
Morphological and Physio-Chemical Responses to PEG-Induced Water Stress in Vanilla planifolia and V. pompona Hybrids

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
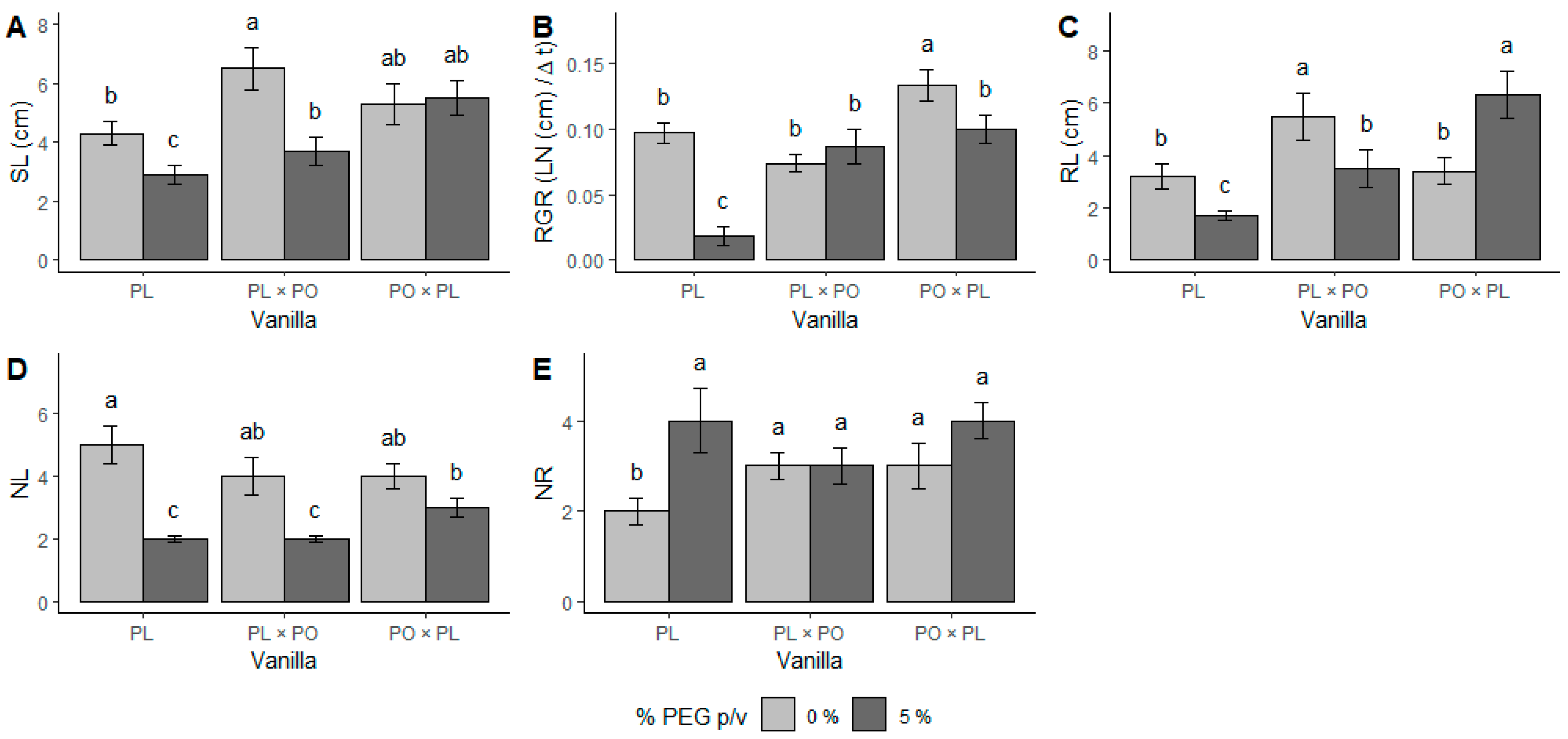
2.1. Morphological and Physiological Responses to Water Stress
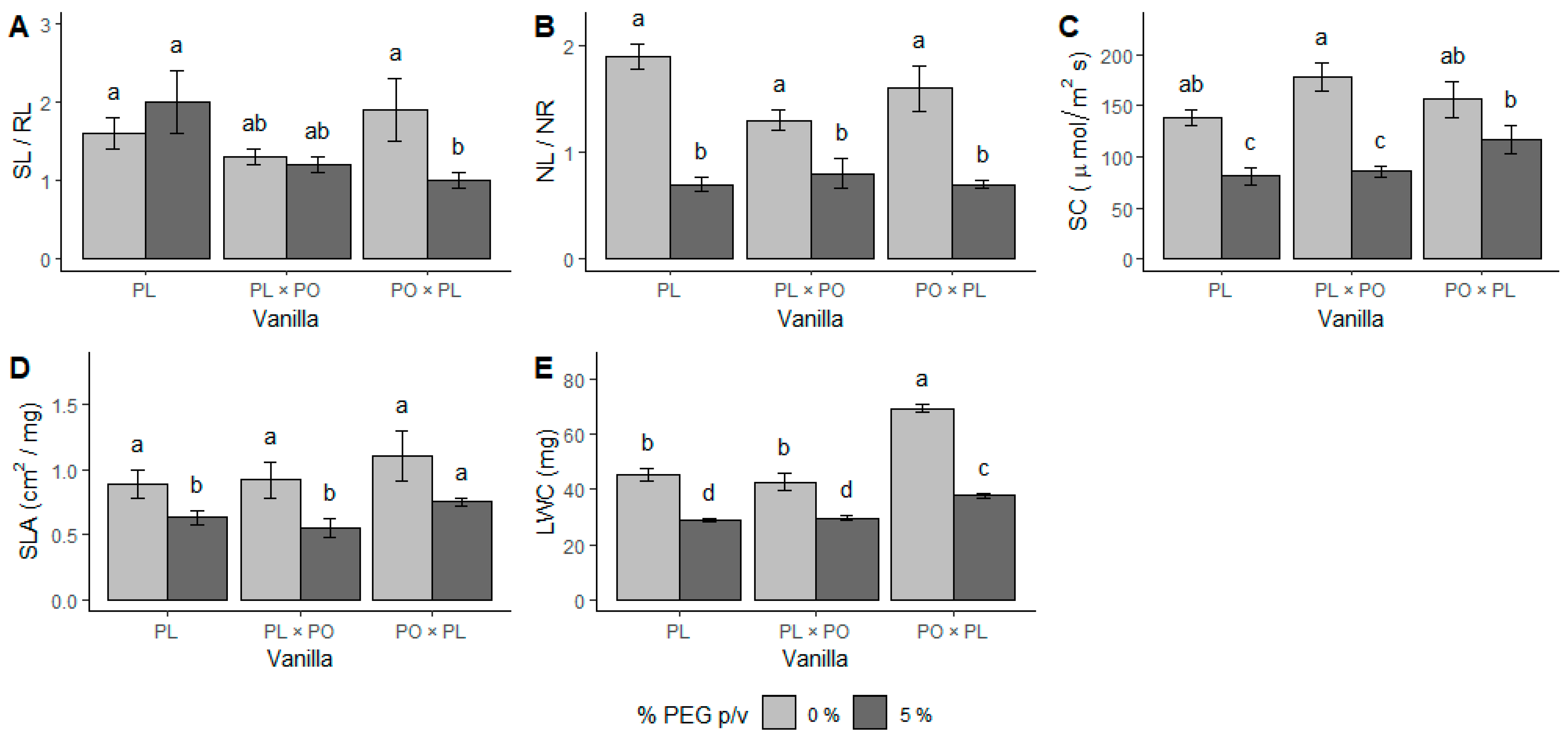
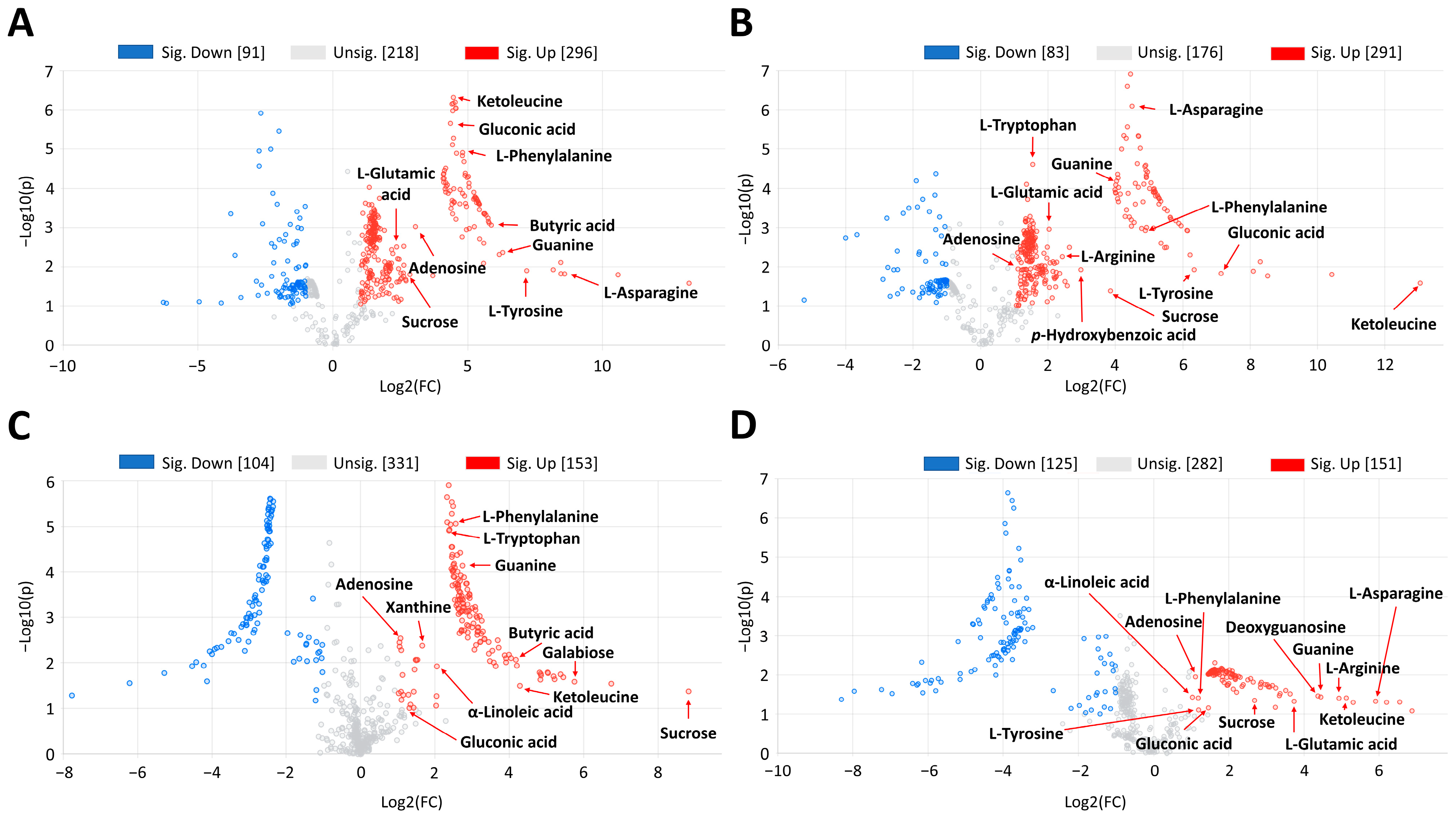
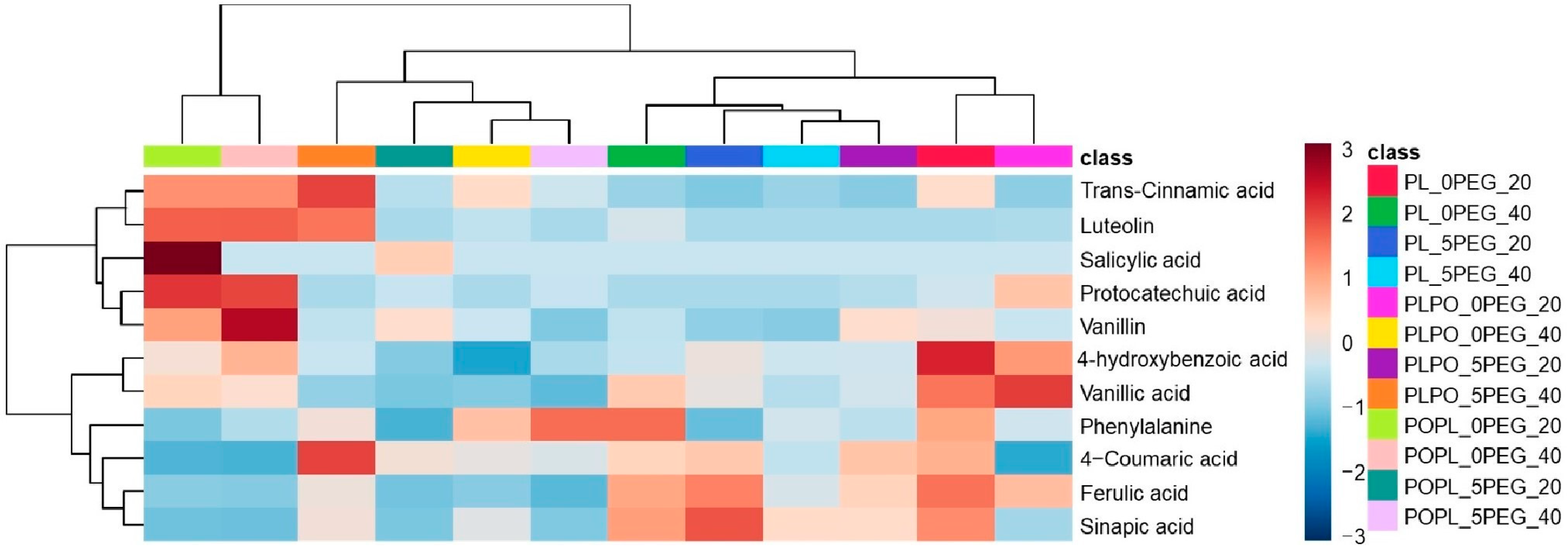
2.2. Physio-Chemical Response to Water Stress
3. Discussion
3.1. Morphological and Physiological Responses
3.2. Physio-Chemical Responses

4. Materials and Methods
4.1. Biological Material
4.2. Morphological and Physiological Responses to Water Stress
4.3. Physio-Chemical Response to Water Stress
4.3.1. Untargeted Metabolomics
4.3.2. Identification of Phenolics Compounds by UPLC-MS-MS
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ranadive, A.S. Quality control of vanilla beans and extracts. In Handbook of Vanilla Science and Technology, 2nd ed.; Havkin-Frenkel, D., Belanger, F., Eds.; Wiley: New York, NY, USA, 2019; pp. 239–260. [Google Scholar] [CrossRef]
- Hernández-Hernández, J. Mexican Vanilla Production. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F., Eds.; Wiley: New York, NY, USA, 2019; Volume 2, pp. 3–26. [Google Scholar] [CrossRef]
- Armenta-Montero, S.; Menchaca-García, R.; Pérez-Silva, A.; Velázquez-Rosas, N. Changes in the Potential Distribution of Vanilla planifolia Andrews under Different Climate Change Projections in Mexico. Sustainability 2022, 14, 2881. [Google Scholar] [CrossRef]
- Menchaca-García, R.A. In vitro germination of Vanilla. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F., Eds.; Wiley: New York, NY, USA, 2019; Volume 2, pp. 181–190. [Google Scholar] [CrossRef]
- Hernández-Hernández, J. Vanilla diseases. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F., Eds.; Wiley: New York, NY, USA, 2019; Volume 2, pp. 27–40. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. Available online: http://www.nature.com/doifinder/10.1038/nclimate2067 (accessed on 20 November 2020). [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extr. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, C.; Ranadive, A.S.; Vázquez, J.T.; Havkin-Frenkel, D. Curing on Vanilla. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F., Eds.; Wiley: New York, NY, USA, 2019; Volume 2, pp. 191–222. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Huang, W.; Hu, H. Physiological diversity of orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Rahman, M.; Rathinasabapathi, B.; Babar, M. UPLC-HRMS-based untargeted metabolic profiling reveals changes in chickpea (Cicer arietinum) metabolome following long-term drought stress. Plant Cell Environ. 2018, 42, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Salehi-Lisar, S.; Bakhshayeshan-Agdam, H. Drought Stress in Plants: Causes, Consequences, and Tolerance. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burrit, D., Tran, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–16. [Google Scholar] [CrossRef]
