google Scholar] [CrossRef]
- Napier, R.J.; Adams, E.J.; Gold, M.C.; Lewinsohn, D.M. The Role of Mucosal Associated Invariant T Cells in Antimicrobial Immunity. Front. Immunol. 2015, 6, 344. [Google Scholar] [CrossRef] [Green Version]
- Nel, I.; Bertrand, L.; Toubal, A.; Lehuen, A. MAIT cells, guardians of skin and mucosa? Mucosal Immunol. 2021, 14, 803–814. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Koay, H.F.; McCluskey, J.; Gherardin, N.A. The biology and functional importance of MAIT cells. Nat. Immunol. 2019, 20, 1110–1128. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Lora, N.E.; Kauffman, K.D.; Dorosky, D.E.; Oh, S.; Namasivayam, S.; Gomez, F.; Fleegle, J.D.; Arlehamn, C.S.L.; Sette, A.; et al. Functional inactivation of pulmonary MAIT cells following 5-OP-RU treatment of non-human primates. Mucosal Immunol. 2021, 14, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Le Bourhis, L.; Martin, E.; Péguillet, I.; Guihot, A.; Froux, N.; Coré, M.; Lévy, E.; Dusseaux, M.; Meyssonnier, V.; Premel, V.; et al. Antimicrobial activity of mucosal-associated invariant T cells. Nat. Immunol. 2010, 11, 701–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jardine, L.; Cytlak, U.; Gunawan, M.; Reynolds, G.; Green, K.; Wang, X.N.; Pagan, S.; Paramitha, M.; Lamb, C.A.; Long, A.K.; et al. Donor monocyte-derived macrophages promote human acute graft-versus-host disease. J. Clin. Investig. 2020, 130, 4574–4586. [Google Scholar] [CrossRef] [PubMed]
- Chakraverty, R.; Sykes, M. The role of antigen-presenting cells in triggering graft-versus-host disease and graft-versus-leukemia. Blood 2007, 110, 9–17. [Google Scholar] [CrossRef]
- Mavers, M.; Maas-Bauer, K.; Negrin, R.S. Invariant Natural Killer T Cells as Suppressors of Graft-versus-Host Disease in Allogeneic Hematopoietic Stem Cell Transplantation. Front. Immunol. 2017, 8, 900. [Google Scholar] [CrossRef] [Green Version]
- Novak, J.; Lehuen, A. Mechanism of regulation of autoimmunity by iNKT cells. Cytokine 2011, 53, 263–270. [Google Scholar] [CrossRef]
- Bendelac, A.; Savage, P.B.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [Green Version]
- Opasawatchai, A.; Matangkasombut, P. iNKT Cells and their Potential Lipid Ligands during Viral Infection. Front. Immunol. 2015, 6, 378. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Kronenberg, M. Activation and Function of iNKT and MAIT Cells. Adv. Immunol. 2015, 127, 145–201. [Google Scholar] [CrossRef]
- Brennan, P.J.; Brigl, M.; Brenner, M.B. Invariant natural killer T cells: An innate activation scheme linked to diverse effector functions. Nat. Rev. Immunol. 2013, 13, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, J.L.; Mallevaey, T.; Scott-Browne, J.; Gapin, L. CD1d-restricted iNKT cells, the ‘Swiss-Army knife’ of the immune system. Curr. Opin. Immunol. 2008, 20, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, T.; Cui, J.; Koezuka, Y.; Toura, I.; Kaneko, Y.; Motoki, K.; Ueno, H.; Nakagawa, R.; Sato, H.; Kondo, E.; et al. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science 1997, 278, 1626–1629. [Google Scholar] [CrossRef] [PubMed]
- Borg, N.A.; Wun, K.S.; Kjer-Nielsen, L.; Wilce, M.C.; Pellicci, D.G.; Koh, R.; Besra, G.S.; Bharadwaj, M.; Godfrey, D.I.; McCluskey, J.; et al. CD1d-lipid-antigen recognition by the semi-invariant NKT T-cell receptor. Nature 2007, 448, 44–49. [Google Scholar] [CrossRef]
