
Roles of RNA Methylations in Cancer Progression, Autophagy, and Anticancer Drug Resistance

Abstract
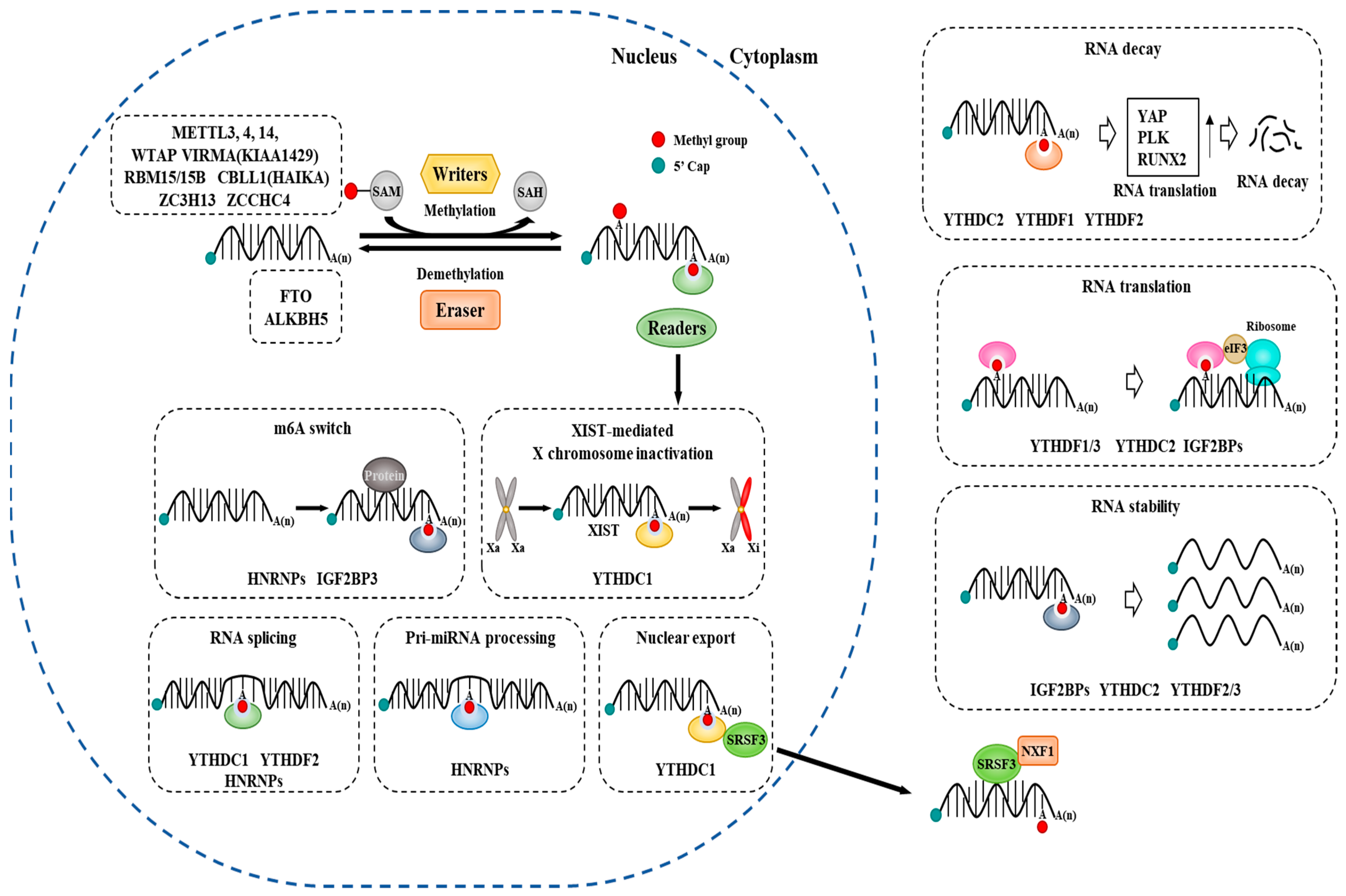
:1. Regulators of RNA Methylation
2. Role of METTL3 in Cancer
3. Role of METTL5 in Cancer
4. Role of METTL14 in Cancer
5. Role of METTL16 in Cancer
6. Roles of m6A-Binding Proteins and m6A Demethylases in Cancer
7. Roles of RNA Methylases and m6A Binding Proteins in Anticancer Drug Resistance and Immune Checkpoint
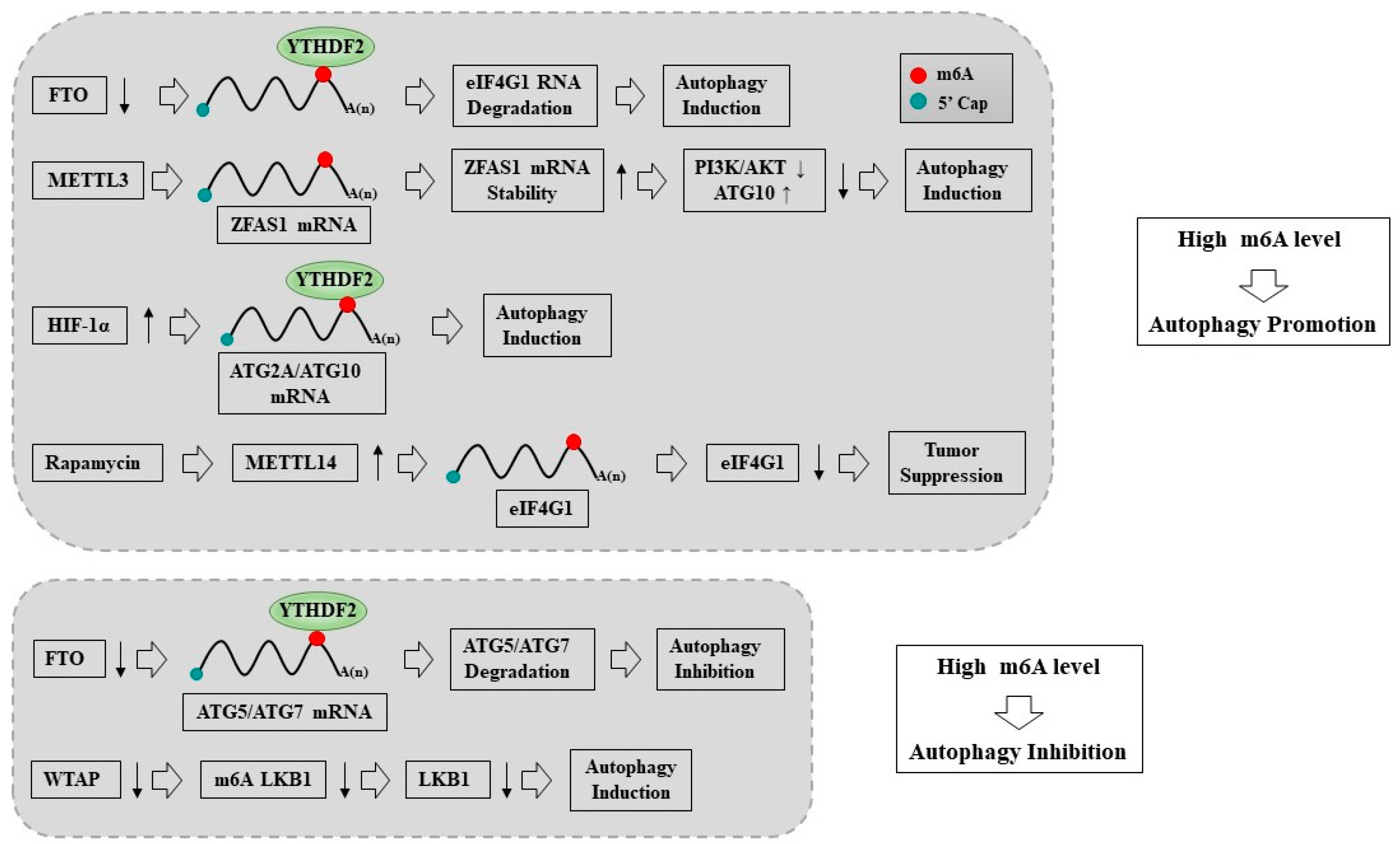
8. The Regulatory Role of m6A Modifications in Autophagy
9. The Regulatory Roles of m6A Writers in Autophagy
9.1. METTL14 Promotes Autophagy
9.2. METTL3 Promotes Autophagy
9.3. METTL1 Acts as Negative Regulator of Autophagy
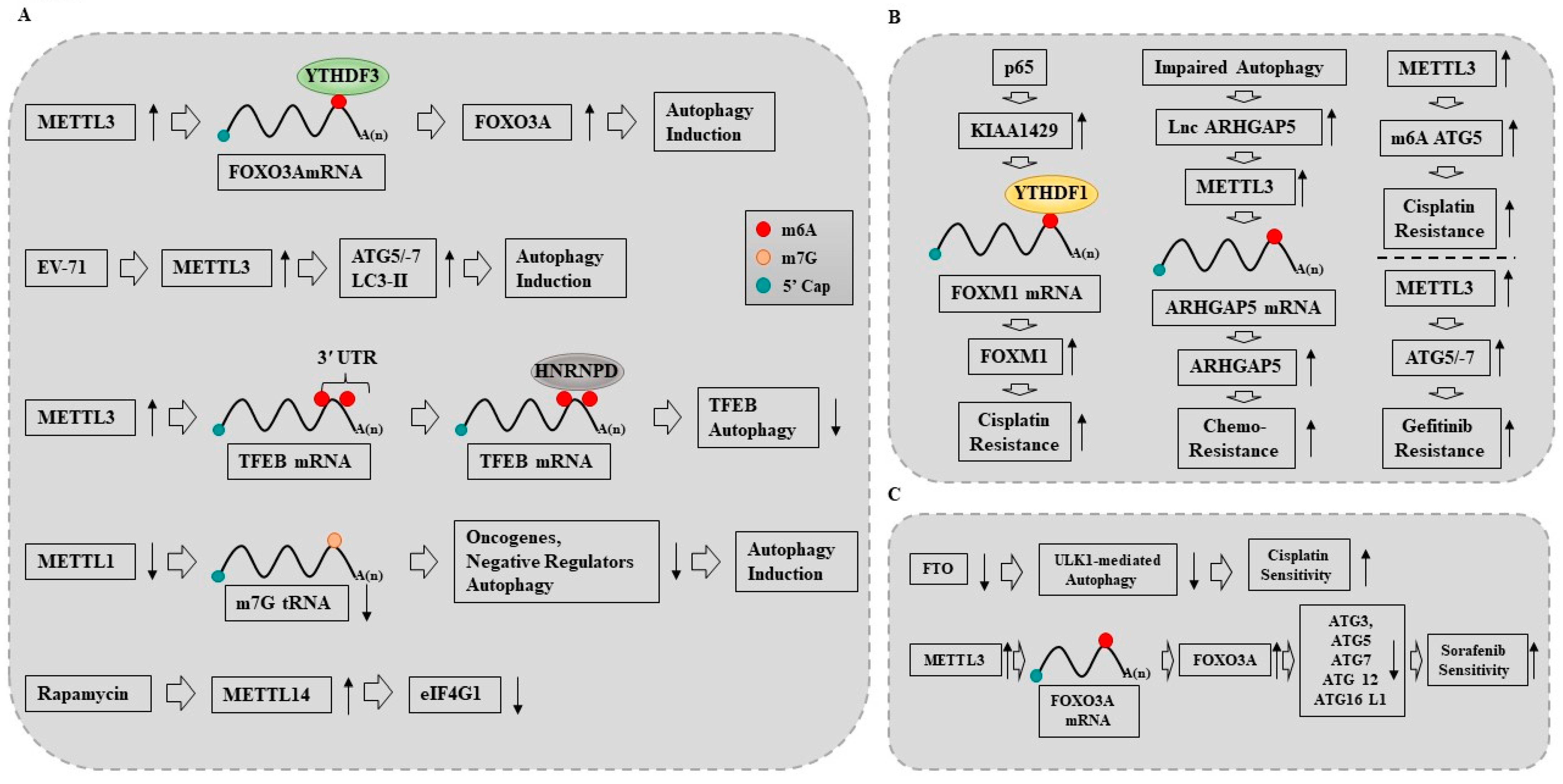
10. Cytoprotective Autophagy Promotes Anticancer Drug Resistance
11. The Roles of METTL3 in Anticancer Drug Resistance and Autophagy
11.1. METTL3 Promotes Autophagy to Induce Anticancer Drug Resistance
11.2. METTL3 Inhibits Autophagy to Reverse Anticancer Drug Resistance
12. RNA Methylome and Cancer Diagnosis
13. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nombela, P.; Miguel-Lopez, B.; Blanco, S. The role of m(6)A, m(5)C and Psi RNA modifications in cancer: Novel therapeutic opportunities. Mol. Cancer 2021, 20, 18. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Guo, J.; Fan, Z. Interactions between m6A modification and miRNAs in malignant tumors. Cell Death Dis. 2021, 12, 598. [Google Scholar] [CrossRef]
- Zhao, W.; Qi, X.; Liu, L.; Ma, S.; Liu, J.; Wu, J. Epigenetic Regulation of m(6)A Modifications in Human Cancer. Mol. Ther. Nucleic Acids 2020, 19, 405–412. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, K.; Song, B.; Ma, J.; Wu, X.; Xu, Q.; Wei, Z.; Su, J.; Liu, G.; Rong, R.; et al. m6A-Atlas: A comprehensive knowledgebase for unraveling the N6-methyladenosine (m6A) epitranscriptome. Nucleic Acids Res. 2021, 49, D134–D143. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, X.; Cai, J.; Dai, Q.; Natchiar, S.K.; Lv, R.; Chen, K.; Lu, Z.; Chen, H.; Shi, Y.G.; et al. N(6-)Methyladenosine methyltransferase ZCCHC4 mediates ribosomal RNA methylation. Nat. Chem. Biol. 2019, 15, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m(6)A RNA methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef]
- Frye, M.; Blanco, S. Post-transcriptional modifications in development and stem cells. Development 2016, 143, 3871–3881. [Google Scholar] [CrossRef] [Green Version]
- Bove, G.; Amin, S.; Babaei, M.; Benedetti, R.; Nebbioso, A.; Altucci, L.; Del Gaudio, N. Interplay between m(6) A epitranscriptome and epigenome in cancer: Current knowledge and therapeutic perspectives. Int. J. Cancer 2022. [Google Scholar] [CrossRef]
- Weng, Y.L.; Wang, X.; An, R.; Cassin, J.; Vissers, C.; Liu, Y.; Liu, Y.; Xu, T.; Wang, X.; Wong, S.Z.H.; et al. Epitranscriptomic m(6)A Regulation of Axon Regeneration in the Adult Mammalian Nervous System. Neuron 2018, 97, 313–325 e316. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Shen, L.; Liu, Y.; Ming, H.; Zhu, X.; Chu, M.; Lin, J. The m6A methyltransferase METTL3 cooperates with demethylase ALKBH5 to regulate osteogenic differentiation through NF-kappaB signaling. Mol. Cell Biochem. 2020, 463, 203–210. [Google Scholar] [CrossRef]
- Xing, M.; Liu, Q.; Mao, C.; Zeng, H.; Zhang, X.; Zhao, S.; Chen, L.; Liu, M.; Shen, B.; Guo, X.; et al. The 18S rRNA m(6) A methyltransferase METTL5 promotes mouse embryonic stem cell differentiation. EMBO Rep. 2020, 21, e49863. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by m(6)A Methylation. Cell 2017, 171, 877–889 e817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Chai, P.; Jia, R.; Jia, R. Novel insights on m(6)A RNA methylation in tumorigenesis: A double-edged sword. Mol. Cancer 2018, 17, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, I.; Much, C.; Di Giacomo, M.; Azzi, C.; Morgan, M.; Moreira, P.N.; Monahan, J.; Carrieri, C.; Enright, A.J.; O’Carroll, D. The RNA m(6)A Reader YTHDF2 Is Essential for the Post-transcriptional Regulation of the Maternal Transcriptome and Oocyte Competence. Mol. Cell 2017, 67, 1059–1067 e1054. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wei, L.; Law, C.T.; Tsang, F.H.; Shen, J.; Cheng, C.L.; Tsang, L.H.; Ho, D.W.; Chiu, D.K.; Lee, J.M.; et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Weng, H.; Chen, J. m(6)A Modification in Coding and Non-coding RNAs: Roles and Therapeutic Implications in Cancer. Cancer Cell 2020, 37, 270–288. [Google Scholar] [CrossRef]
- Anita, R.; Paramasivam, A.; Priyadharsini, J.V.; Chitra, S. The m6A readers YTHDF1 and YTHDF3 aberrations associated with metastasis and predict poor prognosis in breast cancer patients. Am. J. Cancer Res. 2020, 10, 2546–2554. [Google Scholar]
- Lin, Z.; Niu, Y.; Wan, A.; Chen, D.; Liang, H.; Chen, X.; Sun, L.; Zhan, S.; Chen, L.; Cheng, C.; et al. RNA m(6) A methylation regulates sorafenib resistance in liver cancer through FOXO3-mediated autophagy. EMBO J. 2020, 39, e103181. [Google Scholar] [CrossRef]
- Owens, M.C.; Liu, K.F. TRIBE-STAMP reveals new insights into the functions of RNA binding proteins. Genes Dev. 2022, 36, 954–955. [Google Scholar] [CrossRef]
- Yang, B.; Wang, J.Q.; Tan, Y.; Yuan, R.; Chen, Z.S.; Zou, C. RNA methylation and cancer treatment. Pharm. Res. 2021, 174, 105937. [Google Scholar] [CrossRef]
- Song, N.; Cui, K.; Zhang, K.; Yang, J.; Liu, J.; Miao, Z.; Zhao, F.; Meng, H.; Chen, L.; Chen, C.; et al. The Role of m6A RNA Methylation in Cancer: Implication for Nature Products Anti-Cancer Research. Front. Pharm. 2022, 13, 933332. [Google Scholar] [CrossRef]
- Kisan, A.; Chhabra, R. Modulation of gene expression by YTH domain family (YTHDF) proteins in human physiology and pathology. J. Cell Physiol. 2022. [Google Scholar] [CrossRef]
- Du, K.; Zhang, L.; Lee, T.; Sun, T. m(6)A RNA Methylation Controls Neural Development and Is Involved in Human Diseases. Mol. Neurobiol. 2019, 56, 1596–1606. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Li, M.; Xu, Y.; Li, X. N(6)-methyladenosine (m(6)A) writer KIAA1429 accelerates gastric cancer oxaliplatin chemoresistance by targeting FOXM1. J. Cancer Res. Clin. Oncol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.H.; Zhang, G.L.; Jiang, R.F.; Hong, Y.Q.; Zhang, Q.Y.; He, J.P.; Liu, X.R.; Yang, Z.S.; Yang, L.; Jiang, X.; et al. METTL3 is essential for normal progesterone signaling during embryo implantation via m6A-mediated translation control of progesterone receptor. Proc. Natl. Acad. Sci. USA 2023, 120, e2214684120. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Mo, J.; Chen, H.; Hu, Z.; Wang, B.; Wu, J.; Liang, Z.; Xie, W.; Du, K.; Peng, M.; et al. Structural insights into molecular mechanism for N(6)-adenosine methylation by MT-A70 family methyltransferase METTL4. Nat. Commun. 2022, 13, 5636. [Google Scholar] [CrossRef] [PubMed]
