
Upregulation of DJ-1 in Dopaminergic Neurons by a Physically-Modified Saline: Implications for Parkinson’s Disease

Abstract
:1. Introduction
2. Results
2.1. RNS60 Upregulates the Level of DJ-1 in Mouse MN9D Dopaminergic Neuronal Cells
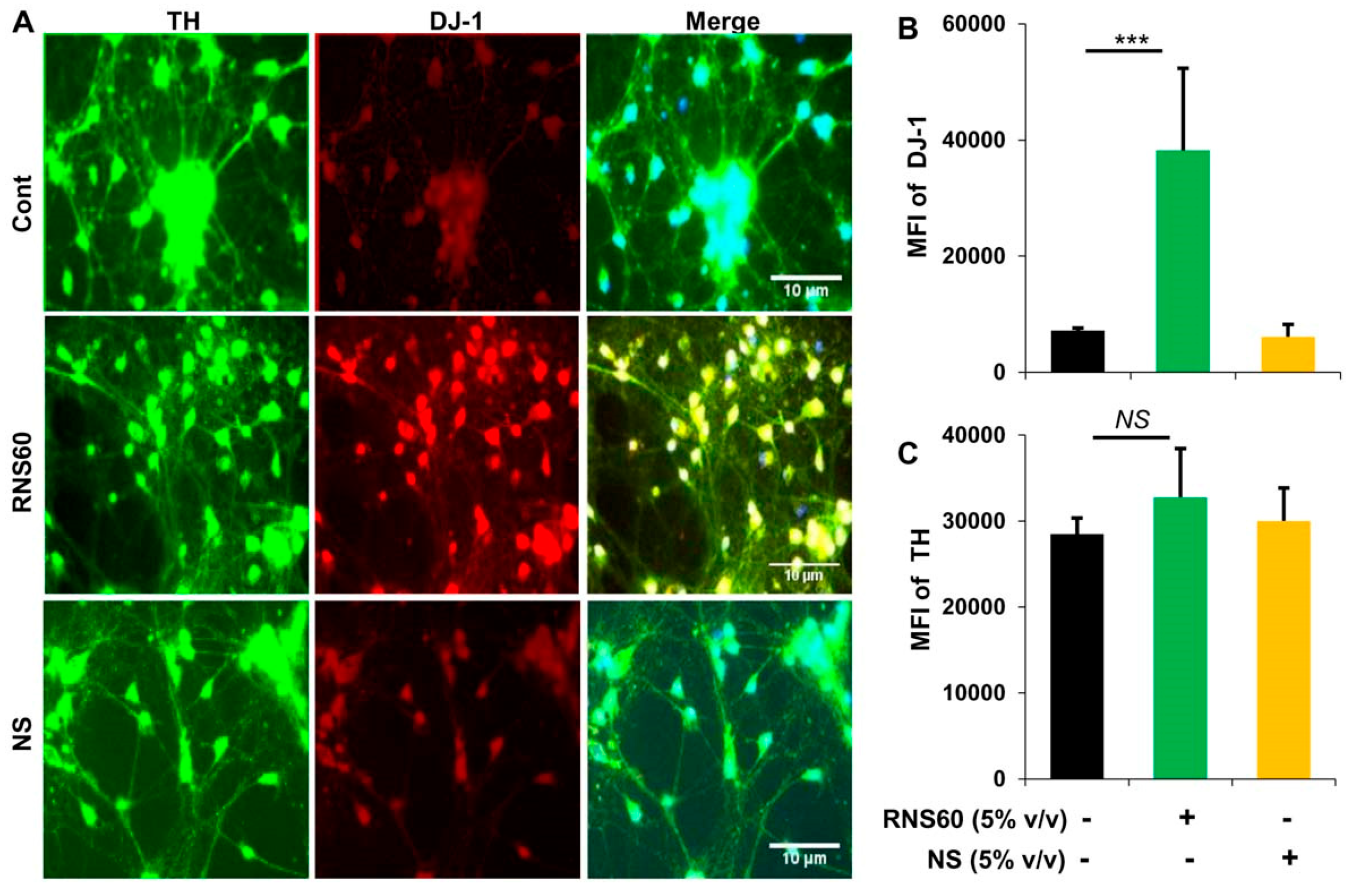
