
Exploration of the Shared Molecular Mechanisms between COVID-19 and Neurodegenerative Diseases through Bioinformatic Analysis

Abstract
:1. Introduction
2. Results
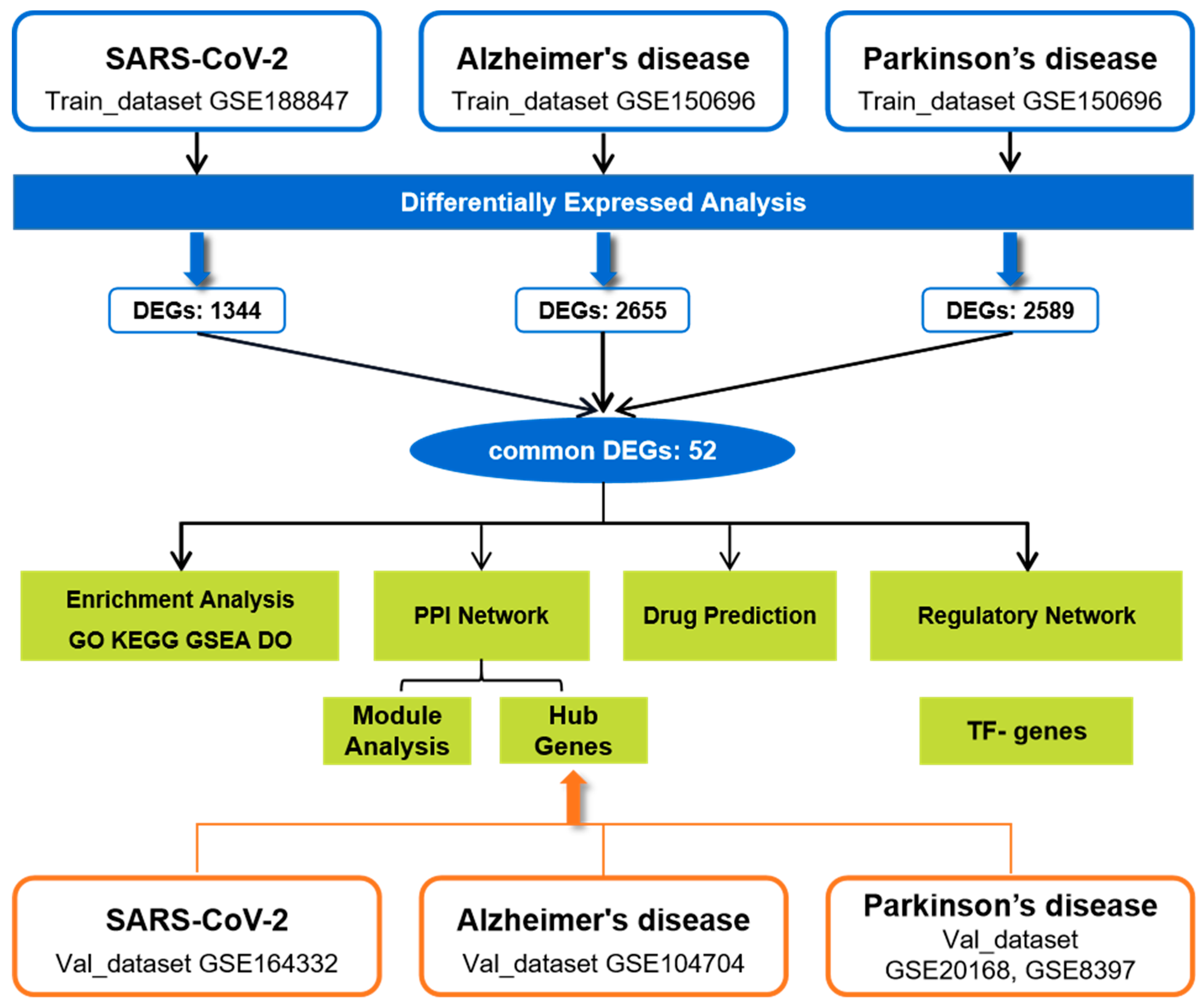
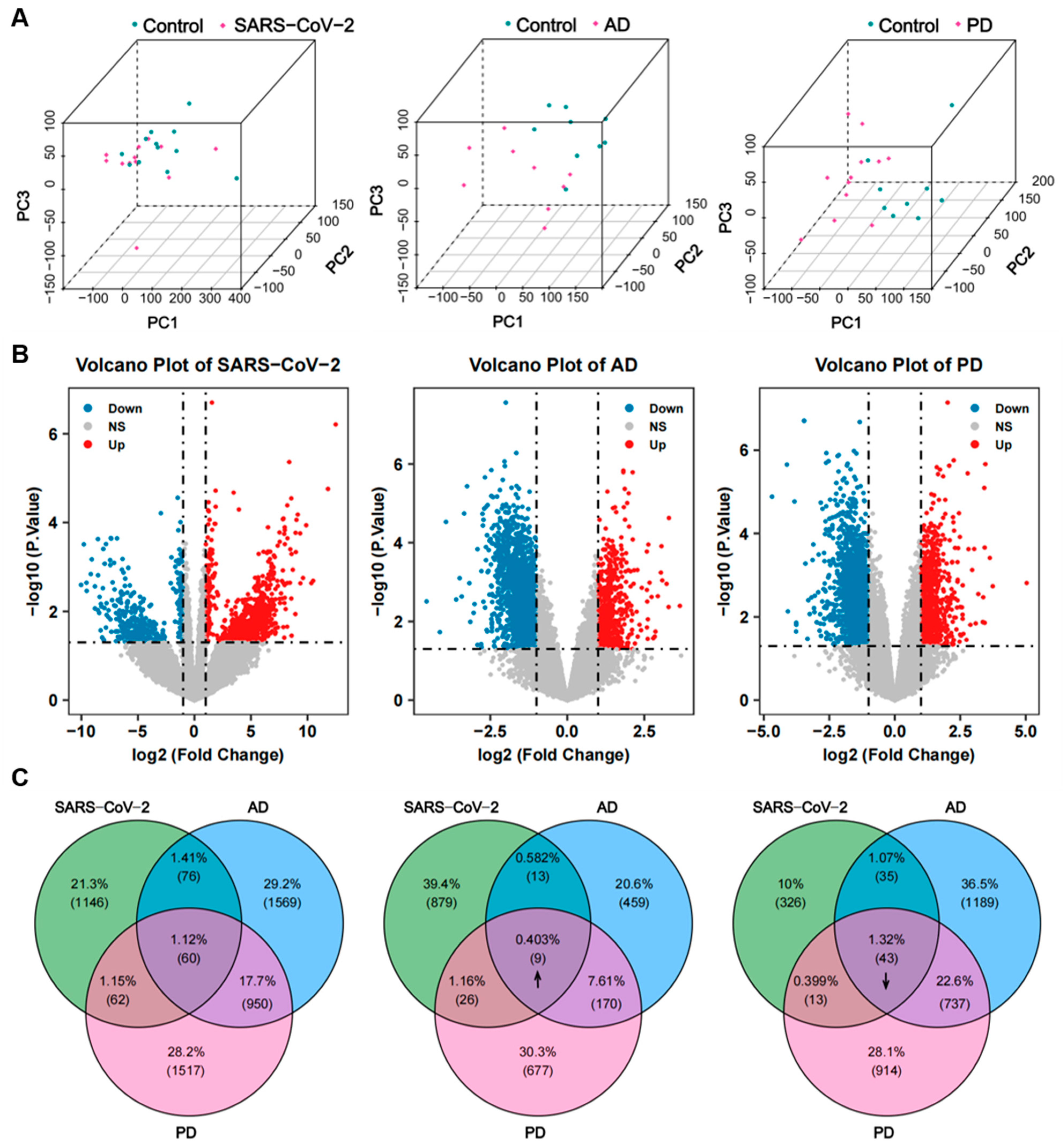
2.1. Identification of DEGs and Common DEGs among COVID-19, AD, and PD
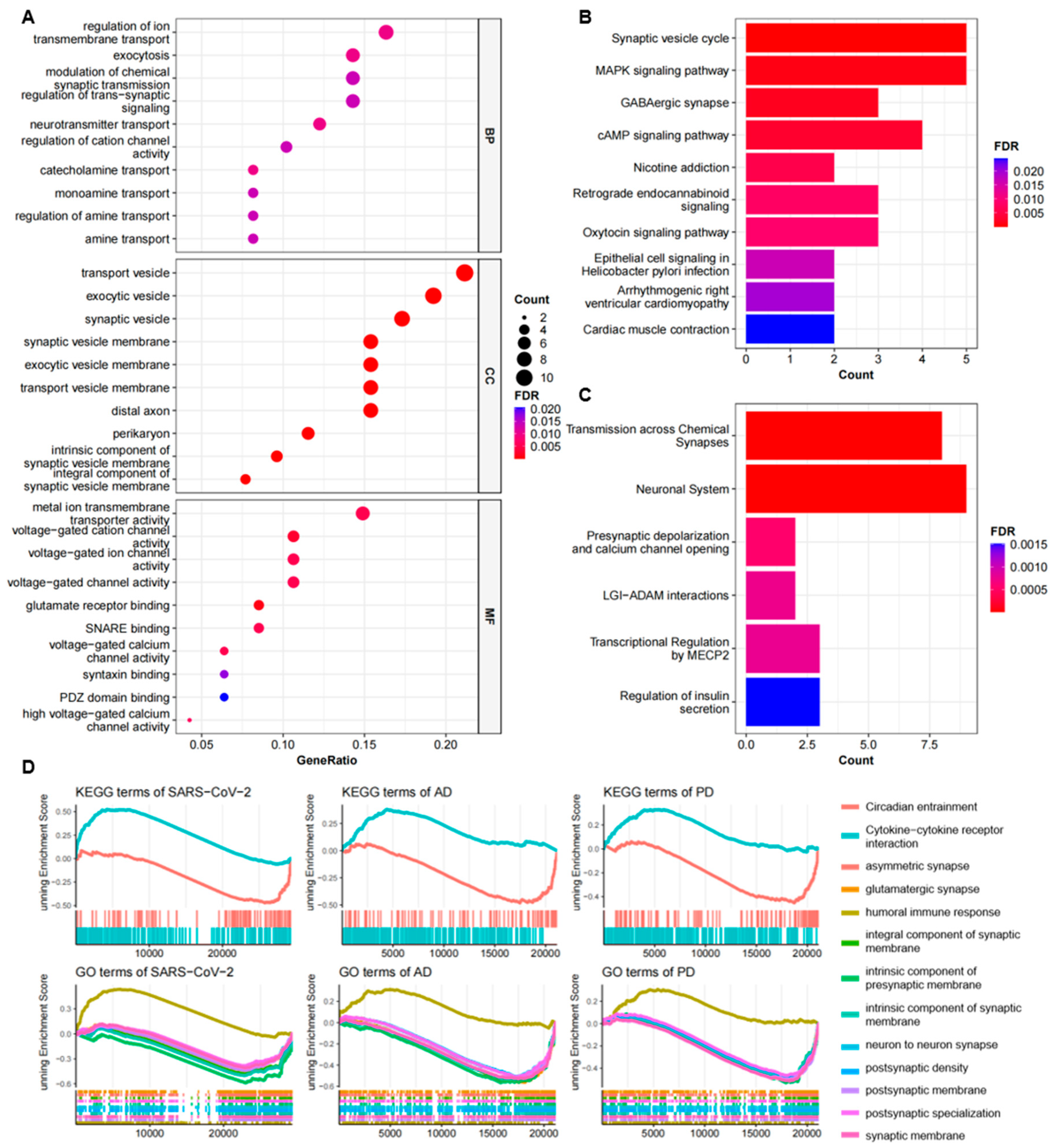
2.2. Functional Annotation and Pathway Enrichment Analysis of Common DEGs
2.3. Gene–Disease Analysis of Common DEGs
2.4. PPI Network Construction and Key Module Analysis
2.5. Identification of Hub Genes
2.6. Identification of Potential Therapeutic Drugs
2.7. Identification of Regulatory Transcript Factors
3. Discussion
4. Materials and Methods
4.1. Datasets Acquired in This Study
4.2. Identification of Common DEGs among COVID-19, AD, and PD
4.3. Functional Annotation and Pathway Enrichment of Common DEGs
4.4. Construction of PPI Network and Key Module Analysis
4.5. Detection of Hub Genes
4.6. Identification of Candidate Drugs
4.7. Prediction of Transcription Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desforges, M.; Le Coupanec, A.; Dubeau, P.; Bourgouin, A.; Lajoie, L.; Dubé, M.; Talbot, P.J. Human Coronaviruses and Other Respiratory Viruses: Underestimated Opportunistic Pathogens of the Central Nervous System? Viruses 2019, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Solomon, I.H.; Normandin, E.; Bhattacharyya, S.; Mukerji, S.S.; Keller, K.; Ali, A.S.; Adams, G.; Hornick, J.L.; Padera, R.F., Jr.; Sabeti, P. Neuropathological Features of Covid-19. N. Engl. J. Med. 2020, 383, 989–992. [Google Scholar] [CrossRef]
- Ceban, F.; Ling, S.; Lui, L.M.W.; Lee, Y.; Gill, H.; Teopiz, K.M.; Rodrigues, N.B.; Subramaniapillai, M.; Di Vincenzo, J.D.; Cao, B.; et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav. Immun. 2022, 101, 93–135. [Google Scholar] [CrossRef]
- Taquet, M.; Sillett, R.; Zhu, L.; Mendel, J.; Camplisson, I.; Dercon, Q.; Harrison, P.J. Neurological and psychiatric risk trajectories after SARS-CoV-2 infection: An analysis of 2-year retrospective cohort studies including 1 284 437 patients. Lancet Psychiatry 2022, 9, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Cavallieri, F.; Fioravanti, V.; Bove, F.; Del Prete, E.; Meoni, S.; Grisanti, S.; Zedde, M.; Pascarella, R.; Moro, E.; Valzania, F. COVID-19 and Parkinsonism: A Critical Appraisal. Biomolecules 2022, 12, 970. [Google Scholar] [CrossRef] [PubMed]
- Zarifkar, P.; Peinkhofer, C.; Benros, M.E.; Kondziella, D. Frequency of Neurological Diseases After COVID-19, Influenza A/B and Bacterial Pneumonia. Front. Neurol. 2022, 13, 904796. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, J.; Lin, J.; Shang, H. COVID-19 and risk of neurodegenerative disorders: A Mendelian randomization study. Transl. Psychiatry 2022, 12, 283. [Google Scholar] [CrossRef] [PubMed]
- Douaud, G.; Lee, S.; Alfaro-Almagro, F.; Arthofer, C.; Wang, C.; McCarthy, P.; Lange, F.; Andersson, J.L.R.; Griffanti, L.; Duff, E.; et al. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature 2022, 604, 697–707. [Google Scholar] [CrossRef]
- Delorme, C.; Paccoud, O.; Kas, A.; Hesters, A.; Bombois, S.; Shambrook, P.; Boullet, A.; Doukhi, D.; Le Guennec, L.; Godefroy, N.; et al. COVID-19-related encephalopathy: A case series with brain FDG-positron-emission tomography/computed tomography findings. Eur. J. Neurol. 2020, 27, 2651–2657. [Google Scholar] [CrossRef]
- Kovacs, G.G. Concepts and classification of neurodegenerative diseases. Handb. Clin. Neurol. 2017, 145, 301–307. [Google Scholar]
- Bak, T.H.; Chandran, S. What wires together dies together: Verbs, actions and neurodegeneration in motor neuron disease. Cortex 2012, 48, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S. Huntington’s Disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a007476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassileff, N.; Cheng, L.; Hill, A.F. Extracellular vesicles—Propagators of neuropathology and sources of potential biomarkers and therapeutics for neurodegenerative diseases. J. Cell Sci. 2020, 133, jcs243139. [Google Scholar] [CrossRef] [PubMed]