- Scott-Browne, J.P.; Matsuda, J.L.; Mallevaey, T.; White, J.; Borg, N.A.; McCluskey, J.; Rossjohn, J.; Kappler, J.; Marrack, P.; Gapin, L. Germline-encoded recognition of diverse glycolipids by natural killer T cells. Nat. Immunol. 2007, 8, 1105–1113. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Patel, O.; Kjer-Nielsen, L.; Pang, S.S.; Sullivan, L.C.; Kyparissoudis, K.; Brooks, A.G.; Reid, H.H.; Gras, S.; Lucet, I.S.; et al. Differential recognition of CD1d-alpha-galactosyl ceramide by the V beta 8.2 and V beta 7 semi-invariant NKT T cell receptors. Immunity 2009, 31, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Mallevaey, T.; Scott-Browne, J.P.; Matsuda, J.L.; Young, M.H.; Pellicci, D.G.; Patel, O.; Thakur, M.; Kjer-Nielsen, L.; Richardson, S.K.; Cerundolo, V.; et al. T cell receptor CDR2 beta and CDR3 beta loops collaborate functionally to shape the iNKT cell repertoire. Immunity 2009, 31, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Rubio, M.T.; Bouillié, M.; Bouazza, N.; Coman, T.; Trebeden-Nègre, H.; Gomez, A.; Suarez, F.; Sibon, D.; Brignier, A.; Paubelle, E.; et al. Pre-transplant donor CD4(-) invariant NKT cell expansion capacity predicts the occurrence of acute graft-versus-host disease. Leukemia 2017, 31, 903–912. [Google Scholar] [CrossRef]
- Lan, F.; Zeng, D.; Higuchi, M.; Higgins, J.P.; Strober, S. Host conditioning with total lymphoid irradiation and antithymocyte globulin prevents graft-versus-host disease: The role of CD1-reactive natural killer T cells. Biol. Blood Marrow Transplant. 2003, 9, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Lowsky, R.; Takahashi, T.; Liu, Y.P.; Dejbakhsh-Jones, S.; Grumet, F.C.; Shizuru, J.A.; Laport, G.G.; Stockerl-Goldstein, K.E.; Johnston, L.J.; Hoppe, R.T.; et al. Protective conditioning for acute graft-versus-host disease. N. Engl. J. Med. 2005, 353, 1321–1331. [Google Scholar] [CrossRef]
- Kohrt, H.E.; Turnbull, B.B.; Heydari, K.; Shizuru, J.A.; Laport, G.G.; Miklos, D.B.; Johnston, L.J.; Arai, S.; Weng, W.K.; Hoppe, R.T.; et al. TLI and ATG conditioning with low risk of graft-versus-host disease retains antitumor reactions after allogeneic hematopoietic cell transplantation from related and unrelated donors. Blood 2009, 114, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Beziat, V.; Nguyen, S.; Exley, M.; Achour, A.; Simon, T.; Chevallier, P.; Sirvent, A.; Vigouroux, S.; Debré, P.; Rio, B.; et al. Shaping of iNKT cell repertoire after unrelated cord blood transplantation. Clin. Immunol. 2010, 135, 364–373. [Google Scholar] [CrossRef] [PubMed]
- de Lalla, C.; Rinaldi, A.; Montagna, D.; Azzimonti, L.; Bernardo, M.E.; Sangalli, L.M.; Paganoni, A.M.; Maccario, R.; Di Cesare-Merlone, A.; Zecca, M.; et al. Invariant NKT cell reconstitution in pediatric leukemia patients given HLA-haploidentical stem cell transplantation defines distinct CD4+ and CD4− subset dynamics and correlates with remission state. J. Immunol. 2011, 186, 4490–4499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellabona, P.; Casorati, G.; de Lalla, C.; Montagna, D.; Locatelli, F. On the use of donor-derived iNKT cells for adoptive immunotherapy to prevent leukemia recurrence in pediatric recipients of HLA haploidentical HSCT for hematological malignancies. Clin. Immunol. 2011, 140, 152–159. [Google Scholar] [CrossRef]
- Erkers, T.; Xie, B.J.; Kenyon, L.J.; Smith, B.; Rieck, M.; Jensen, K.P.; Ji, X.; Basina, M.; Strober, S.; Negrin, R.S.; et al. High-parametric evaluation of human invariant natural killer T cells to delineate heterogeneity in allo- and autoimmunity. Blood 2020, 135, 814–825. [Google Scholar] [CrossRef]
- Vantourout, P.; Hayday, A. Six-of-the-best: Unique contributions of γδ T cells to immunology. Nat. Rev. Immunol. 2013, 13, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, L.C.; Shaw, E.M.; Stankovic, S.; Snell, G.I.; Brooks, A.G.; Westall, G.P. The complex existence of γδ T cells following transplantation: The good, the bad and the simply confusing. Clin. Transl. Immunol. 2019, 8, e1078. [Google Scholar] [CrossRef]
- Khairallah, C.; Chu, T.H.; Sheridan, B.S. Tissue Adaptations of Memory and Tissue-Resident Gamma Delta T Cells. Front. Immunol. 2018, 9, 2636. [Google Scholar] [CrossRef] [Green Version]