2.2. Does RNS60 Upregulate DJ-1 in Mouse Primary Dopaminergic Neurons?
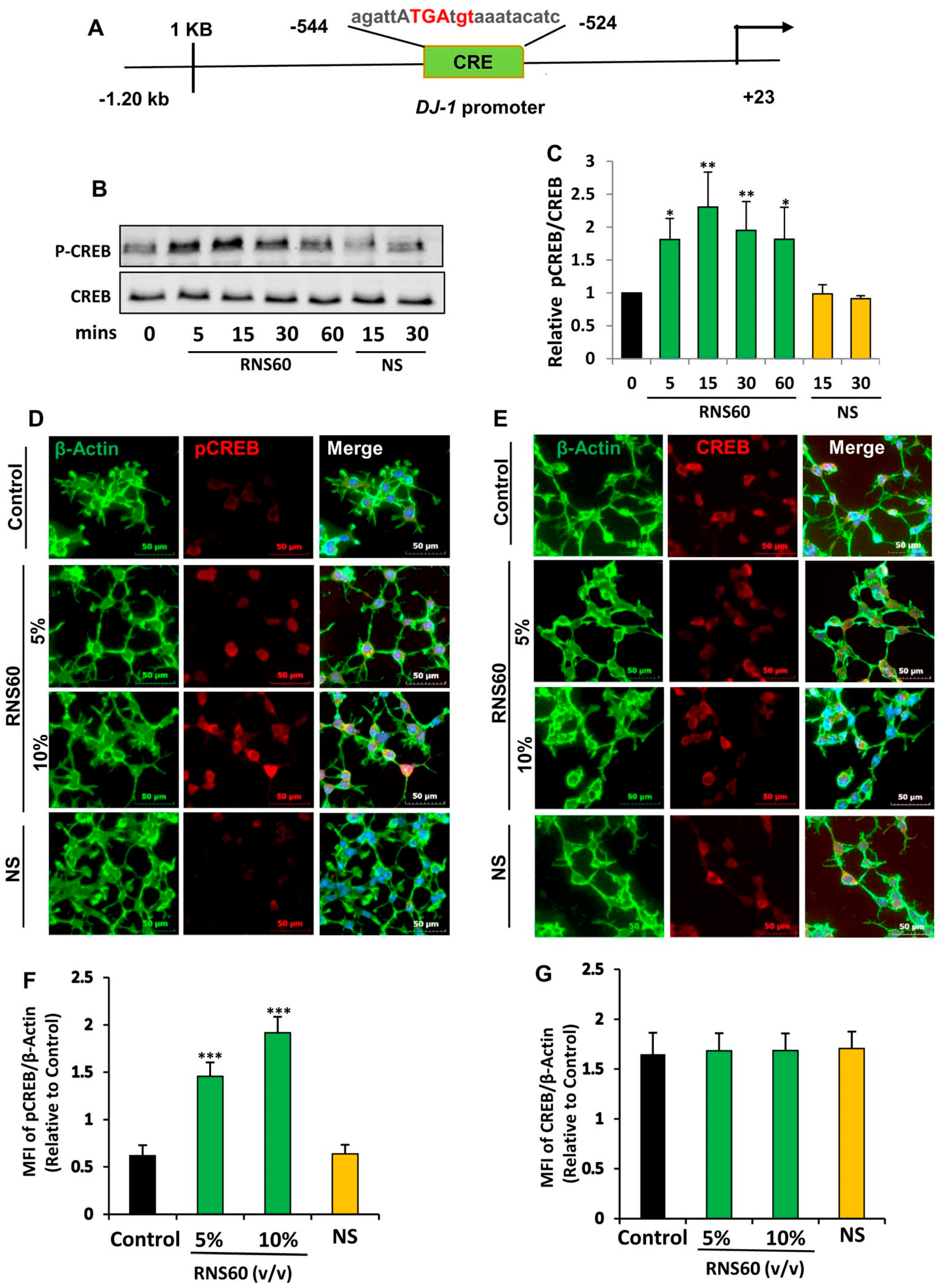
2.3. Activation of cAMP Response Element-Binding Protein (CREB) by RNS60 in MN9D Neuronal Cells
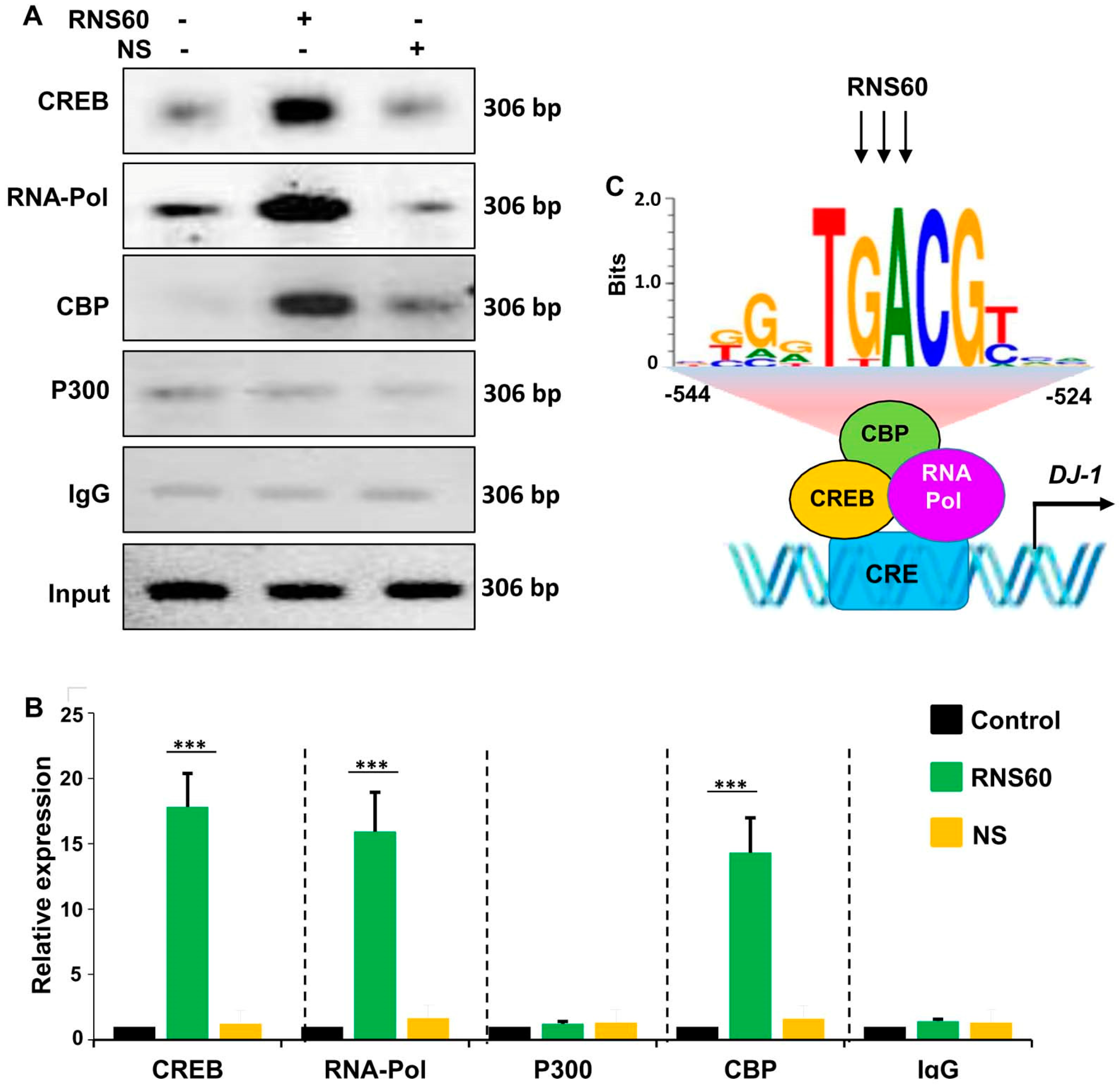