- van Tran, N.; Ernst, F.G.M.; Hawley, B.R.; Zorbas, C.; Ulryck, N.; Hackert, P.; Bohnsack, K.E.; Bohnsack, M.T.; Jaffrey, S.R.; Graille, M.; et al. The human 18S rRNA m6A methyltransferase METTL5 is stabilized by TRMT112. Nucleic Acids Res. 2019, 47, 7719–7733. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Wang, S.; Shao, M. Mechanism of METTL14-mediated m(6)A modification in non-small cell lung cancer cell resistance to cisplatin. J. Mol. Med. 2022, 100, 1771–1785. [Google Scholar] [CrossRef]
- Warda, A.S.; Kretschmer, J.; Hackert, P.; Lenz, C.; Urlaub, H.; Hobartner, C.; Sloan, K.E.; Bohnsack, M.T. Human METTL16 is a N(6)-methyladenosine (m(6)A) methyltransferase that targets pre-mRNAs and various non-coding RNAs. EMBO Rep. 2017, 18, 2004–2014. [Google Scholar] [CrossRef]
- Cai, Z.; He, S.; Liu, R.; Zhou, L.; Zhao, L. Plumbagin rescues the granulosa cell’s pyroptosis by reducing WTAP-mediated N6-methylation in polycystic ovary syndrome. J. Ovarian Res. 2022, 15, 126. [Google Scholar] [CrossRef]
- You, Y.; Liu, J.; Zhang, L.; Li, X.; Sun, Z.; Dai, Z.; Ma, J.; Jiao, G.; Chen, Y. WTAP-mediated m6A modification modulates bone marrow mesenchymal stem cells differentiation potential and osteoporosis. Cell Death Dis. 2023, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Melstrom, L.; Chen, J. RNA N(6)-methyladenosine modification in solid tumors: New therapeutic frontiers. Cancer Gene Ther. 2020, 27, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lin, D.; Wang, X.; Feng, Z.; Zhang, J.; Wang, K. The development of a novel signature based on the m(6)A RNA methylation regulator-related ceRNA network to predict prognosis and therapy response in sarcomas. Front. Genet. 2022, 13, 894080. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Wu, X.; Deng, C.; Zhao, L.; Peng, L.; Zhou, J.; Zhang, W.; Zhao, Y.; Fei, Y. The potential role of RNA N6-methyladenosine in primary Sjogren’s syndrome. Front. Med. 2022, 9, 959388. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qian, Z.; Feng, M.; Liao, W.; Wu, Q.; Wen, F.; Li, Q. Study on the prognosis, immune and drug resistance of m6A-related genes in lung cancer. BMC Bioinform. 2022, 23, 437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, C.; Yang, Z.; Zhang, G.; Wu, P.; Luo, Y.; Zeng, Q.; Wang, L.; Xue, Q.; Zhang, Y.; et al. m(6)A regulators as predictive biomarkers for chemotherapy benefit and potential therapeutic targets for overcoming chemotherapy resistance in small-cell lung cancer. J. Hematol. Oncol. 2021, 14, 190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Sheng, P.; Jiang, Y. m6A regulators are differently expressed and correlated with immune response of pancreatic adenocarcinoma. J. Cancer Res. Clin. Oncol. 2022. [Google Scholar] [CrossRef]
- Horiuchi, K.; Kawamura, T.; Hamakubo, T. Wilms’ tumor 1-associating protein complex regulates alternative splicing and polyadenylation at potential G-quadruplex-forming splice site sequences. J. Biol. Chem. 2021, 297, 101248. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells. Nat. Cell Biol. 2014, 16, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Cai, Y.; Ma, Z.; Jiang, B.; Liu, W.; Cheng, J.; Guo, N.; Wang, Z.; Sealy, J.E.; Song, C.; et al. The RNA helicase DDX5 promotes viral infection via regulating N6-methyladenosine levels on the DHX58 and NFkappaB transcripts to dampen antiviral innate immunity. PLoS Pathog. 2021, 17, e1009530. [Google Scholar] [CrossRef]
- Moindrot, B.; Cerase, A.; Coker, H.; Masui, O.; Grijzenhout, A.; Pintacuda, G.; Schermelleh, L.; Nesterova, T.B.; Brockdorff, N. A Pooled shRNA Screen Identifies Rbm15, Spen, and Wtap as Factors Required for Xist RNA-Mediated Silencing. Cell Rep. 2015, 12, 562–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Y.; Li, J.; Shen, H.; Li, Y.; Hou, Y.; Huang, L.; Song, G.; Qiao, C. WTAP dysregulation-mediated HMGN3-m6A modification inhibited trophoblast invasion in early-onset preeclampsia. FASEB J. 2022, 36, e22617. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.J.; Zhao, X.; Li, H.; Sun, L.P.; Yuan, Y. Expression profiles and prognostic roles of m6A writers, erasers and readers in gastric cancer. Future Oncol. 2021, 17, 2605–2620. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Chen, L.; Liu, Y.; Xu, C.; Jiang, D.; Song, Q.; Wang, H.; Wang, L.; Lin, Y.; et al. Identification of clinical prognostic features of esophageal cancer based on m6A regulators. Front. Immunol. 2022, 13, 950365. [Google Scholar] [CrossRef]
- Su, R.; Dong, L.; Li, Y.; Gao, M.; He, P.C.; Liu, W.; Wei, J.; Zhao, Z.; Gao, L.; Han, L.; et al. METTL16 exerts an m(6)A-independent function to facilitate translation and tumorigenesis. Nat. Cell Biol. 2022, 24, 205–216. [Google Scholar] [CrossRef]
- Ruszkowska, A. METTL16, Methyltransferase-Like Protein 16: Current Insights into Structure and Function. Int. J. Mol. Sci. 2021, 22, 2176. [Google Scholar] [CrossRef] [PubMed]
- Mendel, M.; Chen, K.M.; Homolka, D.; Gos, P.; Pandey, R.R.; McCarthy, A.A.; Pillai, R.S. Methylation of Structured RNA by the m(6)A Writer METTL16 Is Essential for Mouse Embryonic Development. Mol. Cell 2018, 71, 986–1000 e1011. [Google Scholar] [CrossRef] [Green Version]
- Doxtader, K.A.; Wang, P.; Scarborough, A.M.; Seo, D.; Conrad, N.K.; Nam, Y. Structural Basis for Regulation of METTL16, an S-Adenosylmethionine Homeostasis Factor. Mol. Cell 2018, 71, 1001–1011 e1004. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; Zhang, J.; Yang, J.; Lv, Q.; Zhong, C. Overexpression of FTO alleviates osteoarthritis by regulating the processing of miR-515-5p and the TLR4/MyD88/NF-kappaB axis. Int. Immunopharmacol. 2022, 114, 109524. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, X.; Bi, F.; Wang, D.; Zhou, X.; Li, X.; Yang, Q. FTO Inhibits Epithelial Ovarian Cancer Progression by Destabilising SNAI1 mRNA through IGF2BP2. Cancers 2022, 14, 5218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Gu, J.; Su, T.; Gu, X.; Feng, Y. The role of RNA m6A methylation in lipid metabolism. Front. Endocrinol. 2022, 13, 866116. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vagbo, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, Y.P.; Chen, J.A. The m(6)A epitranscriptome on neural development and degeneration. J. Biomed. Sci. 2021, 28, 40. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; He, X.; Zhao, Z.; Zhang, X.; Wang, T.; Jia, L. RNA m6A reader IGF2BP3 promotes metastasis of triple-negative breast cancer via SLIT2 repression. FASEB J. 2022, 36, e22618. [Google Scholar] [CrossRef]