- Flexas, J.; Gallé, A.; Galmés, J.; Ribas-Carbo, M.; Medrano, H. The response of photosynthesis to soil water stress. In Plant Responses to Drought Stress from Morphological to Molecular Features; Aroca, R., Ed.; Springer: Granada, Spain, 2012. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sianuer Associates: Sunderland, MA, USA, 2010; pp. 1–623. [Google Scholar]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, H. Drought stress in plants: An overview. In Plant Responses to Drought Stress from Morphological to Molecular Features; Aroca, R., Ed.; Springer: Granada, Spain, 2012; pp. 1–34. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, J.; Huang, W.; Song, X.; Niu, J. Transcriptomics and Metabolomics Reveal Purine and Phenylpropanoid Metabolism Response to Drought Stress in Dendrobium sinense, an Endemic Orchid Species in Hainan Island. Front. Gen. 2021, 12, 692702. [Google Scholar] [CrossRef]
- Zhang, M.; Yu, Z.; Zeng, D.; Si, C.; Zhao, C.; Wang, H.; Li, C.; He, C.; Duan, J. Transcriptome and Metabolome Reveal Salt-Stress Responses of Leaf Tissues from Dendrobium officinale. Biomolecules 2021, 11, 736. [Google Scholar] [CrossRef] [PubMed]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic Adjustment and Plant Adaptation to Drought Stress. In Drought Stress Tolerance in Plants, Vol. I. Physiology and Chemistry; Hossain, M., Wani, S., Bhattacharjee, S., Burrit, D., Tran, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 104–144. [Google Scholar]
- Samarah, N. Understanding How Plants Respond to Drought Stress at the Molecular and Whole Plant Levels. In Drought Stress Tolerance in Plants, Vol. II.; Hossain, M., Wani, S., Bhattacharjee, S., Burrit, D., Tran, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–38. [Google Scholar] [CrossRef]
- Sami, F.; Yusuf, M.; Faizan, M.; Faraz, A.; Hayat, S. Role of sugars under abiotic stress. Plant. Physiol. 2016, 109, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hassanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2020, 172, 1321–1335. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Gundaraniya, S.; Ambalam, P.; Tomar, R. Metabolomic Profiling of Drought-Tolerant and Susceptible Peanut (Arachis hypogaea L.) Genotypes in Response to Drought Stress. ACS Omega 2020, 5, 31209–31219. [Google Scholar] [CrossRef] [PubMed]
- Pectu, E.; Arsintescu, A.; Stanciu, D. The effect of drought stress on fatty acid composition in some Romanian sunflower hybrids. Rom. Agric. Res. 2001, 15, 39–43. [Google Scholar]
- Ashraf, M.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M. Environmental Stress and Secondary Metabolites in Plants: An Overview. In Plant Metabolites and Regulation Under Environmental Stress; Ahmad, P., Ahanger, M., Singh, V., Tripathi, D., Alam, P., Alyemeni, M., Eds.; Elsevier: London, UK, 2018; pp. 153–167. ISBN 978-0-12-812689-9. [Google Scholar]
