
Transcriptome Analysis of Differentially Expressed Genes Associated with Salt Stress in Cowpea (Vigna unguiculata L.) during the Early Vegetative Stage
 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Physiological Responses to Salt Stress in Cowpea Germplasms
2.2. Illumina Sequencing Pre-Processing, and Read Mapping
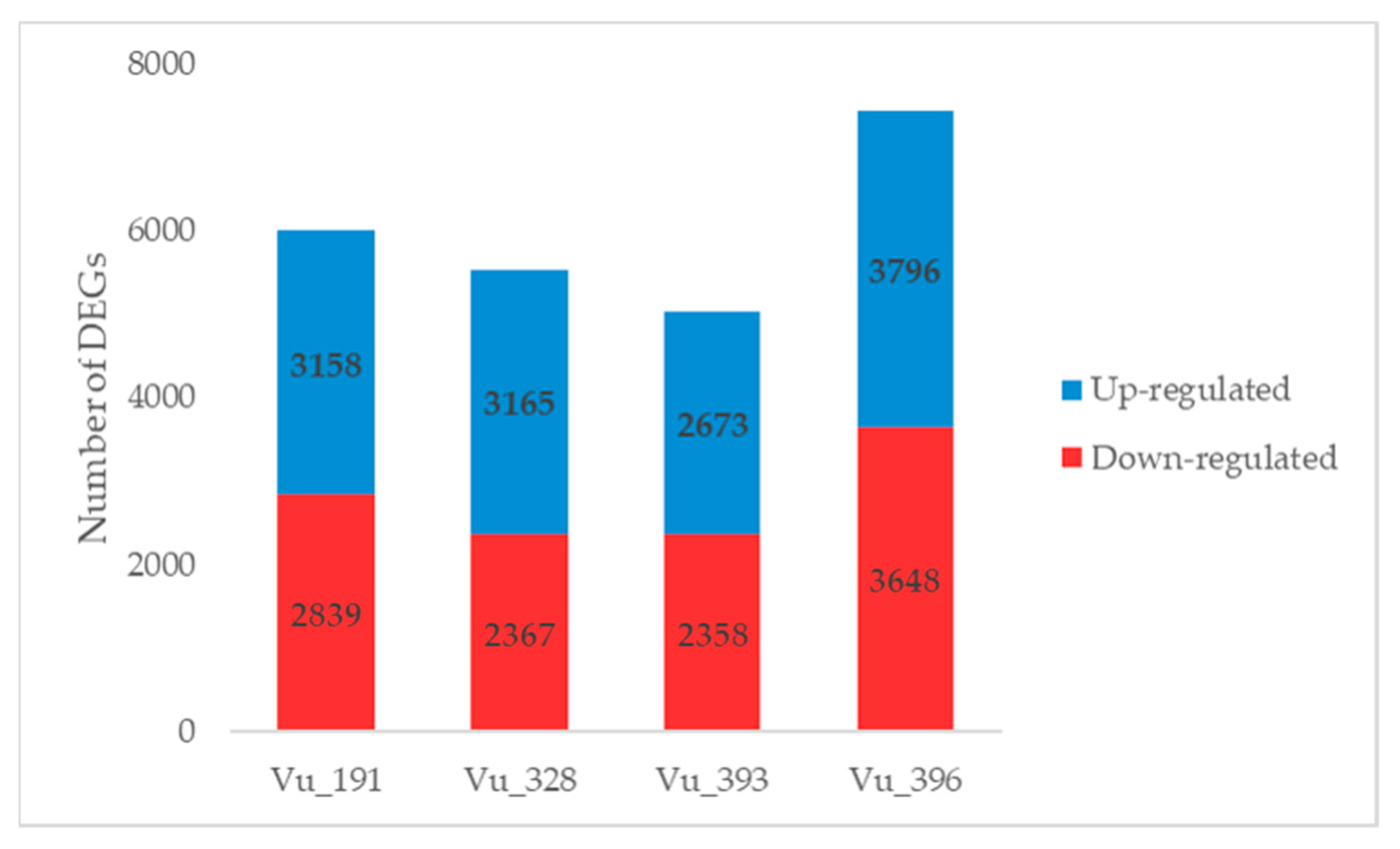
2.3. Identification of DEGs in Cowpea Germplasms with Different Salt Tolerance Levels
2.4. Clustering Analysis of the Identified DEGs
2.5. Identification of Variations in the Target Gene
2.6. Validation of the Variations in the Candidate Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Phenotyping of Salt Tolerance
4.2. Salt Treatment
4.3. RNA Extraction, Construction of cDNA Libraries and Short Read Sequencing
4.4. Sequence Pre-Processing and Mapping of RNA-Seq Reads
4.5. Identification of Differentially Expressed Genes (DEGs)
4.6. Quantitative Reverse Transcription PCR (qRT-PCR) for Validation of DEGs
4.7. Whole-Genome Resequencing and DNA Sequencing
4.8. Kompetitive Allele-Specific PCR(KASP) Primer Design and Validation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ehlers, J.; Hall, A. Cowpea (Vigna unguiculata L. walp.). Field Crops Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Rachie, K.; Roberts, L. Grain legumes of the lowland tropics. Adv. Agron. 1974, 26, 1–132. [Google Scholar]
- Food and Agriculture Organization of the United Nations. FAOSTAT Database 2020. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 16 December 2022).