- Patil, D.P.; Pickering, B.F.; Jaffrey, S.R. Reading m(6)A in the Transcriptome: M(6)A-Binding Proteins. Trends Cell Biol. 2018, 28, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Sun, H.; Xu, C. YTH Domain: A Family of N(6)-methyladenosine (m(6)A) Readers. Genom. Proteom. Bioinform. 2018, 16, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Orouji, E.; Peitsch, W.K.; Orouji, A.; Houben, R.; Utikal, J. Oncogenic Role of an Epigenetic Reader of m(6)A RNA Modification: YTHDF1 in Merkel Cell Carcinoma. Cancers 2020, 12, 202. [Google Scholar] [CrossRef] [Green Version]
- Hao, C.; Zheng, Y.; Jonsson, J.; Cui, X.; Yu, H.; Wu, C.; Kajitani, N.; Schwartz, S. hnRNP G/RBMX enhances HPV16 E2 mRNA splicing through a novel splicing enhancer and inhibits production of spliced E7 oncogene mRNAs. Nucleic Acids Res. 2022, 50, 3867–3891. [Google Scholar] [CrossRef]
- Ying, Y.; Ma, X.; Fang, J.; Chen, S.; Wang, W.; Li, J.; Xie, H.; Wu, J.; Xie, B.; Liu, B.; et al. EGR2-mediated regulation of m(6)A reader IGF2BP proteins drive RCC tumorigenesis and metastasis via enhancing S1PR3 mRNA stabilization. Cell Death Dis. 2021, 12, 750. [Google Scholar] [CrossRef]
- Aitken, M.J.L.; Malaney, P.; Zhang, X.; Herbrich, S.M.; Chan, L.; Benitez, O.; Rodriguez, A.G.; Ma, H.; Jacamo, R.; Duan, R.; et al. Heterogeneous nuclear ribonucleoprotein K is overexpressed in acute myeloid leukemia and causes myeloproliferation in mice via altered Runx1 splicing. NAR Cancer 2022, 4, zcac039. [Google Scholar] [CrossRef]
- Domanski, M.; Dedic, E.; Perez, M.E.; Clery, A.; Campagne, S.; Uldry, A.C.; Braga, S.; Heller, M.; Rabl, J.; Afanasyev, P.; et al. 40S hnRNP particles are a novel class of nuclear biomolecular condensates. Nucleic Acids Res. 2022, 50, 6300–6312. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.N.; Tikhaia, E.; Mourao, A.; Sattler, M. Structural effects of m6A modification of the Xist A-repeat AUCG tetraloop and its recognition by YTHDC1. Nucleic Acids Res. 2022, 50, 2350–2362. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wan, A.H.; Sun, L.; Liang, H.; Niu, Y.; Deng, Y.; Yan, S.; Wang, Q.P.; Bu, X.; Zhang, X.; et al. N6-methyladenosine demethylase FTO enhances chemo-resistance in colorectal cancer through SIVA1-mediated apoptosis. Mol. Ther. 2022. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.; Che, P.; Yang, Z.; Zhang, J.; Yang, L.; Zhuang, M.; Ou, X.; Ji, S.J. m(6)A regulation of cortical and retinal neurogenesis is mediated by the redundant m(6)A readers YTHDFs. iScience 2022, 25, 104908. [Google Scholar] [CrossRef]
- Tsai, K.; Bogerd, H.P.; Kennedy, E.M.; Emery, A.; Swanstrom, R.; Cullen, B.R. Epitranscriptomic addition of m(6)A regulates HIV-1 RNA stability and alternative splicing. Genes Dev. 2021, 35, 992–1004. [Google Scholar] [CrossRef]
- Hao, W.Y.; Lou, Y.; Hu, G.Y.; Qian, C.Y.; Liang, W.R.; Zhao, J.; Wang, X.H. RNA m6A reader YTHDF1 facilitates inflammation via enhancing NLRP3 translation. Biochem. Biophys. Res. Commun. 2022, 616, 76–81. [Google Scholar] [CrossRef]
- Kim, G.W.; Siddiqui, A. Hepatitis B Virus X Protein Expression Is Tightly Regulated by N6-Methyladenosine Modification of Its mRNA. J. Virol. 2022, 96, e0165521. [Google Scholar] [CrossRef]
- Li, P.; Shi, Y.; Gao, D.; Xu, H.; Zou, Y.; Wang, Z.; Li, W. ELK1-mediated YTHDF1 drives prostate cancer progression by facilitating the translation of Polo-like kinase 1 in an m6A dependent manner. Int. J. Biol. Sci. 2022, 18, 6145–6162. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, J.; Jin, Y.; Zheng, Q.; Xing, M.; Tang, Y.; Ma, Y.; Li, L.; Yao, B.; Wu, H.; et al. YTHDF2-mediated FGF14-AS2 decay promotes osteolytic metastasis of breast cancer by enhancing RUNX2 mRNA translation. Br. J. Cancer 2022, 127, 2141–2153. [Google Scholar] [CrossRef]
- Yuan, W.; Chen, S.; Li, B.; Han, X.; Meng, B.; Zou, Y.; Chang, S. The N6-methyladenosine reader protein YTHDC2 promotes gastric cancer progression via enhancing YAP mRNA translation. Transl. Oncol. 2022, 16, 101308. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Doxtader, K.A.; Nam, Y. Structural Basis for Cooperative Function of Mettl3 and Mettl14 Methyltransferases. Mol. Cell 2016, 63, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gregory, R.I. Mining for METTL3 inhibitors to suppress cancer. Nat. Struct. Mol. Biol. 2021, 28, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, L.; Peng, D.; Jiang, A.; He, Y.; Zeng, Y.; Xie, C.; Zhou, H.; Luo, X.; Liu, H.; et al. METTL3 and N6-Methyladenosine Promote Homologous Recombination-Mediated Repair of DSBs by Modulating DNA-RNA Hybrid Accumulation. Mol. Cell 2020, 79, 425–442 e427. [Google Scholar] [CrossRef]
- Zhu, Y.; Peng, X.; Zhou, Q.; Tan, L.; Zhang, C.; Lin, S.; Long, M. METTL3-mediated m6A modification of STEAP2 mRNA inhibits papillary thyroid cancer progress by blocking the Hedgehog signaling pathway and epithelial-to-mesenchymal transition. Cell Death Dis. 2022, 13, 358. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Sheng, L.; Gao, Q.; Xiong, Q.; Zhang, H.; Wu, M.; Liang, Y.; Zhu, F.; Zhang, Y.; Zhang, X.; et al. The m(6)A methyltransferase METTL3 promotes bladder cancer progression via AFF4/NF-kappaB/MYC signaling network. Oncogene 2019, 38, 3667–3680. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, J.Z.; Yang, X.; Yu, H.; Zhou, R.; Lu, H.C.; Yuan, W.B.; Lu, J.C.; Zhou, Z.J.; Lu, Q.; et al. METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner. Mol. Cancer 2019, 18, 110. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Choe, J.; Du, P.; Triboulet, R.; Gregory, R.I. The m(6)A Methyltransferase METTL3 Promotes Translation in Human Cancer Cells. Mol. Cell 2016, 62, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Guo, X.; Li, L.; Gao, Z.; Su, X.; Ji, M.; Liu, J. N(6)-methyladenosine METTL3 promotes cervical cancer tumorigenesis and Warburg effect through YTHDF1/HK2 modification. Cell Death Dis. 2020, 11, 911. [Google Scholar] [CrossRef]
- Li, H.; Zhong, Y.; Cao, G.; Shi, H.; Liu, Y.; Li, L.; Yin, P.; Chen, J.; Xiao, Z.; Du, B. METTL3 promotes cell cycle progression via m(6)A/YTHDF1-dependent regulation of CDC25B translation. Int. J. Biol. Sci. 2022, 18, 3223–3236. [Google Scholar] [CrossRef]