- Quan, N.T.; Anh, L.H.; Khang, D.T.; Tuyen, P.T.; Toan, N.P.; Minh, T.N.; Bach, D.T.; Ha, P.T.T.; Elzaawely, A.A.; Khanh, T.D. Involvement of secondary metabolites in response to drought stress of rice (Oryza sativa L.). Agriculture 2016, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Oh, M.; Sakata, K.; Komatsu, S. Gel-free/label-free proteomic analysis of root tip of soybean over time under flooding and drought stresses. J. Prot. 2016, 130, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Rapala-Kozik, M.; Kowalska, E.; Ostrowska, K. Modulation of thiamine metabolism in Zea mays seedlings under conditions of abiotic stress. J. Exp. Bot. 2008, 59, 4133–4143. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Demesyeux, L.; Brym, M.; Chambers, A.H. Development of species-specific molecular markers in Vanilla for seedling selection of hybrids. Mol. Biol. Rep. 2020, 47, 1905–1920. [Google Scholar] [CrossRef]
- Divakaran, M.; Babu, K.; Peter, K. Cryopreservation of vanilla pollen and its utilization in inter specific hybridization between V. planifolia and V. aphylla. Eur. J. Biomed. Pharm. Sci. 2017, 4, 441–446. [Google Scholar]
- van Esse, H.; Reuber, T.; van der Does, D. Genetic modification to improve disease resistance in crops. New Phytol. 2019, 225, 70–86. [Google Scholar] [CrossRef] [Green Version]
- Flores-Jiménez, A.; Reyes-López, D.; Jiménez-García, D.; Romero-Arenas, O.; Rivera-Tapia, J.A.; Huerta-Lara, J.A.; Pérez-Silva, A. Diversidad de Vanilla spp. (Orchidaceae) y sus perfiles bioclimáticos en México. Rev. Biol. Trop. 2016, 65, 975–987. [Google Scholar] [CrossRef] [Green Version]
- Barreda-Castillo, J.M.; Menchaca-García, R.A.; Pérez-Silva, A.; Sánchez-Coello, N.G.; Luna-Rodríguez, M. Influencia de la temperatura en la infectividad de Fusarium oxysporum f. sp. vanillae en Vanilla planifolia y en híbridos V. planifolia × V. pompona. Biotecnia 2022, 25, 177–183. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.; Zhukov, I.; Tarakhovskaya, E.; Frolov, A.; et al. Methodology of drought stress research experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [Green Version]
- Van der Weele, C.; Spollen, W.; Sharp, R.; Baskin, T. Growth of Arabidopsis thaliana seedlings under water deficit studied by control of water potential in nutrient-agar media. J. Exp. Bot. 2000, 51, 1555–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xian-Pan, B.; Feng-Feng, J.; Feng-Tang, W.; Cai-Zhi, C.; Fu-Sun, Y. Response to Drought Stress Simulated by PEG of Phalaenopsis pulcherrima. J. Trop. Subtrop. Bot. 2019, 28, 53–61. [Google Scholar]
- Martínez-Santos, E.; Cruz-Cruz, C.A.; Spinoso-Castillo, J.L.; Bello-Bello, J.J. In vitro response of vanilla (Vanilla planifolia Jacks. ex Andrews) to PEG-induced osmotic stress. Sci. Rep. 2021, 11, 22611. [Google Scholar] [CrossRef] [PubMed]
- Najla, S.; Sanoubar, R.; Murshed, R. Morphological and biochemical changes in two parsley varieties upon water stress. Physiol. Mol. Biol. Plants 2012, 18, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niechayev, N.A.; Pereira, P.N.; Cushman, J.C. Understanding trait diversity associated with crassulacean acid metabolism (CAM). Curr. Opin. Plant Biol. 2019, 49, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Zotz, G.; Winkler, U. Aerial roots of epiphytic orchids: The velamen radicum and its role in water and nutrient uptake. Oecologia. 2013, 171, 733–741. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Arone, G. Morpho-anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress from Morphological to Molecular Features; Aroca, R., Ed.; Springer: Granada, Spain, 2012; pp. 37–63. [Google Scholar] [CrossRef]