- Szeto, J.Y.; Lewis, S.J. Current Treatment Options for Alzheimer’s Disease and Parkinson’s Disease Dementia. Curr. Neuropharmacol. 2016, 14, 326–338. [Google Scholar] [CrossRef]
- Hely, M.A.; Reid, W.G.; Adena, M.A.; Halliday, G.M.; Morris, J.G. The Sydney multicenter study of Parkinson’s disease: The inevitability of dementia at 20 years. Mov. Disord. 2008, 23, 837–844. [Google Scholar] [CrossRef]
- Martínez-Cué, C.; Rueda, N. Cellular Senescence in Neurodegenerative Diseases. Front. Cell Neurosci. 2020, 14, 16. [Google Scholar] [CrossRef]
- Euston, D.R.; Gruber, A.J.; McNaughton, B.L. The role of medial prefrontal cortex in memory and decision making. Neuron 2012, 76, 1057–1070. [Google Scholar] [CrossRef] [Green Version]
- Klenowski, P.M. Emerging role for the medial prefrontal cortex in alcohol-seeking behaviors. Addict. Behav. 2018, 77, 102–106. [Google Scholar] [CrossRef]
- Zhang, Q.; Schultz, J.L.; Aldridge, G.M.; Simmering, J.E.; Narayanan, N.S. Coronavirus Disease 2019 Case Fatality and Parkinson’s Disease. Mov. Disord. 2020, 35, 1914–1915. [Google Scholar] [CrossRef]
- Zhang, Q.; Schultz, J.L.; Aldridge, G.M.; Simmering, J.E.; Kim, Y.; Ogilvie, A.C.; Narayanan, N.S. COVID-19 Case Fatality and Alzheimer’s Disease. J. Alzheimers Dis. 2021, 84, 1447–1452. [Google Scholar] [CrossRef]
- McAlpine, L.S.; Fesharaki-Zadeh, A.; Spudich, S. Coronavirus disease 2019 and neurodegenerative disease: What will the future bring? Curr. Opin. Psychiatry 2021, 34, 177–185. [Google Scholar] [CrossRef]
- Fionda, V.; Palopoli, L.; Panni, S.; Rombo, S.E. A technique to search for functional similarities in protein-protein interaction networks. Int. J. Data Min. Bioinform. 2009, 3, 431–453. [Google Scholar] [CrossRef]
- Ewing, R.M.; Chu, P.; Elisma, F.; Li, H.; Taylor, P.; Climie, S.; McBroom-Cerajewski, L.; Robinson, M.D.; O’Connor, L.; Li, M.; et al. Large-scale mapping of human protein-protein interactions by mass spectrometry. Mol. Syst. Biol. 2007, 3, 89. [Google Scholar] [CrossRef]
- Howard, P.A. Ibutilide: An antiarrhythmic agent for the treatment of atrial fibrillation or flutter. Ann. Pharmacother. 1999, 33, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Wellington, K.; Scott, L.J. Azelnidipine. Drugs 2003, 63, 2613–2621. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Hirano, T.; Okamoto, S.; Shiraishi, S.; Tomiguchi, S.; Uchino, M. Azelnidipine, a long-acting calcium channel blocker, could control hypertension without decreasing cerebral blood flow in post-ischemic stroke patients. A 123I-IMP SPECT follow-up study. Hypertens. Res. 2010, 33, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czernicki, Z.; Jurkiewicz, J.; Bojanowski, K.; Piechnik, S.; Cervos-Navarro, J. Effects of the calcium channel blockers Dotarizine and Flunarizine on cerebrovascular reactivity. J. Neurol. Sci. 1996, 143, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. Trace elements in human physiology and pathology. Copper. Biomed. Pharmacother. 2003, 57, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.J.; Chia, W.N.; Loh, C.C.; Li, Z.; Lee, Y.M.; He, Y.; Yuan, L.X.; Lim, T.K.; Liu, M.; et al. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef] [Green Version]
- Janky, R.; Verfaillie, A.; Imrichová, H.; Van de Sande, B.; Standaert, L.; Christiaens, V.; Hulselmans, G.; Herten, K.; Naval Sanchez, M.; Potier, D.; et al. iRegulon: From a gene list to a gene regulatory network using large motif and track collections. PLoS Comput. Biol. 2014, 10, e1003731. [Google Scholar] [CrossRef] [Green Version]