- Kalyan, S.; Kabelitz, D. Defining the nature of human γδ T cells: A biographical sketch of the highly empathetic. Cell. Mol. Immunol. 2013, 10, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, I.; Salio, M.; Koo, D.D.; Cerundolo, L.; Shepherd, D.; Cerundolo, V.; Fuggle, S.V. Expression of MHC class I-related Chain B (MICB) molecules on renal transplant biopsies. Transplantation 2006, 81, 1196–1203. [Google Scholar] [CrossRef]
- Yin, S.; Zhang, J.; Mao, Y.; Hu, Y.; Cui, L.; Kang, N.; He, W. Vav1-phospholipase C-γ1 (Vav1-PLC-γ1) pathway initiated by T cell antigen receptor (TCRγδ) activation is required to overcome inhibition by ubiquitin ligase Cbl-b during γδT cell cytotoxicity. J. Biol. Chem. 2013, 288, 26448–26462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willcox, C.R.; Pitard, V.; Netzer, S.; Couzi, L.; Salim, M.; Silberzahn, T.; Moreau, J.F.; Hayday, A.C.; Willcox, B.E.; Déchanet-Merville, J. Cytomegalovirus and tumor stress surveillance by binding of a human γδ T cell antigen receptor to endothelial protein C receptor. Nat. Immunol. 2012, 13, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Marlin, R.; Pappalardo, A.; Kaminski, H.; Willcox, C.R.; Pitard, V.; Netzer, S.; Khairallah, C.; Lomenech, A.M.; Harly, C.; Bonneville, M.; et al. Sensing of cell stress by human γδ TCR-dependent recognition of annexin A2. Proc. Natl. Acad. Sci. USA 2017, 114, 3163–3168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, M.; Mann, B.T.; Chitrakar, A.; Soriano-Sarabia, N. Human Vδ2 T Cells and their Versatility for Immunotherapeutic Approaches. Cells 2022, 11, 3572. [Google Scholar] [CrossRef] [PubMed]
- Sutton, C.E.; Lalor, S.J.; Sweeney, C.M.; Brereton, C.F.; Lavelle, E.C.; Mills, K.H. Interleukin-1 and IL-23 induce innate IL-17 production from gammadelta T cells, amplifying Th17 responses and autoimmunity. Immunity 2009, 31, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Niu, C.; Cui, J. Gamma-delta (γδ) T cells: Friend or foe in cancer development? J. Transl. Med. 2018, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhita, N.; Raikar, S.S. Allogeneic gamma delta T cells as adoptive cellular therapy for hematologic malignancies. Explor. Immunol. 2022, 2, 334–350. [Google Scholar] [CrossRef]
- Gertner-Dardenne, J.; Castellano, R.; Mamessier, E.; Garbit, S.; Kochbati, E.; Etienne, A.; Charbonnier, A.; Collette, Y.; Vey, N.; Olive, D. Human Vγ9Vδ2 T cells specifically recognize and kill acute myeloid leukemic blasts. J. Immunol. 2012, 188, 4701–4708. [Google Scholar] [CrossRef] [Green Version]
- Benyamine, A.; Le Roy, A.; Mamessier, E.; Gertner-Dardenne, J.; Castanier, C.; Orlanducci, F.; Pouyet, L.; Goubard, A.; Collette, Y.; Vey, N.; et al. BTN3A molecules considerably improve Vγ9Vδ2T cells-based immunotherapy in acute myeloid leukemia. Oncoimmunology 2016, 5, e1146843. [Google Scholar] [CrossRef] [Green Version]
- Nishimoto, K.P.; Barca, T.; Azameera, A.; Makkouk, A.; Romero, J.M.; Bai, L.; Brodey, M.M.; Kennedy-Wilde, J.; Shao, H.; Papaioannou, S.; et al. Allogeneic CD20-targeted γδ T cells exhibit innate and adaptive antitumor activities in preclinical B-cell lymphoma models. Clin. Transl. Immunol. 2022, 11, e1373. [Google Scholar] [CrossRef]
- Wu, N.; Liu, R.; Liang, S.; Gao, H.; Xu, L.P.; Zhang, X.H.; Liu, J.; Huang, X.J. γδ T Cells may Aggravate Acute Graft-Versus-Host Disease through CXCR4 Signaling after Allogeneic Hematopoietic Transplantation. Front. Immunol. 2021, 12, 687961. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, S.; Zhang, X.; Liu, L.; Zhou, L.; Shen, Y.; Zhang, R.; He, Y.; Yang, D.; Jiang, E.; et al. Mucosal-associated invariant T cells predict increased acute graft-versus-host-disease incidence in patients receiving allogeneic hematopoietic stem cell transplantation. Cancer Cell. Int. 2022, 22, 297. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Wilson, M.; Yang, L. Target tumor microenvironment by innate T cells. Front. Immunol. 2022, 13, 999549. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Smith, D.J.; Zhou, Y.; Li, Y.R.; Yu, J.; Lee, D.; Wang, Y.C.; Di Biase, S.; Wang, X.; Hardoy, C.; et al. Development of Hematopoietic Stem Cell-Engineered Invariant Natural Killer T Cell Therapy for Cancer. Cell Stem Cell 2019, 25, 542–557.e9. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.H. Shared biology of GVHD and GVT effects: Potential methods of separation. Crit. Rev. Oncol. Hematol. 2006, 57, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, A.R.; Storb, R.F. Separation of graft-vs.-tumor effects from graft-vs.-host disease in allogeneic hematopoietic cell transplantation. J. Autoimmun. 2008, 30, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.S.; Lamb, L.S.; Goldman, F.; Di Stasi, A. Improving the safety of cell therapy products by suicide gene transfer. Front. Pharmacol. 2014, 5, 254. [Google Scholar] [CrossRef]
- Li, Y.R.; Zhou, K.; Wilson, M.; Kramer, A.; Zhu, Y.; Dawson, N.; Yang, L. Mucosal-associated invariant T cells for cancer immunotherapy. Mol. Ther. 2022. [Google Scholar] [CrossRef]
- Li, Y.R.; Zhou, Y.; Kramer, A.; Yang, L. Engineering stem cells for cancer immunotherapy. Trends Cancer 2021, 7, 1059–1073. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Innate T Cell Types | Clinical or Preclinical | GvHD Modulation |
|---|---|---|
| MAIT | Clinical | An increased number of MAIT cells is associated with improved overall survival and less GvHD [21,22,23,24,25]. |
| Preclinical | MAIT cells suppressed conventional CD4+ T-cell proliferation [25]. MAIT cells regulate GvHD in part by the generation of IL17A [26]. | |
| iNKT | Clinical | A higher number of iNKT cells is correlated with better GvHD-free survival [27,28,29,30]. |
| Preclinical | Host iNKT cells trigger the expansion of donor Tregs through an interleukin-4-dependent pathway [31]. CD4+ iNKT cells protect mice from GvHD [32,33,34]. CD4− iNKT cell subset controls GvHD better than its CD4+ counterpart by inducing apoptosis of dendritic cells [35]. iNKT cells induced selective apoptosis of conventional dendritic cells but not beneficial plasmacytoid dendritic cells [36]. Allogeneic iNKT cells could target tumor cells without GvHD risk [37]. Donor iNKT cells could prevent and reverse chronic GvHD in murine models [38]. Host/allogeneic iNKT cells could ameliorate GvHD while preserving antitumor effects [32,39,40,41]. iNKT2 and iNKT17 cells are responsible for the immune regulatory properties instead of iNKT1 cells [42]. | |
| γδ T | Clinical | An increased number of γδ T cells is associated with less acute GvHD [22,43]. Patients receiving higher concentrations of donor γδ T cells have a more frequent incidence of acute GvHD [44,45]. The γδ Treg proportion has a negative correlation with acute GvHD incidence [46]. |
| Preclinical | G-CSF-induced γδ T cells suppress CD4+ T-cell proliferation and regulates acute GvHD [47]. CAR-engineered gdT cells lead to a lower incidence of GvH response [48]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.; Zhu, Y.; Kramer, A.; Chen, Y.; Li, Y.-R.; Yang, L. Graft-versus-Host Disease Modulation by Innate T Cells. Int. J. Mol. Sci. 2023, 24, 4084. https://doi.org/10.3390/ijms24044084
Fang Y, Zhu Y, Kramer A, Chen Y, Li Y-R, Yang L. Graft-versus-Host Disease Modulation by Innate T Cells. International Journal of Molecular Sciences. 2023; 24(4):4084. https://doi.org/10.3390/ijms24044084
Chicago/Turabian StyleFang, Ying, Yichen Zhu, Adam Kramer, Yuning Chen, Yan-Ruide Li, and Lili Yang. 2023. "Graft-versus-Host Disease Modulation by Innate T Cells" International Journal of Molecular Sciences 24, no. 4: 4084. https://doi.org/10.3390/ijms24044084
APA StyleFang, Y., Zhu, Y., Kramer, A., Chen, Y., Li, Y. -R., & Yang, L. (2023). Graft-versus-Host Disease Modulation by Innate T Cells. International Journal of Molecular Sciences, 24(4), 4084. https://doi.org/10.3390/ijms24044084