- Jin, D.; Guo, J.; Wu, Y.; Du, J.; Yang, L.; Wang, X.; Di, W.; Hu, B.; An, J.; Kong, L.; et al. m(6)A mRNA methylation initiated by METTL3 directly promotes YAP translation and increases YAP activity by regulating the MALAT1-miR-1914-3p-YAP axis to induce NSCLC drug resistance and metastasis. J. Hematol. Oncol. 2019, 12, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, X.; Wang, Z.; Lu, W.; Miao, L.; Zhao, Y. METTL3 promotes non-small cell lung cancer (NSCLC) cell proliferation and colony formation in a m6A-YTHDF1 dependent way. BMC Pulm Med 2022, 22, 324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, Y.; Chen, H.; Sun, L.N.; Zhang, B.; Yue, D.S.; Wang, C.L.; Zhang, Z.F. METTL3-induced DLGAP1-AS2 promotes non-small cell lung cancer tumorigenesis through m(6)A/c-Myc-dependent aerobic glycolysis. Cell Cycle 2022, 21, 2602–2614. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yi, Y.; Miao, Y.; Long, W.; Long, T.; Chen, S.; Cheng, W.; Zou, C.; Zheng, Y.; Wu, X.; et al. N(6)-Methyladenosine Modulates Nonsense-Mediated mRNA Decay in Human Glioblastoma. Cancer Res. 2019, 79, 5785–5798. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.X.; Cao, Z.G.; Zhao, S.L. m6A methyltransferase METTL3 promotes the progression of prostate cancer via m6A-modified LEF1. Eur. Rev. Med. Pharm. Sci. 2020, 24, 3565–3571. [Google Scholar] [CrossRef]
- Yuan, D.; Chen, J.; Hao, Q.; Zhang, P.; Chen, Z. Methyltransferase-like 3 Aggravates HCC Development via Mediating N6-Methyladenosine of Ubiquitin-Specific Protease 7. J. Oncol. 2022, 2022, 6167832. [Google Scholar] [CrossRef]
- Hu, J.; Yang, L.; Peng, X.; Mao, M.; Liu, X.; Song, J.; Li, H.; Chen, F. METTL3 promotes m6A hypermethylation of RBM14 via YTHDF1 leading to the progression of hepatocellular carcinoma. Hum. Cell 2022, 35, 1838–1855. [Google Scholar] [CrossRef]
- Ai, Y.; Liu, S.; Luo, H.; Wu, S.; Wei, H.; Tang, Z.; Li, X.; Lv, X.; Zou, C. METTL3 Intensifies the Progress of Oral Squamous Cell Carcinoma via Modulating the m6A Amount of PRMT5 and PD-L1. J. Immunol. Res. 2021, 2021, 6149558. [Google Scholar] [CrossRef]
- Zhao, W.; Cui, Y.; Liu, L.; Ma, X.; Qi, X.; Wang, Y.; Liu, Z.; Ma, S.; Liu, J.; Wu, J. METTL3 Facilitates Oral Squamous Cell Carcinoma Tumorigenesis by Enhancing c-Myc Stability via YTHDF1-Mediated m(6)A Modification. Mol. Ther. Nucleic Acids 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Vu, L.P.; Pickering, B.F.; Cheng, Y.; Zaccara, S.; Nguyen, D.; Minuesa, G.; Chou, T.; Chow, A.; Saletore, Y.; MacKay, M.; et al. The N(6)-methyladenosine (m(6)A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat. Med. 2017, 23, 1369–1376. [Google Scholar] [CrossRef]
- Li, Y.; Wang, N.X.; Yin, C.; Jiang, S.S.; Li, J.C.; Yang, S.Y. RNA Editing Enzyme ADAR1 Regulates METTL3 in an Editing Dependent Manner to Promote Breast Cancer Progression via METTL3/ARHGAP5/YTHDF1 Axis. Int. J. Mol. Sci. 2022, 23, 9656. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.Q.; Zhang, K.; Sheng, J.; Ning, Z.Y.; Li, Y.; Shi, W.D.; Liu, L.M. NUCB1 Suppresses Growth and Shows Additive Effects With Gemcitabine in Pancreatic Ductal Adenocarcinoma via the Unfolded Protein Response. Front. Cell Dev. Biol. 2021, 9, 641836. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wei, J.; Gu, X.Z.; Yao, X.M.; Zhang, D.S.; Li, F.; Sun, Y.Y. Dysregulation of LINC00470 and METTL3 promotes chemoresistance and suppresses autophagy of chronic myelocytic leukaemia cells. J. Cell. Mol. Med. 2021, 25, 4248–4259. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Chen, B.; Wei, W.; Guo, S.; Han, H.; Yang, C.; Ma, J.; Wang, L.; Peng, S.; Kuang, M.; et al. N(6)-methyladenosine (m(6)A) in 18S rRNA promotes fatty acid metabolism and oncogenic transformation. Nat. Metab. 2022, 4, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, H.; Pan, R.; Wang, S.; Khan, A.A.; Zhao, Y.; Zhu, H.; Liu, X. Ribosome 18S m(6)A methyltransferase METTL5 promotes pancreatic cancer progression by modulating c-Myc translation. Int. J. Oncol. 2022, 60. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, S.; Zhang, G.; Liu, J.; Cao, G. Knockdown of METTL5 inhibits the Myc pathway to downregulate PD-L1 expression and inhibits immune escape of hepatocellular carcinoma cells. J. Chemother. 2022, 1–10. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, W.; Hao, C.; He, Q.; Tu, X. The feedback loop of METTL14 and USP38 regulates cell migration, invasion and EMT as well as metastasis in bladder cancer. PLoS Genet. 2022, 18, e1010366. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, X.; Qiao, S. METTL14-mediated N6-methyladenosine modification of Pten mRNA inhibits tumour progression in clear-cell renal cell carcinoma. Br. J. Cancer 2022, 127, 30–42. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, S.; Cui, Y.H.; Wei, J.; Shah, P.; Park, G.; Cui, X.; He, C.; He, Y.Y. METTL14 facilitates global genome repair and suppresses skin tumorigenesis. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Liu, T.; Zhang, Z.; Piao, C.; Ning, H. METTL14 promotes migration and invasion of choroidal melanoma by targeting RUNX2 mRNA via m6A modification. J. Cell Mol. Med. 2022, 26, 5602–5613. [Google Scholar] [CrossRef]
- Weng, H.; Huang, H.; Wu, H.; Qin, X.; Zhao, B.S.; Dong, L.; Shi, H.; Skibbe, J.; Shen, C.; Hu, C.; et al. METTL14 Inhibits Hematopoietic Stem/Progenitor Differentiation and Promotes Leukemogenesis via mRNA m(6)A Modification. Cell Stem. Cell 2018, 22, 191–205 e199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.; Wu, J.; Zhang, F. METTL16 epigenetically enhances GPX4 expression via m6A modification to promote breast cancer progression by inhibiting ferroptosis. Biochem. Biophys. Res. Commun. 2023, 638, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.Z.; Liu, Y.D.; Li, J.; Chen, M.T.; Huang, M.; Wang, F.; Yang, Q.S.; Yuan, J.H.; Sun, S.H. METTL16 promotes hepatocellular carcinoma progression through downregulating RAB11B-AS1 in an m(6)A-dependent manner. Cell Mol. Biol. Lett. 2022, 27, 41. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, Z.; Cai, Y.; Bi, P.; Liang, Q.; Yan, Y.; Xu, Z. YTH domain family: Potential prognostic targets and immune-associated biomarkers in hepatocellular carcinoma. Aging (Albany NY) 2021, 13, 24205–24218. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.; Vagbo, C.B.; Jakobsson, M.E.; Kim, Y.; Baltissen, M.P.; O’Donohue, M.F.; Guzman, U.H.; Malecki, J.M.; Wu, J.; Kirpekar, F.; et al. The human methyltransferase ZCCHC4 catalyses N6-methyladenosine modification of 28S ribosomal RNA. Nucleic Acids Res. 2020, 48, 830–846. [Google Scholar] [CrossRef] [PubMed]