2.4. RNS60 Stimulates the Employment of CREB to the Promoter of DJ-1 Gene in MN9D Neuronal Cells
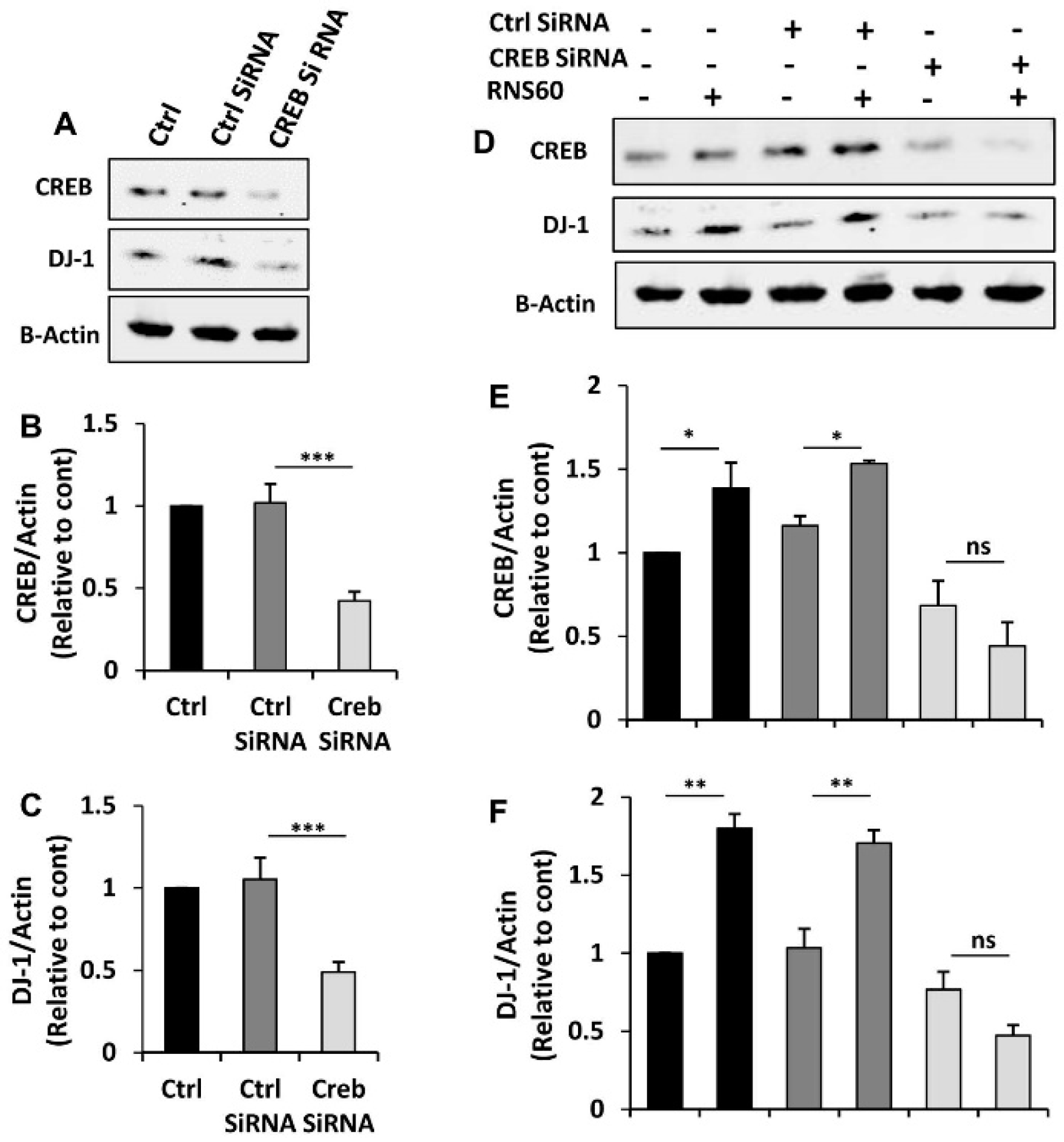
2.5. RNS60 Needs CREB for the Up-Regulation of DJ-1 in MN9D Neuronal Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of RNS60
4.3. MN9D Cells
4.4. Isolation of Mouse Primary Dopaminergic Neurons
4.5. Immunoblot Analysis
4.6. Semi-Quantitative RT-PCR Analysis
- DJ-1:
- Sense: 5′-CCCCGTGCAGTGTAGCCGTG-3′
- Antisense: 5′-CAGGCCGTCCTTCTCCACGC-3′
- GAPDH:
- Sense: 5′-GGTGAAGGTCGGTGTGAACG-3′
- Antisense: 5′-TTGGCTCCACCCTTCAAGTG-3′.
