


Can T Cells Abort SARS-CoV-2 and Other Viral Infections?


{kind=link}
{kind=link}
Abstract
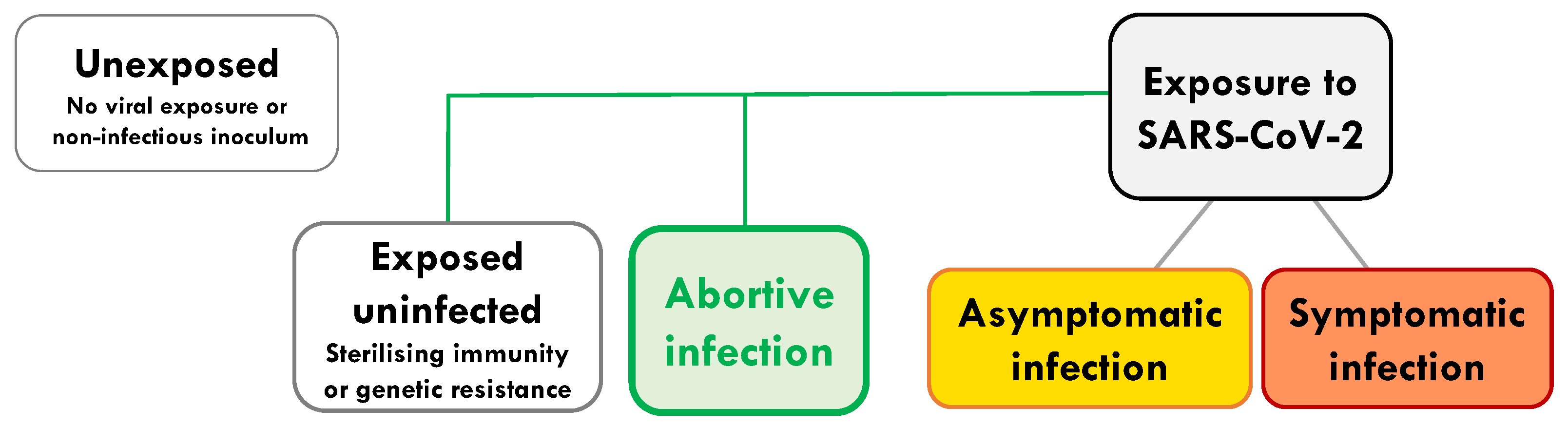
:1. Introduction
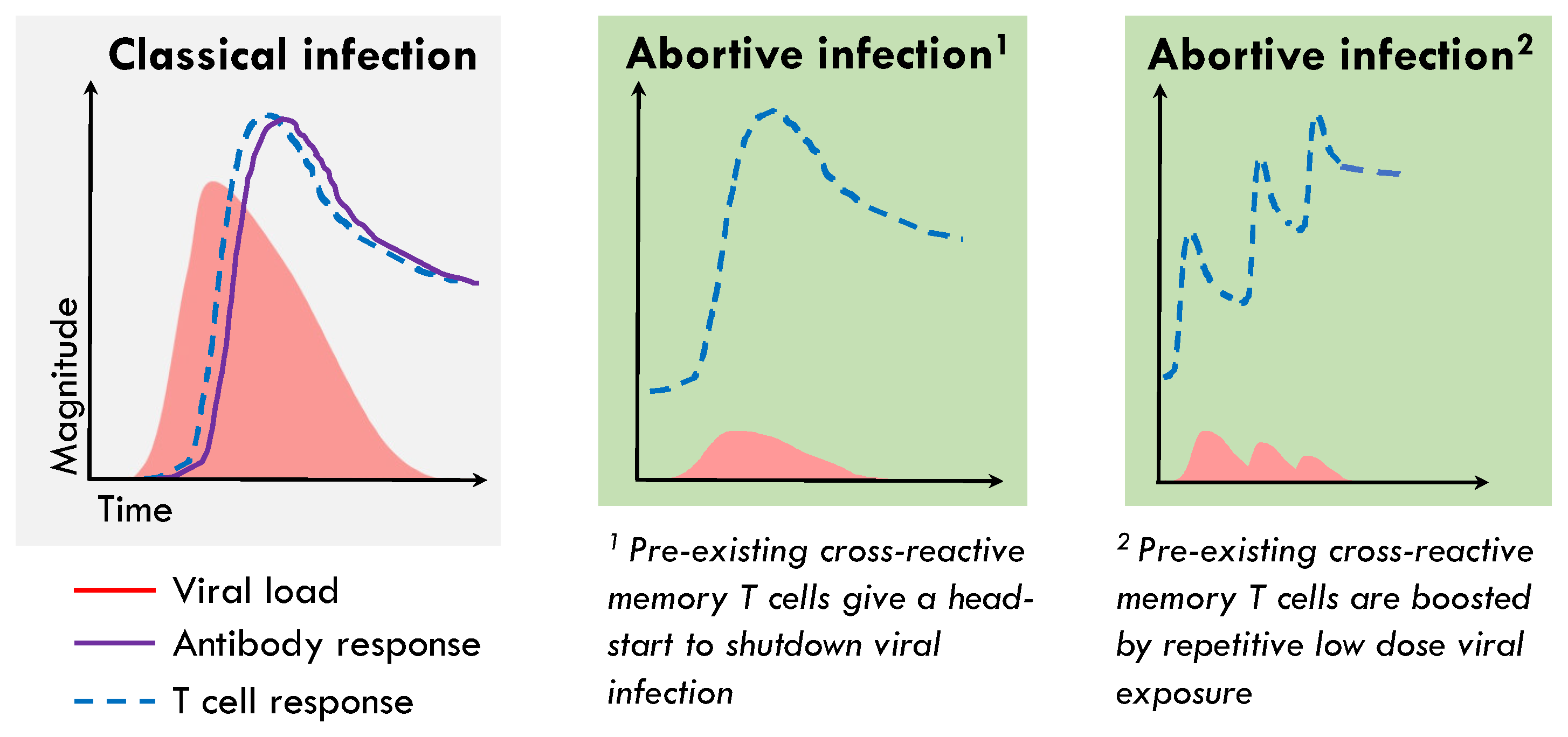
2. Characterisation of Abortive Infection by Early Sampling of a New Pandemic Virus
3. Evidence for Abortive, Seronegative Infection from Other SARS-CoV-2 Studies
4. Seronegative Infections: MERS, SARS-CoV and Beyond
5. Emerging/Unanswered Questions
5.1. Could the Antibody Response Have Been Missed in Seronegative Individuals?
5.2. Could T Cells Be an Epiphenomenon in Abortive Infection?
5.3. What Is the Influence of Variable Viral Inoculum on Outcome?
6. Conclusions: Translational Relevance of Identifying Abortive Infections
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACE-2 | angiotensin-converting enzyme 2 |
| CMV | cytomegalovirus |
| EBV | Epstein-Barr virus |
| ELISpot | enzyme-linked immunosorbent spot assay |
| HBV | hepatitis B virus |
| HBc | hepatitis B virus core protein |
| HBs | hepatitis B virus surface protein |
| HCV | hepatitis C virus |
| HCW | health care worker |
| HEPS | HIV-1-exposed persistently seronegative subjects |
| HIV | human immunodeficiency virus |
| HLA | human leukocyte antigen |
| HSV | herpes simplex virus |
| IFI27 | Interferon alpha inducible protein 27 |
| IFNγ | interferon gamma |
| IgA | Immunoglobulin A |
| IgG | Immunoglobulin G |
| IL-2 | interleukin 2 |
| MAIT | mucosal associated invariant T cells |
| MERS | middle east respiratory syndrome |
| MHC | major histocompatibility complex |
| NK | natural killer cells |
| NSP | non-structural protein |
| NSP7 | cofactor for RNA dependent RNA polymerase |
| NSP12 | RNA dependent RNA polymerase |
| NSP13 | Helicase |
| ORF | open reading frame |
| PBMC | peripheral blood mononuclear cells |
| PCR | polymerase chain reaction |
| PPE | personal protective equipment |
| RIG-I | retinoic acid-inducible gene I |
| RTC | replication-transcription complex |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| SIV | simian immunodeficiency virus |
| TCID50 | tissue culture infectious dose 50 |
References
- Bertoletti, A.; Tan, A.T.; le Bert, N. The T-cell response to SARS-CoV-2: Kinetic and quantitative aspects and the case for their protective role. Oxf. Open Immunol. 2021, 2, iqab006. [Google Scholar] [CrossRef]
- Grifoni, A.; Sette, A. From Alpha to omicron: The response of T cells. Curr. Res. Immunol. 2022, 3, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Niessl, J.; Sekine, T.; Buggert, M. T cell immunity to SARS-CoV-2. Semin. Immunol. 2021, 55, 101505. [Google Scholar] [CrossRef]
- Swadling, L.; Maini, M.K. T cells in COVID-19—United in diversity. Nat. Immunol. 2020, 21, 1307–1308. [Google Scholar] [CrossRef] [PubMed]