- Turkalj, E.M.; Vissers, C. The emerging importance of METTL5-mediated ribosomal RNA methylation. Exp. Mol. Med. 2022, 54, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Rong, B.; Zhang, Q.; Wan, J.; Xing, S.; Dai, R.; Li, Y.; Cai, J.; Xie, J.; Song, Y.; Chen, J.; et al. Ribosome 18S m(6)A Methyltransferase METTL5 Promotes Translation Initiation and Breast Cancer Cell Growth. Cell Rep. 2020, 33, 108544. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Shi, H.; Ye, P.; Li, L.; Qu, Q.; Sun, G.; Sun, G.; Lu, Z.; Huang, Y.; Yang, C.G.; et al. m(6)A RNA Methylation Regulates the Self-Renewal and Tumorigenesis of Glioblastoma Stem Cells. Cell Rep. 2017, 18, 2622–2634. [Google Scholar] [CrossRef]
- Yang, F.; Yuan, W.Q.; Li, J.; Luo, Y.Q. Knockdown of METTL14 suppresses the malignant progression of non-small cell lung cancer by reducing Twist expression. Oncol. Lett. 2021, 22, 847. [Google Scholar] [CrossRef]
- Chen, P.; Liu, X.Q.; Lin, X.; Gao, L.Y.; Zhang, S.; Huang, X. Targeting YTHDF1 effectively re-sensitizes cisplatin-resistant colon cancer cells by modulating GLS-mediated glutamine metabolism. Mol. Ther. Oncolytics 2021, 20, 228–239. [Google Scholar] [CrossRef]
- Chen, H.; Yu, Y.; Yang, M.; Huang, H.; Ma, S.; Hu, J.; Xi, Z.; Guo, H.; Yao, G.; Yang, L.; et al. YTHDF1 promotes breast cancer progression by facilitating FOXM1 translation in an m6A-dependent manner. Cell Biosci. 2022, 12, 19. [Google Scholar] [CrossRef]
- Yarmishyn, A.A.; Yang, Y.P.; Lu, K.H.; Chen, Y.C.; Chien, Y.; Chou, S.J.; Tsai, P.H.; Ma, H.I.; Chien, C.S.; Chen, M.T.; et al. Musashi-1 promotes cancer stem cell properties of glioblastoma cells via upregulation of YTHDF1. Cancer Cell Int. 2020, 20, 597. [Google Scholar] [CrossRef]
- Lou, X.; Ning, J.; Liu, W.; Li, K.; Qian, B.; Xu, D.; Wu, Y.; Zhang, D.; Cui, W. YTHDF1 Promotes Cyclin B1 Translation through m(6)A Modulation and Contributes to the Poor Prognosis of Lung Adenocarcinoma with KRAS/TP53 Co-Mutation. Cells 2021, 10, 1669. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, P.; Zheng, X.; Wang, J.; Peng, Q.; Li, Z.; Wei, L.; Liu, C.; Wu, Y.; Wen, Y.; et al. N6-methyladenosine regulates ATM expression and downstream signaling. J. Cancer 2021, 12, 7041–7051. [Google Scholar] [CrossRef]
- Pi, J.; Wang, W.; Ji, M.; Wang, X.; Wei, X.; Jin, J.; Liu, T.; Qiang, J.; Qi, Z.; Li, F.; et al. YTHDF1 Promotes Gastric Carcinogenesis by Controlling Translation of FZD7. Cancer Res. 2021, 81, 2651–2665. [Google Scholar] [CrossRef]
- Bai, Y.; Yang, C.; Wu, R.; Huang, L.; Song, S.; Li, W.; Yan, P.; Lin, C.; Li, D.; Zhang, Y. YTHDF1 Regulates Tumorigenicity and Cancer Stem Cell-Like Activity in Human Colorectal Carcinoma. Front. Oncol. 2019, 9, 332. [Google Scholar] [CrossRef]
- Luo, X.; Cao, M.; Gao, F.; He, X. YTHDF1 promotes hepatocellular carcinoma progression via activating PI3K/AKT/mTOR signaling pathway and inducing epithelial-mesenchymal transition. Exp. Hematol. Oncol. 2021, 10, 35. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, L.; Wang, L.; Huang, W.; Tan, L.; Liu, H.; Huo, J.; Su, T.; Zhang, M.; Kuang, M.; et al. YTHDF1 promotes intrahepatic cholangiocarcinoma progression via regulating EGFR mRNA translation. J. Gastroenterol. Hepatol. 2022, 37, 1156–1168. [Google Scholar] [CrossRef]
- Chen, J.; Sun, Y.; Xu, X.; Wang, D.; He, J.; Zhou, H.; Lu, Y.; Zeng, J.; Du, F.; Gong, A.; et al. YTH domain family 2 orchestrates epithelial-mesenchymal transition/proliferation dichotomy in pancreatic cancer cells. Cell Cycle 2017, 16, 2259–2271. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Jiang, J.; Huang, Z.; Jin, P.; Peng, L.; Luo, M.; Zhang, Z.; Chen, Y.; Xie, N.; Gao, W.; et al. Hypoxia-induced lncRNA STEAP3-AS1 activates Wnt/beta-catenin signaling to promote colorectal cancer progression by preventing m(6)A-mediated degradation of STEAP3 mRNA. Mol. Cancer 2022, 21, 168. [Google Scholar] [CrossRef]
- Liu, X.; Liu, L.; Dong, Z.; Li, J.; Yu, Y.; Chen, X.; Ren, F.; Cui, G.; Sun, R. Expression patterns and prognostic value of m(6)A-related genes in colorectal cancer. Am. J. Transl. Res. 2019, 11, 3972–3991. [Google Scholar]
- Xu, Y.; He, X.; Wang, S.; Sun, B.; Jia, R.; Chai, P.; Li, F.; Yang, Y.; Ge, S.; Jia, R.; et al. The m(6)A reading protein YTHDF3 potentiates tumorigenicity of cancer stem-like cells in ocular melanoma through facilitating CTNNB1 translation. Oncogene 2022, 41, 1281–1297. [Google Scholar] [CrossRef]
- Yuan, Y.; Yan, G.; He, M.; Lei, H.; Li, L.; Wang, Y.; He, X.; Li, G.; Wang, Q.; Gao, Y.; et al. ALKBH5 suppresses tumor progression via an m(6)A-dependent epigenetic silencing of pre-miR-181b-1/YAP signaling axis in osteosarcoma. Cell Death Dis. 2021, 12, 60. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Li, J.; Chen, Z.; Chen, F.; Tu, J.; Lin, S.; Wang, H. N(6)-methyladenosine regulates glycolysis of cancer cells through PDK4. Nat. Commun. 2020, 11, 2578. [Google Scholar] [CrossRef]
- Li, Z.; Weng, H.; Su, R.; Weng, X.; Zuo, Z.; Li, C.; Huang, H.; Nachtergaele, S.; Dong, L.; Hu, C.; et al. FTO Plays an Oncogenic Role in Acute Myeloid Leukemia as a N(6)-Methyladenosine RNA Demethylase. Cancer Cell 2017, 31, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Romero-Garcia, S.; Prado-Garcia, H.; Carlos-Reyes, A. Role of DNA Methylation in the Resistance to Therapy in Solid Tumors. Front. Oncol. 2020, 10, 1152. [Google Scholar] [CrossRef]
- Liu, Z.; Zou, H.; Dang, Q.; Xu, H.; Liu, L.; Zhang, Y.; Lv, J.; Li, H.; Zhou, Z.; Han, X. Biological and pharmacological roles of m6A modifications in cancer drug resistance. Mol. Cancer 2022, 21, 220. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, Q.; Chen, M.; Li, B.; Wang, N.; Li, C.; Gao, Z.; Zhang, D.; Yang, L.; Xu, Z.; et al. N(6)-methyladenosine modification regulates imatinib resistance of gastrointestinal stromal tumor by enhancing the expression of multidrug transporter MRP1. Cancer Lett. 2022, 530, 85–99. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhou, Y.; Chen, Y.; Zhou, Z.; Liu, W.; Zheng, L.; Pei, Q.; Tan, F.; Pei, H.; Li, Y. m(6)A Methyltransferase KIAA1429 Regulates the Cisplatin Sensitivity of Gastric Cancer Cells via Stabilizing FOXM1 mRNA. Cancers 2022, 14, 5025. [Google Scholar] [CrossRef]
- Li, M.; Ye, J.; Xia, Y.; Li, M.; Li, G.; Hu, X.; Su, X.; Wang, D.; Zhao, X.; Lu, F.; et al. METTL3 mediates chemoresistance by enhancing AML homing and engraftment via ITGA4. Leukemia 2022, 36, 2586–2595. [Google Scholar] [CrossRef]
