
Differential Modulation of Dorsal Raphe Serotonergic Activity in Rat Brain by the Infralimbic and Prelimbic Cortices

 , , and
, , and
Abstract
:
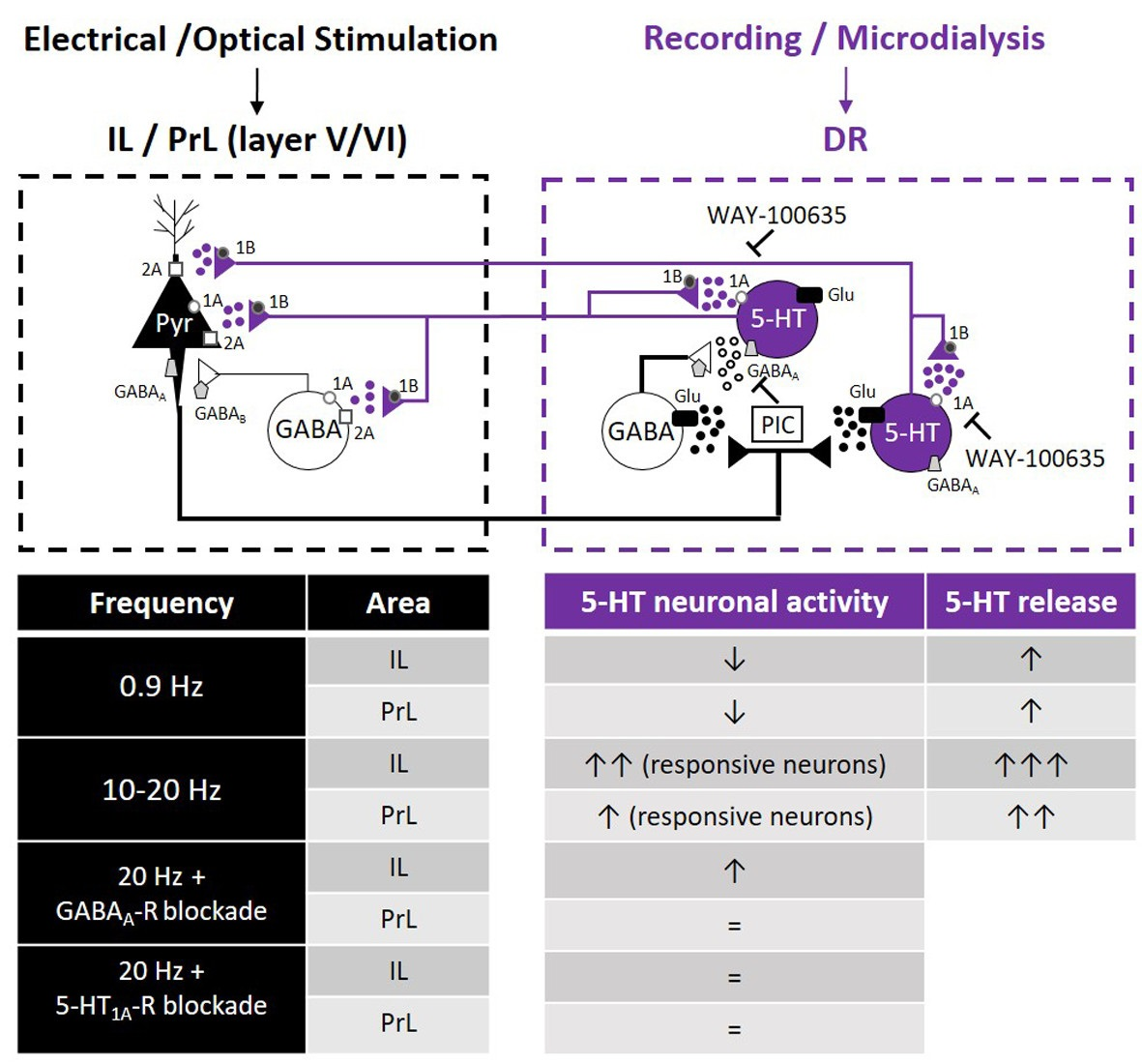{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Modulation of 5-HT Neuronal Responses by Low Frequency Electrical Stimulation of IL and PrL
2.2. Modulation of 5-HT Neuronal Discharge by Electrical Stimulation of IL and PrL
2.3. Pharmacological Characterization of IL- and PrL-Induced Responses in DR 5-HT Neurons: Involvement of 5-HT1A and GABAA Receptors
2.4. Modulation of 5-HT Release in DR by Electrical Stimulation of IL and PrL
2.5. Modulation of 5-HT Release in DR by Optogenetic Stimulation of IL and PrL
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Stereotaxic Surgery
4.4. In Vivo Electrophysiology
Electrophysiology Data Analysis
4.5. In Vivo Microdialysis during Electrical or Optogenetic Stimulation
4.5.1. Electrical Stimulation
4.5.2. Optogenetic Stimulation
4.6. Immunohistochemistry
Image Acquisition and Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, E.K. The prefontral cortex and cognitive control. Nat. Rev. Neurosci. 2000, 1, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Fuster, J. The Prefrontal Cortex, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 9780123736444. [Google Scholar]
- Fuster, J.M. The Prefrontal Cortex—An Update: Time Is of the Essence. Neuron 2001, 30, 319–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.K.; Cohen, J.D. An Integrative Theory of Prefrontal Cortex Function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, M.; Shohamy, D.; Wager, T.D. Ventromedial prefrontal-subcortical systems and the generation of affective meaning. Trends Cogn. Sci. 2012, 16, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Delgado, M.R.; Beer, J.S.; Fellows, L.K.; Huettel, S.A.; Platt, M.L.; Quirk, G.J.; Schiller, D. Viewpoints: Dialogues on the functional role of the ventromedial prefrontal cortex. Nat. Neurosci. 2016, 19, 1545–1552. [Google Scholar] [CrossRef]
- Hiser, J.; Koenigs, M. The Multifaceted Role of theVentromedial Prefrontal Cortex in Emotion, Decision Making, Social Cognition, and Psychopathology. Biol. Psychiatry 2018, 83, 638–647. [Google Scholar] [CrossRef]
- Alexander, L.; Clarke, H.; Roberts, A. A Focus on the Functions of Area 25. Brain Sci. 2019, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Andrewes, D.G.; Jenkins, L.M. The Role of the Amygdala and the Ventromedial Prefrontal Cortex in Emotional Regulation: Implications for Post-traumatic Stress Disorder. Neuropsychol. Rev. 2019, 29, 220–243. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Uylings, H.B. The prefrontal cortex and the integration of sensory, limbic and autonomic information. Prog. Brain Res. 2000, 126, 3–28. [Google Scholar] [CrossRef]