- Kent, S.J.; Khoury, D.S.; Reynaldi, A.; Juno, J.A.; Wheatley, A.K.; Stadler, E.; John Wherry, E.; Triccas, J.; Sasson, S.C.; Cromer, D.; et al. Disentangling the relative importance of T cell responses in COVID-19: Leading actors or supporting cast? Nat. Rev. Immunol. 2022, 22, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Hütter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Müßig, A.; Allers, K.; Schneider, T.; Hofmann, J.; Kücherer, C.; Blau, O.; et al. Long-Term Control of HIV by CCR5 Delta32/Delta32 Stem-Cell Transplantation. N. Engl. J. Med. 2009, 360, 692–698. [Google Scholar] [CrossRef] [Green Version]
- Bunyavanich, S.; Do, A.; Vicencio, A. Nasal Gene Expression of Angiotensin-Converting Enzyme 2 in Children and Adults. JAMA 2020, 323, 2427. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; van de Veerdonk, F.L.; van Crevel, R.; Pulendran, B.; van der Meer, J.W. Natural resistance against infections: Focus on COVID-19. Trends Immunol. 2022, 43, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Pires De Souza, G.A.; Le Bideau, M.; Boschi, C.; Wurtz, N.; Colson, P.; Aherfi, S.; Devaux, C.; La Scola, B. Choosing a cellular model to study SARS-CoV-2. Front. Cell. Infect. Microbiol. 2022, 12, 1583. [Google Scholar] [CrossRef]
- Balloux, F.; Tan, C.; Swadling, L.; Richard, D.; Jenner, C.; Maini, M.; van Dorp, L. The Past, Current and Future Epidemiological Dynamic of SARS-CoV-2. Oxf. Open Immunol. 2022, 3, iqac003. [Google Scholar] [CrossRef]
- Swadling, L.; Diniz, M.O.; Schmidt, N.M.; Amin, O.E.; Chandran, A.; Shaw, E.; Pade, C.; Gibbons, J.M.; Le Bert, N.; Tan, A.T.; et al. Pre-existing polymerase-specific T cells expand in abortive seronegative SARS-CoV-2. Nature 2021, 601, 110–117. [Google Scholar] [CrossRef]
- Moon, J.C.; Augusto, J.B.; Menacho, K.; Andiapen, M.; Bowles, R.; Burton, M.; Welch, S.; Bhuva, A.N.; Seraphim, A.; Pade, C.; et al. Healthcare Workers Bioresource: Study outline and baseline characteristics of a prospective healthcare worker cohort to study immune protection and pathogenesis in COVID-19. Wellcome Open Res. 2020, 5, 179. [Google Scholar]
- Treibel, T.A.; Manisty, C.; Burton, M.; McKnight, Á.; Lambourne, J.; Augusto, J.B.; Couto-Parada, X.; Cutino-Moguel, T.; Noursadeghi, M.; Moon, J.C. COVID-19: PCR screening of asymptomatic health-care workers at London hospital. Lancet 2020, 395, 1608–1610. [Google Scholar] [CrossRef]
- Reynolds, C.J.; Swadling, L.; Gibbons, J.M.; Pade, C.; Jensen, M.P.; Diniz, M.O.; Schmidt, N.M.; Butler, D.K.; Amin, O.E.; Bailey, S.N.; et al. Discordant neutralizing antibody and T cell responses in asymptomatic and mild SARS-CoV-2 infection. Sci. Immunol. 2020, 5, eabf3698. [Google Scholar] [CrossRef]
- Gupta, R.K.; Rosenheim, J.; Bell, L.C.; Chandran, A.; Guerra-Assuncao, J.A.; Pollara, G.; Whelan, M.; Artico, J.; Joy, G.; Kurdi, H.; et al. Blood transcriptional biomarkers of acute viral infection for detection of pre-symptomatic SARS-CoV-2 infection: A nested, case-control diagnostic accuracy study. Lancet Microbe 2021, 2, e508–e517. [Google Scholar] [CrossRef]