4.7. Real-Time PCR
4.8. Immunofluorescence Analysis
4.9. Electrophoretic Mobility Shift Assay (EMSA)
4.10. Chromatin Immunoprecipitation (ChIP) Assay
- Sense: 5′-GAGATCTCATTTACCCTGATTTAA-3′
- Antisense: 5′-GATCCTGATGCTGCTGCACCCACAG-3′
4.11. Measurement of Mean Fluorescence Intensity (MFI)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CBP | CREB-binding protein |
| ChIP | Chromatin immunoprecipitation |
| CREB | cAMP response element-binding |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| IgG | Immunoglobulin |
| MFI | Mean fluorescence intensity |
| PD | Parkinson’s disease |
| RT-PCR | Reverse transcriptase-coupled polymerase chain reaction |
| SNpc | Substantia nigra pars compacta |
| TBST | TBS plus Tween 20 |
| TCP | Taylor-Couette-Poiseuille |
| TH | Tyrosine hydroxylase |
References
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanow, C.W.; Tatton, W.G. Etiology and pathogenesis of Parkinson’s disease. Annu. Rev. Neurosci. 1999, 22, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, J.; Cai, H.; Cookson, M.R.; Gwinn-Hardy, K.; Singleton, A. Genetics of Parkinson’s disease and parkinsonism. Ann. Neurol. 2006, 60, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [Green Version]
- Strauss, K.M.; Martins, L.M.; Plun-Favreau, H.; Marx, F.P.; Kautzmann, S.; Berg, D.; Gasser, T.; Wszolek, Z.; Muller, T.; Bornemann, A.; et al. Loss of function mutations in the gene encoding Omi/HtrA2 in Parkinson’s disease. Hum. Mol. Genet. 2005, 14, 2099–2111. [Google Scholar] [CrossRef] [Green Version]
- Paisan-Ruiz, C.; Jain, S.; Evans, E.W.; Gilks, W.P.; Simon, J.; van der Brug, M.; Lopez de Munain, A.; Aparicio, S.; Gil, A.M.; Khan, N.; et al. Cloning of the gene containing mutations that cause PARK8-linked Parkinson’s disease. Neuron 2004, 44, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Parkinsons Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Pahan, K. Prospects of statins in Parkinson disease. Neuroscientist 2011, 17, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, X.; Li, X.; Liu, Q.; Cheng, Y. Oxidative Stress in Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Mol. Neurosci. 2018, 11, 236. [Google Scholar] [CrossRef] [PubMed]
- Dolgacheva, L.P.; Berezhnov, A.V.; Fedotova, E.I.; Zinchenko, V.P.; Abramov, A.Y. Role of DJ-1 in the mechanism of pathogenesis of Parkinson’s disease. J. Bioenerg. Biomembr. 2019, 51, 175–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, N.; Xu, J. Synergistic activation of the human MnSOD promoter by DJ-1 and PGC-1alpha: Regulation by SUMOylation and oxidation. Hum. Mol. Genet. 2008, 17, 3357–3367. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Freed, C.R. DJ-1 up-regulates glutathione synthesis during oxidative stress and inhibits A53T alpha-synuclein toxicity. J. Biol. Chem. 2005, 280, 43150–43158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Modi, K.K.; Khasnavis, S.; Ghosh, S.; Watson, R.; Pahan, K. Enhancement of morphological plasticity in hippocampal neurons by a physically modified saline via phosphatidylinositol-3 kinase. PLoS ONE 2014, 9, e101883. [Google Scholar] [CrossRef] [Green Version]
- Khasnavis, S.; Jana, A.; Roy, A.; Mazumder, M.; Bhushan, B.; Wood, T.; Ghosh, S.; Watson, R.; Pahan, K. Suppression of nuclear factor-kappaB activation and inflammation in microglia by physically modified saline. J. Biol. Chem. 2012, 287, 29529–29542. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Martinson, J.A.; Ghosh, S.; Watson, R.; Pahan, K. Protection of Tregs, suppression of Th1 and Th17 cells, and amelioration of experimental allergic encephalomyelitis by a physically-modified saline. PLoS ONE 2012, 7, e51869. [Google Scholar] [CrossRef] [Green Version]
- Khasnavis, S.; Roy, A.; Ghosh, S.; Watson, R.; Pahan, K. Protection of dopaminergic neurons in a mouse model of Parkinson’s disease by a physically-modified saline containing charge-stabilized nanobubbles. J. Neuroimmune Pharmacol. 2014, 9, 218–232. [Google Scholar] [CrossRef]