- Celada, P.; Puig, M.V.; Casanovas, J.M.; Guillazo, G.; Artigas, F. Control of Dorsal Raphe Serotonergic Neurons by the Medial Prefrontal Cortex: Involvement of Serotonin-1A, GABAA, and Glutamate Receptors. J. Neurosci. 2001, 21, 9917–9929. [Google Scholar] [CrossRef] [Green Version]
- Maren, S.; Quirk, G.J. Neuronal signalling of fear memory. Nat. Rev. Neurosci. 2004, 5, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Vertes, R.P. Differential projections of the infralimbic and prelimbic cortex in the rat. Synapse 2003, 51, 32–58. [Google Scholar] [CrossRef] [PubMed]
- Gabbott, P.L.; Warner, T.A.; Jays, P.R.; Salway, P.; Busby, S.J. Prefrontal cortex in the rat: Projections to subcortical autonomic, motor, and limbic centers. J. Comp. Neurol. 2005, 492, 145–177. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.V.; Artigas, F.; Celada, P. Modulation of the Activity of Pyramidal Neurons in Rat Prefrontal Cortex by Raphe Stimulation In Vivo: Involvement of Serotonin and GABA. Cereb. Cortex 2004, 15, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Blair, R.J.R. The amygdala and ventromedial prefrontal cortex in morality and psychopathy. Trends Cogn. Sci. 2007, 11, 387–392. [Google Scholar] [CrossRef]
- Sierra-Mercado, D.; Padilla-Coreano, N.; Quirk, G.J. Dissociable Roles of Prelimbic and Infralimbic Cortices, Ventral Hippocampus, and Basolateral Amygdala in the Expression and Extinction of Conditioned Fear. Neuropsychopharmacology 2010, 36, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Lingawi, N.W.; Laurent, V.; Westbrook, R.F.; Holmes, N.M. The role of the basolateral amygdala and infralimbic cortex in (re)learning extinction. Psychopharmacology 2018, 236, 303–312. [Google Scholar] [CrossRef]
- López-Terrones, E.; Celada, P.; Riga, M.S.; Artigas, F. Preferential in vivo inhibitory action of serotonin in rat infralimbic versus prelimbic cortex: Relevance for antidepressant treatments. Cereb. Cortex 2021, 32, 3000–3013. [Google Scholar] [CrossRef]
- Warden, M.R.; Selimbeyoglu, A.; Mirzabekov, J.J.; Lo, M.; Thompson, K.R.; Kim, S.-Y.; Adhikari, A.; Tye, K.M.; Frank, L.M.; Deisseroth, K. A prefrontal cortex–brainstem neuronal projection that controls response to behavioural challenge. Nature 2012, 492, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Burgos-Robles, A.; Vidal-Gonzalez, I.; Santini, E.; Quirk, G.J. Consolidation of Fear Extinction Requires NMDA Receptor-Dependent Bursting in the Ventromedial Prefrontal Cortex. Neuron 2007, 53, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Myers-Schulz, B.; Koenigs, M. Functional Anatomy of Ventromedial Prefrontal Cortex: Implications for Mood and Anxiety Disorders. Mol. Psychiatry 2012, 17, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Mayberg, H.S. Targeted electrode-based modulation of neural circuits for depression. J. Clin. Investig. 2009, 119, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Savitz, J.; Drevets, W.C. Bipolar and major depressive disorder: Neuroimaging the developmental-degenerative divide. Neurosci. Biobehav. Rev. 2009, 33, 699–771. [Google Scholar] [CrossRef] [Green Version]
