

Histaminergic System and Inflammation-Related Genes in Normal Large Intestine and Adenocarcinoma Tissues: Transcriptional Profiles and Relations

Abstract
:1. Introduction
2. Results
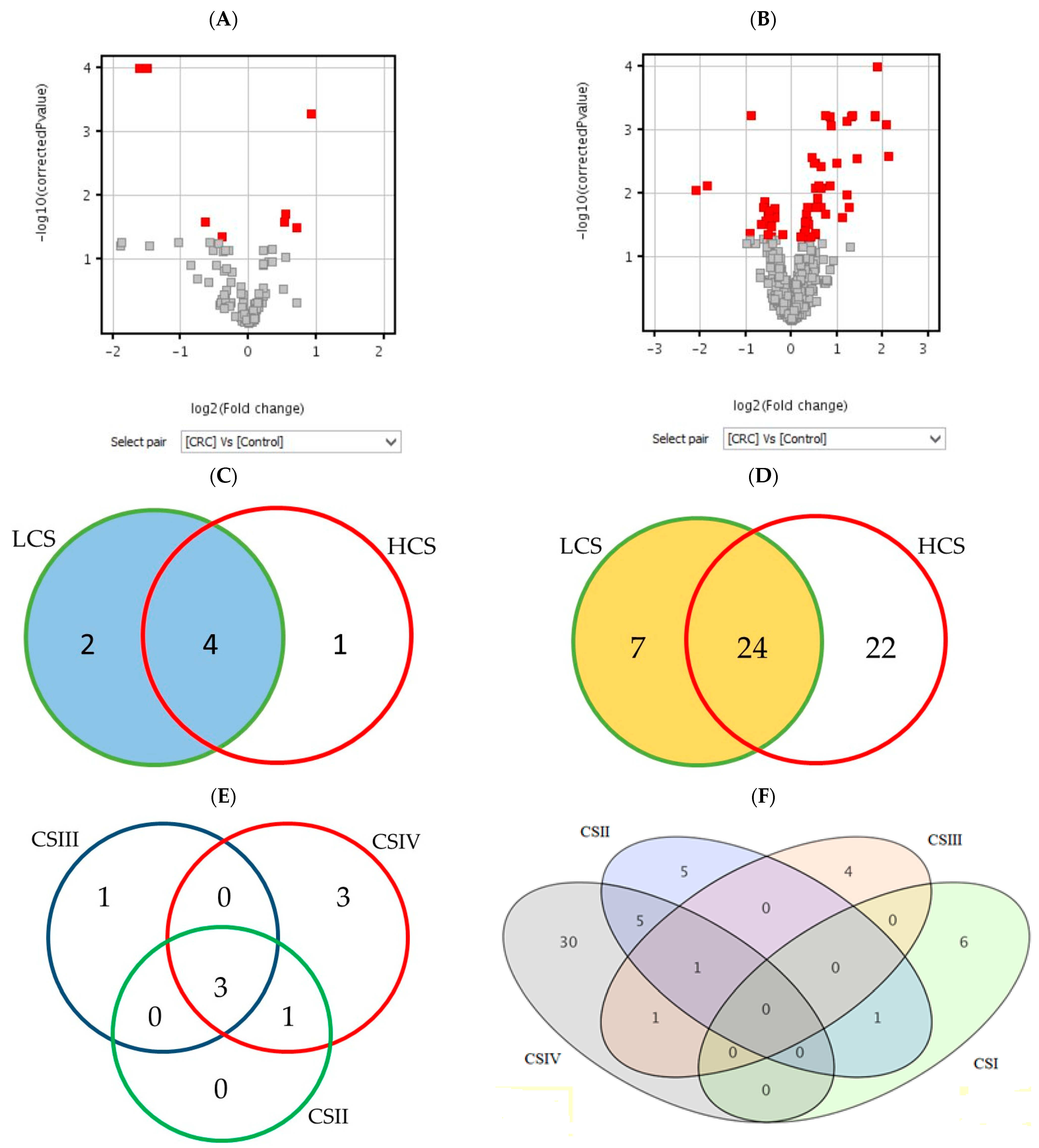
2.1. Microarray Data Analysis
2.1.1. Analysis of Differentially Expressed Genes
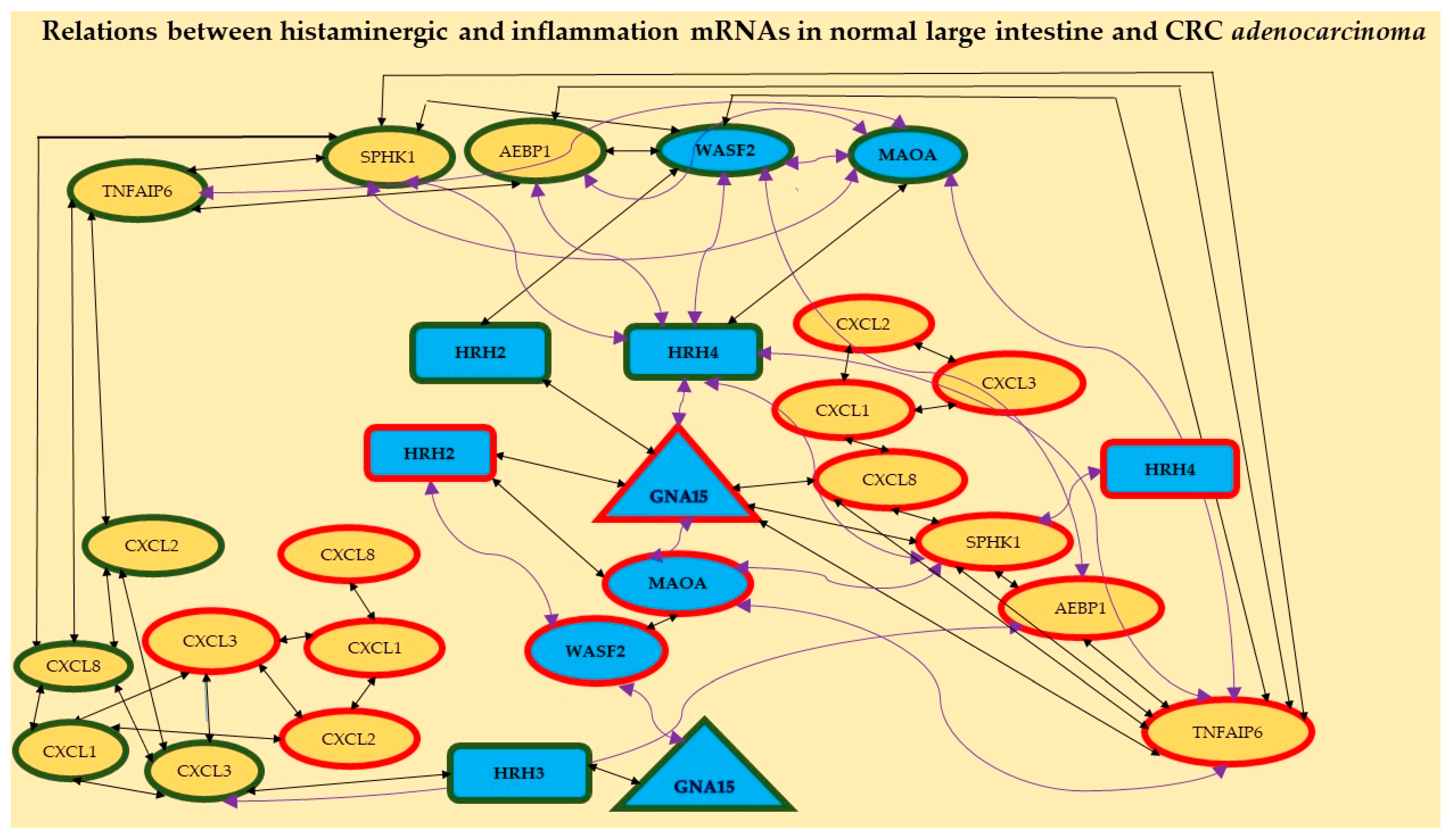
2.1.2. Analysis of Correlation
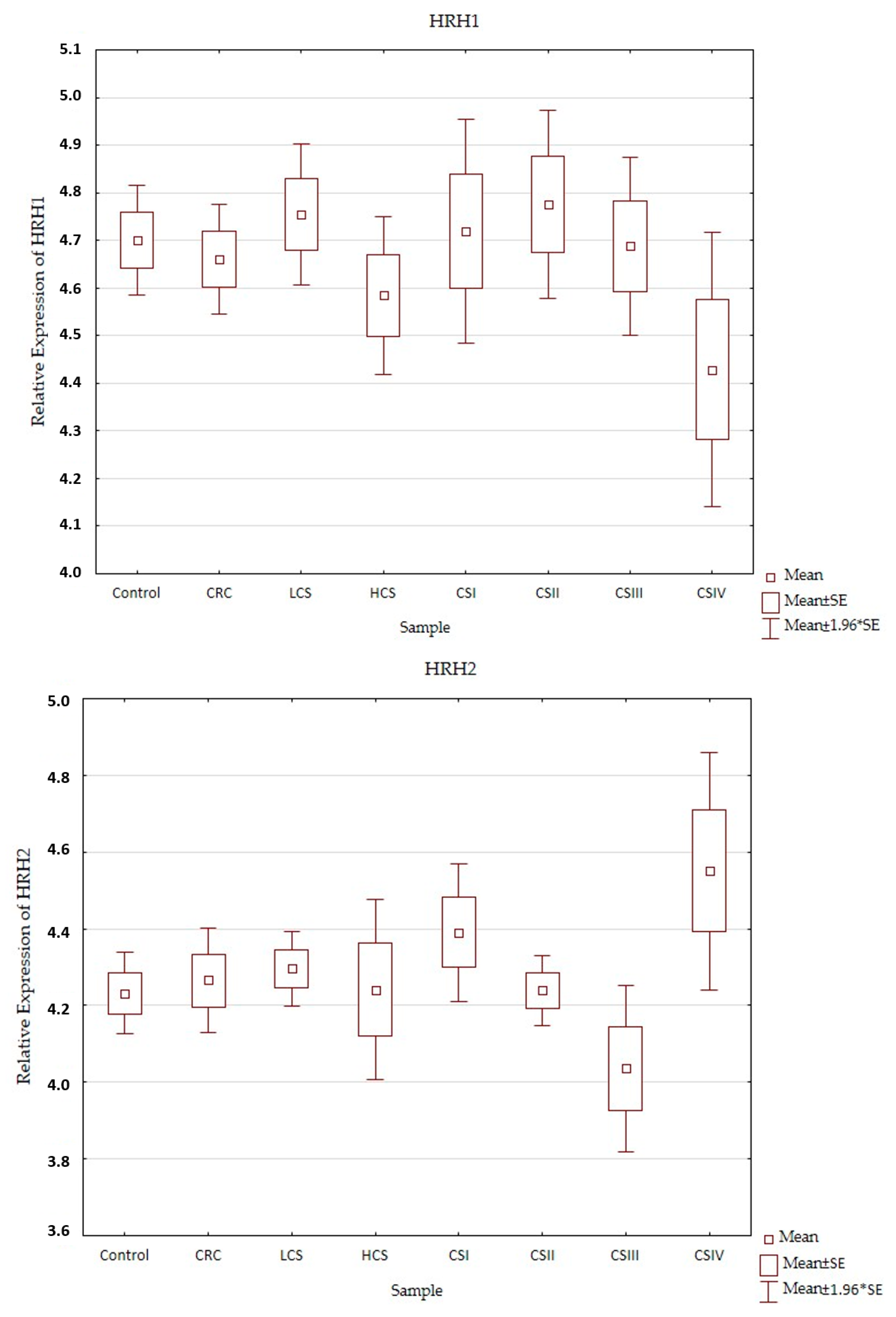
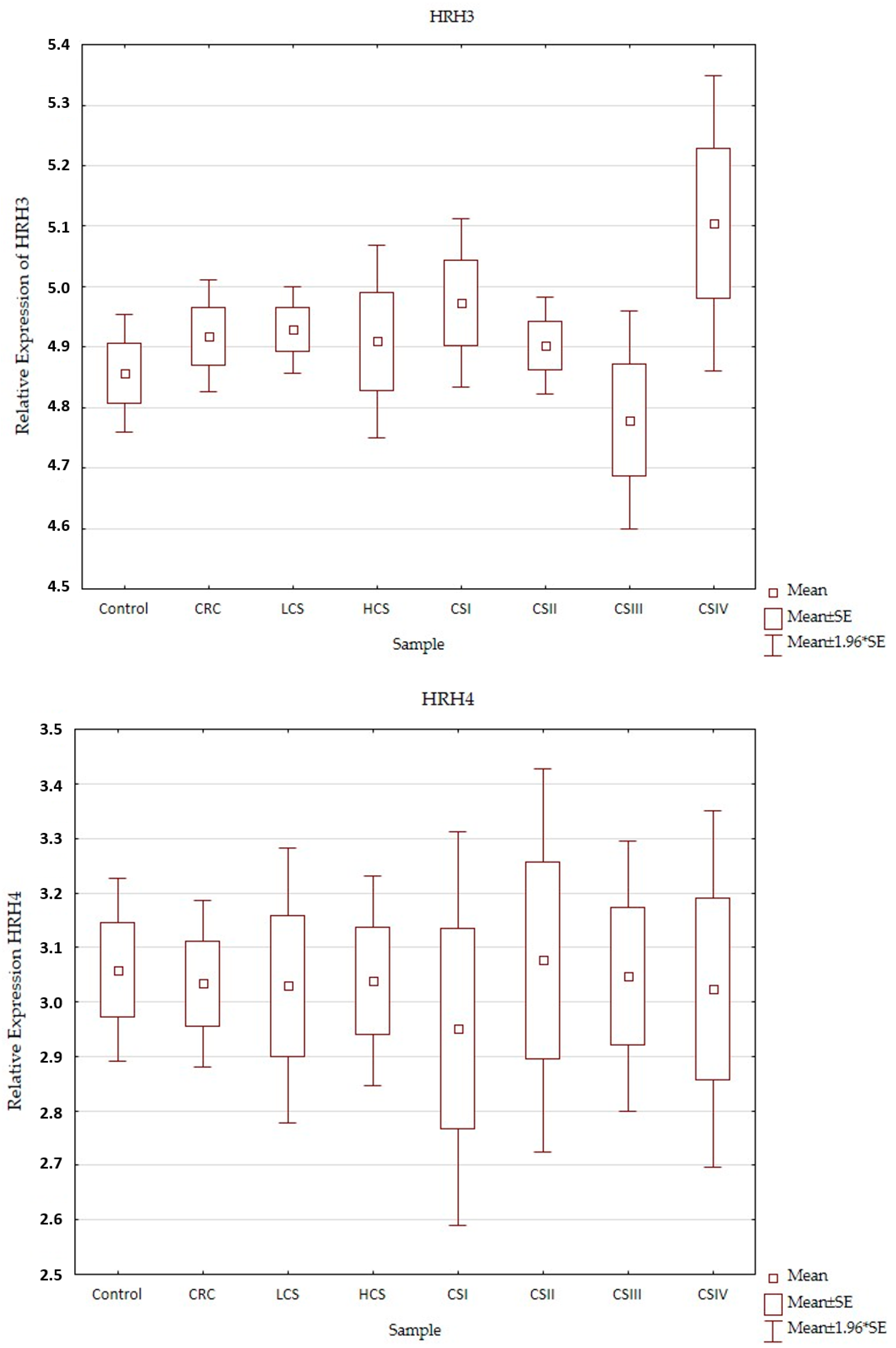
2.2. Expression Profile of Histaminergic Receptors
3. Discussion
4. Materials and Methods
4.1. Selection of Material for Analysis
4.2. Molecular Analysis of Samples
4.2.1. RNA Extraction and Purification
4.2.2. Microarray Hybridization
4.2.3. Relative Expression of Selected Transcripts
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gunter, M.J.; Alhomoud, S.; Arnold, M.; Brenner, H.; Burn, J.; Casey, G.; Chan, A.T.; Cross, A.J.; Giovannucci, E.; Hoover, R.; et al. Meeting report from the joint IARC-NCI international cancer seminar series: A focus on colorectal cancer. Ann. Oncol. 2019, 30, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Thanikachalam, K.; Khan, G. Colorectal cancer and nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Meyerhardt, J.A. Impact of diet and exercise on colorectal cancer. Hematol. Oncol. Clin. N. Am. 2022, 36, 471–489. [Google Scholar] [CrossRef]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Salas, N.; Dominguez, G.; Barderas, R.; Mendiola, M.; García-Albéniz, X.; Maurel, J.; Batlle, J.F. Clinical relevance of colorectal cancer molecular subtypes. Crit. Rev. Oncol. Hematol. 2017, 109, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Carchman, E. Crohn’s disease and the risk of cancer. Clin. Colon Rectal Surg. 2019, 32, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Branco, A.C.C.C.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of histamine in modulating the immune response and inflammation. Mediat. Inflamm. 2018, 2018, 9524075. [Google Scholar] [CrossRef] [Green Version]
- Losurdo, G.; Principi, M.; Girardi, B.; Pricci, M.; Barone, M.; Ierardi, E.; Di Leo, A. Histamine and histaminergic receptors in colorectal cancer: From basic science to evidence-based medicine. Anticancer. Agents Med. Chem. 2018, 18, 15–20. [Google Scholar] [CrossRef]
- Shi, Z.; Fultz, R.S.; Engevik, M.A.; Gao, C.; Hall, A.; Major, A.; Mori-Akiyama, Y.; Versalovic, J. Distinct roles of histamine H1- and H2-receptor signaling pathways in inflammation-associated colonic tumorigenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G205–G216. [Google Scholar] [CrossRef]
