
A Familial Novel Putative-Pathogenic Mutation Identified in Plaque-Psoriasis by a Multigene Panel Analysis

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. NGS Data Analysis
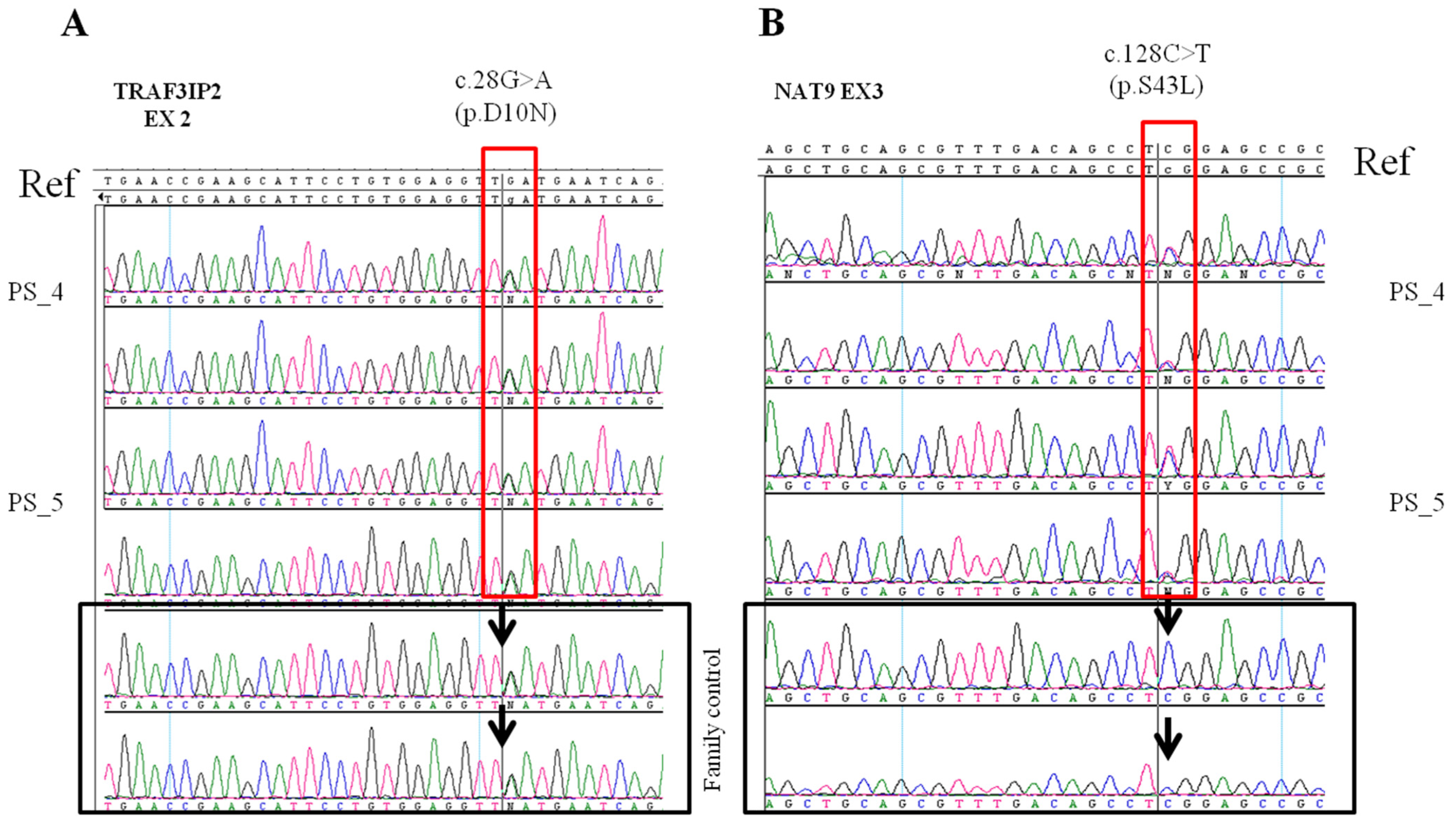
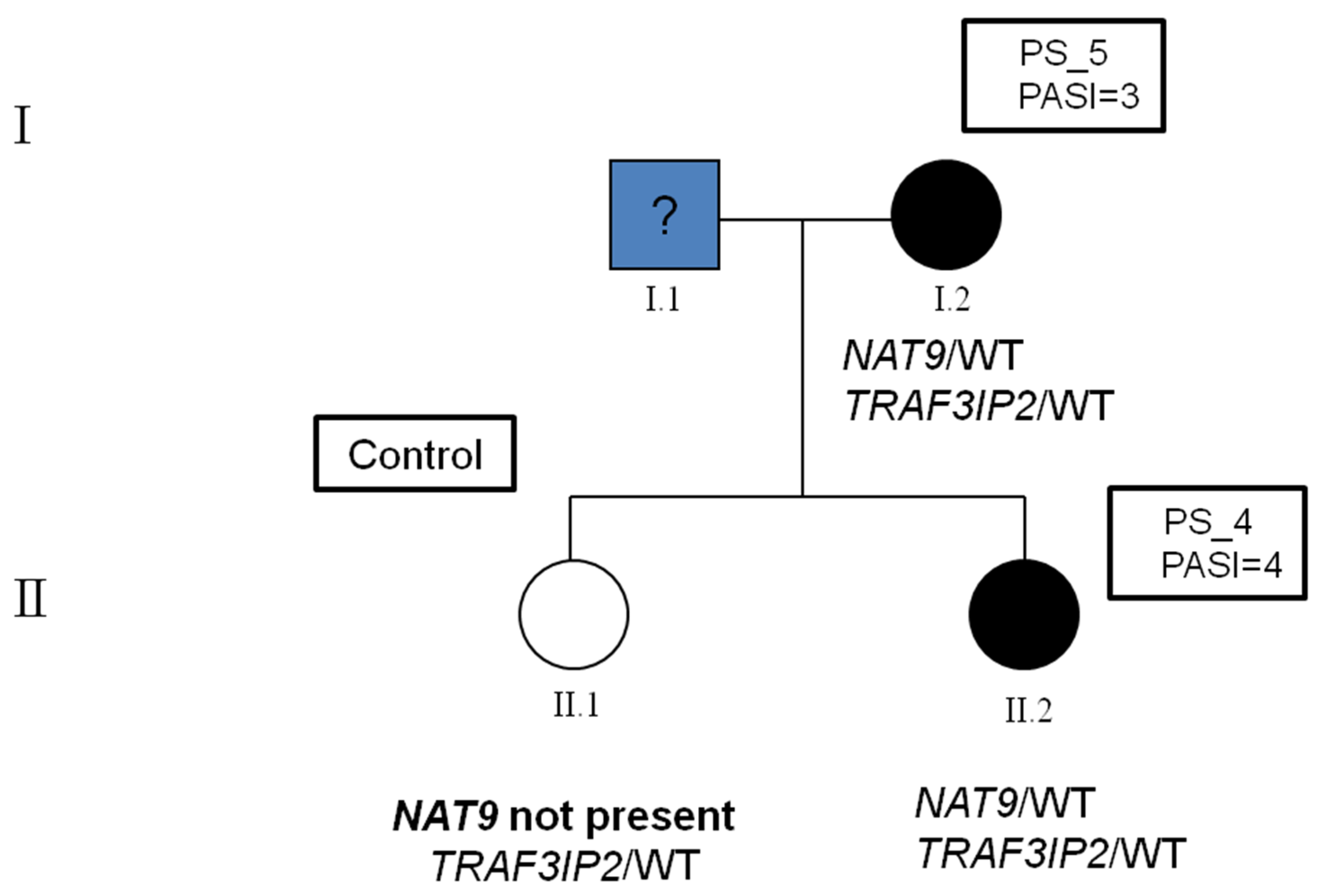
2.2. Identification of Variants in the Family
3. Materials and Methods
3.1. Patient Selection
3.2. Custom Panel Design
3.3. Sample Preparation and Sequencing
3.4. Bioinformatic Analysis
4. Conclusions and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Ghoreschi, K.; Balato, A.; Enerbäck, C.; Sabat, R. Therapeutics targeting the IL-23 and IL-17 pathway in psoriasis. Lancet 2021, 397, 754–766. [Google Scholar] [CrossRef]
- Wu, J.J.; Kavanaugh, A.; Lebwohl, M.G.; Gniadecki, R.; Merola, J.F. Psoriasis and metabolic syndrome: Implications for the management and treatment of psoriasis. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Prinz, J.C.; Choon, S.E.; Griffiths, C.E.M.; Merola, J.F.; Morita, A.; Ashcroft, D.M.; Viguier, M. Prevalence, comorbidities and mortality of generalized pustular psoriasis: A literature review. J. Eur. Acad. Dermatol. Venereol. 2022, 37, 256–273. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Ding, R.; Zhou, L.; Chen, X.; Shen, E. Epidemiology of Psoriasis and Comorbid Diseases: A Narrative Review. Front. Immunol. 2022, 13, 880201. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, A.; Lembo, S.; Di Caprio, R.; Donnarumma, G.; Monfrecola, G.; Balato, N.; Ayala, F.; Balato, A. Psoriatic cutaneous inflammation promotes human monocyte differentiation into active osteoclasts, facilitating bone damage. Eur. J. Immunol. 2017, 47, 1062–1074. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, J.E.; Yan, B.Y.; Chan, T.C.; Krueger, J.G. Discovery of the IL-23/IL-17 Signaling Pathway and the Treatment of Psoriasis. J. Immunol. 2018, 201, 1605–1613. [Google Scholar] [CrossRef] [Green Version]
- Michalek, I.M.; Loring, B.; John, S.M. A systematic review of worldwide epidemiology of psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef]