- Karimi, S.; Rahemi, M.; Rostami, A.; Sedaghat, S. Drought effects on growth, water content and osmoprotectants in four olive cultivars with different drought tolerance. Int. J. Fruit Sci. 2018, 18, 254–267. [Google Scholar] [CrossRef]
- Gantiva, E.; Díez, C.; Moreno, F. Efecto de la interacción luz-agua sobre la fotosíntesis de la Vanilla planifolia (Orchidaceae). Rev. Biol. Trop. 2020, 68, 1250–1261. [Google Scholar]
- Carranza, C.; Lanchero, O.; Miranda, D.; Chaves, B. Growth analysis of ‘Batavia’ lettuce (Lactuca sativa L.) cultivated in a saline soil of the Bogota Plateau. Agron. Colomb. 2009, 27, 41–48. [Google Scholar]
- Arias, R.C.; Reyes, J.J.; Ray, J.V.; Benítez, D.G.; Hernández, L.G.; Ledea, J.L. 2019. Morphometric indicators in new megatermal varieties of cenchrus purpureus tolerant to water stress. Trop. Subtrop. Agroecos. 2019, 22, 115–125. [Google Scholar]
- Mata, A.; Jorge, T.; Pires, M.; Antonio, C. Drought stress tolerance in plants: Insights from metabolomics. In Drought Stress Tolerance in Plants, Vol. II. Molecular and Genetic Perspectives; Hossain, M., Wani, S., Bhattacharjee, S., Burrit, D., Tran, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 187–217. [Google Scholar] [CrossRef]
- Hietz, P.; Wagner, K.; Nunes Ramos, F.; Cabral, J.S.; Agudelo, C.; Benavides, A.M.; Cach-Pérez, M.J.; Cardelús, C.L.; Chilpa Galván, N.; Erickson Nascimento da Costa, L.; et al. Putting vascular epiphytes on the traits map. J. Ecol. 2022, 110, 340–358. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Villar, R. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Leyva, V.; Lopez, J.; Zevallos-Ventura, A.; Cabrera, R.; Cañari-Chumpitaz, C.; Toubiana, D.; Maruenda, H. NMR-based leaf metabolic profiling of V. planifolia and three endemic Vanilla species from the Peruvian Amazon. Food Chem. 2021, 358, 129365. [Google Scholar] [CrossRef]
- Liu, J.; Kang, R.; Liu, Y.; Wu, K.; Yan, X.; Song, Y.; Pan, L.; Thang, Z. Differential Metabolite Accumulation in Different Tissues of Gleditsia sinensis under Water Stress and Rehydration Conditions. Forests 2020, 11, 542–556. [Google Scholar] [CrossRef]
- Rai, V.K. Role of amino acids in plant responses to stresses. Biol. Plant 2002, 45, 481–487. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Emam, Y.; Pessarakli, M. Changes in endogenous hormonal status in corn (Zea mays) hybrids under drought stress. J. Plant Nutr. 2013, 36, 1695–1707. [Google Scholar] [CrossRef]
- Shanker, A.; Maheswari, M.; Yadav, S.; Desai, S.; Bhanu, D.; Attal, N.; Venkateswarlu, B. Drought stress responses in crops. Func. Int. Gen. 2014, 14, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Feizabadi, A.; Noormohammadi, G.; Fatehi, F. Changes in growth, physiology, and fatty acid profile of rapeseed cultivars treated with vermicompost under drought stress. J. Soil Sci. Plant Nutr. 2021, 21, 200–208. [Google Scholar] [CrossRef]