- Schirmer, B.; Neumann, D. The function of the histamine H4 receptor in inflammatory and inflammation-associated diseases of the Gut. Int. J. Mol. Sci. 2021, 22, 6116. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.L.; Akhter, J.; Adams, W.J.; Morris, D.L. Histamine content in colorectal cancer. Are there sufficient levels of histamine to affect lymphocyte function? Eur. J. Surg. Oncol. 1997, 23, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Cricco, G.P.; Mohamad, N.A.; Sambuco, L.A.; Genre, F.; Croci, M.; Gutiérrez, A.S.; Medina, V.; Bergoc, R.; Rivera, E.; Martin, G. Histamine regulates pancreatic carcinoma cell growth through H3 and H4 receptors. Inflamm. Res. 2008, 57, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Tiligada, E.; Ennis, M. Histamine pharmacology: From Sir Henry Dale to the 21st century. Br. J. Pharmacol. 2020, 177, 469–489. [Google Scholar] [CrossRef] [Green Version]
- Esbenshade, T.A.; Kang, C.H.; Krueger, K.M.; Miller, T.R.; Witte, D.G.; Roch, J.M.; Masters, J.N.; Hancock, A.A. Differential activation of dual signaling responses by human H1 and H2 histamine receptors. J. Recept. Signal Transduct. Res. 2003, 23, 17–31. [Google Scholar] [CrossRef]
- Seifert, R.; Strasser, A.; Schneider, E.H.; Neumann, D.; Dove, S.; Buschauer, A. Molecular and cellular analysis of human histamine receptor subtypes. Trends Pharmacol. Sci. 2013, 34, 33–58. [Google Scholar] [CrossRef] [Green Version]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [Green Version]
- Massari, N.A.; Nicoud, M.B.; Medina, V.A. Histamine receptors and cancer pharmacology: An update. Br. J. Pharmacol. 2020, 177, 516–538. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.L.; Cho, J. Pathophysiological roles of histamine receptors in cancer progression: Implications and perspectives as potential molecular targets. Biomolecules 2021, 11, 1232. [Google Scholar] [CrossRef] [PubMed]
- Davio, C.; Madlovan, A.; Shayo, C.; Lemos, B.; Baldi, A.; Rivera, E. Histamine receptors in neo-plastic transformation: Studies in human cell lines. Inflamm. Res. 1996, 45, 62–63. [Google Scholar] [CrossRef] [PubMed]
- Boer, K.; Helinger, E.; Helinger, A.; Pocza, P.; Pos, Z.; Demeter, P.; Baranyai, Z.; Dede, K.; Darvas, Z.; Falus, A. Decreased expression of histamine H1 and H4 receptors suggests disturbance of local regulation in human colorectal tumours by histamine. Eur. J. Cell Biol. 2008, 87, 227–236. [Google Scholar] [CrossRef]
- Medina, V.; Croci, M.; Crescenti, E.; Mohamad, N.; Sanchez-Jiménez, F.; Massari, N.; Nuñez, M.; Cricco, G.; Martin, G.; Bergoc, R.; et al. The role of histamine in human mammary carcinogenesis: H3 and H4 receptors as potential therapeutic targets for breast cancer treatment. Cancer Biol. Ther. 2008, 7, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Saxena, S.; Singh, A.; Singh, P. Tumor associated mast cells: Biological roles and therapeutic applications. Anat. Cell Biol. 2020, 53, 245–251. [Google Scholar] [CrossRef]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pączek, S.; Łukaszewicz-Zając, M.; Gryko, M.; Mroczko, P.; Kulczyńska-Przybik, A.; Mroczko, B. CXCL-8 in Preoperative colorectal cancer patients: Significance for diagnosis and cancer progression. Int. J. Mol. Sci. 2020, 21, 2040. [Google Scholar] [CrossRef] [Green Version]