- Modi, K.K.; Roy, A.; Brahmachari, S.; Rangasamy, S.B.; Pahan, K. Cinnamon and Its Metabolite Sodium Benzoate Attenuate the Activation of p21rac and Protect Memory and Learning in an Animal Model of Alzheimer’s Disease. PLoS ONE 2015, 10, e0130398. [Google Scholar] [CrossRef] [PubMed]
- Chandra, G.; Kundu, M.; Rangasamy, S.B.; Dasarathy, S.; Ghosh, S.; Watson, R.; Pahan, K. Increase in Mitochondrial Biogenesis in Neuronal Cells by RNS60, a Physically-Modified Saline, via Phosphatidylinositol 3-Kinase-Mediated Upregulation of PGC1alpha. J. Neuroimmune Pharmacol. 2018, 13, 143–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, V.T.; Khan, D.; Jones, R.G.; Nakamura, D.S.; Kennedy, T.E.; Cui, Q.L.; Rone, M.B.; Healy, L.M.; Watson, R.; Ghosh, S.; et al. Potential Benefit of the Charge-Stabilized Nanostructure Saline RNS60 for Myelin Maintenance and Repair. Sci. Rep. 2016, 6, 30020. [Google Scholar] [CrossRef] [Green Version]
- Baulac, S.; Lu, H.; Strahle, J.; Yang, T.; Goldberg, M.S.; Shen, J.; Schlossmacher, M.G.; Lemere, C.A.; Lu, Q.; Xia, W. Increased DJ-1 expression under oxidative stress and in Alzheimer’s disease brains. Mol. Neurodegener. 2009, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khasnavis, S.; Pahan, K. Sodium benzoate, a metabolite of cinnamon and a food additive, upregulates neuroprotective Parkinson disease protein DJ-1 in astrocytes and neurons. J. Neuroimmune Pharmacol. 2012, 7, 424–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khasnavis, S.; Pahan, K. Cinnamon treatment upregulates neuroprotective proteins Parkin and DJ-1 and protects dopaminergic neurons in a mouse model of Parkinson’s disease. J. Neuroimmune Pharmacol. 2014, 9, 569–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajjad, M.U.; Green, E.W.; Miller-Fleming, L.; Hands, S.; Herrera, F.; Campesan, S.; Khoshnan, A.; Outeiro, T.F.; Giorgini, F.; Wyttenbach, A. DJ-1 modulates aggregation and pathogenesis in models of Huntington’s disease. Hum. Mol. Genet. 2014, 23, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Saha, R.N.; Jana, M.; Pahan, K. MAPK p38 regulates transcriptional activity of NF-kappaB in primary human astrocytes via acetylation of p65. J. Immunol. 2007, 179, 7101–7109. [Google Scholar] [CrossRef] [Green Version]
- Saha, R.N.; Pahan, K. HATs and HDACs in neurodegeneration: A tale of disconcerted acetylation homeostasis. Cell Death Differ. 2006, 13, 539–550. [Google Scholar] [CrossRef]
- Clements, C.M.; McNally, R.S.; Conti, B.J.; Mak, T.W.; Ting, J.P. DJ-1, a cancer- and Parkinson’s disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc. Natl. Acad. Sci. USA 2006, 103, 15091–15096. [Google Scholar] [CrossRef] [Green Version]
- MacKeigan, J.P.; Clements, C.M.; Lich, J.D.; Pope, R.M.; Hod, Y.; Ting, J.P. Proteomic profiling drug-induced apoptosis in non-small cell lung carcinoma: Identification of RS/DJ-1 and RhoGDIalpha. Cancer Res. 2003, 63, 6928–6934. [Google Scholar] [PubMed]
- Aleyasin, H.; Rousseaux, M.W.; Marcogliese, P.C.; Hewitt, S.J.; Irrcher, I.; Joselin, A.P.; Parsanejad, M.; Kim, R.H.; Rizzu, P.; Callaghan, S.M.; et al. DJ-1 protects the nigrostriatal axis from the neurotoxin MPTP by modulation of the AKT pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 3186–3191. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.H.; Peters, M.; Jang, Y.; Shi, W.; Pintilie, M.; Fletcher, G.C.; DeLuca, C.; Liepa, J.; Zhou, L.; Snow, B.; et al. DJ-1, a novel regulator of the tumor suppressor PTEN. Cancer Cell 2005, 7, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage, S.; Brice, A. Parkinson’s disease: From monogenic forms to genetic susceptibility factors. Hum. Mol. Genet. 2009, 18, R48–R59. [Google Scholar] [CrossRef] [PubMed]
- Zhong, N.; Kim, C.Y.; Rizzu, P.; Geula, C.; Porter, D.R.; Pothos, E.N.; Squitieri, F.; Heutink, P.; Xu, J. DJ-1 transcriptionally up-regulates the human tyrosine hydroxylase by inhibiting the sumoylation of pyrimidine tract-binding protein-associated splicing factor. J. Biol. Chem. 2006, 281, 20940–20948. [Google Scholar] [CrossRef] [Green Version]