- Auler, P.A.; Souza, G.M.; da Silva Engela, M.R.G.; do Amaral, M.N.; Rossatto, T.; da Silva, M.G.Z.; Braga, E.J.B. Stress memory of physiological, biochemical and metabolomic responses in two different rice genotypes under drought stress: The scale matters. Plant Sci. 2021, 311, 110994. [Google Scholar] [CrossRef] [PubMed]
- Mibei, E.K.; Owino, W.O.; Ambuko, J.; Giovannoni, J.J.; Onyango, A.N. Metabolomic analyses to evaluate the effect of drought stress on selected African Eggplant accessions. J. Sci. Food Agric. 2018, 98, 205–216. [Google Scholar] [CrossRef]
- Khazaei, Z.; Esmaielpour, B.; Estaji, A. Ameliorative effects of ascorbic acid on tolerance to drought stress on pepper (Capsicum annuum L.) plants. Phys. Mol. Biol. Plants 2020, 26, 649–1662. [Google Scholar] [CrossRef]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wędzony, M. Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: Accumulation of ferulic acid correlates with drought tolerance. Ann. Bot. 2007, 100, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piasecka, A.; Sawikowska, A.; Kuczyńska, A.; Ogrodowicz, P.; Mikołajczak, K.; Krystkowiak, K.; Kachlicki, P. Drought-related secondary metabolites of barley (Hordeum vulgare L.) leaves and their metabolomic quantitative trait loci. Plant J. 2017, 89, 898–913. [Google Scholar] [CrossRef] [Green Version]
- Xuan, T.D.; Khang, D.T. Effects of exogenous application of protocatechuic acid and vanillic acid to chlorophylls, phenolics and antioxidant enzymes of rice (Oryza sativa L.) in submergence. Molecules 2018, 23, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Gai, Z.; Gui, T.; Chen, J.; Chen, Q.; Li, Y. Antioxidant Effects of Protocatechuic Acid and Protocatechuic Aldehyde: Old Wine in a New Bottle. Evid.-Based Complement. Altern. Med. 2021, 2021, 6139308. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Lupini, A.; Mauceri, A.; Zumbo, A.; Sunseri, F.; Abenavoli, M.R. The allelochemical trans-cinnamic acid stimulates salicylic acid production and galactose pathway in maize leaves: A potential mechanism of stress tolerance. Plant Physiol. Biochem. 2018, 128, 32–40. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Juliano, F.F.; Alvarenga, J.F.R.; Lamuela-Raventos, R.M.; Massarioli, A.P.; Lima, L.M.; Santos, R.C.; Alencar, S.M. Polyphenol analysis using high-resolution mass spectrometry allows differentiation of drought tolerant peanut genotypes. J. Sci. Food Agric. 2020, 100, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Shawon, R.A.; Kang, B.S.; Lee, S.G.; Kim, S.K.; Lee, H.J.; Katrich, E.; Ku, Y.G. Influence of drought stress on bioactive compounds, antioxidant enzymes and glucosinolate contents of Chinese cabbage (Brassica rapa). Food Chem. 2020, 308, 125657. [Google Scholar] [CrossRef] [PubMed]