- Augustin, J.; Wolf, S.; Stephan, B.; Augustin, M.; Andrees, V. Psoriasis comorbidities in Germany: A population-based study on spatiotemporal variations. PLoS ONE 2022, 17, e0265741. [Google Scholar] [CrossRef]
- Dhabale, A.; Nagpure, S. Types of Psoriasis and Their Effects on the Immune System. Cureus 2022, 14, e29536. [Google Scholar] [CrossRef] [PubMed]
- Bachelez, H.; Barker, J.; Burden, A.D.; Navarini, A.A.; Krueger, J.G. Generalized pustular psoriasis is a disease distinct from psoriasis vulgaris: Evidence and expert opinion. Expert Rev. Clin. Immunol. 2022, 18, 1033–1047. [Google Scholar] [CrossRef]
- Iznardo, H.; Puig, L. Beyond plaque psoriasis-pathogenesis and treatment of other psoriasis phenotypes. Curr. Opin. Rheumatol. 2022, 34, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Montes de Oca Pedrosa, A.; Oakley, A.; Rogers, J.; Rangaitaha Epiha, M. PASI vs PO-PASI: Patient-Oriented PASI (PO-PASI) is comparable to clinician score PASI. Australas. J. Dermatol. 2022, 63, e67–e70. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N. Psoriasis: Pathogenesis, comorbidities, and therapy updated. Int. J. Mol. Sci. 2021, 22, 2979. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Okada, Y. The current landscape of psoriasis genetics in 2020. J. Dermatol. Sci. 2020, 99, 2–8. [Google Scholar] [CrossRef]
- Kocaaga, A.; Kocaaga, M. Psoriasis: An Immunogenetic Perspective. Glob. Med. Genet. 2022, 9, 82–89. [Google Scholar] [CrossRef]
- Esposito, M.V.; Fosso, B.; Nunziato, M.; Casaburi, G.; D’Argenio, V.; Calabrese, A.; D’Aiuto, M.; Botti, G.; Pesole, G.; Salvatore, F. Microbiome composition indicate dysbiosis and lower richness in tumor breast tissues compared to healthy adjacent paired tissue, within the same women. BMC Cancer 2022, 22, 30. [Google Scholar] [CrossRef]
- Nardelli, C.; Granata, I.; Nunziato, M.; Setaro, M.; Carbone, F.; Zulli, C.; Pilone, V.; Capoluongo, E.D.; De Palma, G.D.; Corcione, F.; et al. 16s rRNA of mucosal colon microbiome and CCL2 circulating levels are potential biomarkers in colorectal cancer. Int. J. Mol. Sci. 2021, 22, 10747. [Google Scholar] [CrossRef]