- Bossers, K.; Meerhoff, G.; Balesar, R.; van Dongen, J.W.; Kruse, C.G.; Swaab, D.F.; Verhaagen, J. Analysis of gene expression in Parkinson’s disease: Possible involvement of neurotrophic support and axon guidance in dopaminergic cell death. Brain Pathol. 2009, 19, 91–107. [Google Scholar] [CrossRef]
- Beghi, E.; Pupillo, E.; Bianchi, E.; Bonetto, V.; Luotti, S.; Pasetto, L.; Bendotti, C.; Tortarolo, M.; Sironi, F.; Camporeale, L.; et al. Effect of RNS60 in amyotrophic lateral sclerosis: A phase II multicentre, randomized, double-blind, placebo-controlled trial. Eur. J. Neurol. 2023, 30, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Zabielska-Kaczorowska, M.A.; Wierzbicka, B.; Kalmes, A.; Slominska, E.M.; Yacoub, M.H.; Smolenski, R.T. Beneficial Effects of RNS60 in Cardiac Ischemic Injury. Curr. Issues Mol. Biol. 2022, 44, 4877–4887. [Google Scholar] [CrossRef]
- Choi, S.; Yu, E.; Rabello, G.; Merlo, S.; Zemmar, A.; Walton, K.D.; Moreno, H.; Moreira, J.E.; Sugimori, M.; Llinas, R.R. Enhanced synaptic transmission at the squid giant synapse by artificial seawater based on physically modified saline. Front. Synaptic Neurosci. 2014, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, Y.; Ikebe, S.; Hattori, N.; Nakagawa-Hattori, Y.; Mochizuki, H.; Tanaka, M.; Ozawa, T. Role of mitochondria in the etiology and pathogenesis of Parkinson’s disease. Biochim. Biophys. Acta 1995, 1271, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Surmeier, D.J.; Sulzer, D. The pathology roadmap in Parkinson disease. Prion 2013, 7, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jana, M.; Ghosh, S.; Pahan, K. Upregulation of Myelin Gene Expression by a Physically-Modified Saline via Phosphatidylinositol 3-Kinase-Mediated Activation of CREB: Implications for Multiple Sclerosis. Neurochem. Res. 2018, 43, 407–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Rangasamy, S.B.; Ghosh, S.; Watson, R.L.; Pahan, K. Nebulization of RNS60, a Physically-Modified Saline, Attenuates the Adoptive Transfer of Experimental Allergic Encephalomyelitis in Mice: Implications for Multiple Sclerosis Therapy. Neurochem. Res. 2017, 42, 1555–1570. [Google Scholar] [CrossRef] [PubMed]
- Modi, K.K.; Jana, A.; Ghosh, S.; Watson, R.; Pahan, K. A physically-modified saline suppresses neuronal apoptosis, attenuates tau phosphorylation and protects memory in an animal model of Alzheimer’s disease. PLoS ONE 2014, 9, e103606. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, S.B.; Ghosh, S.; Pahan, K. RNS60, a physically-modified saline, inhibits glial activation, suppresses neuronal apoptosis and protects memory in a mouse model of traumatic brain injury. Exp. Neurol. 2020, 328, 113279. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, S.B.; Dasarathi, S.; Nutakki, A.; Mukherjee, S.; Nellivalasa, R.; Pahan, K. Stimulation of Dopamine Production by Sodium Benzoate, a Metabolite of Cinnamon and a Food Additive. J. Alzheimers Dis. Rep. 2021, 5, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, S.B.; Dasarathi, S.; Pahan, P.; Jana, M.; Pahan, K. Low-Dose Aspirin Upregulates Tyrosine Hydroxylase and Increases Dopamine Production in Dopaminergic Neurons: Implications for Parkinson’s Disease. J. Neuroimmune Pharmacol. 2019, 14, 173–187. [Google Scholar] [CrossRef]
- Choi, H.K.; Won, L.; Roback, J.D.; Wainer, B.H.; Heller, A. Specific modulation of dopamine expression in neuronal hybrid cells by primary cells from different brain regions. Proc. Natl. Acad. Sci. USA 1992, 89, 8943–8947. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Pahan, K. Ankyrin repeat and BTB/POZ domain containing protein-2 inhibits the aggregation of alpha-synuclein: Implications for Parkinson’s disease. FEBS Lett. 2013, 587, 3567–3574. [Google Scholar] [CrossRef] [Green Version]