- Uddin, M.B.; Roy, K.R.; Hosain, S.B.; Khiste, S.K.; Hill, R.A.; Jois, S.D.; Zhao, Y.; Tackett, A.J.; Liu, Y.Y. An N(6)-methyladenosine at the transited codon 273 of p53 pre-mRNA promotes the expression of R273H mutant protein and drug resistance of cancer cells. Biochem. Pharm. 2019, 160, 134–145. [Google Scholar] [CrossRef]
- Ji, Y.; Zhao, Q.; Feng, W.; Peng, Y.; Hu, B.; Chen, Q. N6-Methyladenosine Modification of CIRCKRT17 Initiated by METTL3 Promotes Osimertinib Resistance of Lung Adenocarcinoma by EIF4A3 to Enhance YAP1 Stability. Cancers 2022, 14, 5582. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, T.; Yang, Y.; Tu, W.; Huang, H.; Wang, Y.; Chen, Y.; Pan, K.; Chen, Z. N(6)-methyladenosine-mediated LDHA induction potentiates chemoresistance of colorectal cancer cells through metabolic reprogramming. Theranostics 2022, 12, 4802–4817. [Google Scholar] [CrossRef]
- Taketo, K.; Konno, M.; Asai, A.; Koseki, J.; Toratani, M.; Satoh, T.; Doki, Y.; Mori, M.; Ishii, H.; Ogawa, K. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int. J. Oncol. 2018, 52, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Cui, S. METTL3-mediated m6A modification of lnc RNA RHPN1-AS1 enhances cisplatin resistance in ovarian cancer by activating PI3K/AKT pathway. J. Clin. Lab. Anal. 2022, 36, e24761. [Google Scholar] [CrossRef]
- Qiao, X.; Zhu, L.; Song, R.; Shang, C.; Guo, Y. METTL3/14 and IL-17 signaling contribute to CEBPA-DT enhanced oral cancer cisplatin resistance. Oral. Dis. 2021. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Lan, L.; Yan, L.; Li, W.; Evans, I.; Ruiz, E.J.; Su, Q.; Zhao, G.; Wu, W.; et al. METTL3 promotes oxaliplatin resistance of gastric cancer CD133+ stem cells by promoting PARP1 mRNA stability. Cell Mol. Life Sci. 2022, 79, 135. [Google Scholar] [CrossRef]
- Kong, F.; Liu, X.; Zhou, Y.; Hou, X.; He, J.; Li, Q.; Miao, X.; Yang, L. Downregulation of METTL14 increases apoptosis and autophagy induced by cisplatin in pancreatic cancer cells. Int. J. Biochem. Cell Biol. 2020, 122, 105731. [Google Scholar] [CrossRef]
- Hao, L.; Wang, J.M.; Liu, B.Q.; Yan, J.; Li, C.; Jiang, J.Y.; Zhao, F.Y.; Qiao, H.Y.; Wang, H.Q. m6A-YTHDF1-mediated TRIM29 upregulation facilitates the stem cell-like phenotype of cisplatin-resistant ovarian cancer cells. Biochim. Biophy.s Acta Mol. Cell Res. 2021, 1868, 118878. [Google Scholar] [CrossRef]
- Sun, Y.; Dong, D.; Xia, Y.; Hao, L.; Wang, W.; Zhao, C. YTHDF1 promotes breast cancer cell growth, DNA damage repair and chemoresistance. Cell Death Dis. 2022, 13, 230. [Google Scholar] [CrossRef]
- Song, Z.; Wang, X.; Chen, F.; Chen, Q.; Liu, W.; Yang, X.; Zhu, X.; Liu, X.; Wang, P. LncRNA MALAT1 regulates METTL3-mediated PD-L1 expression and immune infiltrates in pancreatic cancer. Front. Oncol. 2022, 12, 1004212. [Google Scholar] [CrossRef]
- Wan, W.; Ao, X.; Chen, Q.; Yu, Y.; Ao, L.; Xing, W.; Guo, W.; Wu, X.; Pu, C.; Hu, X.; et al. METTL3/IGF2BP3 axis inhibits tumor immune surveillance by upregulating N(6)-methyladenosine modification of PD-L1 mRNA in breast cancer. Mol. Cancer 2022, 21, 60. [Google Scholar] [CrossRef]
- Ni, Z.; Sun, P.; Zheng, J.; Wu, M.; Yang, C.; Cheng, M.; Yin, M.; Cui, C.; Wang, G.; Yuan, L.; et al. JNK Signaling Promotes Bladder Cancer Immune Escape by Regulating METTL3-Mediated m6A Modification of PD-L1 mRNA. Cancer Res. 2022, 82, 1789–1802. [Google Scholar] [CrossRef]
- Shu, F.; Xiao, H.; Li, Q.N.; Ren, X.S.; Liu, Z.G.; Hu, B.W.; Wang, H.S.; Wang, H.; Jiang, G.M. Epigenetic and post-translational modifications in autophagy: Biological functions and therapeutic targets. Signal Transduct. Target Ther. 2023, 8, 32. [Google Scholar] [CrossRef]
- Jin, S.; Zhang, X.; Miao, Y.; Liang, P.; Zhu, K.; She, Y.; Wu, Y.; Liu, D.A.; Huang, J.; Ren, J.; et al. m6A RNA modification controls autophagy through upregulating ULK1 protein abundance. Cell Res. 2018, 28, 955–957. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Cui, J.; Wang, H.; Ma, L.; Zhang, X.; Guo, W.; Xue, X.; Wang, Y.; Qiu, S.; Tian, X.; et al. IGF2BP3 is an essential N6-methyladenosine biotarget for suppressing ferroptosis in lung adenocarcinoma cells. Mater. Today Bio. 2022, 17, 100503. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liao, Y.; Zhang, M.; Zhu, Y.; Wang, W.; Cai, H.; Liang, J.; Song, F.; Hou, C.; Huang, S.; et al. N6-methyladenosine demethyltransferase FTO-mediated autophagy in malignant development of oral squamous cell carcinoma. Oncogene 2021, 40, 3885–3898. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zheng, H.; Liu, F.; Wu, Q.; Liu, S. The m6A methyltransferase METTL3 affects autophagy and progression of nasopharyngeal carcinoma by regulating the stability of lncRNA ZFAS1. Infect. Agent Cancer 2022, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ni, Y.; Zhang, L.; Jiang, R.; Xu, J.; Yang, H.; Hu, Y.; Qiu, J.; Pu, L.; Tang, J.; et al. HIF-1alpha-induced expression of m6A reader YTHDF1 drives hypoxia-induced autophagy and malignancy of hepatocellular carcinoma by promoting ATG2A and ATG14 translation. Signal Transduct. Target Ther. 2021, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Deng, L.; Huang, N.; Cui, Z.; Wu, Q.; Ma, J.; Pan, Q.; Sun, F. m(6)A mRNA Methylation Regulates LKB1 to Promote Autophagy of Hepatoblastoma Cells through Upregulated Phosphorylation of AMPK. Genes 2021, 12, 1747. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. m(6)A mRNA methylation controls autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy 2020, 16, 1221–1235. [Google Scholar] [CrossRef]
- Song, H.; Feng, X.; Zhang, H.; Luo, Y.; Huang, J.; Lin, M.; Jin, J.; Ding, X.; Wu, S.; Huang, H.; et al. METTL3 and ALKBH5 oppositely regulate m(6)A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes. Autophagy 2019, 15, 1419–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, W.; Dian, M.; Wang, J.; Sun, Y.; Xiao, D. Epitranscriptomic turbo for autophagy boost: M(6)A reader YTHDF3. Autophagy 2022, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhu, Y.; Cai, H.; Liang, J.; Wang, W.; Liao, Y.; Zhang, Y.; Wang, C.; Hou, J. N6-Methyladenosine Methyltransferase METTL14-Mediated Autophagy in Malignant Development of Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 738406. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Dian, M.; Zhou, Y.; Zhong, Q.; Pang, W.; Li, Z.; Zhao, Y.; Ma, J.; Lin, X.; Luo, R.; et al. Autophagy induction promoted by m(6)A reader YTHDF3 through translation upregulation of FOXO3 mRNA. Nat. Commun. 2022, 13, 5845. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, Y.; Hu, D. Knockdown of METTL3 inhibits enterovirus 71-induced apoptosis of mouse Schwann cell through regulation of autophagy. Pathog. Dis. 2021, 79. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Zheng, S.; Lin, S. N(7)-methylguanosine (m(7)G) tRNA modification: A novel autophagy modulator in cancer. Autophagy 2023, 19, 360–362. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, S.; Ma, B. Autophagy and autophagy-related proteins in cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef]