- Sudhof, T.C. The synaptic vesicle cycle. Annu. Rev. Neurosci. 2004, 27, 509–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.C.; Kern, F.; Losada, P.M.; Agam, M.R.; Maat, C.A.; Schmartz, G.P.; Fehlmann, T.; Stein, J.A.; Schaum, N.; Lee, D.P.; et al. Dysregulation of brain and choroid plexus cell types in severe COVID-19. Nature 2021, 595, 565–571. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Garcia, G., Jr.; Tian, E.; Cui, Q.; Chen, X.; Sun, G.; Wang, J.; Arumugaswami, V.; Shi, Y. ApoE-Isoform-Dependent SARS-CoV-2 Neurotropism and Cellular Response. Cell Stem Cell 2021, 28, 331–342.e335. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A.; et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Fu, A.K.Y.; Ip, N.Y. Synaptic dysfunction in Alzheimer’s disease: Mechanisms and therapeutic strategies. Pharmacol. Ther. 2019, 195, 186–198. [Google Scholar] [CrossRef]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Masliah, E.; Mallory, M.; Alford, M.; DeTeresa, R.; Hansen, L.A.; McKeel, D.W., Jr.; Morris, J.C. Altered expression of synaptic proteins occurs early during progression of Alzheimer’s disease. Neurology 2001, 56, 127–129. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Ana Clara, F.; Verstreken, P. Synaptic vesicle trafficking and Parkinson’s disease. Dev. Neurobiol. 2012, 72, 134–144. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Burré, J.; Sharma, M.; Tsetsenis, T.; Buchman, V.; Etherton, M.R.; Südhof, T.C. Alpha-synuclein promotes SNARE-complex assembly in vivo and in vitro. Science 2010, 329, 1663–1667. [Google Scholar] [CrossRef] [Green Version]
- Funayama, M.; Hasegawa, K.; Kowa, H.; Saito, M.; Tsuji, S.; Obata, F. A new locus for Parkinson’s disease (PARK8) maps to chromosome 12p11.2-q13.1. Ann. Neurol. 2002, 51, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Matta, S.; Van Kolen, K.; da Cunha, R.; van den Bogaart, G.; Mandemakers, W.; Miskiewicz, K.; De Bock, P.J.; Morais, V.A.; Vilain, S.; Haddad, D.; et al. LRRK2 controls an EndoA phosphorylation cycle in synaptic endocytosis. Neuron 2012, 75, 1008–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Trempe, J.F.; Chen, C.X.; Grenier, K.; Camacho, E.M.; Kozlov, G.; McPherson, P.S.; Gehring, K.; Fon, E.A. SH3 domains from a subset of BAR proteins define a Ubl-binding domain and implicate parkin in synaptic ubiquitination. Mol. Cell 2009, 36, 1034–1047. [Google Scholar] [CrossRef]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [Green Version]
- Morais, V.A.; Verstreken, P.; Roethig, A.; Smet, J.; Snellinx, A.; Vanbrabant, M.; Haddad, D.; Frezza, C.; Mandemakers, W.; Vogt-Weisenhorn, D.; et al. Parkinson’s disease mutations in PINK1 result in decreased Complex I activity and deficient synaptic function. EMBO Mol. Med. 2009, 1, 99–111. [Google Scholar] [CrossRef]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Usami, Y.; Hatano, T.; Imai, S.; Kubo, S.; Sato, S.; Saiki, S.; Fujioka, Y.; Ohba, Y.; Sato, F.; Funayama, M.; et al. DJ-1 associates with synaptic membranes. Neurobiol. Dis. 2011, 43, 651–662. [Google Scholar] [CrossRef]
- Milnerwood, A.J.; Raymond, L.A. Early synaptic pathophysiology in neurodegeneration: Insights from Huntington’s disease. Trends Neurosci. 2010, 33, 513–523. [Google Scholar] [CrossRef]
- Murphy, K.P.; Carter, R.J.; Lione, L.A.; Mangiarini, L.; Mahal, A.; Bates, G.P.; Dunnett, S.B.; Morton, A.J. Abnormal synaptic plasticity and impaired spatial cognition in mice transgenic for exon 1 of the human Huntington’s disease mutation. J. Neurosci. 2000, 20, 5115–5123. [Google Scholar] [CrossRef]
- Kanaani, J.; Lissin, D.; Kash, S.F.; Baekkeskov, S. The hydrophilic isoform of glutamate decarboxylase, GAD67, is targeted to membranes and nerve terminals independent of dimerization with the hydrophobic membrane-anchored isoform, GAD65. J. Biol. Chem. 1999, 274, 37200–37209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solas, M.; Puerta, E.; Ramirez, M.J. Treatment Options in Alzheimer’s Disease: The GABA Story. Curr. Pharm. Des. 2015, 21, 4960–4971. [Google Scholar] [CrossRef] [PubMed]