- Bule, P.; Aguiar, S.I.; Aires-Da-Silva, F.; Dias, J.N.R. Chemokine-directed tumor microenvironment modulation in cancer immunotherapy. Int. J. Mol. Sci. 2021, 22, 9804. [Google Scholar] [CrossRef]
- Łukaszewicz-Zając, M.; Mroczko, B. Circulating biomarkers of colorectal cancer (CRC)-their utility in diagnosis and prognosis. J. Clin. Med. 2021, 10, 2391. [Google Scholar] [CrossRef]
- Medina, V.A.; Brenzoni, P.G.; Lamas, D.J.; Massari, N.; Mondillo, C.; Nunez, M.A.; Pignataro, O.; Rivera, E.S. Role of histamine H4 receptor in breast cancer cell proliferation. Front. Biosci. 2011, 3, 1042–1060. [Google Scholar] [CrossRef] [PubMed]
- Zanini, S.; Giovinazzo, F.; Alaimo, D.; Lawrence, B.; Pfragner, R.; Bassi, C.; Modlin, I.; Kidd, M. GNA15 expression in small intestinal neuroendocrine neoplasia: Functional and signalling pathway analyses. Cell. Signal. 2015, 27, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Feng, Q.; Liu, H.; Zhou, Q.; Hu, A.; Yamaguchi, T.; Xia, S.; Kobayashi, H. Bioinformatic evidences and analysis of putative biomarkers in pancreatic ductal adenocarcinoma. Heliyon 2019, 5, e02378. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Lu, J.; Liao, B.; Li, X.; Qian, X.; Li, K. Driver pattern identification over the gene co-expression of drug response in ovarian cancer by integrating high throughput genomics data. Sci. Rep. 2017, 7, 16188. [Google Scholar] [CrossRef] [Green Version]
- Massari, N.A.; Medina, V.A.; Martinel Lamas, D.J.; Cricco, G.P.; Croci, M.; Sambuco, L.; Bergoc, R.M.; Rivera, E.S. Role of H4 receptor in histamine-mediated responses in human melanoma. Melanoma Res. 2011, 21, 395–404. [Google Scholar] [CrossRef]
- Faustino-Rocha, A.I.; Ferreira, R.; Gama, A.; Oliveira, P.A.; Ginja, M. Antihistamines as promising drugs in cancer therapy. Life Sci. 2017, 172, 27–41. [Google Scholar] [CrossRef]
- Schirmer, B.; Rother, T.; Bruesch, I.; Bleich, A.; Werlein, C.; Jonigk, D.; Seifert, R.; Neumann, D. Genetic deficiency of the histamine H4-receptor reduces experimental colorectal carcinogenesis in mice. Cancers 2020, 12, 912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Innamorati, G.; Wilkie, T.M.; Malpeli, G.; Paiella, S.; Grasso, S.; Rusev, B.; Leone, B.E.; Valenti, M.T.; Carbonare, L.D.; Cheri, S.; et al. Gα15 in early onset of pancreatic ductal adenocarcinoma. Sci. Rep. 2021, 11, 14922. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, Q.; Chen, X.; Chen, S.; Yang, J.; Liu, Q.; Cheng, Y. Screening of tumor grade-related mRNAs and lncRNAs for esophagus squamous cell carcinoma. J. Clin. Lab. Anal. 2021, 35, e23797. [Google Scholar] [CrossRef]