- Metternicht, G.I.; Zinck, J. Remote sensing of soil salinity: Potentials and constraints. Remote Sens. Environ. 2003, 85, 1–20. [Google Scholar] [CrossRef]
- Sultan, B.; Gaetani, M. Agriculture in West Africa in the twenty-first century: Climate change and impacts scenarios, and potential for adaptation. Front. Plant Sci. 2016, 7, 1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylla, M.B.; Nikiema, P.M.; Gibba, P.; Kebe, I.; Klutse, N.A.B. Climate change over West Africa: Recent trends and future projections. In Adaptation to Climate Change and Variability in Rural West Africa; Springer: Cham, Sitzerland, 2016; pp. 25–40. [Google Scholar]
- Lacerda, C.F.; Assis Júnior, J.O.; Lemos Filho, L.C.; Oliveira, T.S.d.; Guimarães, F.V.; Gomes-Filho, E.; Prisco, J.T.; Bezerra, M.A. Morpho-physiological responses of cowpea leaves to salt stress. Braz. J. Plant Physiol. 2006, 18, 455–465. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A.M. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-K. Plant salt stress. In eLS; University of California: Riverside, CA, USA, 2007. [Google Scholar]
- Acosta-Motos, J.R.; Penella, C.; Hernández, J.A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Navarro, J.M.; Gómez-Bellot, M.J.; Barba-Espín, G. Towards a sustainable agriculture: Strategies involving phytoprotectants against salt stress. Agronomy 2020, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Chowdhury, N. Salt stress in plants and amelioration strategies: A critical review. Abiotic Stress Plants 2020. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Jagendorf, A.; Zhu, J.K. Understanding and improving salt tolerance in plants. Crop Sci. 2005, 45, 437–448. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Gogoi, N.; Hussain, M.; Barthakur, S.; Paul, S.; Bharadwaj, N.; Migdadi, H.M.; Alghamdi, S.S.; Siddique, K.H. Effects, tolerance mechanisms and management of salt stress in grain legumes. Plant Physiol. Biochem. 2017, 118, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Beszterda, M.; Goliński, P. ABA: Role in plant signaling under salt stress. In Salt Stress in Plants; Springer: New York, NY, USA, 2013; pp. 175–196. [Google Scholar]
- Sudhir, P.; Murthy, S. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 2004, 42, 481–486. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Le, L.T.T.; Kotula, L.; Siddique, K.H.; Colmer, T.D. Na+ and/or Cl− toxicities determine salt sensitivity in soybean (Glycine max (L.) Merr.), mungbean (Vigna radiata (L.) R. Wilczek), cowpea (Vigna unguiculata (L.) Walp.), and common Bean (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2021, 22, 1909. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Turner, N.C.; Colmer, T.D.; Quealy, J.; Pushpavalli, R.; Krishnamurthy, L.; Kaur, J.; Singh, G.; Siddique, K.H.; Vadez, V. Salinity tolerance and ion accumulation in chickpea (Cicer arietinum L.) subjected to salt stress. Plant Soil 2013, 365, 347–361. [Google Scholar] [CrossRef]
- Maas, E.; Poss, J. Salt sensitivity of cowpea at various growth stages. Irrig. Sci. 1989, 10, 313–320. [Google Scholar] [CrossRef]