- Mayberg, H.S.; Lozano, A.M.; Voon, V.; McNeely, H.E.; Seminowicz, D.; Hamani, C.; Schwalb, J.M.; Kennedy, S. Deep Brain Stimulation for Treatment-Resistant Depression. Neuron 2005, 45, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Puigdemont, D.; Pérez-Egea, R.; Portella, M.J.; Molet, J.; de Diego-Adeliño, J.; Gironell, A.; Radua, J.; Gómez-Anson, B.; Rodríguez, R.; Serra, M.; et al. Deep brain stimulation of the subcallosal cingulate gyrus: Further evidence in treatment-resistant major depression. Int. J. Neuropsychopharmacol. 2011, 15, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Holtzheimer, P.E.; Husain, M.M.; Lisanby, S.H.; Taylor, S.F.; Whitworth, L.A.; McClintock, S.; Slavin, K.V.; Berman, J.; McKhann, G.M.; Patil, P.G.; et al. Subcallosal cingulate deep brain stimulation for treatment-resistant depression: A multisite, randomised, sham-controlled trial. Lancet Psychiatry 2017, 4, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, L.S.; Costi, S.; Tan, A.; Stern, E.R.; Charney, D.S.; Murrough, J.W. Ketamine normalizes subgenual cingulate cortex hyper-activity in depression. Neuropsychopharmacology 2020, 45, 975–981. [Google Scholar] [CrossRef]
- Quirk, G.J.; Beer, J.S. Prefrontal involvement in the regulation of emotion: Convergence of rat and human studies. Curr. Opin. Neurobiol. 2006, 16, 723–727. [Google Scholar] [CrossRef]
- Heilbronner, S.R.; Rodriguez-Romaguera, J.; Quirk, G.J.; Groenewegen, H.J.; Haber, S.N. Circuit-Based Corticostriatal Homologies Between Rat and Primate. Biol. Psychiatry 2016, 80, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Fuchikami, M.; Thomas, A.; Liu, R.; Wohleb, E.S.; Land, B.B.; DiLeone, R.J.; Aghajanian, G.K.; Duman, R.S. Optogenetic stimulation of infralimbic PFC reproduces ketamine’s rapid and sustained antidepressant actions. Proc. Natl. Acad. Sci. USA 2015, 112, 8106–8111. [Google Scholar] [CrossRef] [Green Version]
- Gasull-Camós, J.; Martínez-Torres, S.; Tarrés-Gatius, M.; Ozaita, A.; Artigas, F.; Castañé, A. Serotonergic mechanisms involved in antidepressant-like responses evoked by GLT-1 blockade in rat infralimbic cortex. Neuropharmacology 2018, 139, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasull-Camós, J.; Soto-Montenegro, M.L.; Casquero-Veiga, M.; Desco, M.; Artigas, F.; Castañé, A. Differential Patterns of Subcortical Activity Evoked by Glial GLT-1 Blockade in Prelimbic and Infralimbic Cortex: Relationship to Antidepressant-Like Effects in Rats. Int. J. Neuropsychopharmacol. 2017, 20, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fullana, M.N.; Ruiz-Bronchal, E.; Ferrés-Coy, A.; Juárez-Escoto, E.; Artigas, F.; Bortolozzi, A. Regionally selective knockdown of astroglial glutamate transporters in infralimbic cortex induces a depressive phenotype in mice. Glia 2019, 67, 1122–1137. [Google Scholar] [CrossRef] [PubMed]