- Król, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiol. Plant 2014, 36, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, R.D.; Kaur, L.; Srivastava, P. Comparative evaluation of different phenolic acids as priming agents for mitigating drought stress in wheat seedlings. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 1133–1142. [Google Scholar] [CrossRef]
- Yeloojeh, K.A.; Saeidi, G.; Sabzalian, M.R. Drought stress improves the composition of secondary metabolites in safflower flower at the expense of reduction in seed yield and oil content. Ind. Crops Prod. 2020, 154, 112496. [Google Scholar] [CrossRef]
- Guo, X.; Xin, Z.; Yang, T.; Ma, X.; Zhang, Y.; Wang, Z.; Lin, T. Metabolomics response for drought stress tolerance in Chinese wheat genotypes (Triticum aestivum). Plants 2020, 9, 520. [Google Scholar] [CrossRef] [Green Version]
- Jafari, M.; Shahsavar, A. The effect of foliar application of melatonin on changes in secondary metabolite contents in two Citrus species under drought stress conditions. Front. Plant Sci. 2021, 2021, 1509. [Google Scholar] [CrossRef]
- Mechri, B.; Tekaya, M.; Hammami, M.; Chehab, H. Effects of drought stress on phenolic accumulation in greenhouse-grown olive trees (Olea europaea). Biochem. Syst. Ecol. 2020, 92, 104112. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crops Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Kumar, M.; Patel, M.K.; Kumar, N.; Bajpai, A.B.; Siddique, K.H. Metabolomics and molecular approaches reveal drought stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 9108. [Google Scholar] [CrossRef] [PubMed]
- Hein, J.A.; Sherrard, M.E.; Manfredi, K.P.; Abebe, T. The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 2016, 16, 248. [Google Scholar] [CrossRef] [Green Version]
- Koo, Y.M.; Heo, A.Y.; Choi, H.W. Salicylic acid as a safe plant protector and growth regulator. Plant Pat. J. 2020, 36, 1. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 25, 33103–33118. [Google Scholar] [CrossRef]
- Belanger, F.; Havkin-Frenkel, D. Molecular analysis of a vanilla hybrid cultivated in Costa Rica. In Handbook of Vanilla Science and Technology; Havkin-Frenkel, D., Belanger, F., Eds.; Wiley: New York, NY, USA, 2019; Volume 2, pp. 391–402. [Google Scholar] [CrossRef]
- Pontes, M. Orquídeas, Notas Sobre Cultivo y más; Asociación Mexicana de Orquideología: Mexico City, Mexico, 2019; pp. 1–238. [Google Scholar]
- Park, H.S.; Lee, W.K.; Lee, S.C.; Lee, H.O.; Joh, H.J.; Park, J.Y.; Yang, T.J. Inheritance of chloroplast and mitochondrial genomes in cucumber revealed by four reciprocal F1 hybrid combinations. Sci. Rep. 2021, 11, 2506. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Gan, G.; Li, W.; Li, W.; Jiang, Y.; Liang, X.; Wang, Y. Inheritance of Solanum chloroplast genomic DNA in interspecific hybrids. Mitochondrial 2021, 6, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, A. Epigenetic regulation of abiotic stress tolerance in plants. Adv. Plants Agric. Res. 2016, 5, 179. [Google Scholar] [CrossRef]
- Baulcombe, D.C.; Dean, C. Epigenetic regulation in plant responses to the environment. Cold Spring Harb. Perspect. Biol. 2014, 6, a019471. [Google Scholar] [CrossRef]