- Ravelombola, W.S.; Shi, A.; Weng, Y.; Clark, J.; Motes, D.; Chen, P.; Srivastava, V. Evaluation of salt tolerance at germination stage in cowpea [Vigna unguiculata (L.) Walp]. HortScience 2017, 52, 1168–1176. [Google Scholar] [CrossRef] [Green Version]
- Murillo-Amador, B.; Troyo-Diéguez, E.; García-Hernández, J.L.; López-Aguilar, R.; Avila-Serrano, N.Y.; Zamora-Salgado, S.; Rueda-Puente, E.O.; Kaya, C. Effect of NaCl salinity in the genotypic variation of cowpea (Vigna unguiculata) during early vegetative growth. Sci. Hortic. 2006, 108, 423–431. [Google Scholar] [CrossRef]
- Al-Ashkar, I.; Alderfasi, A.; Ben Romdhane, W.; Seleiman, M.F.; El-Said, R.A.; Al-Doss, A. Morphological and genetic diversity within salt tolerance detection in eighteen wheat genotypes. Plants 2020, 9, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Chiodini, R.; Badr, A.; Zhang, G. The impact of next-generation sequencing on genomics. J. Genet. Genom. 2011, 38, 95–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, V.S. RNA Sequencing as a Tool for Understanding Biological Complexity of Abiotic Stress in Plants; Springer: Cham, Sitzerland, 2016; Volume 25, pp. 1–2. [Google Scholar]
- Marguerat, S.; Bähler, J. RNA-seq: From technology to biology. Cell. Mol. Life Sci. 2010, 67, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Bahieldin, A.; Atef, A.; Sabir, J.S.; Gadalla, N.O.; Edris, S.; Alzohairy, A.M.; Radhwan, N.A.; Baeshen, M.N.; Ramadan, A.M.; Eissa, H.F. RNA-Seq analysis of the wild barley (H. spontaneum) leaf transcriptome under salt stress. Comptes Rendus Biol. 2015, 338, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, H.; Chen, L.; Lou, Y.; Amombo, E.; Fu, J. RNA-seq for gene identification and transcript profiling in relation to root growth of bermudagrass (Cynodon dactylon) under salinity stress. BMC Genom. 2015, 16, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, R.; Pan, X.; Zhang, J.; Ma, Y.; He, S.; Zheng, Y.; Ma, Y. Effect of salt-stress on gene expression in citrus roots revealed by RNA-seq. Funct. Integr. Genom. 2018, 18, 155–173. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, G.; Du, L.; Shang, X.; Cheng, C.; Yang, B.; Hu, Y.; Cai, C.; Guo, W. Genetic regulation of salt stress tolerance revealed by RNA-Seq in cotton diploid wild species, Gossypium davidsonii. Sci. Rep. 2016, 6, 20582. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Lei, L.; Zheng, H.; Bi, Y.; Yang, L.; Liu, H.; Wang, J.; Sun, J.; Zhao, H.; Li, X.; Li, J. Identification of a major QTL and candidate gene analysis of salt tolerance at the bud burst stage in rice (Oryza sativa L.) using QTL-Seq and RNA-Seq. Rice 2020, 13, 55. [Google Scholar] [CrossRef]
- James, M.; Masclaux-Daubresse, C.; Marmagne, A.; Azzopardi, M.; Laîné, P.; Goux, D.; Etienne, P.; Trouverie, J. A new role for SAG12 cysteine protease in roots of Arabidopsis thaliana. Front. Plant Sci. 2019, 9, 1998. [Google Scholar] [CrossRef]