- Kargieman, L.; Santana, N.; Mengod, G.; Celada, P.; Artigas, F. Antipsychotic drugs reverse the disruption in prefrontal cortex function produced by NMDA receptor blockade with phencyclidine. Proc. Natl. Acad. Sci. USA 2007, 104, 14843–14848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puig, M.V.; Celada, P.; Diaz-Mataix, L.; Artigas, F. In vivo modulation of the activity of pyramidal neurons in the rat medial prefrontal cortex by 5-HT2A receptors: Relationship to thalamocortical afferents. Cereb. Cortex 2003, 13, 870–882. [Google Scholar] [CrossRef] [Green Version]
- Varga, V.; Székely, A.; Csillag, A.; Sharp, T.; Hajós, M. Evidence for a role of GABA interneurones in the cortical modulation of midbrain 5-hydroxytryptamine neurones. Neuroscience 2001, 106, 783–792. [Google Scholar] [CrossRef]
- Sprouse, J.S.; Aghajanian, G.K. Electrophysiological responses of serotoninergic dorsal raphe neurons to 5-HT1A and 5-HT1B agonists. Synapse 1987, 1, 3–9. [Google Scholar] [CrossRef]
- Cathala, A.; Lucas, G.; López-Terrones, E.; Revest, J.-M.; Artigas, F.; Spampinato, U. Differential expression of serotonin2B receptors in GABAergic and serotoninergic neurons of the rat and mouse dorsal raphe nucleus. Mol. Cell. Neurosci. 2022, 121, 103750. [Google Scholar] [CrossRef]
- Boothman, L.; Raley, J.; Denk, F.; Hirani, E.; Sharp, T. In vivo evidence that 5-HT2C receptors inhibit 5-HT neuronal activity via a GABAergic mechanism. Br. J. Pharmacol. 2006, 149, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Gartside, S.E.; Cole, A.J.; Williams, A.P.; McQuade, R.; Judge, S.J. AMPA and NMDA receptor regulation of firing activity in 5-HT neurons of the dorsal and median raphe nuclei. Eur. J. Neurosci. 2007, 25, 3001–3008. [Google Scholar] [CrossRef]
- Hajós, M.; Richards, C.; Székely, A.; Sharp, T. An electrophysiological and neuroanatomical study of the medial prefrontal cortical projection to the midbrain raphe nuclei in the rat. Neuroscience 1998, 87, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Srejic, L.R.; Hamani, C.; Hutchison, W.D. High-frequency stimulation of the medial prefrontal cortex decreases cellular firing in the dorsal raphe. Eur. J. Neurosci. 2015, 41, 1219–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoover, W.B.; Vertes, R.P. Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Anat. Embryol. 2007, 212, 149–179. [Google Scholar] [CrossRef]