- Dand, N.; Mahil, S.K.; Capon, F.; Smith, C.H.; Simpson, M.A.; Barker, J.N. Psoriasis and genetics. Acta Derm. Venereol. 2020, 100, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Ran, D.; Cai, M.; Zhang, X. Genetics of psoriasis: A basis for precision medicine. Precis. Clin. Med. 2019, 2, 120–130. [Google Scholar] [CrossRef]
- Tsoi, L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; Kang, H.M.; et al. Identification of fifteen new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef] [Green Version]
- Signa, S.; Campione, E.; Rusmini, M.; Chiesa, S.; Grossi, A.; Omenetti, A.; Caorsi, R.; Viglizzo, G.M.; Galluzzo, M.; Bianchi, L.; et al. Whole exome sequencing approach to childhood onset familial erythrodermic psoriasis unravels a novel mutation of CARD14 requiring unusual high doses of ustekinumab. Pediatr. Rheumatol. 2019, 17, 38. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, C.M.; Ramírez, C.P.; Martín, A.S.; Maroun, S.V.; Santiago, S.A.A.; del Carmen Ramírez Tortosa, M.; Morales, A.J. Influence of genetic polymorphisms on response to biologics in moderate-to-severe psoriasis. J. Pers. Med. 2021, 11, 293. [Google Scholar] [CrossRef]
- Ni, X.; Lai, Y. Keratinocyte: A trigger or an executor of psoriasis? J. Leukoc. Biol. 2020, 108, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Cibrian, D.; de la Fuente, H.; Sánchez-Madrid, F. Metabolic Pathways That Control Skin Homeostasis and Inflammation. Trends Mol. Med. 2020, 26, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, H.O.; Kim, J.C.; Park, C.W.; Chung, B.Y. Effects of Autophagy Modulators and Dioxin on the Expression of Epidermal Differentiation Proteins on Psoriasis-Like Keratinocytes in vitro and ex vivo. Clin. Cosmet. Investig. Dermatol. 2022, 15, 1149–1156. [Google Scholar] [CrossRef]
- ELinghaus, E.; ELinghaus, D.; Stuart, P.E.; Nair, R.P.; Debrus, S.; Raelson, J.V.; Belouchi, M.; Fournier, H.; Reinhard, C.; Ding, J.; et al. Genome-wide asociation study identifies a psoriasis susceptibility locus at TRAF3IP2. Nat. Genet. 2010, 42, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, D.; Białecka, M.; Bojko, P.; Kiszkielis, A.; Droździk, M.; Kurzawski, M. Analysis of selected genetic variants in psoriasis susceptibility and response to treatment. Adv. Dermatol. Allergol. 2022, 39, 934–939. [Google Scholar] [CrossRef]