- Grudkowska, M.; Zagdańska, B. Multifunctional role of plant cysteine proteinases. Acta Biochim. Pol. 2004, 51, 609–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagorchev, L.; Kamenova, P.; Odjakova, M. The role of plant cell wall proteins in response to salt stress. Sci. World J. 2014, 2014, 764089. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-C.; Kim, S.-Y.; Paek, K.-H.; Choi, D.; Park, J.M. Suppression of CaCYP1, a novel cytochrome P450 gene, compromises the basal pathogen defense response of pepper plants. Biochem. Biophys. Res. Commun. 2006, 345, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Joshi, M.; Wang, S.; Johnson, C.D.; Joshi, V. Comparative analysis of root transcriptome profiles of sesame (Sesamum indicum L.) in response to osmotic stress. J. Plant Growth Regul. 2021, 40, 1787–1801. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa, A.M.; Martins, C.d.P.S.; Goncalves, L.P.; Costa, M.G.C. Late embryogenesis abundant (LEA) constitutes a large and diverse family of proteins involved in development and abiotic stress responses in sweet orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0145785. [Google Scholar] [CrossRef] [Green Version]
- Wise, M.J.; Tunnacliffe, A. POPP the question: What do LEA proteins do? Trends Plant Sci. 2004, 9, 13–17. [Google Scholar] [CrossRef]
- Chourey, K.; Ramani, S.; Apte, S.K. Accumulation of LEA proteins in salt (NaCl) stressed young seedlings of rice (Oryza sativa L.) cultivar Bura Rata and their degradation during recovery from salinity stress. J. Plant Physiol. 2003, 160, 1165–1174. [Google Scholar] [CrossRef]
- Cuevas-Velazquez, C.L.; Saab-Rincón, G.; Reyes, J.L.; Covarrubias, A.A. The unstructured N-terminal region of Arabidopsis group 4 late embryogenesis abundant (LEA) proteins is required for folding and for chaperone-like activity under water deficit. J. Biol. Chem. 2016, 291, 10893–10903. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Nguyen, C.T.T.; Jung, C.; Cheong, J.-J. AtMYB44 suppresses transcription of the late embryogenesis abundant protein gene AtLEA4-5. Biochem. Biophys. Res. Commun. 2019, 511, 931–934. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.-U.; Abo, M. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Heider, M.R.; Munson, M. Exorcising the exocyst complex. Traffic 2012, 13, 898–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Szczepanek, A.; Abdel-Haleem, H. Genome-wide association study (GWAS) analysis of camelina seedling germination under salt stress condition. Agronomy 2020, 10, 1444. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Nat. Preced. 2010, 11, R106. [Google Scholar]
- Lucas, A. Another Multidimensional Analysis Package. 2006. Available online: http://pbil.univ-lyon1.fr/R/pdf/tdr52.pdf (accessed on 16 December 2022).
- Warnes, M.G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W. Package ‘gplots’. Var. R Program. Tools Plotting Data. 2016. Available online: https://cran.microsoft.com/snapshot/2016-03-30/web/packages/gplots/gplots.pdf (accessed on 16 December 2022).
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols; Springer: Cham, Sitzerland, 2000; pp. 365–386. [Google Scholar]