- Kaul, R.; Dong, T.; Plummer, F.A.; Kimani, J.; Rostron, T.; Kiama, P.; Njagi, E.; Irungu, E.; Farah, B.; Oyugi, J.; et al. CD8+ lymphocytes respond to different HIV epitopes in seronegative and infected subjects. J. Clin. Investig. 2001, 107, 1303–1310. [Google Scholar] [CrossRef]
- Heller, T.; Werner, J.M.; Rahman, F.; Mizukoshi, E.; Sobao, Y.; Gordon, A.M.; Sheets, A.; Sherker, A.H.; Kessler, E.; Bean, K.S.; et al. Occupational exposure to hepatitis C virus: Early T-cell responses in the absence of seroconversion in a longitudinal cohort study. J. Infect. Dis. 2013, 208, 1020–1025. [Google Scholar] [CrossRef] [Green Version]
- Werner, J.M.; Abdalla, A.; Gara, N.; Ghany, M.G.; Rehermann, B. The hepatitis B vaccine protects re-exposed health care workers, but does not provide sterilizing immunity. Gastroenterology 2013, 145, 1026–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland-Jones, S.; Sutton, J.; Ariyoshi, K.; Dong, T.; Gotch, F.; McAdam, S.; Whitby, D.; Sabally, S.; Gallimore, A.; Corrah, T.; et al. HIV-specific cytotoxic T-cells in HIV-exposed but uninfected Gambian women. Nat. Med. 1995, 1, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Promadej, N.; Costello, C.; Wernett, M.M. Broad human immunodeficiency virus (HIV)-specific T cell responses to conserved HIV proteins in HIV-seronegative women highly exposed to a single HIV-infected partner. J. Infect. Dis. 2003, 187, 1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munusamy Ponnan, S.; Thiruvengadam, K.; Kathirvel, S.; Shankar, J.; Rajaraman, A.; Mathaiyan, M.; Dinesha, T.R.; Poongulali, S.; Saravanan, S.; Murugavel, K.G.; et al. Elevated Numbers of HIV-Specific Poly-Functional CD8+ T Cells with Stem Cell-like and Follicular Homing Phenotypes in HIV-Exposed Seronegative Individuals. Front. Immunol. 2021, 12, 572. [Google Scholar] [CrossRef] [PubMed]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Li, C.K.; Wu, H.; Yan, H.; Ma, S.; Wang, L.; Zhang, M.; Tang, X.; Temperton, N.J.; Weiss, R.A.; Brenchley, J.M.; et al. T Cell Responses to Whole SARS Coronavirus in Humans. J. Immunol. 2008, 181, 5490–5500. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.C.S.; Owen, C.J.; Tham, C.Y.; Bertoletti, A.; van Dorp, L.; Balloux, F. Pre-existing T cell-mediated cross-reactivity to SARS-CoV-2 cannot solely be explained by prior exposure to endemic human coronaviruses. Infect. Genet. Evol. 2021, 95, 105075. [Google Scholar] [CrossRef] [PubMed]
- Loyal, L.; Braun, J.; Henze, L.; Kruse, B.; Dingeldey, M.; Reimer, U.; Kern, F.; Schwarz, T.; Mangold, M.; Unger, C.; et al. Cross-reactive CD4+ T cells enhance SARS-CoV-2 immune responses upon infection and vaccination. Science 2021, 374, eabh1823. [Google Scholar] [CrossRef]