- Chambers, A.H. Vanilla (Vanilla spp.) breeding. In Advances in Plant Breeding Strategies: Industrial and Food Crops; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Cincinnati, OH, USA, 2019; pp. 707–734. [Google Scholar] [CrossRef]
- Grisoni, M.; Nany, F. The beautiful hills: Half a century of vanilla (Vanilla planifolia Jacks. ex Andrews) breeding in Madagascar. Genet. Resour. Crop Evol. 2021, 68, 1691–1708. [Google Scholar] [CrossRef]
- Menchaca-García, R.A. Germinación in Vitro de Híbridos de Vanilla planifolia y V. pompona. Ph.D. Thesis, Universidad Veracruzana, Xalapa, Veracruz, Mexico, 2012. [Google Scholar]
- Lee-Espinosa, H.E.; Murguía-González, B.J.; García-Rosas, A.L.; Cordova-Contreras, A.; Luguna-Cerda, J.O.; Mijangos-Cortés, L.F.; Barahona-Pérez, L.G.; Iglesias-Andreu, L.G.; Santana-Buzzy, N. In vitro clonal propagation of Vanilla (Vanilla planifolia ‘Andrews’). Hortscience 2008, 43, 454–458. [Google Scholar] [CrossRef]
- Divakaran, M.; Babu, K.; Peter, K. Protocols for biotechnological interventions in improvement of vanilla (Vanilla planifolia Andrews). In Protocols for In Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants; Jain, M., Ed.; Springer: New York, NY, USA, 2016; pp. 47–63. [Google Scholar] [CrossRef] [Green Version]
- Michel, B.E. Evaluation of the water potentials of solutions of polyethylene glycol 8000 both in the absence and presence of other solutes. Plant Physiol. 1983, 72, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Castellanos, M.; Segura Abril, M.; Ñústez López, C.E. Growth analysis and source-sink relationship of four potato cultivars (Solanum tuberosum L.) in the Zipaquira Town (Cundinamarca, Colombia). Rev. Fac. Nac. Agron. Medellín 2010, 63, 5253–5266. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Met. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Calvete, E.O.; Kämpf, A.N.; Bergamaschi, H.; Daudt, R. Avaliação do crescimento de plantas de morangueiro, durante a aclimatização ex vitro. Hort. Bras. 2000, 18, 188–192. [Google Scholar] [CrossRef] [Green Version]
- Fernández, G.; Johnston, M. Fisiología Vegetal Experimental; Instituto Panamericano para la Agricultura: San José, Costa Rica, 1986; pp. 249–256. [Google Scholar]
- Monribot-Villanueva, J.L.; Altúzar-Molina, A.; Aluja, M.; Zamora-Briseño, J.A.; Elizalde-Contreras, J.M.; Bautista-Valle, M.; Arellano, J.; Sánchez-Martínez, D.; Rivera-Reséndiz, F.; Vázquez-Rosas-Landa, M.; et al. Integrating proteomics and metabolomics approaches to elucidate the ripening process in white Psidium guajava. Food Chem. 2022, 367, 130656. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Trujillo, N.; Monribot-Villanueva, J.L.; Alvarado-Olivarez, M.; Luna-Solano, G.; Guerrero-Analco, J.A.; Jiménez-Fernández, M. Phenolic profile and antioxidative properties of Pulp and sedes of Randia monatha Benth. Ind. Crops Prod. 2018, 124, 53–58. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 12 September 2022).
- Rmisc: Ryan Miscellaneous. Available online: https://CRAN.R-project.org/package=Rmisc (accessed on 12 September 2022).