- Amorim, L.L.B.; Ferreira-Neto, J.R.C.; Bezerra-Neto, J.P.; Pandolfi, V.; de Araújo, F.T.; da Silva Matos, M.K.; Santos, M.G.; Kido, E.A.; Benko-Iseppon, A.M. Cowpea and abiotic stresses: Identification of reference genes for transcriptional profiling by qPCR. Plant Methods 2018, 14, 88. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Juhos, S.; Larsson, M.; Olason, P.I.; Martin, M.; Eisfeldt, J.; DiLorenzo, S.; Sandgren, J.; De Ståhl, T.D.; Ewels, P. Sarek: A portable workflow for whole-genome sequencing analysis of germline and somatic variants. F1000Research 2020, 9, 63. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene id | Position | Log2FoldChange (FC) for Identified DEGs | Annotation | |||
|---|---|---|---|---|---|---|
| Vu_191 | Vu_328 | Vu_393 | Vu_396 | |||
| Vigun01g124200.1 | 30,107,016–30,107,917 | 3.71 | 2.51 | −0.04 | 0.66 | LATE EMBRYOGENESIS ABUNDANT PROTEIN 4–5 |
| Vigun02g076100.1 | 22,812,209–22,818,056 | 3.35 | −0.53 | 0.03 | −0.91 | POTASSIUM TRANSPORTER 6 |
| Vigun02g087000.1 | 24,173,651–24,175,542 | −2.55 | −1.58 | −1.40 | 1.73 | ALCOHOL DEHYDROGENASE-RELATED |
| Vigun02g150700.1 | 29,780,314–29,782,539 | 3.70 | 0.77 | 0.71 | −1.12 | N-TERMINAL ACETYLTRANSFERASE |
| Vigun02g156800.1 | 30,300,350–30,306,429 | 4.74 | −0.79 | 0.76 | −2.85 | OLIGOPEPTIDE TRANSPORTER-RELATED |
| Vigun03g036000.1 | 2,770,941–2,774,105 | 0.19 | −0.98 | −1.24 | −2.64 | RING-H2 FINGER PROTEIN ATL69-RELATED |
| Vigun03g195700.1 | 27,574,151–27,575,892 | −0.49 | −0.86 | −1.45 | −2.13 | CYTOCHROME P450 89A2-RELATED |
| Vigun03g323700.1 | 51,942,392–51,948,451 | 3.10 | −0.56 | 0.25 | −2.84 | ANION EXCHANGE PROTEIN |
| Vigun03g411400.1 | 61,890,298–61,892,363 | −1.54 | 0.18 | −0.65 | 1.31 | PEROXIDASE 40 |
| Vigun06g049900.1 | 17,502,581–17,504,135 | 1.89 | 1.00 | 0.25 | −2.16 | Hydroxycinnamate 4-beta-glucosyltransferase |
| Vigun06g206600.1 | 32,059,029–32,059,206 | 0.84 | −0.07 | −1.34 | −1.61 | Unknown |
| Vigun07g005500.1 | 439,864–440,793 | 3.34 | 1.49 | 1.55 | −2.61 | Hemopexin |
| Vigun07g044100.1 | 4,471,515–4,474,926 | −2.91 | 0.52 | 0.32 | 2.81 | 3-oxoacyl-[acyl-carrier-protein] reductase |
| Vigun07g065400.1 | 7,663,218–7,665,263 | −0.93 | −0.34 | 2.22 | 1.80 | Myb/SANT-like DNA-binding domain (Myb_DNA-bind_4) |
| Vigun07g164700.1 | 27,692,362–27,695,281 | 0.64 | −3.29 | −4.63 | −4.02 | ALUMINUM-ACTIVATED MALATE TRANSPORTER 10 |
| Vigun07g217900.1 | 33,993,108–33,994,160 | 0.14 | −4.35 | −4.41 | −5.92 | Uncharacterized membrane protein |
| Vigun08g025300.1 | 2,190,472–2,192,518 | −0.24 | −1.85 | −0.80 | −3.35 | HEAT STRESS TRANSCRIPTION FACTOR B-4 |
| Vigun08g090000.1 | 20,808,945–20,813,724 | 4.14 | −0.69 | −0.46 | −1.53 | EamA-like transporter family (EamA) |
| Vigun08g116200.1 | 28,318,383–28,321,227 | 2.68 | −1.19 | −0.60 | −2.22 | COPPER TRANSPORT PROTEIN ATOX1-RELATED |
| Vigun08g125100.1 | 29,512,430–29,514,230 | 0.25 | 0.42 | 5.48 | 7.06 | EXOCYST COMPLEX PROTEIN EXO70 |
| Vigun09g086300.1 | 11,333,702–11,336,177 | −2.42 | 1.31 | 2.73 | 3.33 | INACTIVE POLY [ADP-RIBOSE] POLYMERASE SRO4-RELATED |
| Vigun10g015100.1 | 1,666,906–1,668,606 | 1.63 | −0.20 | −1.32 | −1.96 | CYCLIN-U4-1 |