- Helms, C.; Cao, L.; Krueger, J.G.; Wijsman, E.M.; Chamian, F.; Gordon, D.; Heffernan, M.; Wright Daw, J.A.; Robarge, J.; Ott, J.; et al. A putative RUNX1 binding site variant between SLC9A3R1 and NAT9 is associated with susceptibility to psoriasis. Nat. Genet. 2003, 35, 349–356. [Google Scholar] [CrossRef]
- Filer, C.; Ho, P.; Bruce, I.N.; Worthington, J.; Barton, A. Investigation of association of genes NAT9, SLC9A3R1 and RAPTOR on chromosome 17q25 with psoriatic arthritis. Ann. Rheum. Dis. 2009, 68, 291–292. [Google Scholar] [CrossRef]
- Bowcock, A.M.; Cookson, W.O.C.M. The genetics of psoriasis, psoriatic arthritis and atopic dermatitis. Hum. Mol. Genet. 2004, 13, 43–55. [Google Scholar] [CrossRef]
- Nunziato, M.; Esposito, M.V.; Starnone, F.; Diroma, M.A.; Calabrese, A.; del Monaco, V.; Buono, P.; Frasci, G.; Botti, G.; D’Aiuto, M.; et al. A multi-gene panel beyond BRCA1/BRCA2 to identify new breast cancer-predisposing mutations by a picodroplet PCR followed by a next-generation sequencing strategy: A pilot study. Anal. Chim. Acta 2018, 1046, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Nunziato, M.; Di Maggio, F.; Pensabene, M.; Esposito, M.V.; Starnone, F.; De Angelis, C.; Calabrese, A.; D’Aiuto, M.; Botti, G.; De Placido, S.; et al. Multi-gene panel testing increases germline predisposing mutations’ detection in a cohort of breast/ovarian cancer patients from Southern Italy. Front. Med. 2022, 9, 894358. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, V.; Esposito, M.V.; Nunziato, M.; De Simone, A.; Buono, P.; Salvatore, F.; Frisso, G. Molecular diagnosis of Brugada syndrome via next-generation sequencing of a multigene panel in a young athlete Diagnosi molecolare di sindrome di Brugada in un. Med. Dello Sport 2018, 71, 27–34. [Google Scholar] [CrossRef]
- Rezaei, N.; Rajabi, F. The Immunogenetics of Dermatologic Diseases; Springer: Cham, Switzerland, 2022; ISBN 9783030926151. [Google Scholar]