- Tan, F.; He, D.; Hu, K.; Wang, D.; Zhang, S.; Li, J.; Wang, Z.; Tao, Y. WAVE2 enhanced hepatic stellate cells activity in colorectal liver metastases. Cancer Manag. Res. 2020, 12, 7671–7680. [Google Scholar] [CrossRef]
- Sun, L.N.; Xing, C.; Zhi, Z.; Liu, Y.; Chen, L.Y.; Shen, T.; Zhou, Q.; Liu, Y.H.; Gan, W.J.; Wang, J.R.; et al. Dicer suppresses cytoskeleton remodeling and tumorigenesis of colorectal epithelium by miR-324–5p mediated suppression of HMGXB3 and WASF-2. Oncotarget 2017, 8, 55776–55789. [Google Scholar] [CrossRef] [PubMed]
- Iwaya, K.; Oikawa, K.; Semba, S.; Tsuchiya, B.; Mukai, Y.; Otsubo, T.; Nagao, T.; Izumi, M.; Kuroda, M.; Domoto, H.; et al. Correlation between liver metastasis of the colocalization of actin-related protein 2 and 3 complex and WAVE2 in colorectal carcinoma. Cancer Sci. 2007, 98, 992–999. [Google Scholar] [CrossRef]
- Semba, S.; Iwaya, K.; Matsubayashi, J.; Serizawa, H.; Kataba, H.; Hirano, T.; Kato, H.; Matsuoka, T.; Mukai, K. Coexpression of actin-related protein 2 and Wiskott-Aldrich syndrome family verproline-homologous protein 2 in adenocarcinoma of the lung. Clin. Cancer Res. 2006, 12, 2449–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaya, K.; Norio, K.; Mukai, K. Coexpression of Arp2 and WAVE2 predicts poor outcome in invasive breast carcinoma. Mod. Pathol. 2007, 20, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Taniuchi, K.; Furihata, M.; Naganuma, S.; Saibara, T. WAVE2 is associated with poor prognosis in pancreatic cancers and promotes cell motility and invasiveness via binding to ACTN4. Cancer Med. 2018, 7, 5733–5751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.Y.; Tao, Y.M.; Ou, D.P.; Wang, W.; Chang, Z.G.; Wu, F. Increased expression of Wiskott-Aldrich syndrome protein family verprolin-homologous protein 2 correlated with poor prognosis of hepatocellular carcinoma. Clin. Cancer Res. 2006, 12, 5673–5679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Ding, Y.; Sun, L.; Shi, M.; Zhang, P.; He, A.; Zhang, X.; Huang, Z.; Li, R. WASF2 serves as a potential biomarker and therapeutic target in ovarian cancer: A Pan-Cancer analysis. Front. Oncol. 2022, 12, 840038. [Google Scholar] [CrossRef]
- Pang, Y.Y.; Li, J.D.; Gao, L.; Yang, X.; Dang, Y.W.; Lai, Z.F.; Liu, L.M.; Yang, J.; Wu, H.Y.; He, R.Q.; et al. The clinical value and potential molecular mechanism of the downregulation of MAOA in hepatocellular carcinoma tissues. Cancer Med. 2020, 9, 8004–8019. [Google Scholar] [CrossRef]