- Fontes, M.A.P.; Vaz, G.C.; Cardoso, T.Z.D.; de Oliveira, M.F.; Campagnole-Santos, M.J.; Dos Santos, R.A.S.; Sharma, N.M.; Patel, K.P.; Frézard, F. GABA-containing liposomes: Neuroscience applications and translational perspectives for targeting neurological diseases. Nanomedicine 2018, 14, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Yoon, J.; Lee, S.H. The role of neuropeptide somatostatin in the brain and its application in treating neurological disorders. Exp. Mol. Med. 2021, 53, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.; Katzman, R.; Terry, R.D. Reduced somatostatin-like immunoreactivity in cerebral cortex from cases of Alzheimer disease and Alzheimer senile dementa. Nature 1980, 288, 279–280. [Google Scholar] [CrossRef]
- Iwasawa, C.; Kuzumaki, N.; Suda, Y.; Kagawa, R.; Oka, Y.; Hattori, N.; Okano, H.; Narita, M. Reduced expression of somatostatin in GABAergic interneurons derived from induced pluripotent stem cells of patients with parkin mutations. Mol. Brain 2019, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Kremer, H.P.; Roos, R.A.; Dingjan, G.; Marani, E.; Bots, G.T. Atrophy of the hypothalamic lateral tuberal nucleus in Huntington’s disease. J. Neuropathol. Exp. Neurol. 1990, 49, 371–382. [Google Scholar] [CrossRef]
- Rubinow, D.R.; Gold, P.W.; Post, R.M.; Ballenger, J.C. CSF somatostatin in affective illness and normal volunteers. Prog. Neuropsychopharmacol. Biol. Psychiatry 1985, 9, 393–400. [Google Scholar] [CrossRef]
- Hoftman, G.D.; Volk, D.W.; Bazmi, H.H.; Li, S.; Sampson, A.R.; Lewis, D.A. Altered cortical expression of GABA-related genes in schizophrenia: Illness progression vs developmental disturbance. Schizophr. Bull. 2015, 41, 180–191. [Google Scholar] [CrossRef]
- Arnaud, L.; Benech, P.; Greetham, L.; Stephan, D.; Jimenez, A.; Jullien, N.; García-González, L.; Tsvetkov, P.O.; Devred, F.; Sancho-Martinez, I.; et al. APOE4 drives inflammation in human astrocytes via TAGLN3 repression and NF-κB activation. Cell Rep. 2022, 40, 111200. [Google Scholar] [CrossRef]
- Gincel, D.; Shoshan-Barmatz, V. The synaptic vesicle protein synaptophysin: Purification and characterization of its channel activity. Biophys. J. 2002, 83, 3223–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarsa, L.; Goda, Y. Synaptophysin regulates activity-dependent synapse formation in cultured hippocampal neurons. Proc. Natl. Acad. Sci. USA 2002, 99, 1012–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.E.; Chapman, E.R. Synaptophysin regulates the kinetics of synaptic vesicle endocytosis in central neurons. Neuron 2011, 70, 847–854. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, U.; Tanimoto, N.; Seeliger, M.; Schaeffel, F.; Leube, R.E. Detection of behavioral alterations and learning deficits in mice lacking synaptophysin. Neuroscience 2009, 162, 234–243. [Google Scholar] [CrossRef]
- Wu, X.Y.; Yu, X.Y. Overexpression of KCNJ4 correlates with cancer progression and unfavorable prognosis in lung adenocarcinoma. J. Biochem. Mol. Toxicol. 2019, 33, e22270. [Google Scholar] [CrossRef]
- Szuts, V.; Ménesi, D.; Varga-Orvos, Z.; Zvara, Á.; Houshmand, N.; Bitay, M.; Bogáts, G.; Virág, L.; Baczkó, I.; Szalontai, B.; et al. Altered expression of genes for Kir ion channels in dilated cardiomyopathy. Can. J. Physiol. Pharmacol. 2013, 91, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Kim, Y.; Choi, J.W.; Oh, H.E.; Lee, J.H. Genetic variants and risk of prostate cancer using pathway analysis of a genome-wide association study. Neoplasma 2016, 63, 629–634. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, T. Network-Based Data Analysis Reveals Ion Channel-Related Gene Features in COVID-19: A Bioinformatic Approach. Biochem. Genet. 2022, 1–35. [Google Scholar] [CrossRef]