- Diniz, M.O.; Mitsi, E.; Swadling, L.; Rylance, J.; Johnson, M.; Goldblatt, D.; Ferreira, D.; Maini, M.K. Airway-resident T cells from unexposed individuals cross-recognize SARS-CoV-2. Nat. Immunol. 2022, 23, 1324–1329. [Google Scholar] [CrossRef]
- Killingley, B.; Mann, A.J.; Kalinova, M.; Boyers, A.; Goonawardane, N.; Zhou, J.; Lindsell, K.; Hare, S.S.; Brown, J.; Frise, R.; et al. Safety, tolerability and viral kinetics during SARS-CoV-2 human challenge in young adults. Nat. Med. 2022, 28, 1031–1041. [Google Scholar] [CrossRef]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lübke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Schulien, I.; Kemming, J.; Oberhardt, V.; Wild, K.; Seidel, L.M.; Killmer, S.; Daul, F.; Salvat Lago, M.; Decker, A.; Luxenburger, H.; et al. Characterization of pre-existing and induced SARS-CoV-2-specific CD8+ T cells. Nat. Med. 2021, 27, 78–85. [Google Scholar] [CrossRef]
- Low, J.S.; Vaqueirinho, D.; Mele, F.; Foglierini, M.; Jerak, J.; Perotti, M.; Jarrossay, D.; Jovic, S.; Perez, L.; Cacciatore, R.; et al. Clonal analysis of immunodominance and crossreactivity of the CD4 T cell response to SARS-CoV-2. Science 2021, 372, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, X.; Zhong, J.; Zhou, Y.; Tang, Z.; Zhou, H.; He, J.; Mei, X.; Tang, Y.; Lin, B.; et al. Exposure to SARS-CoV-2 generates T-cell memory in the absence of a detectable viral infection. Nat. Commun. 2021, 12, 1724. [Google Scholar] [CrossRef] [PubMed]
- Ogbe, A.; Kronsteiner, B.; Skelly, D.T.; Pace, M.; Brown, A.; Adland, E.; Adair, K.; Akhter, H.D.; Ali, M.; Ali, S.E.; et al. T cell assays differentiate clinical and subclinical SARS-CoV-2 infections from cross-reactive antiviral responses. Nat. Commun. 2021, 12, 2055. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef]
- Kundu, R.; Narean, J.S.; Wang, L.; Fenn, J.; Pillay, T.; Fernandez, N.D.; Conibear, E.; Koycheva, A.; Davies, M.; Tolosa-Wright, M.; et al. Cross-reactive memory T cells associate with protection against SARS-CoV-2 infection in COVID-19 contacts. Nat. Commun. 2022, 13, 80. [Google Scholar] [CrossRef]
- Alshukairi, A.N.; Zheng, J.; Zhao, J.; Nehdi, A.; Baharoon, S.A.; Layqah, L.; Bokhari, A.; Al Johani, S.M.; Samman, N.; Boudjelal, M.; et al. High Prevalence of MERS-CoV Infection in Camel Workers in Saudi Arabia. mBio 2018, 9, e01985-18. [Google Scholar] [CrossRef] [Green Version]
- Mok, C.K.P.; Zhu, A.; Zhao, J.; Lau, E.H.; Wang, J.; Chen, Z.; Zhuang, Z.; Wang, Y.; Alshukairi, A.N.; Baharoon, S.A.; et al. T-cell responses to MERS coronavirus infection in people with occupational exposure to dromedary camels in Nigeria: An observational cohort study. Lancet Infect. Dis. 2021, 21, 385–395. [Google Scholar] [CrossRef]