- Flamand, V.; Zhao, H.; Peehl, D.M. Targeting monoamine oxidase A in advanced prostate cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 1761–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikula, M.; Rubel, T.; Karczmarski, J.; Goryca, K.; Dadlez, M.; Ostrowski, J. Integrating proteomic and transcriptomic high-throughput surveys for search of new biomarkers of colon tumors. Funct. Integr. Genom. 2011, 11, 215–224. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Massri, M.; Ro, H.S. AEBP1 is a novel oncogene: Mechanisms of action and signaling pathways. J. Oncol. 2020, 2020, 8097872. [Google Scholar] [CrossRef]
- Yorozu, A.; Yamamoto, E.; Niinuma, T.; Tsuyada, A.; Maruyama, R.; Kitajima, H.; Numata, Y.; Kai, M.; Sudo, G.; Kubo, T.; et al. Upregulation of adipocyte enhancer-binding protein 1 in endothelial cells promotes tumor angiogenesis in colorectal cancer. Cancer Sci. 2020, 111, 1631–1644. [Google Scholar] [CrossRef]
- Shen, Z.; Feng, X.; Fang, Y.; Li, Y.; Li, Z.; Zhan, Y.; Lin, M.; Li, G.; Ding, Y.; Deng, H. POTEE drives colorectal cancer development via regulating SPHK1/p65 signaling. Cell Death Dis. 2019, 10, 863. [Google Scholar] [CrossRef] [Green Version]
- Long, J.; Xie, Y.; Yin, J.; Lu, W.; Fang, S. SphK1 promotes tumor cell migration and invasion in colorectal cancer. Tumour. Biol. 2016, 37, 6831–6836. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, L.; Chen, X.; Chen, X.; Diao, L.; Zeng, Y.; Xu, J. TNFAIP6 defines the MSC subpopulation with enhanced immune suppression activities. Stem Cell Res. Ther. 2022, 13, 479. [Google Scholar] [CrossRef] [PubMed]
- Yip, K.T.; Das, P.K.; Suria, D.; Lim, C.R.; Ng, G.H.; Liew, C.C. A case-controlled validation study of a blood-based seven-gene biomarker panel for colorectal cancer in Malaysia. J. Exp. Clin. Cancer Res. 2010, 29, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offenberg, H.; Brünner, N.; Mansilla, F.; Orntoft Torben, F.; Birkenkamp-Demtroder, K. TIMP-1 expression in human colorectal cancer is associated with TGF-B1, LOXL2, INHBA1, TNF-AIP6 and TIMP-2 transcript profiles. Mol. Oncol. 2008, 2, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Yao, W.; Xiong, Y.; Li, J.; Liu, L.; Shi, L.; Zhang, W.; Zhang, C.; Nie, L.; Wan, J. Attenuated expression of HRH4 in colorectal carcinomas: A potential influence on tumor growth and progression. BMC Cancer 2011, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Cianchi, F.; Cortesini, C.; Schiavone, N.; Perna, F.; Magnelli, L.; Fanti, E.; Bani, D.; Messerini, L.; Fabbroni, V.; Perigli, G.; et al. The role of cyclooxygenase-2 in mediating the effects of histamine on cell proliferation and vascular endothelial growth factor production in colorectal cancer. Clin. Cancer Res. 2005, 11, 6807–6815. [Google Scholar] [CrossRef] [Green Version]
- Cricco, G.; Martín, G.; Medina, V.; Núñez, M.; Gutiérrez, A.; Cocca, C.; Bergoc, R.; Rivera, E. Histamine regulates the MAPK pathway via the H(2) receptor in PANC-1 human cells. Inflamm. Res. 2004, 53, S65–S66. [Google Scholar] [CrossRef]