- Ji, G.; Neugebauer, V. Modulation of medial prefrontal cortical activity using in vivo recordings and optogenetics. Mol. Brain 2012, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Aerde, K.I.; Heistek, T.S.; Mansvelder, H.D. Prelimbic and Infralimbic Prefrontal Cortex Interact during Fast Network Oscillations. PLoS ONE 2008, 3, e2725. [Google Scholar] [CrossRef] [Green Version]
- Marek, R.; Xu, L.; Sullivan, R.K.P.; Sah, P. Excitatory connections between the prelimbic and infralimbic medial prefrontal cortex show a role for the prelimbic cortex in fear extinction. Nat. Neurosci. 2018, 21, 654–658. [Google Scholar] [CrossRef]
- Mukherjee, A.; Caroni, P. Infralimbic cortex is required for learning alternatives to prelimbic promoted associations through reciprocal connectivity. Nat. Commun. 2018, 9, 2727. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Uematsu, A.; Johansen, J.P. Enhanced synchronization between prelimbic and infralimbic cortices during fear extinction learning. Mol. Brain 2021, 14, 175. [Google Scholar] [CrossRef]
- Srejic, L.R.; Wood, K.M.; Zeqja, A.; Hashemi, P.; Hutchison, W.D. Modulation of serotonin dynamics in the dorsal raphe nucleus via high frequency medial prefrontal cortex stimulation. Neurobiol. Dis. 2016, 94, 129–138. [Google Scholar] [CrossRef]
- Levine, E.S.; Jacobs, B.L. Microiontophoresis and single-unit recordings of serotonergic neurons in the awake cat. J. Chem. Neuroanat. 1992, 5, 335–337. [Google Scholar] [CrossRef]
- Bagdy, E.; Király, I.; Harsing, J.L.G. Reciprocal Innervation between Serotonergic and GABAergic Neurons in Raphe Nuclei of the Rat. Neurochem. Res. 2000, 25, 1465–1473. [Google Scholar] [CrossRef]
- Sharp, T.; McQuade, R.; Bramwell, S.; Hjorth, S. Effect of acute and repeated administration of 5-HT1A receptor agonists on 5-HT release in rat brain in vivo. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1993, 348, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Artigas, F.; Celada, P.; Laruelle, M.; Adell, A. How does pindolol improve antidepressant action? Trends Pharmacol. Sci. 2001, 22, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Sánchez, L.; Castañé, A.; Pérez-Caballero, L.; Grifoll-Escoda, M.; López-Gil, X.; Campa, L.; Galofré, M.; Berrocoso, E.; Adell, A. Activation of AMPA Receptors Mediates the Antidepressant Action of Deep Brain Stimulation of the Infralimbic Prefrontal Cortex. Cereb. Cortex 2015, 26, 2778–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fullana, M.N.; Covelo, A.; Bortolozzi, A.; Araque, A.; Artigas, F. In vivo knockdown of astroglial glutamate transporters GLT-1 and GLAST increases excitatory neurotransmission in mouse infralimbic cortex: Relevance for depressive-like phenotypes. Eur. Neuropsychopharmacol. 2019, 29, 1288–1294. [Google Scholar] [CrossRef] [PubMed]