- Rowland-Jones, S.L.; McMichael, A. Immune responses in HIV-exposed seronegatives: Have they repelled the virus? Curr. Opin. Immunol. 1995, 7, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Shacklett, B.L. Understanding the ‘Lucky Few’: The Conundrum of HIV-exposed, Seronegative Individuals. Curr. HIV AIDS Rep. 2006, 3, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.C.; Markee, J.; Akridge, R.E.; Meldorf, M.; Musey, L.; Karchmer, T.; Krone, M.; Collier, A.; Corey, L.; Emerman, M.; et al. Protection against Human Immunodeficiency Virus Type 1 Infection in Persons with Repeated Exposure: Evidence for T Cell Immunity in the Absence of Inherited CCR5 Coreceptor Defects. J. Infect. Dis. 1999, 179, 548–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, A.J.; Campion, S.L.; Kopycinski, J.; Moodie, Z.; Wang, Z.M.; Pandya, K.; Moore, S.; Liu, M.K.; Brackenridge, S.; Kuldanek, K.; et al. Differences in HIV-Specific T Cell Responses between HIV-Exposed and -Unexposed HIV-Seronegative Individuals. J. Virol. 2011, 85, 3507–3516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clerici, M.; Levin, J.M.; Kessler, H.A.; Harris, A.; Berzofsky, J.A.; Landay, A.L.; Shearer, G.M. HIV-Specific T-Helper Activity in Seronegative Health Care Workers Exposed to Contaminated Blood. JAMA 1994, 271, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Missale, G.; Papagno, L.; Penna, A.; Pilli, M.; Zerbini, A.; Vitali, P.; Pieroni, G.; Urbani, S.; Uggeri, J.; Pinheiro, S.; et al. Parenteral exposure to high HIV viremia leads to virus-specific T cell priming without evidence of infection. Eur. J. Immunol. 2004, 34, 3208–3215. [Google Scholar] [CrossRef]
- Clerici, M.; Berzofsky, J.A. T-cell proliferation to subinfectious SIV correlates with lack of infection after challenge of macaques. AIDS 1994, 8, 1391–1395. [Google Scholar] [CrossRef]
- Posavad, C.M.; Remington, M.; Mueller, D.E.; Zhao, L.; Magaret, A.S.; Wald, A.; Corey, L. Detailed Characterization of T Cell Responses to Herpes Simplex Virus-2 in Immune Seronegative Persons. J. Immunol. 2010, 184, 3250–3259. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, J.; Meya, S.; Schlaphoff, V.; Manns, M.P.; Mössner, J.; Wedemeyer, H.; Tillmann, H.L. HBV-specific T-cell responses in healthy seronegative sexual partners of patients with chronic HBV infection. J. Viral Hepat. 2010, 17, 631–639. [Google Scholar] [CrossRef] [PubMed]
- McElhaney, J.E.; Xie, D.; Hager, W.D.; Barry, M.B.; Wang, Y.; Kleppinger, A.; Ewen, C.; Kane, K.P.; Bleackley, R.C. T Cell Responses Are Better Correlates of Vaccine Protection in the Elderly. J. Immunol. 2006, 176, 6333–6339. [Google Scholar] [CrossRef] [Green Version]
- Tsang, T.K.; Lam, K.T.; Liu, Y.; Fang, V.J.; Mu, X.; Leung, N.H.; Peiris, J.S.; Leung, G.M.; Cowling, B.J.; Tu, W. Investigation of CD4 and CD8 T cell-mediated protection against influenza A virus in a cohort study. BMC Med. 2022, 20, 230. [Google Scholar] [CrossRef]