| Vigun10g180000.1 | 39,812,625–39,813,195 | −0.79 | 0.97 | 1.66 | 2.02 | Cotton fiber expressed protein (DUF761) |
| Vigun11g017700.1 | 2,206,680–2,207,648 | −1.29 | 0.42 | 2.80 | 1.52 | Unknown |
| Vigun11g018800.1 | 2,332,367–2,339,763 | −0.10 | −0.35 | 1.67 | 1.88 | protein regulator of cytokinesis 1 (PRC1) |
| Vigun11g126800.1 | 33,386,966–33,389,552 | 1.96 | −0.74 | 0.59 | −2.28 | MYB FAMILY TRANSCRIPTION FACTOR-RELATED |
| Vigun11g182400.1 | 38,574,316–38,575,114 | 0.20 | −0.76 | −1.17 | −1.24 | SAUR family protein (SAUR) |
| Sample | Salt Tolerance Type | Vigun_02G076100 | Vigun_08G125100 |
|---|---|---|---|
| Vu_035 | Sensitive | REF (AAA, K) | Existence |
| Vu_266 | Sensitive | REF (AAA, K) | - |
| Vu_296 | Sensitive | REF (AAA, K) | Existence |
| Vu_318 | Sensitive | REF (AAA, K) | Existence |
| Vu_319 | Sensitive | REF (AAA, K) | - |
| Vu_343 | Sensitive | REF (AAA, K) | - |
| Vu_348 | Sensitive | REF (AAA, K) | - |
| Vu_393 * | Sensitive | REF (AAA, K) | Existence |
| Vu_396 * | Sensitive | REF (AAA, K) | Existence |
| Vu_403 | Sensitive | REF (AAA, K) | - |
| Vu_055 | Resistant | REF (AAA, K) | - |
| Vu_095 | Resistant | REF (AAA, K) | - |
| Vu_111 | Resistant | SNP (GAA, E) | - |
| Vu_129 | Resistant | REF (AAA, K) | - |
| Vu_147 | Resistant | REF (AAA, K) | - |
| Vu_166 | Resistant | REF (AAA, K) | - |
| Vu_191 * | Resistant | SNP (GAA, E) | - |
| Vu_328 * | Resistant | REF (AAA, K) | - |
| Vu_336 | Resistant | REF (AAA, K) | - |
| Vu_352 | Resistant | REF (AAA, K) | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, B.H.; Kim, W.J.; Chowdhury, S.; Moon, C.Y.; Kang, S.; Kim, S.-H.; Jo, S.-H.; Jun, T.-H.; Kim, K.D.; Ha, B.-K. Transcriptome Analysis of Differentially Expressed Genes Associated with Salt Stress in Cowpea (Vigna unguiculata L.) during the Early Vegetative Stage. Int. J. Mol. Sci. 2023, 24, 4762. https://doi.org/10.3390/ijms24054762
Kang BH, Kim WJ, Chowdhury S, Moon CY, Kang S, Kim S-H, Jo S-H, Jun T-H, Kim KD, Ha B-K. Transcriptome Analysis of Differentially Expressed Genes Associated with Salt Stress in Cowpea (Vigna unguiculata L.) during the Early Vegetative Stage. International Journal of Molecular Sciences. 2023; 24(5):4762. https://doi.org/10.3390/ijms24054762
Chicago/Turabian StyleKang, Byeong Hee, Woon Ji Kim, Sreeparna Chowdhury, Chang Yeok Moon, Sehee Kang, Seong-Hoon Kim, Sung-Hwan Jo, Tae-Hwan Jun, Kyung Do Kim, and Bo-Keun Ha. 2023. "Transcriptome Analysis of Differentially Expressed Genes Associated with Salt Stress in Cowpea (Vigna unguiculata L.) during the Early Vegetative Stage" International Journal of Molecular Sciences 24, no. 5: 4762. https://doi.org/10.3390/ijms24054762
APA StyleKang, B. H., Kim, W. J., Chowdhury, S., Moon, C. Y., Kang, S., Kim, S. -H., Jo, S. -H., Jun, T. -H., Kim, K. D., & Ha, B. -K. (2023). Transcriptome Analysis of Differentially Expressed Genes Associated with Salt Stress in Cowpea (Vigna unguiculata L.) during the Early Vegetative Stage. International Journal of Molecular Sciences, 24(5), 4762. https://doi.org/10.3390/ijms24054762