- Wang, G.; Zeng, J.; Shen, C.Y.; Wang, Z.Q.; Chen, S.D. Overexpression of Kir2.3 in PC12 cells resists rotenone-induced neurotoxicity associated with PKC signaling pathway. Biochem. Biophys. Res. Commun. 2008, 374, 204–209. [Google Scholar] [CrossRef]
- Drożdżal, S.; Rosik, J.; Lechowicz, K.; Machaj, F.; Szostak, B.; Przybyciński, J.; Lorzadeh, S.; Kotfis, K.; Ghavami, S.; Łos, M.J. An update on drugs with therapeutic potential for SARS-CoV-2 (COVID-19) treatment. Drug Resist. Updates 2021, 59, 100794. [Google Scholar] [CrossRef]
- Niknam, Z.; Jafari, A.; Golchin, A.; Danesh Pouya, F.; Nemati, M.; Rezaei-Tavirani, M.; Rasmi, Y. Potential therapeutic options for COVID-19: An update on current evidence. Eur. J. Med. Res. 2022, 27, 6. [Google Scholar] [CrossRef] [PubMed]
- Mølhave, M.; Agergaard, J.; Wejse, C. Clinical Management of COVID-19 Patients—An Update. Semin. Nucl. Med. 2022, 52, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Molavi, Z.; Razi, S.; Mirmotalebisohi, S.A.; Adibi, A.; Sameni, M.; Karami, F.; Niazi, V.; Niknam, Z.; Aliashrafi, M.; Taheri, M.; et al. Identification of FDA approved drugs against SARS-CoV-2 RNA dependent RNA polymerase (RdRp) and 3-chymotrypsin-like protease (3CLpro), drug repurposing approach. Biomed. Pharmacother. 2021, 138, 111544. [Google Scholar] [CrossRef] [PubMed]
- Naccarelli, G.V.; Lee, K.S.; Gibson, J.K.; VanderLugt, J. Electrophysiology and pharmacology of ibutilide. Am. J. Cardiol. 1996, 78, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.H.; Wilde, M.I.; Markham, A. Ibutilide. A review of its pharmacological properties and clinical potential in the acute management of atrial flutter and fibrillation. Drugs 1997, 54, 312–330. [Google Scholar] [CrossRef] [PubMed]
- Kuridze, N.; Gajkowska, B.; Czernicki, Z.; Jurkiewicz, J.; Cervos-Navarro, J. The effect of Dotarizine--(Ca2+ channel blocker)--on vascular reactivity and ultrastructure of cerebral capillaries in animals subjected to anoxia. Folia Neuropathol. 1998, 36, 101–108. [Google Scholar]
- Nada, T.; Nomura, M.; Koshiba, K.; Kawano, T.; Mikawa, J.; Ito, S. Clinical study with azelnidipine in patients with essential hypertension. Antiarteriosclerotic and cardiac hypertrophy-inhibitory effects and influence on autonomic nervous activity. Arzneimittelforschung 2007, 57, 698–704. [Google Scholar]
- Yamashita, T.; Deguchi, K.; Sehara, Y.; Lukic-Panin, V.; Zhang, H.; Kamiya, T.; Abe, K. Therapeutic strategy for ischemic stroke. Neurochem. Res. 2009, 34, 707–710. [Google Scholar] [CrossRef]
- Omote, Y.; Deguchi, K.; Kono, S.; Liu, W.; Kurata, T.; Hishikawa, N.; Yamashita, T.; Ikeda, Y.; Abe, K. Synergistic neuroprotective effects of combined treatment with olmesartan plus azelnidipine in stroke-prone spontaneously hypertensive rats. J. Neurosci. Res. 2014, 92, 1330–1337. [Google Scholar] [CrossRef]
- Vijayan, M.; Kumar, S.; Bhatti, J.S.; Reddy, P.H. Molecular Links and Biomarkers of Stroke, Vascular Dementia, and Alzheimer’s Disease. Prog. Mol. Biol. Transl. Sci. 2017, 146, 95–126. [Google Scholar]
- Madsen, E.; Gitlin, J.D. Copper and iron disorders of the brain. Annu. Rev. Neurosci. 2007, 30, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Zucconi, G.G.; Cipriani, S.; Scattoni, R.; Balgkouranidou, I.; Hawkins, D.P.; Ragnarsdottir, K.V. Copper deficiency elicits glial and neuronal response typical of neurodegenerative disorders. Neuropathol. Appl. Neurobiol. 2007, 33, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Desai, V.; Kaler, S.G. Role of copper in human neurological disorders. Am. J. Clin. Nutr. 2008, 88, 855s–858s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatta, P.; Frank, A. Copper deficiency and neurological disorders in man and animals. Brain Res. Rev. 2007, 54, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gilmore, K.; Ramirez, S.; Settels, E.; Gammeltoft, K.A.; Pham, L.V.; Fahnøe, U.; Feng, S.; Offersgaard, A.; Trimpert, J.; et al. In vitro efficacy of artemisinin-based treatments against SARS-CoV-2. Sci. Rep. 2021, 11, 14571. [Google Scholar] [CrossRef]