- Edge, S.B.; Byrd, S.R.; Compton, C.C.; Fritz, A.G.; Greene, F.L.; Trotti, A. AJCC Cancer Staging Manual, 7th ed.; Springer: New York, NY, USA, 2010; pp. 143–164. [Google Scholar]
- Amin, M.B.; Edge, S.B.; Greene, F.L.; Byrd, D.R.; Brookland, R.K.; Washington, M.K.; Gershenwald, J.E.; Compton, C.C.; Hess, K.R.; Sullivan, D.C.; et al. AJCC Cancer Staging Manual, 8th ed.; Springer: New York, NY, USA, 2017. [Google Scholar]
- Reiner, A.; Yekutieli, D.; Benjamini, Y. Identifying differentially expressed genes using false discovery rate controlling procedures. Bioinformatics 2003, 19, 368–375. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probes | Design | p-Value | p < 0.05 | p < 0.02 | p < 0.01 | p < 0.005 |
|---|---|---|---|---|---|---|
| Histaminergic system | CRC | uncorr | 33 | 25 | 18 | 13 |
| BH | 10 | 4 | 4 | 4 | ||
| LCS–HCS | uncorr | 31 | 20 | 14 | 9 | |
| BH | 7 | 6 | 6 | 6 | ||
| CSI–CSIV | uncorr | 24 | 18 | 13 | 10 | |
| BH | 10 | 9 | 7 | 5 | ||
| Inflammation | CRC | uncorr | 154 | 108 | 78 | 62 |
| BH | 62 | 34 | 24 | 18 | ||
| LCS–HCS | uncorr | 156 | 99 | 80 | 59 | |
| BH | 53 | 26 | 22 | 14 | ||
| CSI–CSIV | uncorr | 181 | 114 | 84 | 66 | |
| BH | 71 | 39 | 32 | 24 |
| Probes | Gene Symbol | Probe Set ID | Clinical Stage, Fold Change, Regulation | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FC CRC | p | BH | FC LCS | FC HCS | p | BH | FC CSI | FC CSII | FC CSIII | FC CSIV | p | BH | |||
| Histaminergic system | GNA15 | 205349_at | 1.4 ↑ | 1 × 10−3 | 3 × 10−2 | 1.3 ↑ | 1.6 ↑ | 2 × 10−2 | 3 × 10−2 | 1.2 ↑ | 1.3 ↑ | 1.4 ↑ | 2.0 ↑ | 1 × 10−3 | 2 × 10−2 |
| HRH2 | 220805_at | 1.1 ↑ | 1.0 ↑ | −1.1 ↓ | 1.2 ↑ | 3 × 10−2 | |||||||||
| HRH3 | 220447_at | 1.1 ↑ | 1.1 ↑ | −1.0 ↓ | 1.4 ↑ | 3 × 10−2 | |||||||||
| MAOA | 204388_s_at | −2.8 ↓ | 8 × 10−9 | 6 × 10−7 | −3.2 ↓ | −2.4 ↓ | 3 × 10−8 | 4 × 10−6 | −1.6 ↓ | −3.1 ↓ | −2.7 ↓ | −4.1 ↓ | 5 × 10−9 | 3 × 10−7 | |
| MAOA | 204389_at | −2.8 ↓ | 3 × 10−7 | 1 × 10−5 | −3.3 ↓ | −2.8 ↓ | 6 × 10−8 | 4 × 10−6 | −1.6 ↓ | −3.9 ↓ | −2.5 ↓ | −4.8 ↓ | 8 × 10−10 | 1 × 10−8 | |
| MAOA | 212741_at | −3.0 ↓ | 9 × 10−9 | 6 × 10−7 | −3.2 ↓ | −2.5 ↓ | 1 × 10−6 | 5 × 10−5 | −1.6 ↓ | −3.4 ↓ | −2.5 ↓ | −4.6 ↓ | 1 × 10−7 | 6 × 10−5 | |
| WASF2 | 221725_at | −1.5 ↓ | 5 × 10−3 | −1.4 ↓ | −1.6 ↓ | 2 × 10−2 | −1.7 ↓ | −4.2 ↓ | −1.2 ↓ | −2.4 ↓ | 5 × 10−3 | 9 × 10−3 | |||
| Inflammation | AEBP1 | 201792_at | 2.4 ↑ | 9 × 10−4 | 2 × 10−2 | 1.5 ↑ | 3.4 ↑ | 3 × 10−4 | 9 × 10−3 | −1.1 ↓ | 2.1 ↑ | 3.5 ↑ | 3.2 ↑ | 8 × 10−5 | 1 × 10−2 |
| CXCL1 | 204470_at | 3.5 ↑ | 8 × 10−6 | 6 × 10−4 | 4.8 ↑ | 2.8 ↑ | 1 × 10−5 | 1 × 10−3 | 6.7 ↑ | 3.9 ↑ | 2.6 ↑ | 3.1 ↑ | 2 × 10−5 | 1 × 10−3 | |
| CXCL2 | 209774_x_at | 4.2 ↑ | 1 × 10−5 | 8 × 10−4 | 5.2 ↑ | 3.5 ↑ | 4 × 10−5 | 2 × 10−3 | 9.2 ↑ | 3.7 ↑ | 4.4 ↑ | 2.4 ↑ | 3 × 10−5 | 1 × 10−3 | |