- Raposo, R.A.; Gupta, R.; Abdel-Mohsen, M.; Dimon, M.; Debbaneh, M.; Jiang, W.; York, V.A.; Leadabrand, K.S.; Brown, G.; Malakouti, M.; et al. Antiviral gene expression in psoriasis. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 1951–1957. [Google Scholar] [CrossRef] [Green Version]
- Alibardi, L. Immunolocalization of serpins in the regenerating tail of lizard suggests a role for epidermal and neural barrier formation. Zoology 2018, 131, 1–9. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ID NGS | Mean Region Coverage Depth | Target Coverage at 50× | Percent Aligned Reads | Variants in Genes Presented in the Panel | Variants in Coding Regions |
|---|---|---|---|---|---|
| PS_4 | 271.9 | 97.50% | 99.30% | 3135 | 456 |
| PS_5 | 222 | 96.60% | 99.40% | 3048 | 456 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunziato, M.; Balato, A.; Ruocco, A.; D’Argenio, V.; Di Caprio, R.; Balato, N.; Ayala, F.; Salvatore, F. A Familial Novel Putative-Pathogenic Mutation Identified in Plaque-Psoriasis by a Multigene Panel Analysis. Int. J. Mol. Sci. 2023, 24, 4743. https://doi.org/10.3390/ijms24054743
Nunziato M, Balato A, Ruocco A, D’Argenio V, Di Caprio R, Balato N, Ayala F, Salvatore F. A Familial Novel Putative-Pathogenic Mutation Identified in Plaque-Psoriasis by a Multigene Panel Analysis. International Journal of Molecular Sciences. 2023; 24(5):4743. https://doi.org/10.3390/ijms24054743
Chicago/Turabian StyleNunziato, Marcella, Anna Balato, Anna Ruocco, Valeria D’Argenio, Roberta Di Caprio, Nicola Balato, Fabio Ayala, and Francesco Salvatore. 2023. "A Familial Novel Putative-Pathogenic Mutation Identified in Plaque-Psoriasis by a Multigene Panel Analysis" International Journal of Molecular Sciences 24, no. 5: 4743. https://doi.org/10.3390/ijms24054743
APA StyleNunziato, M., Balato, A., Ruocco, A., D’Argenio, V., Di Caprio, R., Balato, N., Ayala, F., & Salvatore, F. (2023). A Familial Novel Putative-Pathogenic Mutation Identified in Plaque-Psoriasis by a Multigene Panel Analysis. International Journal of Molecular Sciences, 24(5), 4743. https://doi.org/10.3390/ijms24054743