- Nair, M.S.; Huang, Y.; Fidock, D.A.; Polyak, S.J.; Wagoner, J.; Towler, M.J.; Weathers, P.J. Artemisia annua L. extracts inhibit the in vitro replication of SARS-CoV-2 and two of its variants. J. Ethnopharmacol. 2021, 274, 114016. [Google Scholar] [CrossRef]
- Sehailia, M.; Chemat, S. Antimalarial-agent artemisinin and derivatives portray more potent binding to Lys353 and Lys31-binding hotspots of SARS-CoV-2 spike protein than hydroxychloroquine: Potential repurposing of artenimol for COVID-19. J. Biomol. Struct. Dyn. 2021, 39, 6184–6194. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Anjum, A.; Jaggi, S.; Varghese, E.; Lall, S.; Bhowmik, A.; Rai, A. Identification of Differentially Expressed Genes in RNA-seq Data of Arabidopsis thaliana: A Compound Distribution Approach. J. Comput. Biol. 2016, 23, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Gene Ontology Consortium. Gene Ontology Consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- Antonov, A.V.; Schmidt, E.E.; Dietmann, S.; Krestyaninova, M.; Hermjakob, H. R spider: A network-based analysis of gene lists by combining signaling and metabolic pathways from Reactome and KEGG databases. Nucleic Acids Res. 2010, 38, W78–W83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schriml, L.M.; Munro, J.B.; Schor, M.; Olley, D.; McCracken, C.; Felix, V.; Baron, J.A.; Jackson, R.; Bello, S.M.; Bearer, C.; et al. The Human Disease Ontology 2022 update. Nucleic Acids Res. 2022, 50, D1255–D1261. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [Green Version]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Aibar, S.; González-Blas, C.B.; Moerman, T.; Huynh-Thu, V.A.; Imrichova, H.; Hulselmans, G.; Rambow, F.; Marine, J.C.; Geurts, P.; Aerts, J.; et al. SCENIC: Single-cell regulatory network inference and clustering. Nat. Methods 2017, 14, 1083–1086. [Google Scholar] [CrossRef] [Green Version]
- Aerts, S.; Quan, X.J.; Claeys, A.; Naval Sanchez, M.; Tate, P.; Yan, J.; Hassan, B.A. Robust target gene discovery through transcriptome perturbations and genome-wide enhancer predictions in Drosophila uncovers a regulatory basis for sensory specification. PLoS Biol. 2010, 8, e1000435. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Sample, N (%) | COVID-19, N = 12 (50%), N (%) | Control, N = 12 (50%), N (%) | Statistics/df | p-Value | |
|---|---|---|---|---|---|
| Gender (% female) | 10 (41.67%) | 5 (41.67%) | 5 (41.67%) | 0/1 | 1.0000 |
| Age, in years, mean ± SD | 66.79 ± 10.06 | 66.7 ± 10.38 | 66.9 ± 9.72 | −0.0487/22 | 0.9616 |
| Total Sample, N (%) | AD, N = 9 (50%), N (%) | Control, N = 9 (50%), N (%) | Statistics/df | p-Value | |
|---|---|---|---|---|---|
| Gender (% female) | 10 (55.56%) | 5 (55.56%) | 5 (55.56%) | 0/1 | 1.0000 |
| Age, in years, mean ± SD | 85.17 ± 6.15 | 85.67 ± 6.36 | 84.67 ± 5.89 | 0.3461/16 | 0.7338 |
| Postmortem interval, in hours, mean ± SD | 44.11 ± 25.89 | 38.22 ± 23.30 | 50.0 ± 26.99 | −0.9911/16 | 0.3364 |
| Total Sample, N (%) | PD, N = 12 (57%), N (%) | PD, N = 9 (43%), N (%) | Statistics/df | p-Value | |
|---|---|---|---|---|---|
| Gender (% female) | 11 (52.4%) | 6 (50%) | 5 (55.56%) | 0.0636/1 | 0.8008 |
| Age, in years, mean ± SD | 80.9 ± 6.78 | 78.08 ± 5.99 | 84.67 ± 5.89 | −2.5125/19 | 0.0212 |
| Postmortem interval, in hours, mean ± SD | 40.48 ± 21.16 | 33.33 ± 10.87 | 50.0 ± 26.99 | −1.9519/19 | 0.0659 |
| Category | GO ID | Term | p-Value | Gene ID |
|---|---|---|---|---|