| CXCL3 | 207850_at | 3.5 ↑ | 4 × 10−6 | 6 × 10−4 | 5.0 ↑ | 2.7 ↑ | 4 × 10−6 | 6 × 10−4 | 8.7 ↑ | 3.6 ↑ | 2.6 ↑ | 2.8 ↑ | 2 × 10−6 | 2 × 10−4 | |
| CXCL8 | 202859_x_at | 4.4 ↑ | 5 × 10−5 | 3 × 10−3 | 4.2 ↑ | 4.6 ↑ | 2 × 10−4 | 8 × 10−3 | 3.9 ↑ | 4.3 ↑ | 3.3 ↑ | 7.3 ↑ | 4 × 10−4 | 7 × 10−3 | |
| SPHK1 | 219257_s_at | 1.7 ↑ | 5 × 10−6 | 6 × 10−4 | 1.3 ↑ | 2.1 ↑ | 4 × 10−8 | 2 × 10−5 | 1.2 ↑ | 1.3 ↑ | 1.8 ↑ | 2.6 ↑ | 6 × 10−9 | 4 × 10−6 | |
| TNFAIP6 | 206025_s_at | 2.3 ↑ | 4 × 10−4 | 1 × 10−2 | 1.5 ↑ | 3.3 ↑ | 8 × 10−5 | 4 × 10−3 | 1.4 ↑ | 1.6 ↑ | 2.8 ↑ | 4.2 ↑ | 3 × 10−6 | 5 × 10−3 | |
| TNFAIP6 | 206026_s_at | 1.8 ↑ | 3 × 10−4 | 8 × 10−3 | 1.3 ↑ | 2.3 ↑ | 3 × 10−5 | 2 × 10−3 | 1.1 ↑ | 1.4 ↑ | 2.0 ↑ | 2.8 ↑ | 5 × 10−5 | 2 × 10−3 | |
| A | B | C | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control vs. Control | Control vs. CRC | CRC vs. CRC | |||||||||||||||
| Gene | vs. | Gene | r | p | Gene | vs. | Gene | r | p | Gene | vs. | Gene | r | p | |||
| GNA15 | ⇄ | HRH3 | + | 0.63 | 0.00 | HRH2 | ⇄ | GNA15 | + | 0.50 | 0.03 | GNA15 | ⇄ | HRH2 | + | 0.53 | 0.02 |
| HRH3 | ⇄ | CXCL3 | + | 0.48 | 0.04 | HRH4 | ⇄ | GNA15 | − | −0.58 | 0.01 | GNA15 | ⇄ | MAOA | − | −0.48 | 0.04 |
| HRH4 | ⇄ | MAOA | + | 0.56 | 0.02 | HRH4 | ⇄ | SPHK1 | − | −0.48 | 0.05 | GNA15 | ⇄ | CXCL8 | + | 0.68 | 0.00 |
| HRH4 | ⇄ | WASF2 | − | −0.48 | 0.04 | HRH4 | ⇄ | TNFAIP6 | − | −0.67 | 0.00 | GNA15 | ⇄ | SPHK1 | + | 0.68 | 0.00 |
| HRH4 | ⇄ | AEBP1 | − | −0.63 | 0.01 | MAOA | ⇄ | TNFAIP6 | − | −0.58 | 0.01 | GNA15 | ⇄ | TNFAIP6 | + | 0.57 | 0.01 |
| HRH4 | ⇄ | SPHK1 | − | −0.58 | 0.04 | WASF2 | ⇄ | GNA15 | + | 0.47 | 0.05 | HRH2 | ⇄ | WASF2 | − | −0.55 | 0.02 |
| MAOA | ⇄ | WASF2 | − | −0.60 | 0.01 | WASF2 | ⇄ | AEBP1 | + | 0.56 | 0.02 | HRH4 | ⇄ | SPHK1 | − | −0.48 | 0.05 |
| MAOA | ⇄ | AEBP1 | − | −0.71 | 0.00 | WASF2 | ⇄ | SPHK1 | + | 0.54 | 0.02 | MAOA | ⇄ | WASF2 | + | 0.54 | 0.02 |
| MAOA | ⇄ | SPHK1 | − | −0.57 | 0.01 | WASF2 | ⇄ | TNFAIP6 | + | 0.60 | 0.01 | MAOA | ⇄ | SPHK1 | − | −0.61 | 0.01 |
| MAOA | ⇄ | TNFAIP6 | − | −0.51 | 0.03 | AEBP1 | ⇄ | TNFAIP6 | + | 0.55 | 0.02 | MAOA | ⇄ | TNFAIP6 | − | −0.48 | 0.04 |
| WASF2 | ⇄ | AEBP1 | + | 0.74 | 0.00 | CXCL1 | ⇄ | CXCL2 | + | 0.48 | 0.04 | WASF2 | ⇄ | SPHK1 | + | 0.54 | 0.02 |
| WASF2 | ⇄ | SPHK1 | + | 0.60 | 0.00 | CXCL1 | ⇄ | CXCL3 | + | 0.54 | 0.02 | AEBP1 | ⇄ | SPHK1 | + | 0.69 | 0.00 |
| AEBP1 | ⇄ | SPHK1 | + | 0.69 | 0.00 | CXCL3 | ⇄ | CXCL2 | + | 0.52 | 0.03 | AEBP1 | ⇄ | TNFAIP6 | + | 0.50 | 0.04 |
| AEBP1 | ⇄ | TNFAIP6 | + | 0.47 | 0.05 | CXCL3 | ⇄ | CXCL3 | + | 0.54 | 0.02 | CXCL1 | ⇄ | CXCL2 | + | 0.79 | 0.00 |
| CXCL1 | ⇄ | CXCL2 | + | 0.63 | 0.00 | CXCL3 | ⇄ | AEBP1 | − | −0.51 | 0.03 | CXCL1 | ⇄ | CXCL3 | + | 0.85 | 0.00 |
| CXCL1 | ⇄ | CXCL3 | + | 0.79 | 0.00 | CXCL1 | ⇄ | CXCL8 | + | 0.62 | 0.00 | ||||||