- Agricolae: Statistical Procedures for Agricultural Research. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 12 September 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biomet. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; pp. 1–182. [Google Scholar]
- Xia, J.; Sinelnikov, I.; Han, B.; Wishart, D. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, 251–257. [Google Scholar] [CrossRef] [Green Version]
- KEGG PATHWAY Database. Wiring Diagrams of Molecular Interactions, Reactions, and Relations. Available online: https://www.genome.jp/kegg/pathway.html (accessed on 12 September 2022).
- Adler, M.; Alon, U. Fold-change detection in biological systems. Curr. Opt. Syst. Biol. 2018, 8, 81–89. [Google Scholar] [CrossRef]
- Gaffney, I.; Sallach, J.; Wilson, J.; Bergstrom, E.; Thomas-Oates, J. Metabolomic Approaches to Studying the Response to Drought Stress in Corn (Zea mays) Cobs. Metabolites 2021, 11, 438. [Google Scholar] [CrossRef] [PubMed]
- Melandri, G.; AbdElgawad, H.; Riewe, D.; Hageman, J.A.; Asard, H.; Beemster, G.; Bouwmeester, H. Biomarkers for grain yield stability in rice under drought stress. J. Exp. Bot. 2020, 71, 669–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaletti, A.; Naghavi, M.R.; Toorchi, M.; Zolla, L.; Rinalducci, S. Metabolomics and proteomics reveal drought-stress responses of leaf tissues from spring-wheat. Sci. Rep. 2018, 8, 5710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KEGG COMPOUND Database. Chemical Substances Integrated with Genomics. Available online: https://www.genome.jp/kegg/compound/ (accessed on 12 September 2022).






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barreda-Castillo, J.M.; Monribot-Villanueva, J.L.; Velázquez-Rosas, N.; Bayman, P.; Guerrero-Analco, J.A.; Menchaca-García, R.A. Morphological and Physio-Chemical Responses to PEG-Induced Water Stress in Vanilla planifolia and V. pompona Hybrids. Int. J. Mol. Sci. 2023, 24, 4690. https://doi.org/10.3390/ijms24054690
Barreda-Castillo JM, Monribot-Villanueva JL, Velázquez-Rosas N, Bayman P, Guerrero-Analco JA, Menchaca-García RA. Morphological and Physio-Chemical Responses to PEG-Induced Water Stress in Vanilla planifolia and V. pompona Hybrids. International Journal of Molecular Sciences. 2023; 24(5):4690. https://doi.org/10.3390/ijms24054690
Chicago/Turabian StyleBarreda-Castillo, José Martín, Juan L. Monribot-Villanueva, Noé Velázquez-Rosas, Paul Bayman, José A. Guerrero-Analco, and Rebeca Alicia Menchaca-García. 2023. "Morphological and Physio-Chemical Responses to PEG-Induced Water Stress in Vanilla planifolia and V. pompona Hybrids" International Journal of Molecular Sciences 24, no. 5: 4690. https://doi.org/10.3390/ijms24054690
APA StyleBarreda-Castillo, J. M., Monribot-Villanueva, J. L., Velázquez-Rosas, N., Bayman, P., Guerrero-Analco, J. A., & Menchaca-García, R. A. (2023). Morphological and Physio-Chemical Responses to PEG-Induced Water Stress in Vanilla planifolia and V. pompona Hybrids. International Journal of Molecular Sciences, 24(5), 4690. https://doi.org/10.3390/ijms24054690