| BP | GO:0006836 | neurotransmitter transport | 1.64 × 10−5 | SYT5/SLC6A17/RAB3B/SYP/SLC17A7/SLC6A7 |
| GO:0006887 | exocytosis | 3.41 × 10−5 | SYT5/PAK1/RAB3B/SYP/CRHBP/NSF/SYT13 | |
| GO:0034765 | regulation of ion transmembrane transport | 3.70 × 10−5 | KCNJ4/CACNG3/CACNB1/THY1/CRHBP/RASGRF1/KCNC2/CACNA1A | |
| GO:0051937 | catecholamine transport | 3.95 × 10−5 | SYT5/RAB3B/SYT13/VIP | |
| GO:0015844 | monoamine transport | 6.85 × 10−5 | SYT5/RAB3B/SYT13/VIP | |
| GO:0051952 | regulation of amine transport | 9.36 × 10−5 | SYT5/RAB3B/SYT13/VIP | |
| GO:2001257 | regulation of cation channel activity | 0.0001 | CACNG3/CACNB1/CRHBP/RASGRF1/KCNC2 | |
| GO:0015837 | amine transport | 0.0001 | SYT5/RAB3B/SYT13/VIP | |
| GO:0050804 | modulation of chemical synaptic transmission | 0.0001 | SRGN/CACNG3/SYP/ADCYAP1/RASGRF1/NTNG1/CACNA1A | |
| GO:0099177 | regulation of trans-synaptic signaling | 0.0001 | SRGN/CACNG3/SYP/ADCYAP1/RASGRF1/NTNG1/CACNA1A | |
| CC | GO:0070382 | exocytic vesicle | 3.31 × 10−10 | SYT5/SLC6A17/RAB3B/SYP/SVOP/SLC17A7/WDR7/ GAD2/ATP6V1G2/SYT13 |
| GO:0030672 | synaptic vesicle membrane | 4.42 × 10−10 | SYT5/SLC6A17/RAB3B/SYP/SVOP/SLC17A7/GAD2/ ATP6V1G2 | |
| GO:0099501 | exocytic vesicle membrane | 4.42 × 10−10 | SYT5/SLC6A17/RAB3B/SYP/SVOP/SLC17A7/GAD2/ ATP6V1G2 | |
| GO:0008021 | synaptic vesicle | 3.34 × 10−9 | SYT5/SLC6A17/RAB3B/SYP/SVOP/SLC17A7/WDR7/ GAD2/ATP6V1G2 | |
| GO:0030133 | transport vesicle | 8.86 × 10−9 | SYT5/NRSN1/SLC6A17/RAB3B/SYP/SVOP/SLC17A7/WDR7/GAD2/ATP6V1G2/SYT13 | |
| GO:0030658 | transport vesicle membrane | 8.37 × 10−8 | SYT5/SLC6A17/RAB3B/SYP/SVOP/SLC17A7/GAD2/ ATP6V1G2 | |
| GO:0098563 | intrinsic component of synaptic vesicle membrane | 1.54 × 10−7 | SLC6A17/RAB3B/SYP/SLC17A7/ATP6V1G2 | |
| GO:0150034 | distal axon | 1.01 × 10−6 | NRSN1/SYP/THY1/CRHBP/ADCYAP1/RASGRF1/ KCNC2/OLFM1 | |
| GO:0030285 | integral component of synaptic vesicle membrane | 2.19 × 10−6 | SLC6A17/SYP/SLC17A7/ATP6V1G2 | |
| GO:0043204 | perikaryon | 3.79 × 10−6 | SERPINI1/CRHBP/ADCYAP1/KCNC2/OLFM1/VIP | |
| MF | GO:0035254 | glutamate receptor binding | 6.19 × 10−6 | CACNG3/NECAB2/NSF/RASGRF1 |
| GO:0022843 | voltage-gated cation channel activity | 3.14 × 10−5 | KCNJ4/CACNG3/CACNB1/KCNC2/CACNA1A | |
| GO:0046873 | metal ion transmembrane transporter activity | 0.0001 | KCNJ4/CACNG3/CACNB1/SLC17A7/KCNC2/ SLC6A7/CACNA1A | |
| GO:0005244 | voltage-gated ion channel activity | 0.0002 | KCNJ4/CACNG3/CACNB1/KCNC2/CACNA1A | |
| GO:0022832 | voltage-gated channel activity | 0.0002 | KCNJ4/CACNG3/CACNB1/KCNC2/CACNA1A | |
| GO:0000149 | SNARE binding | 0.0002 | SYT5/NSF/SYT13/CACNA1A | |
| GO:0005245 | voltage-gated calcium channel activity | 0.0002 | CACNG3/CACNB1/CACNA1A | |
| GO:0008331 | high voltage-gated calcium channel activity | 0.0003 | CACNB1/CACNA1A | |
| GO:0019905 | syntaxin binding | 0.0009 | SYT5/NSF/CACNA1A | |
| GO:0030165 | PDZ domain binding | 0.0013 | KCNJ4/CACNG3/NSF |
| Category | ID | Pathway | p-Value | Gene ID |
|---|---|---|---|---|
| KEGG | hsa04721 | Synaptic vesicle cycle | 2.06 × 10−6 | NSF/SLC17A7/ATP6V1G2/SLC6A7/ CACNA1A |
| hsa04010 | MAPK signaling pathway | 0.0011 | PAK1/CACNG3/CACNB1/RASGRF1/ CACNA1A | |
| hsa04727 | GABAergic synapse | 0.0019 | NSF/GAD2/CACNA1A | |
| hsa04024 | cAMP signaling pathway | 0.0031 | PAK1/SST/ADCYAP1/VIP | |
| hsa05033 | Nicotine addiction | 0.0056 | SLC17A7/CACNA1A | |
| hsa04723 | Retrograde endocannabinoid signaling | 0.0079 | NDUFV2/SLC17A7/CACNA1A | |
| hsa04921 | Oxytocin signaling pathway | 0.0089 | KCNJ4/CACNG3/CACNB1 | |
| hsa05120 | Epithelial cell signaling in Helicobacter pylori infection | 0.0163 | PAK1/ATP6V1G2 | |
| hsa05412 | Arrhythmogenic right ventricular cardiomyopathy | 0.0196 | CACNG3/CACNB1 | |
| hsa04260 | Cardiac muscle contraction | 0.0246 | CACNG3/CACNB1 | |