- Yeon, M.; Kim, Y.; Pathak, D.; Kwon, E.; Kim, D.Y.; Jeong, M.S.; Jung, H.S.; Jeoung, D. The CAGE-MiR-181b-5p-S1PR1 Axis Regulates Anticancer Drug Resistance and Autophagy in Gastric Cancer Cells. Front. Cell Dev. Biol. 2021, 9, 666387. [Google Scholar] [CrossRef]
- Yeon, M.; Byun, J.; Kim, H.; Kim, M.; Jung, H.S.; Jeon, D.; Kim, Y.; Jeoung, D. CAGE Binds to Beclin1, Regulates Autophagic Flux and CAGE-Derived Peptide Confers Sensitivity to Anti-cancer Drugs in Non-small Cell Lung Cancer Cells. Front. Oncol. 2018, 8, 599. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Zhu, M.; Wang, Q.; Zhou, X.; Wang, J.; Wang, T.; Zhang, J. DARS-AS1 recruits METTL3/METTL14 to bind and enhance DARS mRNA m(6)A modification and translation for cytoprotective autophagy in cervical cancer. RNA Biol. 2022, 19, 751–763. [Google Scholar] [CrossRef]
- Liu, S.; Li, Q.; Li, G.; Zhang, Q.; Zhuo, L.; Han, X.; Zhang, M.; Chen, X.; Pan, T.; Yan, L.; et al. The mechanism of m(6)A methyltransferase METTL3-mediated autophagy in reversing gefitinib resistance in NSCLC cells by beta-elemene. Cell Death Dis. 2020, 11, 969. [Google Scholar] [CrossRef]
- Lv, D.; Gimple, R.C.; Zhong, C.; Wu, Q.; Yang, K.; Prager, B.C.; Godugu, B.; Qiu, Z.; Zhao, L.; Zhang, G.; et al. PDGF signaling inhibits mitophagy in glioblastoma stem cells through N(6)-methyladenosine. Dev. Cell 2022, 57, 1466–1481 e1466. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiang, Y.; Yin, Y.; Peng, J.; Peng, D.; Li, D.; Kitazawa, R.; Tang, Y.; Yang, J. The m6A methyltransferase METTL3 regulates autophagy and sensitivity to cisplatin by targeting ATG5 in seminoma. Transl. Androl. Urol. 2021, 10, 1711–1722. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhu, Y.; Han, S.; Chen, M.; Song, P.; Dai, D.; Xu, W.; Jiang, T.; Feng, L.; Shin, V.Y.; et al. Impaired autophagic degradation of lncRNA ARHGAP5-AS1 promotes chemoresistance in gastric cancer. Cell Death Dis. 2019, 10, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Gao, L.X.; Wang, W.; Zhang, T.; Dong, F.Y.; Ding, W.P. M(6) A demethylase fat mass and obesity-associated protein regulates cisplatin resistance of gastric cancer by modulating autophagy activation through ULK1. Cancer Sci. 2022, 113, 3085–3096. [Google Scholar] [CrossRef] [PubMed]
- Lothion-Roy, J.; Haigh, D.B.; Harris, A.E.; Metzler, V.M.; Alsaleem, M.; Toss, M.S.; Kariri, Y.; Ntekim, A.; Robinson, B.D.; Khani, F.; et al. Clinical and molecular significance of the RNA m6A methyltransferase complex in prostate cancer. Front. Genet. 2023, 13, 1096071. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Xu, L.; Dai, X.; Zhu, M.; Chen, X.; Li, Y.; Li, J.; Ge, R.; Cheng, B.; Wang, Y. Prognostic and clinicopathological value of m6A regulators in human cancers: A meta-analysis. Aging (Albany NY) 2022, 14, 8818–8838. [Google Scholar] [CrossRef]
- Chen, S.; Su, X.; Wang, J.; Zheng, N.; Tang, Y.; Peng, G.; Dong, R.; Lu, F.; Liu, M.; Zhao, Y.; et al. Identification and Validation of METTL3-Related Molecules for Predicting Prognosis and Efficacy of Immunotherapy in Gastric Cancer Based on m6A Methylome and Transcriptome Sequencing Analysis. Front. Oncol. 2022, 12, 935239. [Google Scholar] [CrossRef]
- Qiu, X.; Yang, S.; Wang, S.; Wu, J.; Zheng, B.; Wang, K.; Shen, S.; Jeong, S.; Li, Z.; Zhu, Y.; et al. M6A Demethylase ALKBH5 Regulates PD-L1 Expression and Tumor Immunoenvironment in Intrahepatic Cholangiocarcinoma. Cancer Res. 2021, 81, 4778–4793. [Google Scholar] [CrossRef]
- Mao, W.; Yu, Q.; Wang, K.; Ma, Q.; Zheng, Y.; Zhang, G.; Luo, W.; Wang, N.; Wang, Y. Comprehensive Analysis of the Transcriptome-wide m6A Methylome in Lung Adenocarcinoma by MeRIP Sequencing. Front. Oncol. 2022, 12, 791332. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.B.; Hu, B.B.; He, R.; He, L.; Zou, C.; Man, C.F.; Fan, Y. Analysis of N6-Methyladenosine Methylome in Adenocarcinoma of Esophagogastric Junction. Front. Genet. 2022, 12, 787800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, Q.; Zhang, M.; Zhang, W.; Zhao, L.; Yang, C.; Wang, B.; Jiang, K.; Ye, Y. Comprehensive analysis of the transcriptome-wide m6A methylome in colorectal cancer by MeRIP sequencing. Epigenetics 2021, 16, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Gan, Y.; Cao, C.; Ma, B.; Zhang, Q.; Zhang, Q.; Yao, L. Transcriptome-Wide Map of N6-Methyladenosine Methylome Profiling in Human Bladder Cancer. Front. Oncol. 2021, 11, 717622. [Google Scholar] [CrossRef]
- Deng, Y.; Duan, R.; Ding, W.; Gu, Q.; Liu, M.; Zhou, J.; Sun, J.; Zhu, J. Astrocyte-derived exosomal nicotinamide phosphoribosyltransferase (Nampt) ameliorates ischemic stroke injury by targeting AMPK/mTOR signaling to induce autophagy. Cell Death Dis. 2022, 13, 1057. [Google Scholar] [CrossRef]
- Chi, B.; Zheng, Y.; Xie, F.; Fu, W.; Wang, X.; Gu, J.; Yang, J.; Yin, J.; Cai, L.; Tang, P.; et al. Increased expression of miR-194-5p through the circPVRL3/miR-194-5p/SOCS2 axis promotes proliferation and metastasis in pancreatic ductal adenocarcinoma by activating the PI3K/AKT signaling pathway. Cancer Cell Int. 2022, 22, 415. [Google Scholar] [CrossRef]
- Huang, W.; Huang, T.; Liu, Y.; Fu, J.; Wei, X.; Liu, D.; Ma, W.; Gu, H.; Yuan, Z. Nuclear factor I-C disrupts cellular homeostasis between autophagy and apoptosis via miR-200b-Ambra1 in neural tube defects. Cell Death Dis. 2021, 13, 17. [Google Scholar] [CrossRef]
- Xuan, X.; Wang, Z.; Wang, Y. Circ_0058608 contributes to the progression and taxol resistance of non-small cell lung cancer by sponging miR-1299 to upregulate GBP1. Anticancer Drugs 2023, 34, 103–114. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Name | Functional Roles in m6A Regulation | Ref |
|---|---|---|---|
| m6A writers and regulators | VIRMA | Promotes m6A methylation of mRNAs in the 3′-UTR near the stop codons. A component of RNA methyl transferase complex | [24] |