- Jeffery-Smith, A.; Burton, A.R.; Lens, S.; Rees-Spear, C.; Davies, J.; Patel, M.; Gopal, R.; Muir, L.; Aiano, F.; Doores, K.J.; et al. SARS-CoV-2-specific memory B cells can persist in the elderly who have lost detectable neutralizing antibodies. J. Clin. Investig. 2022, 132, e152042. [Google Scholar] [CrossRef]
- Chandran, A.; Rosenheim, J.; Nageswaran, G.; Swadling, L.; Pollara, G.; Gupta, R.K.; Burton, A.R.; Guerra-Assuncao, J.A.; Woolston, A.; Ronel, T.; et al. Rapid synchronous type 1 IFN and virus-specific T cell responses characterize first wave non-severe SARS-CoV-2 infections. Cell Rep. Med. 2022, 3, 100557. [Google Scholar] [CrossRef]
- Wilkinson, T.M.; Li, C.K.; Chui, C.S.; Huang, A.K.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Begom, S.; Bermingham, A.; Hoschler, K.; Adamson, W.; Carman, W.; Bean, T.; Barclay, W.; Deeks, J.J.; Lalvani, A. Cellular immune correlates of protection against symptomatic pandemic influenza. Nat. Med. 2013, 19, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Allie, S.R.; Bradley, J.E.; Mudunuru, U.; Schultz, M.D.; Graf, B.A.; Lund, F.E.; Randall, T.D. The establishment of resident memory B cells in the lung requires local antigen encounter. Nat. Immunol. 2018, 20, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Barker, K.A.; Etesami, N.S.; Shenoy, A.T.; Arafa, E.I.; de Ana, C.L.; Smith, N.M.; Martin, I.M.; Goltry, W.N.; Barron, A.M.; Browning, J.L.; et al. Lung-resident memory B cells protect against bacterial pneumonia. J. Clin. Investig. 2021, 131, e141810. [Google Scholar] [CrossRef]
- Gregoire, C.; Spinelli, L.; Villazala-Merino, S.; Gil, L.; Holgado, M.P.; Moussa, M.; Dong, C.; Zarubica, A.; Fallet, M.; Navarro, J.M.; et al. Viral infection engenders bona fide and bystander subsets of lung-resident memory B cells through a permissive mechanism. Immunity 2022, 55, 1216–1233.e9. [Google Scholar] [CrossRef]
- Hasselrot, K.; Säberg, P.; Hirbod, T.; Söderlund, J.; Ehnlund, M.; Bratt, G.; Sandström, E.; Broliden, K. Oral HIV-exposure elicits mucosal HIV-neutralizing antibodies in uninfected men who have sex with men. AIDS 2009, 23, 329–333. [Google Scholar] [CrossRef]
- Kaul, R.; Trabattoni, D.; Bwayo, J.J.; Arienti, D.; Zagliani, A.; Mwangi, F.M.; Kariuki, C.; Ngugi, E.N.; MacDonald, K.S.; Ball, B.T.; et al. HIV-1-specific mucosal IgA in a cohort of HIV-1-resistant Kenyan sex workers. AIDS 1999, 13, 23–29. [Google Scholar] [CrossRef]
- Cervia, C.; Nilsson, J.; Zurbuchen, Y.; Valaperti, A.; Schreiner, J.; Wolfensberger, A.; Raeber, M.E.; Adamo, S.; Weigang, S.; Emmenegger, M.; et al. Systemic and mucosal antibody responses specific to SARS-CoV-2 during mild versus severe COVID-19. J. Allergy Clin. Immunol. 2021, 147, 545–557.e9. [Google Scholar] [CrossRef]