| Reactome | R-HSA-112315 | Transmission across chemical synapses | 8.21 × 10−7 | KCNJ4/CACNG3/CACNB1/NSF/SLC17A7/ RASGRF1/GAD2/CACNA1A |
| R-HSA-112316 | Neuronal system | 1.84 × 10−6 | KCNJ4/CACNG3/CACNB1/NSF/SLC17A7/ RASGRF1/KCNC2/GAD2/CACNA1A | |
| R-HSA-112308 | Presynaptic depolarization and calcium channel opening | 0.0005 | CACNB1/CACNA1A | |
| R-HSA-5682910 | LGI-ADAM interactions | 0.0007 | CACNG3/LGI2 | |
| R-HSA-8986944 | Transcriptional regulation by MECP2 | 0.0008 | FKBP5/SST/GAD2 | |
| R-HSA-422356 | Regulation of insulin secretion | 0.0015 | SYT5/KCNC2/CACNA1A | |
| R-HSA-373080 | Class B/2 (secretin family receptors) | 0.0027 | CRHBP/ADCYAP1/VIP | |
| R-HSA-112314 | Neurotransmitter receptors and postsynaptic signal transmission | 0.0029 | KCNJ4/CACNG3/NSF/RASGRF1 | |
| R-HSA-399719 | Trafficking of AMPA receptors | 0.0037 | CACNG3/NSF | |
| R-HSA-399721 | Glutamate binding, activation of AMPA receptors, and synaptic plasticity | 0.0037 | CACNG3/NSF |
| Drug | Type | p-Value | Target | Expression | Structure/Formula | Indication |
|---|---|---|---|---|---|---|
| Ibutilide | activator | 0.0058 | CACNB1 | Down | C20H36N2O3S  | Indicated for the rapid conversion of atrial fibrillation or atrial flutter of recent onset to sinus rhythm. |
| Azelnidipine | agonist | 0.0058 | CACNB1 | Down | C33H34N4O6  | For the treatment of hypertension. |
| Dotarizine | unknow | 0.0068 | CACNA1A | Down | C29H34N2O2  | Investigated for use/treatment in migraine and cluster headaches. |
| Copper | unknow | 0.0233 | SFPQ | Up | Cu | For use in the supplementation of total parenteral nutrition and in contraception with intrauterine devices. |
| Artenimol | ligand | 0.0233 | SFPQ | Up | C15H24O5  | For the treatment of uncomplicated Plasmodium falciparum infection in adults, children, and infants aged 6 months and up and weighing over 5 kg. Used in combination with Piperaquine. |
| Disease | GEO Accession | Tissue Source | Data | DEG Count | GEO GPL | Assay Type | ||
|---|---|---|---|---|---|---|---|---|
| Case | Control | Up | Down | |||||
| COVID-19 Train_dataset | GSE188847 | frontal cortex | 12 | 12 | 927 | 417 | GPL24676 | RNA-Seq |
| AD Train_dataset | GSE150696 | frontal cortex | 9 | 9 | 651 | 2004 | GPL17585 | Array |
| PD Train_dataset | GSE150696 | frontal cortex | 12 | 9 | 882 | 1707 | GPL17585 | Array |
| COVID-19 Val_dataset | GSE164332 | frontal cortex | 9 | 7 | 658 | 349 | GPL18573 | RNA-Seq |
| AD Val_dataset | GSE104704 | lateral temporal lobe | 12 | 18 | 683 | 2114 | GPL18573 | RNA-Seq |
| PD Val_dataset | GSE8397 | frontal cortex | 5 | 3 | 552 | 676 | GPL96 | Array |
| GSE20168 | frontal cortex | 14 | 15 | 156 | 667 | GPL96 | Array | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Liu, W.; Yang, Y.; Ci, Y.; Shi, L. Exploration of the Shared Molecular Mechanisms between COVID-19 and Neurodegenerative Diseases through Bioinformatic Analysis. Int. J. Mol. Sci. 2023, 24, 4839. https://doi.org/10.3390/ijms24054839
Shi Y, Liu W, Yang Y, Ci Y, Shi L. Exploration of the Shared Molecular Mechanisms between COVID-19 and Neurodegenerative Diseases through Bioinformatic Analysis. International Journal of Molecular Sciences. 2023; 24(5):4839. https://doi.org/10.3390/ijms24054839
Chicago/Turabian StyleShi, Yingchao, Wenhao Liu, Yang Yang, Yali Ci, and Lei Shi. 2023. "Exploration of the Shared Molecular Mechanisms between COVID-19 and Neurodegenerative Diseases through Bioinformatic Analysis" International Journal of Molecular Sciences 24, no. 5: 4839. https://doi.org/10.3390/ijms24054839
APA StyleShi, Y., Liu, W., Yang, Y., Ci, Y., & Shi, L. (2023). Exploration of the Shared Molecular Mechanisms between COVID-19 and Neurodegenerative Diseases through Bioinformatic Analysis. International Journal of Molecular Sciences, 24(5), 4839. https://doi.org/10.3390/ijms24054839