- Saha, R.N.; Liu, X.; Pahan, K. Up-regulation of BDNF in astrocytes by TNF-alpha: A case for the neuroprotective role of cytokine. J. Neuroimmune Pharmacol. 2006, 1, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Roy, A.; Liu, X.; Kordower, J.H.; Mufson, E.J.; Hartley, D.M.; Ghosh, S.; Mosley, R.L.; Gendelman, H.E.; Pahan, K. Selective inhibition of NF-kappaB activation prevents dopaminergic neuronal loss in a mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 18754–18759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Roy, A.; Matras, J.; Brahmachari, S.; Gendelman, H.E.; Pahan, K. Simvastatin inhibits the activation of p21ras and prevents the loss of dopaminergic neurons in a mouse model of Parkinson’s disease. J. Neurosci. 2009, 29, 13543–13556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jana, A.; Pahan, K. Oxidative stress kills human primary oligodendrocytes via neutral sphingomyelinase: Implications for multiple sclerosis. J. Neuroimmune Pharmacol. 2007, 2, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Paidi, R.K.; Jana, M.; Mishra, R.K.; Dutta, D.; Pahan, K. Selective Inhibition of the Interaction between SARS-CoV-2 Spike S1 and ACE2 by SPIDAR Peptide Induces Anti-Inflammatory Therapeutic Responses. J. Immunol. 2021, 207, 2521–2533. [Google Scholar] [CrossRef]
- Paidi, R.K.; Jana, M.; Mishra, R.K.; Dutta, D.; Raha, S.; Pahan, K. ACE-2-interacting Domain of SARS-CoV-2 (AIDS) Peptide Suppresses Inflammation to Reduce Fever and Protect Lungs and Heart in Mice: Implications for COVID-19 Therapy. J. Neuroimmune Pharmacol. 2021, 16, 59–70. [Google Scholar] [CrossRef]
- Jana, A.; Modi, K.K.; Roy, A.; Anderson, J.A.; van Breemen, R.B.; Pahan, K. Up-regulation of neurotrophic factors by cinnamon and its metabolite sodium benzoate: Therapeutic implications for neurodegenerative disorders. J. Neuroimmune Pharmacol. 2013, 8, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Kundu, M.; Mondal, S.; Roy, A.; Martinson, J.L.; Pahan, K. Sodium Benzoate, a Food Additive and a Metabolite of Cinnamon, Enriches Regulatory T Cells via STAT6-Mediated Upregulation of TGF-beta. J. Immunol. 2016, 197, 3099–3110. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Prorok, T.; Roy, A.; Patel, D.; Dasarathi, S.; Pahan, K. Upregulation of IL-1 Receptor Antagonist by Aspirin in Glial Cells via Peroxisome Proliferator-Activated Receptor-Alpha. J. Alzheimers Dis. Rep. 2021, 5, 647–661. [Google Scholar] [CrossRef]
- Chandra, G.; Roy, A.; Rangasamy, S.B.; Pahan, K. Induction of Adaptive Immunity Leads to Nigrostriatal Disease Progression in MPTP Mouse Model of Parkinson’s Disease. J. Immunol. 2017, 198, 4312–4326. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Manufacturer | Catalog | Host | Application Dilution |
|---|---|---|---|---|
| TH | Abcam | ab137869 | Rabbit | WB 1:1000 IF 1:500 |
| DJ-1 | Cell Signaling | 35743 | Mouse | WB 1:1000 IF 1:500 |
| p-CREB | Cell Signaling | 9198L | Rabbit | WB 1:500 |
| CREB | Cell Signaling | 9197S | Rabbit | WB 1:500 |
| β-actin | Abcam | Ab6276 | Mouse | WB 1:6000 |
| CBP | Santa Cruz | SC-369 | Rabbit | ChIP 2 µg |
| p300 | Santa Cruz | SC-585 | Rabbit | ChIP 2 µg |
| CREB | Millipore | CS203204 | Rabbit | ChIP 2 µg |
| IgG | Santa Cruz | SC-3888 | Rabbit | ChIP 2 µg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jana, M.; Dasarathy, S.; Ghosh, S.; Pahan, K. Upregulation of DJ-1 in Dopaminergic Neurons by a Physically-Modified Saline: Implications for Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 4652. https://doi.org/10.3390/ijms24054652
Jana M, Dasarathy S, Ghosh S, Pahan K. Upregulation of DJ-1 in Dopaminergic Neurons by a Physically-Modified Saline: Implications for Parkinson’s Disease. International Journal of Molecular Sciences. 2023; 24(5):4652. https://doi.org/10.3390/ijms24054652
Chicago/Turabian StyleJana, Malabendu, Sridevi Dasarathy, Supurna Ghosh, and Kalipada Pahan. 2023. "Upregulation of DJ-1 in Dopaminergic Neurons by a Physically-Modified Saline: Implications for Parkinson’s Disease" International Journal of Molecular Sciences 24, no. 5: 4652. https://doi.org/10.3390/ijms24054652
APA StyleJana, M., Dasarathy, S., Ghosh, S., & Pahan, K. (2023). Upregulation of DJ-1 in Dopaminergic Neurons by a Physically-Modified Saline: Implications for Parkinson’s Disease. International Journal of Molecular Sciences, 24(5), 4652. https://doi.org/10.3390/ijms24054652