| CXCL1 | ⇄ | CXCL8 | + | 0.53 | 0.02 | CXCL2 | ⇄ | CXLC3 | + | 0.88 | 0.00 | ||||||
| CXCL2 | ⇄ | CXCL3 | + | 0.73 | 0.00 | CXCL8 | ⇄ | SPHK1 | + | 0.65 | 0.00 | ||||||
| CXCL2 | ⇄ | CXCL8 | + | 0.82 | 0.00 | CXCL8 | ⇄ | TNFAIP6 | + | 0.58 | 0.01 | ||||||
| CXCL2 | ⇄ | TNFAIP6 | + | 0.61 | 0.01 | SPHK1 | ⇄ | TNFAIP6 | + | 0.72 | 0.00 | ||||||
| CXCL3 | ⇄ | CXCL8 | + | 0.50 | 0.03 | ||||||||||||
| CXCL8 | ⇄ | SPHK1 | + | 0.55 | 0.02 | ||||||||||||
| CXCL8 | ⇄ | TNFAIP6 | + | 0.78 | 0.00 | ||||||||||||
| SPHK1 | ⇄ | TNFAIP6 | + | 0.55 | 0.02 | ||||||||||||
| Parameters | CRC | Control |
|---|---|---|
| Age [years] | 51–79 | 52–76 |
| Male [%] | 50 | 56.6 (6) |
| Haemoglobin [g/dL] | 9.27–11.59 | 10.15–11.62 |
| WBC ×103/µL | 7–10.5 | 5–10 |
| Platelete [number/µL] | 150,000–350,000 | 150,000–400,000 |
| Glycemia [mg/dL] | 73–115 | 80–110 |
| Creatinine [mg/dL] | 0.86–1.10 | 0.95–1.15 |
| PT INR | 0.8–1.2 | 0.85–1.15 |
| APTT [s] | 26–36 | 27–38 |
| CRP [mg/dL] | 2–6 | 1–5 |
| Samples | Clinical Stage | Clinical features | Histopathological Grading (% of All) | |
|---|---|---|---|---|
| CRC | LCS | CSI | T1N0M0 | G2 (75), G1 (25) |
| CSII | T3N0M0 | G1 (60), G2 (40) | ||
| HCS | CSIII | T3N1M0, T3N2M0 | G1 (60), G3 (40) | |
| CSIV | Any T Any N M1a, Any T Any N M1b | G2 (75), G3 (25) | ||
| Control | Tissues without signs of neoplastic changes under the microscope, histopathologically normal large intestine | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janikowska, G.; Janikowski, T.; Plato, M.; Mazurek, U.; Orchel, J.; Opiłka, M.; Lorenc, Z. Histaminergic System and Inflammation-Related Genes in Normal Large Intestine and Adenocarcinoma Tissues: Transcriptional Profiles and Relations. Int. J. Mol. Sci. 2023, 24, 4913. https://doi.org/10.3390/ijms24054913
Janikowska G, Janikowski T, Plato M, Mazurek U, Orchel J, Opiłka M, Lorenc Z. Histaminergic System and Inflammation-Related Genes in Normal Large Intestine and Adenocarcinoma Tissues: Transcriptional Profiles and Relations. International Journal of Molecular Sciences. 2023; 24(5):4913. https://doi.org/10.3390/ijms24054913
Chicago/Turabian StyleJanikowska, Grażyna, Tomasz Janikowski, Marta Plato, Urszula Mazurek, Joanna Orchel, Mieszko Opiłka, and Zbigniew Lorenc. 2023. "Histaminergic System and Inflammation-Related Genes in Normal Large Intestine and Adenocarcinoma Tissues: Transcriptional Profiles and Relations" International Journal of Molecular Sciences 24, no. 5: 4913. https://doi.org/10.3390/ijms24054913
APA StyleJanikowska, G., Janikowski, T., Plato, M., Mazurek, U., Orchel, J., Opiłka, M., & Lorenc, Z. (2023). Histaminergic System and Inflammation-Related Genes in Normal Large Intestine and Adenocarcinoma Tissues: Transcriptional Profiles and Relations. International Journal of Molecular Sciences, 24(5), 4913. https://doi.org/10.3390/ijms24054913