| METTL3 | Induces RNA GAC, AAC methylation. A component of RNA methyl transferase complex | [25] | |
| METTL4 | Induces N6-methylation of snRNA | [26] | |
| METTL5 | Induces methylation of 18S rRNA, enhances translation | [27] | |
| METTL14 | Induces GAC methylation. A component of RNA methyl transferase complex | [28] | |
| METTL16 | Induces m6A methylation on S-adenosyl methionine (SAM) synthetase pre-mRNA | [29] | |
| WTAP | Enhances efficient binding of methyl transferase complex to RNA. A regulatory subunit of RNA methyl transferase complex | [30,31] | |
| RBM15/15B | Mediates X inactivation promoted by Xist. A component of m6A-METTL associated complex | [32] | |
| CBLL1 | A component of m6A-METTL associated complex | [33] | |
| ZC3H13 | Modulates RNA m6A methylation. A component of m6A-METTL associated complex | [34,35] | |
| ZCCHC4 | Acts as rRNA N6-methyl transferase | [34,35] | |
| m6A eraser | FTO | Involved in processing of miRNAs, RNA stability, RNA metabolism | [50,51,52] |
| ALKBH5 | mRNA export, methylation ↓ | [53] | |
| m6A binding proteins | IGF2BP1/2/3 | Enhances tumorigenesis by stabilizing S1PR3 | [60] |
| HNRNPs | mRNA splicing | [61,62] | |
| YTHDC1 | RNA splicing, X-inactivation | [63] | |
| YTHDF1 | Enhances protein translation | [67] | |
| YTHDF2 | RNA splicing, nuclear export, RNA degradation | [69] |
| Methylase | Target | Function | Mechanism | Cancer Type | Ref |
|---|---|---|---|---|---|
| METTL3 | STEAP2 | Tumor suppression | STEAP2 ↑, EMT ↑, Hedgehog signaling ↑ | Papillary thyroid cancer | [75] |
| NF-kB, MYC, AFF4 | Tumor promotion | NF-kB ↑, MYC ↑, AFF4 ↑ | Bladder cancer | [76] | |
| CDC25 | Tumor promotion | m6A CDC25B ↑ | Cervical cancer | [80] | |
| MALAT1 | Tumor promotion | m6A MALAT1 ↑ YAP ↑ | Non-small cell lung cancer | [81] | |
| DLGAP1-AS2 | Tumor promotion | m6ADLGAP1-AS2 ↑, c-MYC ↑ | Non-small cell lung cancer | [83] | |
| SRSF | Tumor promotion | m6ASRSF ↑ | Glioblastoma | [84] | |
| LEF1, Wnt signaling | Tumor promotion | LEF1 ↑, Wnt signaling ↑ | Prostate cancer | [85] | |
| C-Myc | Tumor promotion | C-Myc ↑, PRM5 ↑, PD-L1 ↑ | Oral squamous cell carcinoma | [89] | |
| C-Myc, BCL2 | Tumor promotion | C-Myc ↑, BCL2 ↑ | AML | [90] | |
| NUCB1 | Tumor promotion | NUCB1 ↓ | Pancreatic ductal adenocarcinoma | [92] | |
| METTL5 | ACSL4 | Tumor promotion | ACSL4 ↑ | Hepatocellular carcinoma | [94] |
| C-Myc | Tumor promotion | C-Myc ↑ | Pancreatic cancer | [95] | |
| C-Myc, PD-L1 | Tumor promotion | C-Myc ↑, PD-L1 ↑ | Hepatocellular carcinoma | [96] | |
| METTL14 | USP38 | Tumor suppression | USP38 ↑ | Bladder cancer | [97] |
| PTEN | Tumor suppression | PTEN ↑, PI3 kinase signaling ↓ | Clear cell renal cell carcinoma | [98] | |
| GGR, DDB2 | Tumor suppression | GGR ↑, DDB2 ↑ | Skin tumor | [99] | |
| Wnt/β-catenin signaling | Tumor promotion | Wnt/β-catenin signaling ↑ | Choroidal melanoma | [100] | |
| Myb, Myc | Tumor promotion | Myb ↑, Myc ↑ | AML | [101] | |
| METTL16 | GPX4 | Tumor promotion | GPX4 ↑ | Breast cancer | [102] |
| Cyclin D1 | Tumor promotion | Cyclin D1 ↑ | Gastric cancer | [103] | |
| RAB11B-AS1 | Tumor promotion | RAB11B-AS1 ↓ | Hepatocellular carcinoma | [104] |
| m6A-Binding Protein/m6A Demethylase | Target | Function | Mechanism | Cancer Type | Ref |
|---|---|---|---|---|---|
| IGF2BP3 | SLIT2 | Tumor promotion | SLIT2 ↓, PI3K/AKT ↑, MAPK ↑ | Breast cancer | [55] |
| IGF2BP2 | S1PR3 | Tumor promotion | S1PR3 ↑ | Renal cell carcinoma | [60] |
| YTHDF1 | eIF3 | Tumor promotion | eIF3 ↑ | Merkel cell carcinoma | [58] |
| YTHDF1 | Polo-like kinase/PI3K/AKT | Tumor promotion | Polo-like kinase/PI3K/AKT ↑ | Prostate cancer | [69] |
| YTHDF1 | FOXM1 | Tumor promotion | FOXM1 ↑ | Breast cancer | [111] |
| YTHDF1 | Cyclin B1 | Tumor promotion | Cyclin B1 ↑ | Lung adenocarcinoma | [113] |
| YTHDF1 | FZD7 | Tumor promotion | Wnt/β-catenin signaling ↑ | Gastric cancer | [115] |
| YTHDF1 | PI3K/Akt/mTOR | Tumor promotion | PI3K/Akt/mTOR signaling ↑ | Hepatocellular carcinoma | [117] |
| YTHDF1 | EGFR | Tumor promotion | EGFR ↑ | Intrahepatic cholangiocarcinoma | [118] |
| ALKBH5 | YAP, miR-181b-5p | Tumor suppression | YAP ↓, miR-181b-5p ↑ | Osteosarcoma | [123] |
| FTO | ASB2 PARA | Tumor promotion | ASB2 ↓ PARA ↓ | AML | [125] |
| Methylase/m6A Demethylase/m6A-Binding Protein | Function | Mechanism | Cancer Type | Ref |
|---|---|---|---|---|
| METTL14 | Enhances cisplatin resistance | m6A level of pri-miR-19a ↑ | Non-small cell lung cancer | [28] |
| Enhances cisplatin resistance | AMPKα/ERK1/2/mTOR signaling ↑ | Pancreatic cancer | [138] | |
| FTO | Enhances 5-FU resistance | SIVA-1 ↓ | Colorectal cancer | [64] |
| YTHDF1 | Enhances cisplatin synthesis | GLS1 ↑ | Colon cancer | [110] |
| Enhances cisplatin resistance | TRIM29 ↑ | Ovarian cancer | [139] | |
| KIAA1429 | Enhances cisplatin resistance | FOXM1 ↑ | Gastric cancer | [129] |
| METTL3 | Enhances cisplatin resistance | ITGA4 ↑ | AML | [130] |
| Enhances multidrug resistance | m6A installation at mutant p53 | Colon cancer | [131] | |
| Enhances osimertinib resistance | CircKRT17 ↑ | Lung cancer | [132] | |
| Enhances 5-FU resistance | Glycolysis ↑ | Colorectal cancer | [133] | |
| Enhances cisplatin resistance | RHPN1-AS1 ↑, PI3K/AKT↑ | Ovarian cancer | [135] | |
| Enhances oxaliplatin resistance | PARP1 ↑ | Gastric cancer | [137] | |
| Enhances resistance to anti-PD-L1 blockage | PD-L1 ↑ | Breast cancer | [142] | |
| METTL3/ METTL14 | Enhances cisplatin resistance | Functions as downstream targets of CEBPA-DT | Oral cancer | [136] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, H.; Shim, K.; Jeoung, D. Roles of RNA Methylations in Cancer Progression, Autophagy, and Anticancer Drug Resistance. Int. J. Mol. Sci. 2023, 24, 4225. https://doi.org/10.3390/ijms24044225
Jo H, Shim K, Jeoung D. Roles of RNA Methylations in Cancer Progression, Autophagy, and Anticancer Drug Resistance. International Journal of Molecular Sciences. 2023; 24(4):4225. https://doi.org/10.3390/ijms24044225
Chicago/Turabian StyleJo, Hyein, Kyeonghee Shim, and Dooil Jeoung. 2023. "Roles of RNA Methylations in Cancer Progression, Autophagy, and Anticancer Drug Resistance" International Journal of Molecular Sciences 24, no. 4: 4225. https://doi.org/10.3390/ijms24044225
APA StyleJo, H., Shim, K., & Jeoung, D. (2023). Roles of RNA Methylations in Cancer Progression, Autophagy, and Anticancer Drug Resistance. International Journal of Molecular Sciences, 24(4), 4225. https://doi.org/10.3390/ijms24044225