- Tosif, S.; Neeland, M.R.; Sutton, P.; Licciardi, P.V.; Sarkar, S.; Selva, K.J.; Do, L.A.H.; Donato, C.; Quan Toh, Z.; Higgins, R.; et al. Immune responses to SARS-CoV-2 in three children of parents with symptomatic COVID-19. Nat. Commun. 2020, 11, 5703. [Google Scholar] [CrossRef] [PubMed]
- Shrock, E.; Fujimura, E.; Kula, T.; Timms, R.T.; Lee, I.H.; Leng, Y.; Robinson, M.L.; Sie, B.M.; Li, M.Z.; Chen, Y.; et al. Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. Science 2020, 370, eabd4250. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Z.; Lei, Q.; Lai, D.Y.; Hou, H.; Jiang, H.W.; Zheng, Y.X.; Wang, X.N.; Wu, J.; Ma, M.L.; et al. Antibody landscape against SARS-CoV-2 reveals significant differences between non-structural/accessory and structural proteins. Cell Rep. 2021, 36, 109391. [Google Scholar] [CrossRef]
- Peddireddy, S.P.; Rahman, S.A.; Cillo, A.R.; Vijay, G.M.; Somasundaram, A.; Workman, C.J.; Bain, W.; McVerry, B.J.; Methe, B.; Lee, J.S.; et al. Antibodies targeting conserved non-canonical antigens and endemic coronaviruses associate with favorable outcomes in severe COVID-19. Cell Rep. 2022, 39, 111020. [Google Scholar] [CrossRef] [PubMed]
- Neeland, M.R.; Bannister, S.; Clifford, V.; Dohle, K.; Mulholland, K.; Sutton, P.; Curtis, N.; Steer, A.C.; Burgner, D.P.; Crawford, N.W.; et al. Innate cell profiles during the acute and convalescent phase of SARS-CoV-2 infection in children. Nat. Commun. 2021, 12, 1084. [Google Scholar] [CrossRef] [PubMed]
- Habibi, M.S.; Thwaites, R.S.; Chang, M.; Jozwik, A.; Paras, A.; Kirsebom, F.; Varese, A.; Owen, A.; Cuthbertson, L.; James, P.; et al. Neutrophilic inflammation in the respiratory mucosa predisposes to RSV infection. Science 2020, 370, eaba9301. [Google Scholar] [CrossRef] [PubMed]
- Flament, H.; Rouland, M.; Beaudoin, L.; Toubal, A.; Bertrand, L.; Lebourgeois, S.; Rousseau, C.; Soulard, P.; Gouda, Z.; Cagninacci, L.; et al. Outcome of SARS-CoV-2 infection is linked to MAIT cell activation and cytotoxicity. Nat. Immunol. 2021, 22, 322–335. [Google Scholar] [CrossRef]
- Parrot, T.; Gorin, J.B.; Ponzetta, A.; Maleki, K.T.; Kammann, T.; Emgård, J.; Perez-Potti, A.; Sekine, T.; Rivera-Ballesteros, O. MAIT cell activation and dynamics associated with COVID-19 disease severity. Sci. Immunol. 2020, 5, eabe1670. [Google Scholar] [CrossRef]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural killer cell immunotypes related to COVID-19 disease severity. Sci. Immunol. 2020, 5, 6832. [Google Scholar] [CrossRef]
- Santer, D.M.; Li, D.; Ghosheh, Y.; Zahoor, M.A.; Prajapati, D.; Hansen, B.E.; Tyrrell, D.L.J.; Feld, J.J.; Gehring, A.J. Interferon-λ treatment accelerates SARS-CoV-2 clearance despite age-related delays in the induction of T cell immunity. Nat. Commun. 2022, 13, 6992. [Google Scholar] [CrossRef]