- Fullana, N.; Gasull-Camós, J.; Tarrés-Gatius, M.; Castañé, A.; Bortolozzi, A.; Artigas, F. Astrocyte control of glutamatergic activity: Downstream effects on serotonergic function and emotional behavior. Neuropharmacology 2019, 166, 107914. [Google Scholar] [CrossRef] [PubMed]
- Garro-Martínez, E.; Fullana, M.N.; Florensa-Zanuy, E.; Senserrich, J.; Paz, V.; Ruiz-Bronchal, E.; Adell, A.; Castro, E.; Díaz, Á.; Pazos, Á.; et al. mTOR Knockdown in the Infralimbic Cortex Evokes A Depressive-like State in Mouse. Int. J. Mol. Sci. 2021, 22, 8671. [Google Scholar] [CrossRef]
- Lim, L.W.; Prickaerts, J.; Huguet, G.; Kadar, E.; Hartung, H.; Sharp, T.; Temel, Y. Electrical stimulation alleviates depressive-like behaviors of rats: Investigation of brain targets and potential mechanisms. Transl. Psychiatry 2015, 5, e535. [Google Scholar] [CrossRef] [Green Version]
- Haj-Dahmane, S.; Béïque, J.C.; Shen, R.-Y. GluA2-Lacking AMPA Receptors and Nitric Oxide Signaling Gate Spike-Timing–Dependent Potentiation of Glutamate Synapses in the Dorsal Raphe Nucleus. Eneuro 2017, 4, ENEURO.0116-17. [Google Scholar] [CrossRef] [Green Version]
- Weissbourd, B.; Ren, J.; DeLoach, K.E.; Guenthner, C.J.; Miyamichi, K.; Luo, L. Presynaptic Partners of Dorsal Raphe Serotonergic and GABAergic Neurons. Neuron 2014, 83, 645–662. [Google Scholar] [CrossRef] [Green Version]
- Allers, K.; Sharp, T. Neurochemical and anatomical identification of fast- and slow-firing neurons in the rat dorsal raphe nucleus using juxtacellular labelling methods in vivo. Neuroscience 2003, 122, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Terrones, E.; Paz, V.; Campa, L.; Conde-Berriozabal, S.; Masana, M.; Artigas, F.; Riga, M.S. Differential Modulation of Dorsal Raphe Serotonergic Activity in Rat Brain by the Infralimbic and Prelimbic Cortices. Int. J. Mol. Sci. 2023, 24, 4891. https://doi.org/10.3390/ijms24054891
López-Terrones E, Paz V, Campa L, Conde-Berriozabal S, Masana M, Artigas F, Riga MS. Differential Modulation of Dorsal Raphe Serotonergic Activity in Rat Brain by the Infralimbic and Prelimbic Cortices. International Journal of Molecular Sciences. 2023; 24(5):4891. https://doi.org/10.3390/ijms24054891
Chicago/Turabian StyleLópez-Terrones, Elena, Verónica Paz, Leticia Campa, Sara Conde-Berriozabal, Mercè Masana, Francesc Artigas, and Maurizio S. Riga. 2023. "Differential Modulation of Dorsal Raphe Serotonergic Activity in Rat Brain by the Infralimbic and Prelimbic Cortices" International Journal of Molecular Sciences 24, no. 5: 4891. https://doi.org/10.3390/ijms24054891
APA StyleLópez-Terrones, E., Paz, V., Campa, L., Conde-Berriozabal, S., Masana, M., Artigas, F., & Riga, M. S. (2023). Differential Modulation of Dorsal Raphe Serotonergic Activity in Rat Brain by the Infralimbic and Prelimbic Cortices. International Journal of Molecular Sciences, 24(5), 4891. https://doi.org/10.3390/ijms24054891