- Yamada, T.; Sato, S.; Sotoyama, Y.; Orba, Y.; Sawa, H.; Yamauchi, H.; Sasaki, M.; Takaoka, A. RIG-I triggers a signaling-abortive anti-SARS-CoV-2 defense in human lung cells. Nat. Immunol. 2021, 22, 820–828. [Google Scholar] [CrossRef]
- Asabe, S.; Wieland, S.F.; Chattopadhyay, P.K.; Roederer, M.; Engle, R.E.; Purcell, R.H.; Chisari, F.V. The Size of the Viral Inoculum Contributes to the Outcome of Hepatitis B Virus Infection. J. Virol. 2009, 83, 9652–9662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gujar, S.A.; Mulrooney-Cousins, P.M.; Michalak, T.I. Repeated Exposure to Trace Amounts of Woodchuck Hepadnavirus Induces Molecularly Evident Infection and Virus-Specific T Cell Response in the Absence of Serological Infection Markers and Hepatitis. J. Virol. 2013, 87, 1035–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland-Jones, S.L.; Nixon, D.F.; Gotch, F.; McMichael, A.; Kroll, J.S.; Hallam, N.; Aldhous, M.C.; Froebel, K.; Ariyoshi, K. HIV-specific cytotoxic T-cell activity in an HIV-exposed but uninfected infant. Lancet 1993, 341, 860–861. [Google Scholar] [CrossRef] [PubMed]
- Scurr, M.J.; Lippiatt, G.; Capitani, L.; Bentley, K.; Lauder, S.N.; Smart, K.; Somerville, M.S.; Rees, T.; Stanton, R.J.; Gallimore, A.; et al. Magnitude of venous or capillary blood-derived SARS-CoV-2-specific T cell response determines COVID-19 immunity. Nat. Commun. 2022, 13, 5422. [Google Scholar] [CrossRef]
- Tan, A.T.; Lim, J.M.; Le Bert, N.; Kunasegaran, K.; Chia, A.; Qui, M.D.; Tan, N.; Chia, W.N.; de Alwis, R.; Ying, D.; et al. Rapid measurement of SARS-CoV-2 spike T cells in whole blood from vaccinated and naturally infected individuals. J. Clin. Investig. 2021, 131, e152379. [Google Scholar] [CrossRef]
- Schwarz, M.; Torre, D.; Lozano-Ojalvo, D.; Tan, A.T.; Tabaglio, T.; Mzoughi, S.; Sanchez-Tarjuelo, R.; Le Bert, N.; Lim, J.M.E.; Hatem, S.; et al. Rapid, scalable assessment of SARS-CoV-2 cellular immunity by whole-blood PCR. Nat. Biotechnol. 2022, 40, 1680–1689. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swadling, L.; Maini, M.K. Can T Cells Abort SARS-CoV-2 and Other Viral Infections? Int. J. Mol. Sci. 2023, 24, 4371. https://doi.org/10.3390/ijms24054371
Swadling L, Maini MK. Can T Cells Abort SARS-CoV-2 and Other Viral Infections? International Journal of Molecular Sciences. 2023; 24(5):4371. https://doi.org/10.3390/ijms24054371
Chicago/Turabian StyleSwadling, Leo, and Mala K. Maini. 2023. "Can T Cells Abort SARS-CoV-2 and Other Viral Infections?" International Journal of Molecular Sciences 24, no. 5: 4371. https://doi.org/10.3390/ijms24054371
APA StyleSwadling, L., & Maini, M. K. (2023). Can T Cells Abort SARS-CoV-2 and Other Viral Infections? International Journal of Molecular Sciences, 24(5), 4371. https://doi.org/10.3390/ijms24054371